行政院國家科學委員會專題研究計畫 成果報告
抗血清因子及標識蛋白研究
計畫類別: 個別型計畫
計畫編號: NSC94-2320-B-006-067-
執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日
執行單位: 國立成功大學生物化學暨分子生物學科(所)
計畫主持人: 張敏政
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 95 年 10 月 31 日
行政院國家科學委員會專題研究計劃成果報告 計畫名稱:抗血清因子及標識蛋白研究
計畫編號:NSC 94-2320-B-006-067
執行期限:94 年 8 月 1 日~95 年 7 月 31 日
主持人:張敏政 執行機構及單位名稱:成大醫學院生化所
一、中文摘要
我們由創傷弧菌臨床菌珠篩選出其外泌性酵素脂解酶及其脂解活化蛋白外泌性
溶血蛋白,我們也利用跳躍子 Tn5 變異篩選法由此臨床菌珠選殖出其抗血清因
子而與鉀離子輸送有關之蛋白,TrkA 及 σ54 依存性雙份子系統之反應蛋白 polR,進一步的實驗證明 TrkA 不僅參考鉀離子進出菌體本身的調控,並在此菌 感染宿主時,亦擔任抵抗宿主防禦系統的角色。此外我們利用相似重組法分別構
築 polR 剔除菌及 σ54 剔除菌。此兩剔除菌對血清、抗菌小胜肽、吞噬菌及嗜中
性的抵抗能力比原菌珠低,且兩剔除菌皮下注射感染實驗小白鼠的菌數致死量也
比原菌珠的菌數量高很多,此結果顯示抗血清因子 σ54 及其調控蛋白 polR 是創
傷弧菌的重要致病因子。
關鍵詞:創傷弧菌,抗血清,雙份子系統,致病因子
Abstract
The genes encoding extracellular lipase, lipase activator, and extracellular hemolysin III, were cloned from V. vulnificus CKM-1, respectively. We also cloned the gene encoding a K+ uptake protein, TrkA, and the gene encoding aσ54-dependent response regulator (PolR) of two-component systems, from the Tn5 mutants of V.
vulnificus. Further experiments demonstrated that TrkA may modulate the transport of
potassium and resistance to host innate defenses. The polR isogenic and theσ54 isogenic mutant were individually constructed by using homologous recombination and both the two isogenic mutants showed more sensitive than parental strain to killing by serum, polymyxin B, macrophages, and polymorphonuclear leukocytoes (PMN). Mice inoculated with polR orσ54mutant demonstrated virulence attenuation when the strains were administered subcutaneously, suggesting theσ54 and its regulating protein PolR are the important virulent factors.
Keywords: Vibrio vulnificus, serum resistance, two-component system, virulent
factor
二、計畫緣由與目的
Vibrio vulnificus is a halophilic gram-negative bacterium notorious for its ability
to cause both serious wound infections and fatal septicemia in humans, particularly in those with certain underlying diseases especially in patients with chronic liver disease (1, 2). Primary septicemia may be acquired upon consumption of seafood containing this organism, and wound infections are often associated with exposure of wounds to seawater. Cases of V. vulnificus infections have been reported from many areas of the world (3-6). In the United Stated, this organism is responsible for 95% of the seafood-related deaths, most often affecting men over the age of 40 (7). The cases of primary bacteremia are often complicated by severe soft tissue infections such as necrotizing fasciitis and myonecrosis coupled with hemorrhagic bullae formation on the skin (8). The mortality rate of patients with primary septicemia in our series and those reported by others exceeds 50%, and death occurs mostly within 48 h of hospitalization (9, 10). Because V. vulnificus often contaminate in the seafood and/or seawater, the prevalence rate of liver cirrhosis is very high in Taiwan, acquaculture and the consumption of seafood especially raw seafood is very popular, V. vulnificus infection is apparently more prevalent in this island than in many areas of the world.
Several virulence factors or properties of V. vulnificus strains have been described.
Wright et al demonstrated that injection of iron into mice could lower the LD
50of a virulent strain of V. vulnificus (11). A catechol siderophore synthesis mutant of V.
vulnificus incapable of acquiring iron from transferrin had reduced virulence in an
animal model (12). The presence of a polysaccharide capsule has also been associated with virulence for mouse (13-16). Other purported determinants of virulence of V.
vulnificus include extracellular cytolytic and cytotoxic factors (17-20), a
phospholipase (21) and a metalloprotease (22, 23), although two exotoxins, a metalloprotease and a cytolysin were not confirmed as virulence factors by genetic analysis (24, 25). Recently, several regulators, including cyclic AMP receptor protein (CRP)/LuxR, ToxRS/CRP, Fur, RpoS, SmcR, and LuxS (26), have been reported to control expression of virulence factors.
Rapid adaptation to hostile environments faced during the course of infection
is essential for the survival of many bacterial pathogens. This adaptive response is
often effectively mediated by two-component regulatory systems that typically consist
of a membrane-associated sensor kinase and a DNA-binding response regulator. The
sensor kinase responses to specific environmental stimuli by phosphorylating its
cognate response-regulator protein, thereby the activated phosphoprotein activates or represses the target genes involved in responding to the individual environmental signal in order to facilitate survival of the cell. It is also generally thought that Gram-negative bacterial pathogens evade the action of cationic, amphipathic, antimicrobial polypeptide (CAMP) found within neutrophil granules, macrophage phagosomes, and secretions of mucosal epithelia (27) by modifying the lipid A part of their lipopolysaccharides (LPS). CAMP contains notable anti-microbial factors including defensins, azurocidin, melittin, magainins, serum complement and polymyxin B (28). Salmonellae resistance to polymyxin appears to be regulated by PhoP, a response-regulator protein which controls gene expression in response to environmental signals. The two-component regulatory system, phoPQ operon in
Salmonella typhimurium is required for virulence in mice and survival withinmacrophage (29). Although there are a wide number of two-component regulatory systems and some of which are involved in control virulence gene expression within bacterial pathogens have been described, no information is available about this system in V. vulnificus. However, since infection caused by V. vulnificus is remarkable for its invasiveness and rapidly fulminating course of disease, it is reasonable to predict that
V. vulnificus is likely to posses the ability of rapid adaptation to hostile environments,and to have developed method effectively to evade host defenses. This report described the cloning of the genes encoding extracellular lipase, lipase activator, extracellular hemolysin III, a K+ uptake protein, TrkA, and aσ54-dependent response regulator (PolR) of two-component systems from the V. vulnificus as well as the characterization of them by genetic analysis
三、結果與討論
(1).Cloning and characterization of the lipase and lipase activator protein from Vibrio vulnificus CKM-1
The gene (lipA) encoding the extracellular lipase and its downstream gene (lipB)
from V. vulnificus CKM-1 were cloned. Nucleotide sequence analysis suggest that
LipA is a member of bacterial lipase family I.1 and that LipB is a lipase activator of
LipA (Fig.1). The active LipA was produced in recombinant E. coli cells only in the
presence of the lipB. The precise functions of the LipA in V. vulnificus are unclear.
Further studies with genetic modifications for functional analysis may help to elucidate whether LipA is a virulence determinant in the pathogenesis of V. vulnificus
(2). Characterization and Virulence of Hemolysin III from V.vulnificus
A gene (hlyIII) encoding a hemolysin was cloned and HlyIII showed 48% sequence identity to the hemolysin III of Bacillus cereus. When hlyIII was expressed in E. coli, crude extracts exhibited hemolytic activity. A hlyIII isogenic mutant showed an attenuated virulence (the LD50 value was 16-fold increase) compared with the wild-type when administered intraperitoneally in mice. The results indicated that Hly III is a virulence factor in mice, although it has not yet been investigated whether Hly III is an exotoxin during infection.
(3). A k+ uptake protein, TrkA, is required for serum, protamine, and polymyxin B resistance in V. vulnificus
To identify the genes required for resistance to human serum, we constructed a library of Tn5 mutants of V. vulnificus and screened them for hypersensitivity to serum. One of the isolated serum-susceptible mutants had a mutation in an ORF identified as trkA, whose gene product showing high identity to that of TrkA of Vibrio alginolyticus, a protein required for the uptake of potassium. A trkA isogenic mutant was significantly more easily killed by human serum, protamine, or polymyxin B than was the wild type. Infection experiments demonstrated virulence attenuation when this mutant was administered intraperitoneally or subcutaneously to both normal and iron-treated mice (Table 1), indicating that TrkA may modulate the transport of potassium and resistance to host innate defenses and that it is important for virulence in mice.
(4). V. vulnificus responses to host during infection--- virulence factors that are required for serum, antimicrobial peptides and neutrophil killing resistance in V. vulnificus
A DNA fragment that is involved in serum susceptivity in another Tn5 mutant was
cloned. This fragment was found to potentially encode two genes, polS and polR,
which are arranged in a putative polSR operon and encode products with homology to
the sensor and σ54 response regulator proteins of two-component systems. Both
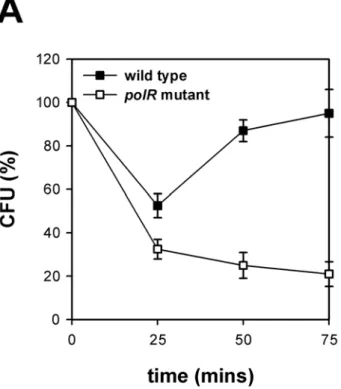
polR and σ54 isogenic mutant showed more sensitive than parental strain to killingby serum, polymyxin B, macrophages (data not shown), and polymorphonuclear
leukocytoes (PMN) (Fig.2).
Mice inoculated with polR or σ 54 isogenic mutant demonstrated virulence attenuation when the strains were administered subcutaneously (data not shown).
Promoter fusion experiments indicated that the polSR was regulated by cell growth phase and specifically induced during exposure of bacteria to serum, macrophages, or PMN (data not shown), suggesting that polSR was up-regulated during PMN or macrophages phagocytosis and that this transcriptional regulatory system controls gene that assist V. vulnificus immune evasion.
5). Vibrio vulnificus in Taiwan
We analyze a clinical-case series of 84 patients with V. vulnificus infection from 1995 to 2000 and describe the molecular epidemiologic features of pathogens isolated from these patients. The spectrum of clinical manifestations and outcomes, options of antimicrobial therapy, and virulence mechanisms were investigated. Results of molecular typing of isolates from humans and marine environment in this country had a high genetic divergence among these isolates. Education and measures are needed to prevent this emerging disease.
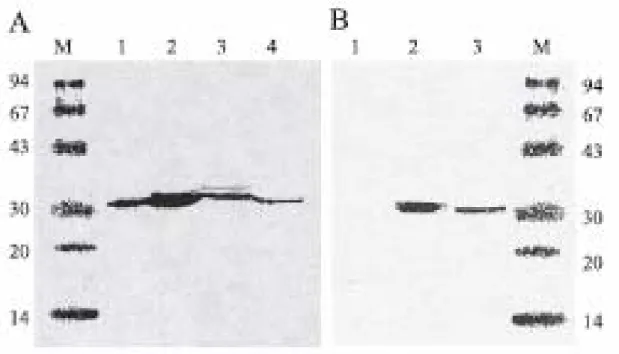
Fig. 1. (A) Western blot analysis of LipA. Lanes 1 and 2, crude cell extractsfrom E.
coli XL1B carrying pUA and pUAB, respectively; lanes 3 and 4, crude cell extracts
and oncentrated supernatants of culture broth from V. vulnificus CKM-1, respectively.
(B) Western blot analysis of LipB. Lanes 1 and 2, crude cell extracts from E. coli
XL1B carrying pUA and pUAB, respectively; lane 3, crude cell extracts from V.
vulnificus CKM-1. The culture medium was supplemented with IPTG. Samples were
separated by SDS-PAGE, blotted and incubated with antisera. Lane M, molecular mass standards (Pharmacia); values in kilodaltons.
Fig.2. Bacterial killing kinetics of wild type and polR mutant by neutrophils. The numbers of viable bacteria (CFU) after
incubation with neutrophils are expressed as percentage of the original counts. ..
TABLE 1. Virulence of V. vulnificus strains in untreated or irontreated mice
a ND, not done.
AKK-1: a trkA isogenic mutant TRK-1: AKK-1 containing trkA gene
四、計畫結果與自評
由於創傷弧菌可引起嚴重且蔓延迅速之傷口感染及敗血症,且近來台灣地區此菌 所引起的感染日愈增多而頗受矚目,我們在本計畫之執行中已先後選殖出其抗血 清致病因子,及其外泌蛋白因子之基因,同定並探討其與致病機制之關聯性。另 外由於又發現此菌含一新型之螢光蛋白,在本計畫中也詳細研究此蛋白,並以定 向演化及隨機突變改良其螢光蛋白強度,也獲得了預期成果,經本計畫之研究成 果可期待其作用在分生領域有應用潛力之報導蛋白。
五、參考文獻
1. Morris, J.G., Jr., and R.E. Black. 1985. Cholera and other vibrioses in the United States. N. Engl. J. Med. 312:343-350.
2. Tison, D.L., and M.T. Kelly. 1984. Vibrio species of medical importance. Diagn.
Microbiol. Infect. Dis. 2:263-276.
3. Blake, P.A., M.H. Merson, R.E.Weaver, D.G. Hollis, and P.C. Heublein. 1979.
Disease caused by a marine Vibrio: clinical characteristics and epidemiology. N.
Engl. J. Med. 300:1-5.
4. Bonner, J.R., A.S. Coker, C.R. Berryman, and H.M. Pollock. 1983. Spectrum of vibrio infections in a Gulf Coast community. Ann. Intern. Med. 99:464-469.
5. Kelly, M.T., and W.F. McCormick. 1981. Acute bacterial myositis caused by Vibrio
vulnificus. JAMA 246:72-73.6. Park, S.D., H.S. Shon, and N.J. Joh. 1991. Vibrio vulnificus septicemia in Korea:
clinical and epidemiologic findings in seventy patients. J. Am. Acad. Dermatol.
24:397-403.
7. Oliver, J.D. 1989. Vibrio vulnificus. p. 569-600. In M.P. Doyle (ed.), Foodborne
bacterial pathogens. Marcel Dekker, New York.
8. Chuang, Y.C., C. Young, and C.W. Chen. 1989. Vibrio vulnificus infection. Scand. J.
Infect. Dis. 21:721-726.
9. Chuang, Y.C., C.Y. Yuan, C.Y. Liu, C.K. Lan, and A.H.M. Huang. 1992. Vibrio
vulnificus infection in Taiwan: report of 28 cases and review of clinicalmanifestations and treatment. Clin. Infect. Dis. 15:271-276.
10.Klontz, K.C., S. Lieb, M. Schreiber, H.T. Janowski, L.M. Baldy, and R.A. Gunn.
1988. Syndromes of Vibrio vulnificus infections: clinical and epidemiologic features in Florida cases, 1981-1987. Ann Intern. Med. 109:318-323.
11.Brennaman, B., D. Soucy, and R.J. Howard. 1987. Effect of iron and liver injury on the
pathogenesis of Vibrio vulnificus. J. Surg. Res. 43:527-531.
12.Litwin, C.M., T.W. Rayback, J. Skimer. 1996. Role of catechol siderophore synthesis in Vibrio
vulnificus virulence. Infect. Immun. 64:2834-2838.
13.Simpson, L.M., V.K. White, S.F. Zane, and J.D. Oliver. 1987. Correlation between virulence and colony morphology in Vibrio vulnificus. Infect. Immun. 55:269-272.
14.Wright, A.C., L.M. Simpson, J.D. Oliver, and J.G. Morris, Jr. 1990. Phenotypic evaluation of
acapsular transposon mutants of Vibrio vulnificus. Infect. Immun. 58:1769-1773.
15.Zuppardo, A.B., and R.J. Siebeling. 1998. An epimerase gene essential for capsule synthesis in Vibrio vulnificus. Infect. Immun. 66:2601-2606.
16.Gray, L.D., and A.S. Kreger. 1985. Purification and characterization of an extracellular cytolysin produced by Vibrio vulnificus. Infect. Immun. 48:62-72.
17.Gray, L.D., and A.S. Kreger. 1987. Mouse skin damage caused by cytolysin from
Vibrio vulnificus and by V. vulnificus infection. J. Infect. Dis. 155:236-241.18.Gray, L.D., and A.S. Kreger. 1989. Detection of Vibrio vulnificus cytolysin in V.
vulnificus-infected mice. Toxicon. 27:459-464.
19.Park, J.W., S.N. Ma, E.S. Song, C.H. Song, M.R. Chae, B.H. Park, H.W. Rho, S.D.
Park, and H.R. Kim. 1996. Pulmonary damage by Vibrio vulnificus cytolysin.
Infect. Immun. 64:2873-2876.
20.Park, J.W., S.N. Ma, E.S. Song, C.H. Song, M.R. Chae, B.H. Park, H.W. Rho, S.D.
Park, and H.R. Kim. 1996. Pulmonary damage by Vibrio vulnificus cytolysin.
Infect. Immun. 64:2873-2876.
21.Testa, J., W. Daniet, and A.S. Kreger. 1984. Extracellular pxhospholipase A2 and lysophospholipase produced by Vibrio vulnificus. Infect. Immun. 45:458-463.
22.Kothary, M.H., and A.S. Kreger. 1985. Production of partial characterization of an elastolytic protease of Vibrio vulnificus. Infect. Immun. 50:534-540.
23.Hirofumi, W., S.I. Miyoshi, T. Kawase, K.I. Tomochika, and S. Shinoda. 2004.
High growing ability of Vibrio vulnificus biotype 1 is essential for production of a
toxic metalloprotease causing systemic disease in humans. Microbiol Pathogen.
36:117-123.
24.Shao, C.P., and L.I. Ho. 2000. Metalloprotease is not essential for Vibrio vulnificus virulence in mice. Infect. Immun. 68:3569-3573.
25.Fan, J.J., C.P. Shao, Y.C. Ho, C.K. Yu, and L.I. Hor. 2001. Isolation and characterization of a Vibrio vulnificus mutant deficient in both extracellular metalloprotease and cytolysin. Infect. Immun. 69:5943-5948.
26.Shao, C.P. and Hor, L. 2001. Regulation of metalloproteasegene expression in
Vibrio vulnificus by a Vibrio harveyi LuxR homologue. J. Bacteriol.183:1369-1375.
27.Boman, H.G. 1995 Peptide antibiotics and their role in innate immunity. Annu Rev
Immunol 13:61-92.28.Vaata, M. 1992 Agents that increase the permeability of the outer membrane.
Microb Rev 56:395-411.