The phagocytic recognition/clearance of senescent HL60-A1 transfectants is mediated by apoptosis-dependent and -independent mechanisms
Ling-Ling Chiang*, Chien-Ying Liu, Hao-Cheng Chen, Shu-Min Lin, Chien-Da Huang, Chun-Hua Wang, Horng-Chyuan Lin, Chih-Ten Yu and Han-Pin Kuo
*Department of Respiratory Therapy, Taipei Medical University, and Departments of Thoracic Medicine, Chang Gung Memorial Hospital, Taipei, Taiwan
Running title: An apoptosis-independent mechanism mediating phagocytic clearance
Address correspondence to Dr. Han-Pin Kuo
Professor and Head
Department of Thoracic Medicine
Chang-Gung Memorial Hospital and Chang-Gung University 199, Tun-Hwa N. Rd. Taipei 105, Taiwan
Tel. +886 3 328-1200, ext. 8468 Fax +886 3 327-2474
E-Mail [email protected]
Abstract
Objective: To investigate the phagocytic recognition/clearance of the senescent cells using promyelocytic cell line, HL60, overexpressing Bcl-2 homilogue, A1, as a model.
Materials and Methods: Apoptotic and senescent, nonapoptotic HL60-A1 transfectants were isolated using the MACS Apoptotic Cell Isolation Kit and labeled with a fluorescent marker.
Monocyte-derived macrophages (MDM) were cultured to perform the phagocytosis assay, and flow cytometry was used to determine the phagocytic index. Blocking agents, including anti- CD14, anti-CD36, and anti-αvβ3 antibodies, as well as the scavenger receptor inhibitor, Oxi- LDL were used to differentiate the mechanism of recognition between apoptotic and nonapoptotic cells.
Results: Under the serum-free condition, the phagocytic index of HL60-A1 transfectants was little different from that of the HL60-EGFP control, despite having a significantly lower degree of apoptosis. While the phagocytic index of HL60-EGFP control was significantly correlated with the degree of apoptosis, that index of HL60-A1 transfectants was less relevant to apoptosis. The two cell types did not differ in the phagocytic index of annexin V-positive cells.
However, the phagocytic index of annexin V-negative cells from HL60-A1 transfectants was significantly higher than that of the control cells and increased with the age in days.
Preincubation of MDM with scavenger receptor inhibitor, Oxi-LDL, and the inhibitory antibodies of αvβ3, CD14 and CD36 could attenuate the phagocytic recognition of annexin V- positive HL60 cells but not senescent, annexin V-negative A1 transfectants.
Conclusions: This study suggests that a different recognition mechanism exists, which mediates the phagocytic clearance of the senescent cells and may play a role in the resolution of inflammation.
Keywords
Lung inflammation Macrophage Phagocytosis Apoptosis Bcl-2 A1
Background
Recruitment of inflammatory cells, including neutrophils, lymphocytes and monocytes is important in activating and modulating tissue inflammation [1]. Although the factors regulating the fate of the transmigrated cells in tissue are poorly undersood, the phagocytic recognition and clearance of the recruited inflammatory cells has been shown to play a pivotal role in relieving tissue inflammation and maintaining tissue homeostasis [2, 3]. Among the inflammatory cells, neutrophils were thought to have an inherently limited life-span in tissue (namely constitutively programmed cell death), but recent evidence suggests that their survival in tissue is partially regulated by local factors, including adhesion, cytokines, and chemokines [3, 4]. Neutropil persistence in tissue may be a major determinant of acute organ injury since the possibility of organ injury caused by the release of proteases and reactive oxygen intermediates increases with the time that neutrophils are present in the tissue [5]. While the resolution of acute inflammation ultimately depends on neutrophil clearance, the clearance mechanism may also influence the duration and severity of tissue inflammation [6, 7]. Necrosis of neutrophils releases toxins extracellularly, thus perpetuating the inflammatory response and further damaging tissue [8, 9]. In contrast, apoptosis of neutrophils and their subsequent phagocytosis by resident macrophages or elicited monocyte-derived macrophages may terminate the inflammation [8, 9]. It has been widely accepted that factors promoting neutrophil apoptosis and engulfment by macrophages can accelerate the resolution of tissue inflammation, while those preventing apoptosis may prolong inflammatoion and increase the likelihood of acute organ injury [8-10]. However, there are studies reporting that apoptotic neurophils release cytotoxic mediators and cause tissue cell damage [6], and the progression of inflammation is prevented by the inhibition of apoptosis [11], suggesting the importance of apoptosis- independent mechanisms in the resolution of inflammation.
The apoptotic cells are destined to be cleared; however, the clearance of recruited inflammatory cells may operate through an apoptosis-independent mechanism. Lagasee et al.
found that over-expression of human Bcl-2 in mouse neutrophils prevented apoptosis but did not prevent phagocytosis by macrophages ex vivo or retard clearance in thioglycollate-induced peritonitis [12, 13]. Their study also revealed that Bcl-2 attenuated neutrophil apoptosis, while the homeostasis of the neutrophil population was essentially unaffected [13]. All these findings suggested that phagocytosis of the aging neutrophils exists and is independent of the process of apoptosis regulated by Bcl-2. Endogenous expression of Bcl-2 is restricted to early myeloid cells in bone marrow, and is not present in mature polymorphonuclear neutrophils of the blood [12, 14]. However, A1 is a constitutively expressed- and inducible-Bcl-2 homologue in neutrophils [5]. Further examination of the paradoxical phenomen found by Lagasse and Weissman using the endogenous Bcl-2 homologue in mature neutrophils, A1, is particularly important given its implications for understanding the relationship between neutrophil senescence and recognition/phagocytosis. In the present study, we use promyelocytic cell line, HL60, over-expressing A1 as a model to investigate the mechanism of the phagocytic recognition/clearance of the senescent cells. The stable expression of the mRNA and the fusion protein of the EGFP-A1 molecule in HL60-A1 transfectants, demonstrated by ribonyclease- protection assay and Western blotting respectively, had already shown in our previous publication [15]. Additionally, a simple labeling procedure combined with flow cytometric analysis to quantify the percentage of phagocytic cells [16] was used to facilitate the research.
Materials and Methods
Reagents and Cells
The solutions for cell culture, DMEM, EMEM, RPMI-1640, fetal calf serum (FCS, 10 x), non-essential amino acids (NEAA, 100 x), sodium pyruvate (100 x), penicillin-streptomycin (100 x), L-glutamine (100 x), and granulocyte monocyte-colony stimulating factor (GM-CSF) were purchased from Life Technologies (Grand Island, NY). FITC conjugated annexin-V, and propidium iodide were purchased from BD PharMingen (San Diegi, CA), and red fluorescent cell linker was obtained from DAKO (Carpinteria, CA). Meanwhile, blocking antibodies, anti- CD14-, anti-CD36-, anti-αvβ3-antibodies, scavenger receptor inhibitor, Oxi-LDL, were obtained from Calbiochem (La Jolla, CA), and the human promyelocytic HL60 cells were obtained from the American Tissue and Cell Collection (ATCC) and maintained in EMEM with the above additives. Finally, the monocytes were isolated from peripheral blood obtained from healthy volunteers.
Construct of Plasmids
The fragment containing the open reading frame of the cloned human A1 cDNA [17, 18]
was inserted in-fame into the EGFP-C1 plasmid (Clontech, Palo Alto, CA). The resulting pEGFP-A1 plasmid was then sequenced to confirm the in-frame fusion of EGFP cDNA5’ to the A1 open reading frame.
Transfection and selection of stable clones
HL-60 cells were transfected according to the optimized protocol described previously [19, 20], with minor modifications. Briefly, growth phase HL-60 cells were resuspended at 5 x 107 cells/ml in DMEM with 1.25% DMSO. Meanwhile, 250 microliters of the cells were mixed
with 20 μg of previously linearized p-EGFP-A1 plasmid DNA in 50 μl of the same medium and placed in a pre-chilled 4 mm gap electroporation cuvette (BTX, San Diego, CA).
Electroporation was conducted at 290V, R1 (minimal resistance), and 960 μF on a BTX 600 instrument, and the electroporated cells were rapidly transferred to a pre-warmed 25 cm2 flask containing growth medium but no selection antibiotic. G418 (Life Technologies, Grand Island, NY), at a final concentration of 1 mg/ml, was added 36 hours after transfection, and the selection medium was replaced as needed for the next month. Using mock-transfected (no plasmid DNA) HL-60 cells cultured in parallel as a guide, pEGFP-A1 transfected cells were removed from the selection after approximately one month to allow out-growth of the surviving cells. These cells were sorted into polyclonal pools and individual clones (referred to as the ‘A1 transfectants’ elsewhere in this study based on green fluorescence. Similarly, HL-60 cells with stable EGFP expression and no A1 fusion were generated using the pEGFP-C1 plasmid, sorted into a polyclonal pool (‘EGFP Control’). All the sorted HL-60 transfectants were subsequently cultured in supplemented EMEM medium.
Confocal microscopy
Fluorescent images of the HL60-A1 transfectants were obtained using a confocal laser scanning microscope (MRC 1000; Bio-Rad, Hemel Hempstead,UK) with COMOS software (version 7.0; Bio-Rad). The excitationwavelengths of the krypton/argon laser were 488 nm for EGFP. The live cells were opticallysectioned by scanning at increasing focus depths (typically, in steps of 0.5 µm), and the maximum intensity of thecorresponding pixels in an optical section displaying maximal cell size was selected adjusted to generate a two-dimentional projected image.
Preparation of monocyte-derived macrophages
Human monocytes were isolated as described previously [21]. Briefly, 50 ml of peripheral venous blood was drawn from a single volunteer, with citrate (0.31% w/v) being used as an anticoagulant. The blood was diluted 2-fold with RPMI-1640 (pH 7.4) before centrifugation at 300 x g at 20°C for 10 min. The cell pellet was layered on a Percoll gradient (p=1.077 g/ml), and the interphase containing the peripheral blood mononuclear cells (PBMC) was obtained following 20 min of centrifugation at 450 x g and 20°C. The mononuclear cells were then resuspended (106 cells/ml) in RPMI-1640 (GIBCO) medium containing 5% fetal calf serum (FCS), 100 U/ml penicillin, and 100 μg/ml streptomycin. After resuspension, the cells were seeded onto the plastic surface of a 75 cm2 culture dish for 2 h at 37 °C to let the monocytes adhere. Following the incubation, non-adherent cells were removed with three rinses of 5 ml RPMI-1640 containing 5% FCS, and the adherent cells were then detached by scrapping with rubber policemen. Isolated monocytes were constantly >98% pure on Luko-stat stain (Fisher Scientific, Pittsburgh, PA) and >99% viable as determined by trypan blue exclusion. The monocytes were resuspended in RPMI-1640 containing GM-CSF (1 ng/ml) and then plated at an initial density of 2 x 105 cells/well on 48-well culture dishes. Medium was refreshed 5 days after plating and before the experiments. Human monocyte-derived macrophages (HMDM) were used for experiments between days seven and ten after plating.
Isolation of Annexin V-positive and negative HL60 cells
Apoptotic HL60 cells were isolated using the MACS Apoptotic Cell Isolation Kit (Miltenyi Biotech, Auburn, CA, USA) following the manufacturer’s instructions. Briefly, HL60 cells in serum-free medium were centrifuged for 10 min at 300 x g. After the supernatant had been completely removed, the cells were resuspended in 80 μl of binding buffer and 20 μl of annexin V-conjugated paramagnetic microbeads/107 cells, and then incubated for 15 min at
6°C. After washing once in 5 ml of binding buffer, the cells were loaded onto a Midi-MACS column, washed with binding buffer, and both annexin V-selected (by a magnet) and flow- through (AV-negative) fractions were collected. To ensure quality, the purity of each fraction was measured by double-labeling with annexin V-FITC (PharMingen, San Diego, CA, USA) and propidium iodide (PI) (Molecular Probes, Oregon, USA). Briefly, fractions of HL60 cells were washed once with phosphate-buffered saline, then incubated with annexin V- FITC and PI in binding buffer containing 10 mM HEPES, 1.5 mM MgCl2, and 2.5 mM CaCl2 in normal saline for 15 min at room temperature. Flow cytometry revealed that the purity of the annexin V-positive and negative fractions constantly exceeded 95%.
Cell survival and apoptosis assay
HL-60 cells were resuspended at 1x106 cells/ml in EMEM, in the presence or absence of serum, and cultured for various periods in 24-well plates. Cell death was assessed using various complementary methods. After culturing, cells were counted using a hematocytometer and viability determined by exclusion of trypan blue. Meanwhile, DNA fragmentation was quantitated by flow cytometric analysis of propidium iodide stained DNA in ethanol-fixed cells. Phosphoserine exteriorization of the cell surface was quantitated using flow cytometric analysis of annexin-V-PE (Pharmingen) staining, and all procedures were performed according to the manufactuer’s protocols.
Cell cycle analysis
Cell cycle progression was quantitated using flow cytometric analysis of propidium iodide stained DNA in ethanol-fixed cells. Meanwhile, the G0, G1, synthetic (S), G2, mitotic (M) phases of the cell cycle were automatically calculated using the CellQuest Software. The proliferation index was defined as (S/M+S2)/(G0+G1).
Phagocytosis assay
At the end of various treatments, HL60-cells were suspended in buffer A (108 cells/ml) and labeled with 5 μM final concentration of red fluorescent cell linker for 15 min at 37 °C.
The cells were then washed twice with 15 ml of ice-cold buffer A and resuspended in RPMI- 1640 containing 10% FBS. To determine the phagocytic clearance of cells by MDM, 1x106 of HL-60 cells in 200 μl of RPMI (10% FBS) were incubated with MDM for 1 hr at 37 , and ℃ phagocytosis was proceeded for 1 h at 37°C. Non-phagocytosed HL-60 cells were removed by five extensive washings in PBS. The MDM were then resuspended by exposure to trypsin (0.005% w/v) for 5 min at 37°C.
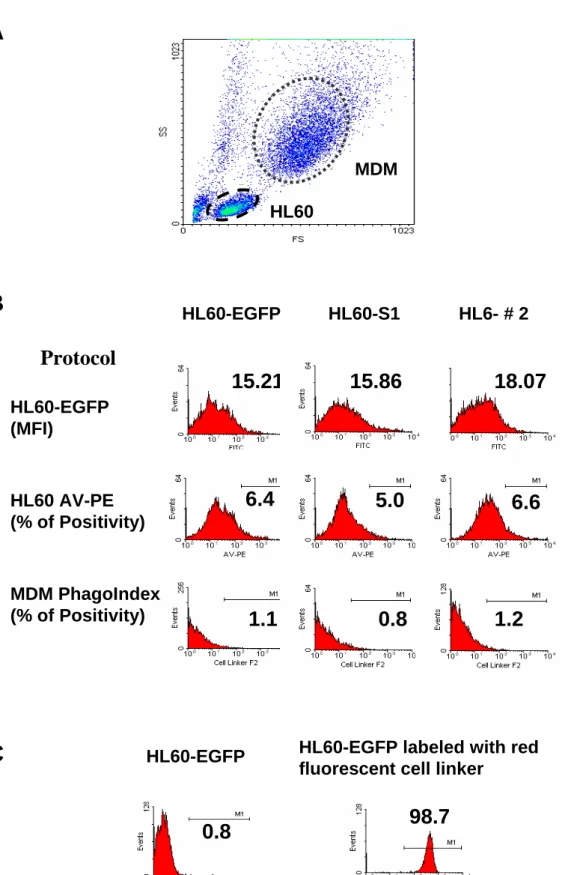
Flow cytometry was used to perform the phagocytosis assay [16]. Flow cytometric analysis utilizing density plots of forward scatter (FSC) versus side scatter (SSC) clearly distinguished the two populations of cells in co-cultures containing HL60 cells and MDM, (Fig. 1A). The green fluorescence profiles for the HL60-A1 transfectants and the control EGFP-transfectants were determined before examining the cell-co-cultures and analysing the phagocytosis assay. The fluorescence intensity of EGFP at 530 nm (FL1) did not differ significantly between cells, and no significant fluorescent interference of EGFP occurred at 585 nm (FL2) when using protocols to analyze the apoptosis and the phagocytic index of cells (Fig.
1B). Following labelling with the red fluorescent cell linker, the fluorescent intensity of HL60 cells detected at FL2 was two to three logs higher than that of the unlabeled cells. The cut-off value for defining red fluorescent positivity was set arbitrarily to include over 98 % of the labelled cells and exclude over 99 % of the unlabelled cells (Fig. 1C). The percentage of red fluorescence-positive MDM detected at 585 nm under this setting was defined as the phagocytic index.
Statistical analysis
Data are expressed as means ± SEM. The Mann-Whitney U test was used to analyze the non-normally distributed data, and Spearman rank correlation was used to assay the relevance between annexin V positivity and phagocytic index. GraphPad Prism (version 3.0; GraphPad Software, San Diego, CA) was used for all the statistical analyses. Statistical significance is set at p<0.05.
Results
HL-60 transfectants are able to stably express EGFP-A1 fusion protein.
The green fluorescence that gauged the expression of the ectopic EGFP-A1 fusion protein was detected by confocal microscopy (Fig. 2).
Overexpression of A1 in HL-60 provides cytoprotection against serum withdrawal-induced cell death.
To confirm the cytoprotective function of the EGFP-A1 fusion protein, this investigation first challenged the A1 transfectants with serum starvation. The data showed that A1 transfectants provided protection against cell death death triggered by 48 hours of serum- deprivation, as measured by total dye-excluding cell counts (Fig. 3A), cell hypodiploidy (Fig.
3B), and surface annexin-V binding activity (Fig. 3C) - complementary methodologies that assess viability, DNA fragmentation, and phosphoserine exteriorization during apoptosis, respectively.
Phagocytosis index was correlated with the degree of apoptosis in the HL60-EGFP control but not in the HL60-A1 transfectants.
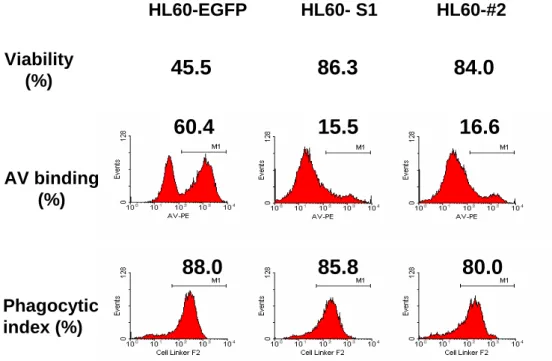
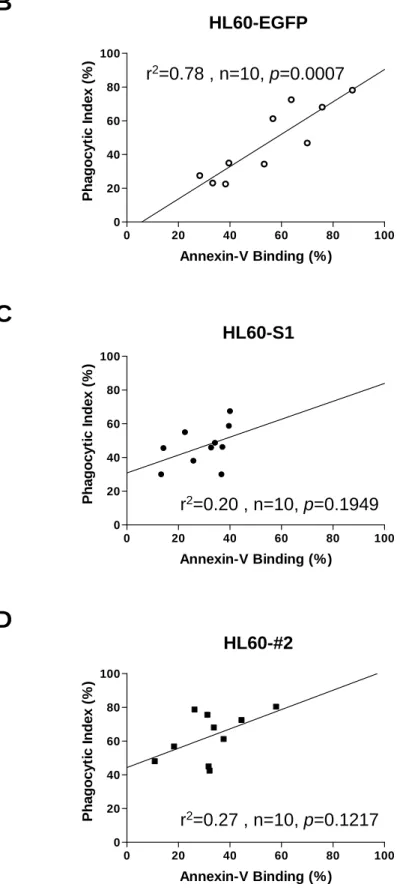
Using a serum-free medium to trigger HL60 apoptosis, the data presented here indicated that the phagocytic index of the HL60-A1 transfectants, despite having a significantly lower degree of apoptosis, did not differ from that of the HL60-EGFP control (Fig. 4A). Further study revealed that while the phagocytic index of the HL60-EGFP control was significantly correlated with the degree of apoptosis, the phagocytic index of the HL60-A1 transfectants was less relevant to apoptosis (HL60-A1 transfectants: S1, r2=0.20, n=10, p=0.1049; #2, r2=0.27, n=10, p=0.1217, versus HL60-EGFP control: r2=0.78, n=10, p=0.0007, Spearman rank
correlation). (Figs. 4B, 4C, and 4D). Furthermore, although the HL60-A1 transfectants had lower levels of annexin V-positive cells than the HL60-EGFP control, their phagocytic indices were comparable or higher (Figs. 4B, 4C, and 4D). To determine whether the difference in the phagocytic index arises from the difference in the efficiency of uptaking the apoptotic cells by MDM, apoptotic HL60 cells of the EGFP control and A1 transfectrants were isolated using magnetic beads conjugated with annexin V and then labeled with the red fluorescent cell linker.
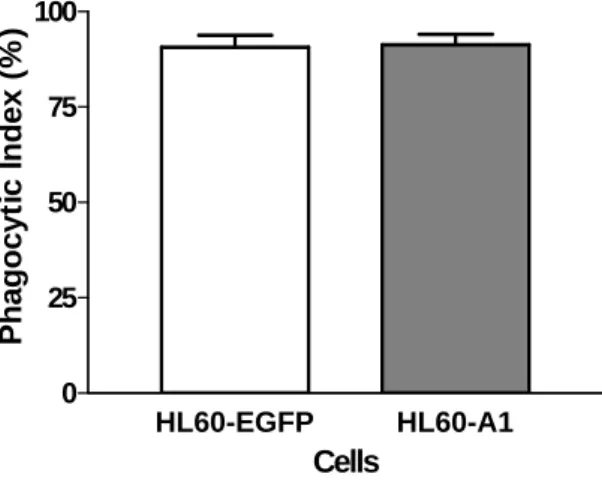
This study revealed no significant difference in the phagocytic index of annexin V-positive cells between the HL60-A1 transfectants and the control cells, suggesting that the annexin V- negative non-apoptotic fraction contributed to the enhanced phagocytic recognition in the HL60-A1 transfectants (Fig. 5).
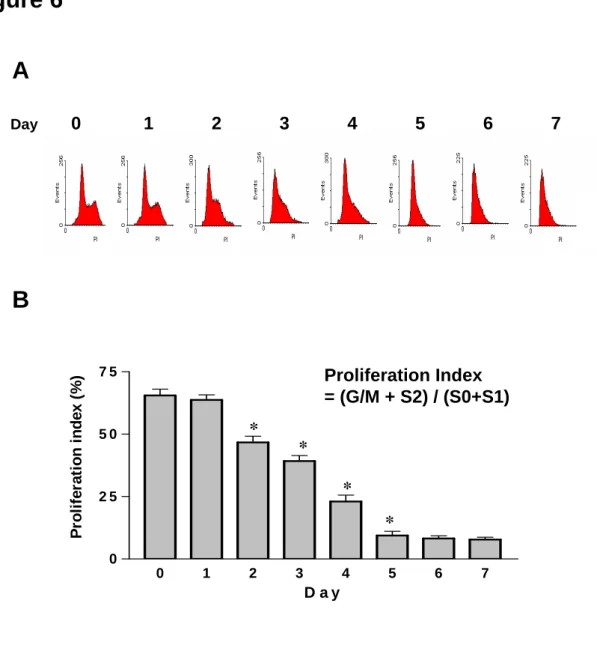
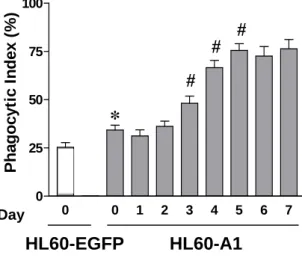
Phgocytosis index was increased in the senescent non-apoptotic HL60-A1 transfectants Further analysis of the cell cycle progression using flow cytometry revealed that the proliferation index decreased after two days of serum withdrawal, while proliferation ceased and the cells became quiescent on day five in HL60-A1 transfectants under serum-free conditions (Figs. 6A and 6B), suggesting an aging process during the serum-withdrawal period. There was no visible intracellular granule under light microscopy. Phagocytosis assay was performed in the isolated annexin V-negative A1 transfectants cultured in serum-free medium for various periods. The data showed that the phagocytic index of these cells increased with the number of days in culturing (Fig. 7).
Phagocytosis of the non-apoptotic senescent HL60-A1 transfectants was mediated by the CD14-, CD36-, avb3-, and scavenger receptor-independent mechanism
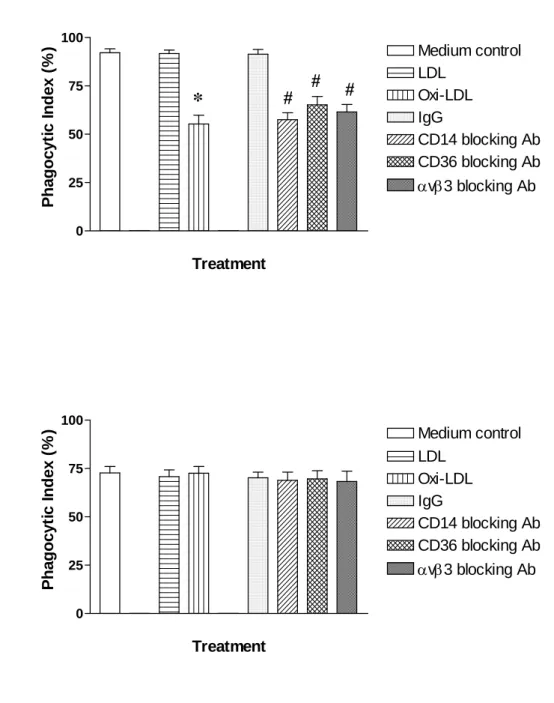
To clarify the mechanisms of MDM-mediated phagocytic clearance in the apoptotic and the non-apoptotic senescent HL60 cells, the MDMs were pretreated for one hour with blocking
agents, including anti-CD14, anti-CD36, and anti-αvβ3 antibodies, as well as the scavenger receptor inhibitor, Oxi-LDL. Cells were cultured for three days to isolate the annexin V- positive cells from the HL60-A1 transfectants and the EGFP control. Meanwhile, the HL60-A1 cells were cultured for five days to isolate the senescent annexin V-negative cells (The HL60- EGFP control was intolerable for serum-withdrawal, so the senescent non-apoptotic cells was unable to obtain). The data revealed that these inhibiting molecules significantly attenuated the phagocytic index in the annexin V-positive cells of HL60-A1 transfectants (Fig. 8A) and EGFP-control cells, both to a similar degree (data not shown), while no significant blocking effect existed for the senescent, annexin V-negative cells (Fig. 8B). The results presented herein thus suggest that the phagocytic clearance of the senescent non–apoptotic HL60-A1 transfectants was mediated by a different mechanism from that for apoptotic cells.
Discussion
The experiments conducted herein demonstrated that senescent HL60-A1 transfectants was able to be recognized and phagocytosed through an apoptosis-independent mechanism.
Lagasse’s study revealed significantly enhanced survival of the Bcl-2 transgenic neutrophils, making it likely that neutrophils homeostasis would also be influenced. However, flow cytometry analysis demonstrated that the proportion of neutrophils in the blood did not differ significantly between the transgenic and control mice [13]. Nor did significant differences exist in the total number of neutrophils or the proportion of myeloid cells in the bone marrow and spleen. Consequently, constitutive expression of Bcl-2 during myeloid cell maturation did not cause cell accumulation [13], suggesting that a mechanism of phagocytic clearance independent of apoptosis may operate for the homeostasis of blood cells [13]. As an anti-apoptotic Bcl-2 homologue, A1 is important in regulating the hematopoiesis of lymphoid and myeloid lineages, as well as in embryonic development [22, 23]. A1 is induced during the neutrophilic differentiation driven by G-CSF and GM-CSF in murine bone marrow cell lineage.
[22]. Studies have implied that A1 may have important role in modulating cell proliferation, transformation and inflammatory responses [17, 18]. Furthermore, the cytoprotective effect of A1 may provide enhanced survival capacity to neutrophils in inflammatory situations [5]. This study found the EGFP control and A1-expressing HL-60 cells did not differ in the phagocytic index of apoptotic cells. However, although freshly isolated, non-apoptotic HL60 cells were recognized and phagocytosed by the macrophages in a small degree, the senescent, non- apoptotic HL60-A1 tansfectants triggerred by serum deprivation were vigorously phagocytosed. Furthermore, the baseline degree of phagocytic recognition of non-apoptotic A1-transfected cells also exceeded that of the control cells. The results of this study thus confirmed that, independently of apoptosis, senescent non-apoptotic cells may exhibit cell
surface changes that allow them to be recognized and engulfed by macrophages and is, possibly, associated with the function of Bcl-2 homologue, A1.
To date, several different mechanisms have been described for recognizing apoptotic cells. Recognition of apoptotic cells is mediated by various receptors, including lectins, CD14, scavenger receptor A (SR-A), and CD36, in conjunction with the vitronectin receptor (αvβ3) [24-31]. Ligands on the apoptotic cell that are recognized by these receptors include sugars, phosphatidylserine (PS), and surface-bound thrombospondin (TSP) [32, 33]. Both N-acetyl glucosamine or N-acetyl galgactosamine can inhibit the uptake of apoptptic murine thymocytes, sugesting a lectin-type interaction [34, 35]. Apoptotic human neutrophils were phagocytosed by blood- or bone marrow-macrophages using thrombospodin-1 (TSP-1) bridging between a charge-sensitive moiety on the apoptotic cell and a complex formed by the intergrin αvβ3 (the fibronectin receptor) and CD36 on the macrophages [36-41] . Meanwhile, phosphatidylserine (PS) externalized on apoptotic neutrophils was recognized by a still uncharacterized receptor on certain murine macrophages from peritoneal exudates, whereas blood-derived and bone marrow-derived macrophages utilized TSP/αVβ3/CD36 to recognize apoptotic cells [26, 42-44]. Human fibroblasts were demonstrated to bind apoptotic neutrophils partially by a mechanism inhibited by mannose or fucose, suggesting a mannose/fucose- specific lectin on the fibroblast [31, 45]. A macrophage receptor identified by the 61D3 mAb is involved in the recognition of apoptotic neutrophils [46, 47]. The class A macrophage scavenger receptor was involved in the phagocytosis of apoptotic thymocytes [20, 48-50].
Apoptotic endothelial cells (EC) are recognized by mononucleatr phagocytes through a novel β1 integrin-dependent mechanism [5, 51] Additionally, the unexpected finding that Bcl-2 over- expression reduced cell apoptosis but did not block the recognition of of cells by macrophages [13] suggests that recognition may result from changes produced by senescence only. Although investigations have demonstrated that aging of cell membrane molecules could lead to the
emergence of an aging antigen and the removal of senescent cells [52-54], the mechanism by which the senescent non-apoptotic cells are recognized remains poorly understood [11, 55].
Our data showed that blocking recognition molecules on MDM, including CD14, CD36, vibronectin and scavanger receptors, could attenuate the phagocytic clearance of apoptotic HL60 cells but not of the senescent non-apoptotic HL60 A1 transfectants, further confirming that an apoptosis-independent recognition mechanism exists in the senescent cell clearance.
With the findings that a variety of mediators in an inflammatory environment were able to prolong the neutrophil survival [5] and that the apoptosing neutrophils were still releasing active mediators and causing lung epithelial cells damage [6], our results speculated an apoptosis-independent phagocytic clearance mechanism to attenulate the progression of inflammation.
In conclusion, the experiments conducted in this study demonstrated that Bcl-2-related protein, A1 could inhibit programmed cell death of HL60 cells, but could not prevent the cells from being engulfed by macrophages. The recognition and ingestion of senescent, non- apoptotic cells by macrophages is, possibly, a primary mechanism of eradicating aged cells, maintaining tissue homeostasis and relieving inflammation.
Acknowledgments
The authors would like to thank the National Science Council of Taiwan for financially supporting this research under Contract No. NSC 93-2314-B-182A-137 and 93-2314-B-038- 020, and Chang-Gung Memorial Hospital under Contract No. CMRPG32113
References
1. Schlegel, R. A., S. Krahling, M. K. Callahan, P. Williamson. CD14 is a component of multiple recognition systems used by macrophages to phagocytose apoptotic lymphocytes.
Cell Death Differ 1999; 6:583-592.
2. Grigg JM, Savill JS, Sarraf C, Haslett C, Silverman M. Neutrophil apoptosis and clearance from neonatal lungs. Lancet 1991; 338:720-2.
3. Jones ST, Denton J, Holt PJ, Freemont AJ. Possible clearance of effete polymorphonuclear leucocytes from synovial fluid by cytophagocytic mononuclear cells:
implications for pathogenesis and chronicity in inflammatory arthritis. Ann Rheum Dis 1993; 52:121-6.
4. Savill J. Recognition and phagocytosis of cells undergoing apoptosis. Br Med Bull 1997;
53:491-508.
5. Chuang PI, Yee E, Karsan A, Winn RK, Harlan JM. A1 is a constitutive and inducible Bcl-2 homologue in mature human neutrophils. Biochem Biophys Res Commun 1998;
249:361-5.
6. Liu CY, Liu YH, Lin SM, Yu CT, Wang CH, Lin HC, Lin CH, and Kuo HP. Apoptotic neutrophils undergoing secondary necrosis induce human lung epithelial cell detachment.
J Biomed Sci 2003; 10:746-56.
7. Ren Y, Savill J: Proinflammatory cytokines potentiate thrombospondin-mediated phagocytosis of neutrophils undergoing apoptosis. J Immunol 1995; 154:2366-74.
8. Savill J. Apoptosis: Phagocytic docking without shocking. Nature 1998; 392:442-3; 1998.
9. Savill J, Fadok V. Corpse clearance defines the meaning of cell death. Nature 2000;
407:784-8.
10. Savill JS, Wyllie AH, Henson JE, Walport MJ, Henson PM, Haslett C. Macrophage phagocytosis of aging neutrophils in inflammation. Programmed cell death in the neutrophil leads to its recognition by macrophages. J Clin Invest 1989; 83:865-75.
11. Daemen MA, van 't Veer C, Denecker G, Heemskerk VH, Wolfs TG, Clauss M, Vandenabeele P, Buurman WA. Inhibition of apoptosis induced by ischemia-reperfusion prevents inflammation. J Clin Invest 1999; 104:541-9.
12. Hockenbery DM, Zutter M, Hickey W, Nahm M, Korsmeher SJ. Bcl-2 protein is topographically restricted in tissues characterized by apoptotic cell death. Proc Natl Acad Sci USA 1991; 88:6961-5.
13. Lagasse E, Weissman IL.Bcl-2 inhibits apoptosis of neutrophils but not their engulfment by macrophages. J Exp Med 1994; 179:1047-52.
14. Savill J. Macrophages recognition of senescent neutrophils. Clin Sci 1992; 83:649-55.
(14)
15. Cocco RE, Ucker DS. Distinct Modes of Macrophage Recognition for Apoptotic and Necrotic Cells Are Not Specified Exclusively by Phosphatidylserine Exposure. Mol Biol Cell 2001; 12: 919-30.
16. Hess KL, Babcock GF, Askew DS, Cook-Mills JM. A novel flow cytometric method for quantifying phagocytosis of apoptotic cells. Cytometry 1997; 27:145-52.
17. Karsan A, Yee E, Harlan JM. Endothelial cell death induced by tumor necrosis factor-α is inhibited by the Bcl-2 family member, A1. J Biol Chem 1996; 271:27201-4.
18. Karsan A, Yee E, Kaushansky K, and Harlan JM. Cloning of a human bcl-2 homologue:
inflammatory cytokines induce human A1 in cultured endothelial cells. Blood 1996;
87:3089-96.
19. Liu CY, Chuang PI, Chou CL, Lin SM, Chen CH, Chou PC, Liu YH, Yu CT, Wang CH, Lin HC, Kuo HP. Cytoprotective response of A1, a Bcl-2 homologue expressed in mature
human neutrophils and promyelocytc HL-60 cells, to oxidant stress-induced cell death. J Biomed Sci 2004; 11:214-27.
20. Pahl HL, Burn TC, Tenen DG. Optimization of transient transfection into human myeloid cell lines using a luciferase reporter gene. Exp Hematol 1991; 19:1038-41.
21. Liu CY, Wang CH, Chen TC, Lin HC, Yu CT, Kuo HP. Increased level of exhaled nitric oxide and upregulation of nitric oxide synthase in alveolar macrophages in patients with primary lung cancer. British Journal of Cancer 1998; 78:534-41.
22. Lin EY, Orlofsky A, Berger MS and Prystowsky MB. Characterization of A1, a novel hemopoietic-specific early-response gene with sequence similarity to bcl-2. J Immunol 1993; 151:1979-88.
23. Lin EY, Orlofsky A, Wang HG, Reed JC, Prystowsky MB. A1, a Bcl-2 family member, prolongs cell survival and permits myeloid differentiation. Blood 1996; 87:983-92.
24. Albert ML, Pearce SF, Francisco LM, Sauter B, Roy P, Silverstein RL, Bhardwaj N.
Immature dendritic cells phagocytose apoptotic cells via αvβ5 and CD36, and cross- present antigens to cytotoxic T lymphocytes. J Exp Med 1998; 188:1359-68.
25. Devitt A, Moffatt OD, Raykundalia C, Capra JD, Simmons DL, Gregory CD. Human CD14 mediates recognition and phagocytosis of apoptotic cells. Nature 1998; 392:505-9.
26. Fadok VA, de Cathelineau A, Daleke DL, Henson PM, Bratton DL. Loss of phospholipid asymmetry and surface exposure of phosphatidylserine is required for phagocytosis of apoptotic cells by macrophages and fibroblasts. J Biol Chem 2001; 276:1071-7.
27. Gregory CD. CD14-dependent clearance of apoptotic cells: relevance to the immune system. Curr Opin Immunol 2000; 12:27-34.
28. Hughes J, Liu Y, Van Damme J, Savill J. Human glomerular mesangial cell phagocytosis of apoptotic neutrophils: mediation by a novel CD36-independent vitronectin
receptor/thrombospondin recognition mechanism that is uncoupled from chemokine secretion. J Immunol 1997; 158:4389-97.
29. Krieger M, Herz J. Structures and functions of multiligand lipoprotein receptors:
macrophage scavenger receptors and LDL receptor-related protein (LRP). Annu Rev Biochem 1994; 63:601-37.
30. Ogden CA, deCathelineau A, Hoffmann PR, Bratton D, Ghebrehiwet B, Fadok VA, Henson PM. C1q and mannose binding lectin (MBL) engagement of cell surface calreticulin and CD91 to initiates macropinocytosis and uptake of apoptotic cells. J Exp Med 2001; 194:781-95.
31. Schlesinger LS. Macrophage phagocytosis of virulent but not attenuated stains of Mycobacterium tuberculosis is mediated by mannose receptors in addition to complement receptors. J Immunol 1993; 150:2920-30.
32. Balasubramanian K, Chandra J, Schroit AJ. Immune clearance of phosphatidylserine- expressing cells by phagocytes: The role of beta2-glycoprotein I in macrophage recognition. J Biol Chem 1997; 272:31113-7.
33. Bennett MR, Gibson DF, Schwartz SM, Tait JF. Binding and phagocytosis of apoptotic vascular smooth muscle cells is mediated in part by exposure of phosphatidylserine. Circ Res 1995; 77:1136-42.
34. Duvall E, Wyllie AH, Morris RG. Macrophage recognition of cells undergoing programmed cell death (apoptosis). Immunology 1985; 56:351-8.
35. Pearson AM. Scavenger receptors in innate immunity. Curr Opin Immunol 8:20-28; 1996.
36. Greenwalt DE, Lipsky RH, Ockenhouse CF, Ikeda H, Tandon NN, Jamieson GA Membrane glycoprotein CD36: a review of its roles in adherence, signal transduction, and transfusion medicine. Blood 1992; 80:1105-15.
37. Pommier CG, Inada S, Fries LF, Takahashi T, Frank MM, Brown EJ. Plasma fibronectin enhances phagocytosis of opsonized particles by human peripheral blood monocytes. J Exp Med 1983; 157:1844-54.
38. Pradhan D, Krahling S, Williamson P, Schlegel RA. Multiple systems for recognition of apoptotic lymphocytes by macrophages. Mol Biol Cell 1997; 8:767-78.
39. Savill J, Dransfield I, Hogg N, Haslett C. Vitronectin receptor-mediated phagocytosis of cells undergoing apoptosis. Nature 1990; 343:170-3.
40. Savill J, Fadok V, Henson P, Haslett C. Phagocyte recognition of cells undergoing apoptosis. Immunol Today 1993; 14:131-6.
41. Savill J, Hogg N, Ren Y, Haslett C. Thrombospondin cooperates with CD36 and the vitronectin receptor in macrophage recognition of neutrophils undergoing apoptosis. J Clin Invest 1992; 90:1513-22.
42. Fadok VA, Bratton DL, Henson PM. Phagocyte receptors for apoptotic cells: recognition, uptake, and consequences. J Clil Invest 2001; 108:957-62.
43. Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DL, Henson PM. Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. J. Immunol 1992; 148:2207-16.
44. Fadok VA, Warner ML, Bratton DL, Henson PM. CD36 is required for phagocytosis of apoptotic cells by human macrophages that use either a phosphatidylserine receptor or the vitronectin receptor (αvß3). J Immunol 1998; 161:6250-7.
45. Taylor PR, Brown GD, Herre J, Williams DL, Willment JA, Gordon S.The role of SIGNR1 and the beta-glucan receptor (dectin-1) in the nonopsonic recognition of yeast by specific macrophages. J Immunol 2004; 172:1157-62.
46. Murphy JF, McGregor JL, Leung LL. Senescent human neutrophil binding to thrombospondin (TSP): evidence for a TSP-independent pathway of phagocytosis by macrophages. Br J Haematol 1998; 102:957-64.
47. Navazo MD, Daviet L, Savill J, Ren Y, Leung LL, McGregor JL. Identification of a domain (155-1830) on CD36 implicated in the phagocytosis of apoptotic neutrophils. J Bio Chem 1996; 271:15381-5.
48. Platt N, da Silva RP, Gordon S. Recognizing death: the phagocytosis of apoptotic cells.
Trends Cell Biol 1998; 8:365-72.
49. Platt N, Gordon S. Is the class A macrophage scavenger receptor (SR-A) multifunctional?
- The mouse’s tale. J Clin Invest 2001; 108:649-54.
50. Platt N, Haworth R, Darley L, Gordon S. The many roles of the class A macrophage scavenger receptor. Int Rev Cytol 2002; 212:1-40.
51. Zen K, Karsan A, Stempien-Otero A, Yee E, Tupper J, Li X, Eunson T, Kay MA, Wilson CB, Winn RK, Harlan JM. NF-kappaB activation is required for human endothelial survival during exposure to tumor necrosis factor-alpha but not to interleukin-1beta or lipopolysaccharide. J Biol Chem 1999; 274:28808-15.
52. Hirt UA, Gantner F, Leist M. Phagocytosis of nonapoptotic cells dying by caspase- independent mechanisms. J Immunol 2000; 164:6520-9.
53. Kay MM. Aging of cell membrane molecules leads to appearance of an aging antigen and removal of senescent cells. Gerontology 1985; 31:215-35.
54. Newman SL, Henson SE, Henson PM. Phagocytois of senescent neutrophils by human monocyte-derived macrophages and rabbit inflammatory macrophages. J Exp Med 1982;
156:430-5.
55. Stern M, Savill J, Haslett C. Human monocyte-derived macrophage phagocytosis of senescent eosinophils undergoing apoptosis. Mediation by αvβ3/CD36/thrombospondin recognition mechanism and lack of phlogistic response. Am J Pathol 1996; 149:911-21.
Legends
Figure 1. Flow cytometry can determine the extent of phagocytosis of MDM. (A) Flow cytometric analysis utilizing density plots of forward scatter (FSC) versus sideward scatter (SSC) clearly distinguished the MDM from the HL60 cells owing to their different size and granularity. (B) To prevent bias resulting from the innate green fluorescence in the transfectants, three clones of HL60-EGFP control and HL60-A1 transfectants-S1 and #2 with similar fluorescent intensity at FL1 (530nm) were chosen for study (upper panel). Further investigaton of the fluorescent interference of EGFP at FL2 (585 nm, Protocols for detemining the annexin V positivity and phagocytic index, MDM PhagoIndex) revealed no significant scattering of green fluorescence on FL2 (middle and lower panels). (C) HL60 cells were heavily labeled with the red fluorescent cell linker, resuling in a two to three log increase of the fluorescent intensity of the cells. An arbitrary positivity cutoff value was established to include more than 98 % of fluorescent positive events and less than 1 % of fluorescent negative events.
Consequently, the percentage of MDM gated from the density plots (FSC vs. SSC) containing red fluorescence indicated the extent of phagocytosis (phagocytic index).
Figure 2. HL-60 transfectants exhibited stable EGFP-A1 fusion protein expression. The green fluorescence that indicated the expression of the ectopic EGFP-A1 fusion protein was detected in the cytoplasma of cells using confocal microscopy.
Figure 3. Overexpression of A1 protected against serum withdrawal-induced cell death. Cells of parental HL-60, EGFP-HL-60 control, pooled A1 and single A1 transfectants, at initial cell number of 1 x 106, were cultured in 2 ml of serum-free EMEM medium for 24 and 48 h. Cell death was assayed by total viable cell counts (A), DNA content analysis (B), and annexin V
binding (C). Data were expressed as the mean ± SEM of five independent experiments.
*p<0.01compared with the parental or EGFP-transfected HL-60 cells in the same subgroup;
#p<0.01 compared with the parental or EGFP-transfected HL-60 cells in the same subgroup.
Figure 4. The phagocytic index was correlated with the degree of apoptosis in HL60-EGFP control but not in HL60-A1 transfectants. (A) To perform the phagocytosis assay, HL60 cells were co-cultured with MDM for one hour, and serum-free medium was used to trigger HL60 apoptosis. Although the HL60-A1 transfectants had a significantly lower rate of cell death (upper and middle panels), their phagocytic indices were similar to those of the HL60-EGFP control (lower panel). (One reprsentative data from at least five experiments is shown here). A significant correlation existed between the degree of apoptosis (annexin V positivity) and the extent of phagocytic clearance of cells (phagocytic index) in the HL60-EGFP control (B), but not in the HL60-A1 transfectants (C and D). Spearman rank correlation was used to study the relavance between the degree of apoptosis and the phagocytic index.
Figure 5. Following three days of serum withdrawal, annexin V-positive cells were isolated using annexin V-conjugated paramagnetic microbeads. A phagocytosis assay was performed after labeling the apoptotic cells with the red fluorescent cell linker, and revealed no significant difference in the phagocytic recognition of the annexin V-positive cells between the EGFP- and A1- transfectants. (Data shown are means ± SEM, N=6, Mann-Whitney U test).
Figure 6. Serum withdrawal reduced cell growth and stopped cell proliferation. (A) Flow cytometric analysis using CellQuest Software demonstrated that the S/M and S2 fractions of HL60A1 transfectants reduced following serum-deprivation, suggesting the cell cycle was suppressed under serum-withdrawal. (B) Further quantitative analysis of the cell cycle
progression indicated that the proliferation index significantly declined after two days of serum withdrawal, and the cells turned quiescent on day five of culture, suggesting the occurance of an aging process during the serum-withdrawal period. Data shown are means ± SEM. * p<0.05, N=6, compared to the index one day earlier, Mann-Whitney U test.
Figure 7. HL60-A1 transfectants were cultured in serum-deprived medium for periods ranging from several days to one week. The annexin V-negative fraction of cells was isolated using annexin V-conjugated paramagnetic microbeads. Following labeling with the red fluorescent cell linker, cells were co-cultureed with MDM for phagocytosis assay. The phagocytic index of the annexin V-negative HL60-A1 transfectants was significantly higher than that of the EGFP- control cells and increased with the number of days of culturing. Data shown are means ± SEM. * p<0.05, compared to HL60-EGFP control; # p<0.05, compared to the index one day earlier; N=6, Mann-Whitney U test.
Figure 8. The mechanisms of phagocytic recognition operate differently in apoptotic and senescent non-apoptic HL60 cells. HL60-A1 transfectants were cutured in serum-withdrawal medium for three days to isolate the annexin V-positive cells and for five days to isolate the annexin V-negative fraction. Precubation of the MDM with agents blocking the recognition molecules of CD14-, CD36-, avb3-, and scavenger receptor could attenuate the extent of phagocytic clearance of annexin V-positive HL60-A1 transfectants (A) but not of the annexin V-negative cells (B). Data shown are means ± SEM. * p<0.05, compared to LDL control; # p<0.05, compared to the IgG control; N=6, Mann-Whitney U test.
Figure 1
MDM HL60
A
B
HL60-EGFP HL60-S1 HL6- # 2HL60-EGFP (MFI)
15.21 18.07
HL60 AV-PE (% of Positivity)
6.4
MDM PhagoIndex
(% of Positivity) 0.8
Protocol
1.1
0.8
HL60-EGFP HL60-EGFP labeled with red fluorescent cell linker
98.7
C
5.0 6.6
15.86
1.2
Figure 2
A Figure 3
24 48
0 25 50 75 100 125 150
*
#*
#Time (hr) Total viable cell count (% of initial)
24 48
0 25 50 75
100 HL 60-ATCC
HL60-GFP Pooled clone Individual clone
*
#*
#Time (hr)
Hypodiploidy (%)
24 48
0 25 50 75 100
*
#*
#Annexin-V binding (%)
B
C
Figure 4
A
HL60-EGFP HL60- S1 HL60-#2
Viability
(%) 45.5 86.3 84.0
AV binding (%)
60.4
Phagocytic index (%)
88.0 85.8 80.0
16.6 15.5
Figure 4 B
C
D
HL60-EGFP
0 20 40 60 80 100
0 20 40 60 80 100
Annexin-V Binding (%)
Phagocytic Index (%)
r2=0.78 , n=10, p=0.0007 HL60-EGFP
HL60-S1
0 20 40 60 80 100
0 20 40 60 80 100
Annexin-V Binding (%)
Phagocytic Index (%)
HL60-#2
0 20 40 60 80 100
0 20 40 60 80 100
Annexin-V Binding (%)
Phagocytic Index (%)
r2=0.27 , n=10, p=0.1217 r2=0.20 , n=10, p=0.1949
HL60-S1
HL60-#2
Figure 5
HL60-EGFP HL60-A1 0
25 50 75 100
Cells
Phagocytic Index (%)
Figure 6 A
B
0 1 2 3 4 5 6 7
0 2 5 5 0 7 5
D a y
Proliferation index (%)
*
*
*
Proliferation Index
= (G/M + S2) / (S0+S1)
* 1
Day 0 2 3 4 5 6 7
Figure 7
0 0 1 2 3 4 5 6 7
0 25 50 75 100
Phagocytic Index (%)
*
#
#
#
HL60-EGFP HL60-A1
Day
Figure 8 A
B
0 25 50 75 100
Medium control LDL
Oxi-LDL IgG
CD14 blocking Ab CD36 blocking Ab αvβ3 blocking Ab
Treatment
Phagocytic Index (%)
* #
#
#
0 25 50 75 100
Medium control LDL
Oxi-LDL IgG
CD14 blocking Ab CD36 blocking Ab αvβ3 blocking Ab
Treatment
Phagocytic Index (%)