Correspondence and requests for reprints:Dr. Shih-Chung Chen
Address:Division of Internal Medicine, Taipei County Hospital, Sanchong Branch, No. 2, Jhongshan Rd., Sanchong City, Taipei County 241, Taiwan
Fenofibrate Represses Matrix
Metalloproteinase-2 in Endothelial Cells
Chia-Hui Yang
1, Yin-Cih Chao
2, Ting-Wuan Lin
3, Jau-Kang Huang
3, Hsi-Che Shen
4, Danny Ling Wang
5, and Shih-Chung Chen
3,61
Division of Endocrinology and Metabolism,
2
Division of Allergy, Immunology and Rheumatology,
3
Division of Cardiovascular Medicine,
4
Division of Emergency Medicine, Taipei County Hospital;
5
Cardiovascular Division, Institute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan;
6
Graduate Institute of Medical Sciences, College of Medicine, Taipei Medical University
Introduction
Fenofibrate is mainly used to reduce trigly- cerides levels in patients at risk of cardiovascular disease (CVD), including atheroclerosis and co- ronary heart disease. Several clinical trials, such as the Diabetes Atherosclerosis Intervention Study (DAIS) revealed that treatment with fenofibrate
reduces the angiographic progression of coronary- artery disease in type 2 diabetes
1. Similar to other fibric acid, it reduces low-density lipoprotein (LDL), very low density lipoprotein (VLDL) and triacylglycerides (TG) levels, whereas increases high-density lipoprotein (HDL) level. Endothelial cells (ECs), constituting the endothelium, the first
Abstract
Fenofibrate is a widely used to ameliorate hyperlipidemia and/or hypercholesterolemia in patients at risk of cardiovascular disease. To date, most of its favorable effects have been attributed to its activation of peroxisome proliferator activated receptor alpha (PPARα), which alters lipid metabolism, contributing to an improved lipid profile. Lines of evidence suggest pleotrophic effects of fenofibrate in cardiovascular system. Given that endothelium is the frontier of vasculature and its dysfunction is prelude of various vascular disorders, e.g. atherosclerosis and arterial stiffening, we investigated the effect of fenofibrate in human endothelial cell (EC) line (Eahy926). Fenofibrate treatment in ECs repressed matrix metalloproteinase-2 (MMP-2) expression in ECs as well as endothelial MMP-2 secretion, associated with decreased MMP-2 activity. In parallel, fenofibrate activated endothelial nitric oxide synthase (eNOS), demonstrated by Ser1177 phosphorylation. Inhibition of eNOS by L-NAME attenuated fenofibrate- suppressed MMP-2, indicating fenofibrate inhibits MMP-2 requires eNOS-derived NO. Collectively, these findings suggest that fenofibrate activates eNOS and increases NO-bioavailability, which in turn suppresses overactivation of MMP-2, contributing to maintenance of endothelial homeostasis. The present study provides a molecular rationale in which fenofibrate exerts vascular protective effects, facilitating the clinical application of its derived medicine.
( J Intern Med Taiwan 2010; 21: 270-276 )Key Words: Fenofibrate, MMP-2, Endothelial Cell, eNOS, Nitric oxide
layer of blood vessels, provide the interface between the vessel wall and the blood circulation.
Endothelial homeostasis exerts various vasoprote- ctive effects, including vasodilation, anti-inflamm- ation, as well as suppression of vascular smooth muscle cell (VSMC) proliferation and hypertrophy.
The impaired endothelial homeostasis is an early marker for vascular pathogenesis, leading to atherosclerosis or arterial stiffening
2,3. To prevent these pathophysiological alterations, two molecular components, i.e. endothelial nitric oxide synthase (eNOS)-derived NO bioavailability and matric metalloproteinases (MMPs), play major roles in maintaining the homeostasis of ECs, and extra- cellular matrix (ECM), the dynamic interactive milieu between ECs and VSMCs
4,5.
Vascular ECM is tightly regulated by MMPs, a family of zinc-dependent protease, which catalyze the degradation of collagen and other ECM com- ponents. Under stimulation such as oxidative stress and plasminogen activators, MMPs are upregulated or activated. i.e. the pro-MMPs converted to active MMPs
6,7. Lines of evidence suggest a hierarchical pathway in which the eNOS-derived NO modulates MMP activity in the ECs and VSMCs
8,9.
In the present study, we investigated the ef- fects of fenofibrate in ECs, in terms of MMP expression and activity. Due to the role of eNOS in MMP modulation throught its effector NO, we also examined the possible mechanism by which fenofibrate may inhibit overactivation of MMPs, i.e. through posttranslational regulation of eNOS on its activation site Ser-1177. Collectively, our study provides an additional explanation for the vascular protective effects exerted by fenofibrate.
Methods
Antibodies and reagents
N
G-Nitro-L-arginine Methyl Ester (L-NAME) and mouse anti-human MMP-2 monoclonal anti- body were purchased from Calbiochem (San Diego,
CA). eNOS, and phospho-eNOS (Ser-1177) were purchased from BD Bioscience Pharmingen (San Diego, CA). anti- pan actin Ab-5 antibody (ACTN05) was obtained from NeoMarkers (Fremont, CA). Fenofibrate was obtained from Synkem (France). Gelatin was purchased from Sigma (St Louis, MO).
Cell culture
Eahy926 cell line was obtained commercially from American Type Culture Collection (Rockvi- lle, MD) and cultured in medium consisting of Dulbecco's Modified Eagle's Medium (DMEM;
Gibco, Gaithersburg, MD) supplemented with 10%
fetal bovine serum (FBS; Gibco). Cells were seeded on 60 mm Petri dishes until ~90% confluence, and the culture medium was then changed to DMEM containing 0.1% BSA prior to experiments.
Western blot analysis
ECs were washed in PBS and lysed with RIPA buffer containing Nonidet P-40 (1%), SDS (0.5%) and protease inhibitor cocktail. Cells were disrupted by intermittent sonication. After centrifugation, the protein level of supernatant was measured using BSA as standard. Cell lysates were then subjected to SDS-PAGE followed by Western blotting. Antigen- antibody complexes were detected by horseradish peroxidase-labeled rabbit anti-mouse or goat anti- rabbit IgG with an ECL detection system (Pierce, Rockford, IL).
Zymographic analysis of MMP-2
Confluent ECs were cultured in serum free
DMEM with or without fenofibrate (10 or 50μl)
for 24 hours. An equal amount (30μl) of con-
ditional culture medium was applied to gel. The
activities of MMP-2 were analyzed by gelatin
zymography. In brief, the conditional medium was
applied to 10 % SDS-polyacrylamide gel containing
0.1 % gelatin. Commercially available MMP-2 was
used as a reference. After electrophoresis, gels were
incubated in Zymogram Renaturing Buffer (2.5 %
triton X-100) for 1 hour, and then equilibrate the
gels in 1X Zymogram developing buffer (0.05 M Tris-HCl, pH 7.5, 0.02 M NaCl, 5 mM CaCl
2, and 0.02% Brij-35) at room temperature with gentle agitation for 30minutes. Gels were transferred to fresh 1X Zymogram Developing Buffer and incubated at 37 ℃ for 24 hours. The gels were stained with 0.5 % coomassie brilliant blue and destained with destaining solution ( methanol : acetic acid : water = 50 : 10 : 40 ). The gels were scanned using the UMAX 1220S scanner with a photo-mask (UMAX Corp.).
RNA isolation and real-time quantitative PCR Total cellular RNA was extracted from ECs (~1.5 x 10
6cells) using TRI Reagent (Molecular Research Center, Cincinnati, OH) according to manufacturer's instruction. One microgram of total RNA was reverse-transcribed by M-MuLV reverse transcriptase (Finnzyme, Finland) in a total volume of 20μl. The reverse transcriptase products were amplified with DyNAmo HS SYBR Green qPCR Kit (Finnzyme, Finland) in the reaction mix co- ntaining DyNAmo SYBR Green master mix and primers. Primers were designed for detection of human MMP-2 and GAPDH gene expression, as below:
For MMP-2,
forward: 5'-TGGCAAGTACGGCTTCTGTC-3';
reverse: 5'-TTCTTGTCGCGGTCGTAGTC -3'.
For GAPDH:
forward: 5'-GAGTCAACGGATTTGGTCGT -3';
reverse: 5'-GACAAGCTTCCCGTTCTCAG-3'.
Statistical analysis
Results are expressed as means ± SEM.
Statistical analysis was performed by using an independent Student's t-test for two groups of data.
P < 0.05 was considered statistically significant.
Results
Fenofibrate downregulates MMP-2 expression in ECs
Because MMP-2 is the predominant MMP
secreted from ECs
10, we examined the expression of MMP-2 in ECs with or withour fenofibrate treatment. As seen in Fig. 1, the MMP-2 protein level was significantly reduced by fenofibrate treatment in a dose-dependent manner. To further examine whether MMP-2 expression is regulated at transcriptional level, the quantitative realtime PCR revealed that MMP-2 mRNA level was also decreased by fenofibrate treatment (Fig. 2), con- sistent with the protein level change.
Fenofibrate suppresses MMP-2 secretion from ECs
To evaluate whether fenofibrate's inhibitory effect in endothelial MMP-2 expression is as- sociated with decreased MMP-2 secretion as well as suppressed proteinase activity, we detected the MMP-2 released into EC culture medium, using gelatinase zymography. As expected, the activity Fig. 1.Fenofibrate decreases MMP-2 protein expre-
ssion in ECs.
Eahy926 cells were starved in 0.1% BSA DMEM for 16 hours before fenofibrate tre- atment. Eahy926 cells were cultured in the presence of fenofibrate (0, 10, 50μM) for 24 hr, and cells were lysed. 50μg cell lysate was loaded in each lane for Western blotting. The expression of MMP-2 was normalized to the expression of actin. Data are mean ± s.e.m.
of three independent experiments.
(*:p value < 0.05 **:p value < 0.01)
of secreted MMP-2 was also decreased in a dose- dependent manner by fenofibrate, as observed in (Fig. 3).
Fenofibrate repressed MMP-2 is requires eNOS-derived NO
To investigate the possible upstream signaling event for MMP-2 repression by fenofibrate, we asked whether NO bioavailability controlled by eNOS may play a central role. First, we detected the phosphorylation of eNOS Ser-1177, which was increased in both a dose-dependent(in a) as well as a time-dependent(in b) manner by fenofibrate treatment (Fig. 4). This increased eNOS pho- sphorylation indicates the activation of eNOS, which expectedly leads to an enhanced NO pr- oduction in ECs
11. Next, to find out whether eNOS- NO is a key component mediating fenofibrate- inhibited MMP-2, L-NAME was used to block the
activation of eNOS in ECs. As shown in (Fig. 5), L-NAME attenuated the fenofibrate-repressed MMP-2 secretion from ECs (in a); as well, L-NAME also reversed the fenofibrate-reduced MMP-2 expression in ECs (in b). Taken together, this finding suggests that fenofibrate requires eNOS-derived NO to repress MMP-2, both in its expression and its activation.
Discussion
In this study, we demonstrated that fenofibrate decreased MMP-2 level in human ECs, which process requires the activation of eNOS and hence enhanced NO bioavailability. Our observation that eNOS Ser-1177 phosphorylation is increased by fenofibrate is consistent with the previous finding that fenofibrate activates AMP-acitivated protein Fig. 2.Fenofibrate inhibits MMP-2 mRNA expression.
Eahy926 cells were starved in serum free me- dium overnight prior to fenofibrate stimulation.
Cells were then treated with fenofibrate (0, 10, 50μM) for 24 hours. RNA was isolated and was converted to cDNA. The expression of MMP-2 mRNA was analyzed by real time PCR. GADPH mRNA expression was used for normalization in real time PCR. Data are mean
± s.e.m. of three independent experiments.
( #:p value < 0.05 *:p value < 0.01 )
Fig. 3.Fenofibrate suppresses MMP-2 secretion in ECs. Eahy926 cells were starved in 0.1% BSA DMEM for 16 hours before fenofibrate tre- atment. After cells were treated with fenofibrate (10 or 50μM) for 24 hr, culture medium was collected and 30μl culture medium was used in each lane for zymography. Data are mean
± s.e.m. of four independent experiments.
(#:p value < 0.05 *:p value < 0.01),
P.C.:recombinant MMP-2 as positive control.
kinase (AMPK) and eNOS phosphorylation in human umbilical vein ECs (HUVECs)
12. Due to the multiple upstream kinases of eNOS activation, such as protein kinase B/Akt
13,14, it is conceivable that other protein kinases may also be activated by fenofibrate and mediate the activation of eNOS.
Meanwhile, eNOS Ser-633 is another critical site, the phosphorylation of which has been suggested to contribute to the sustained activation of eNOS, associated with enhanced NO production
15. The- refore, it is expectable that this site may also play a role in the repressive effect in MMP-2, due to the relatively slow effects of fenofibrate (24 hr treatment in ECs).
The interplay of NO and MMPs in various contexts have been intensively studied. Our finding that NO negatively regulates MMPs is in line with the previous notion that enhanced NO bioavail- ability prohibits the pathophysiological activation of MMPs, and thus inhibits ECM degradation and vascular remodeling
8. Never-theless, there is also evidence implying a positive correlation of NO and MMP function
9. Actually, NO/reactive nitrogen species can mediate activation and expression of MMPs by modulating cystein residues
16. One
possible explanation for the seemingly paradox is that under physiological condition, NO is required for the normal function of MMPs and the NO- centered endothelial homeostasis keeps the MMP activation in check. The endothelial dysfunction occurring in metabolic disorder or CVDs is marked by an impaired NO bioavailability. The amelioration exerted by fenofibrate in enhancing NO could attenuate the detrimental effects downstream, such as overactivation of MMPs. However, under patho- physiological context, such as severe oxidative stress, the eNOS uncoupling may result in over- production of reactive nitrogen species, which becomes an undesirable events leading to MMP activation.
Previous study showed hypoxia significantly enhanced MMP-2 expression and activity in EC
17,18. Studies of circulating level of MMPs and their inhibitors have been conducted previously during Fig. 4.Fenofibrate treatment increases phospho-
rylation of eNOS Ser1177:
(a) Eahy926 cells were cultured in serum- free condition for 16 hours before fenofibrate treatment. Eahy926 cells were treated with fenofibrate (0, 10, 50μM) for 2 hours, and cell lysates were collected. 50μg cell lysate was used in each lane for western blot analysis. (b) Eahy 926 cells were cultured in serum-free condition for 16 hours. Cells were treated with 50μM fenofibrate for the time indicated in each lane. Cells were lysed and analyzed by Western blot.
Fig. 5.Fenofibrate mediated MMP-2 suppression is nitric oxide-dependent.
(a) Eahy 926 cells were pretreated with 500μM L-NAME in serum free medium over night(16hr). Eahy 926 cells were then treated with fenofibrate (10, 50μM), fenofibrate and L-NAME, or L-NAME for 24hr. Culture medium was collected and analyzed by zymograpghy.
(b) Cells were lysed and cell lysate was used
in western blotting.
hemodialysis. Serum MMP-2 and increased oxidative stress are associated with carotid ath- erosclerosis in hemodialyzed patients
19. Our study demonstrated that fenofibrate represses MMP-2 expression in EC and it is reasonable to propose that fenofibrate provides a protective role in EC under pathological conditions, eventhough the contrary effect was reported previously
20,21. Therefore, the physiological function of MMP-2 raised by fenofibrate between normal and patient could be further discussed.
In this focused study, we provide one clue for the vascular beneficial effects of fenofibrate, in terms of its modulation in MMPs in ECs. The hypothetical scheme was demonstrated (Fig. 6). It is very likely that such effect may also be observed in VSMC, the other key constituent of vasculature.
We would expect to observe inhibitory effects in SMC hypertrophy and proliferation, as well as overall preventive effects in atherogenesis, vascular remodeling and arterial stiffness.
Conclusion
In the present study, we demonstrated that fenofibrate, one of the most prominent fibrates, negatively regulates MMP-2 expression as well as activity in ECs. This regulation is largely, if not all, mediated by eNOS-derived NO. Taken together, these findings provide further understanding of beneficial effects provided by fenofibrate in ECs, which may contribute to clinical use of this medicine in reducing CVD risk in patients with metabolic symptoms.
References
1.Diabetes Atherosclerosis Intervention Study Investigators.
Effect of fenofibrate on progression of coronary-artery disease in type 2 diabetes: the Diabetes Atherosclerosis Intervention Study, a randomized study. Lancet 2001; 357:
905-10.
2. Davignon and Ganz. Role of endothelial dysfunction in atherosclerosis. Circulation 2004; 109: III27-32.
3. Lacolly P, Challande P, Osborne-Pellegrin M, Regnault V. Genetics and pathophysiology of arterial stiffness.
Cardiovasc Res 2009; 81: 637-48.
4. Yetik-Anacak G, Catravas JD. Nitric oxide and the en- dotheliu: history and impact on cardiovascular disease.
Vascul Pharmacol 2006; 45: 268-76.
5. Raffetto JD, Khalil RA. Matric Metalloproteinases and their inhibitors in vascular remodeling and vascular disease.
Biochem Pharmacol 2008; 75: 346-59.
6. Valentin F, Bueb JL, Kieffer P, Tschirhart E, Atkinson J. Oxidative stress activates MMP-2 in cultured human coronary smooth muscle cells. Fundam Clin Pharmacol 2005; 19: 661-7.
7. Davis GE, Pintar Allen KA, Salazar R, Maxwell SA. Matrix metalloproteinase-1 and -9 activation by plasmin regulates a novel endothelial cell-mediated mechanism of collagen gel contraction and capillary tube regression in three-dimensional collagen matrices. J Cell Sci 2001; 114: 917-30.
8. Gurjar MV, Sharma RV, Bhalla RC. eNOS gene transfer inhibits smooth muscle cell migration and MMP-2 and MMP-9 activity. Arterioscler Thromb Vasc Biol 1999; 19:
2871-7.
9. Genis L, Gonzalo P, Tutor AS, et al. Functional interplay between endothelial nitric oxide synthase and membrane type 1 matrix metalloproteinase in migrating endothelial cells.
Blood 2007; 110: 2916-23.
10.Nguyen M, Arkell J, Jackson C. Activated protein kinase C directly activates human endothelial gelatinase A. J Biol Chem 2000; 275: 9095-8.
11.Mount PF, Kemp BE, Power DA. Regulation of endothelial and myocardial NO synthesis by multi-site eNOS pho- sphorylation. J Mol Cell Cardiol 2006; 42: 271-9.
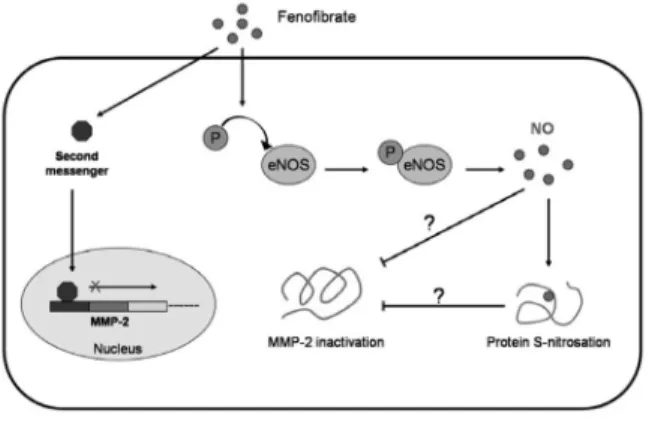
Fig. 6.The hypothetical scheme of fenofibrate su- ppressed MMP-2 function.
Fenofibrate might triggers second messen- ger and suppresses MMP-2 transcription.
Fenofibrate can also activates eNOS and
increases nitric oxide bioavailability. MMP-2
is then possibly be S-nitrosated directly and
loss its activity, or maybe, the other proteins
alter their function to inactivate MMP-2 via
S-nitrosation.
12.Murakami H, Murakami R, Kambe F, et al. Fenofibrate activates AMPK and increases eNOS phosphorylation in HUVEC. Biochem Biophys Res Commun 2006; 341: 973-8.
13.Fulton D, Gratton JP, McCabe TJ, et al. Regulation of endothelium-derived nitric oxide production by the protein kinase Akt. Nature 1999; 399: 597-601.
14.Dimmeler S, Fleming I, Fisslthaler B, et al. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999; 399: 601-5.
15.Chen Z, Peng IC, Sun W, et al. AMP-activated protein kinase functionally phosphorylates endothelial nitric oxide synthase Ser633. Circ Res 2009; 104: 496-505.
16.Okamoto T, Akuta T, Tamura F, van Der Vliet A, Akaike T.
Molecular mechanism for activation and regulation of matrix metalloproteinases during bacterial infections and respiratory inflammation. Biol Chem 2004; 385: 997-1006.
17.Ben-Yosef Y, Lahat N, Shapiro S, Bitterman H, Miller A.
Regulation of endothelial matrix metalloproteinase-2 by hypoxia/reoxygenation. Cir Res 2002; 90: 784-91.
18.Ben-Yosef Y, Miller A, Shapiro S, Lahat N. Hypoxia of endothelial cells leads to MMP-2-dependent survival and death. Am J Physiol Cell Physiol 2005; 289: 1321-31.
19.Krystyna P, Dariusz P, Michal M. Serum matrix meta- lloproteinase-2 and increased oxidative stress are associated with carotid atherosclerosis in hemodialyzed patients Atherosclerosis 2007; 190: 199-204.
20.Herouy Y, Mellios P, Bandemir E, Dichmann S, Nockowski P, Schopf E. Inflammation in stasis dermatitis upregulates MMP-1, MMP-2 and MMP-13 expression. J Dermatol Sci 2001; 25: 198-205.
21.Matsusaka H, Tsutsui H, Ikeuchi M, Matsushima S, Wen J, Kubota T. J Cardiac Failure 2004; 5(Suppl 1): 151.
Fenofibrate抑制內皮細胞基質金屬蛋白 -2 (Matrix Metalloproteinase-2)之表現
楊家慧1
趙尹慈
2林廷遠
3黃兆康
3沈希哲
4王寧
5陳識中
3,6臺北縣立醫院 1內科部新陳代謝科 2內科部風濕免疫科
3內科部心臟科 4急診醫學科
5中央研究院 6臺北醫學大學 內科學系
摘 要
Fenofibrate 已廣泛使用於心血管疾病病患之高酯血症。現今之研究發現,fenofibrate
經由活化過氧化體增生活化接受體α(Peroxisome Proliferator Activated Receptor type alpha, PPARα),改變酯質代謝而達到血管保護作用,而多數証據亦證實fenofibrate經由多效作用 (Pleiotropic effect)對血管造成保護作用。以往研究顯示,血管內皮細胞失調對於動脈硬化之
成因扮演極重要之角色,本實驗探討fenofibrate對於血管內皮細胞保護作用之機轉。血管內皮細胞經
fenofibrate處理後可抑制基質金屬蛋白 -2之表現、內皮細胞基質金屬蛋白 -2之分
泌與活性。Fenofibrate可經由磷酸化Ser1177造成活化,而經由L-NAME抑制內皮型一氧化氮
合成
(eNOS)可減緩fenofibrate造成抑制基質金屬蛋白 -2之表現。本實驗證實了fenofibrate
抑制基質金屬蛋白
-2需要一氧化氮(NO)之參與。總之,本研究發現fenofibrate可活化eNOS,
增加一氧化氮生物利用度,進而抑制基質金屬蛋白