A Solution Study on the Local and Global Structure Changes of Cytochrome c: An
Unfolding Process Induced by Urea
†I-Jui Hsu,‡ Ying-Jen Shiu,§U-Ser Jeng,*,£Tung-Ho Chen,£Yu-Shan Huang,£ Ying-Huang Lai,£ Ling-Na Tsai,# Ling-Yun Jang,£Jyh-Fu Lee,£Li-Jiaun Lin,#Sheng-Hsien Lin,§and Yu Wang‡ Department of Chemistry, National Taiwan UniVersity, Taipei 106, Taiwan, Institute of Atomic and Molecular Sciences, Academia Sinica, P. O. Box 23-166, Taipei 106, Taiwan, National Synchrotron Radiation Research Center, Hsinchu, Taiwan 300, and Industrial Technology Research Institute, Hsinchu, Taiwan 310
ReceiVed: April 19, 2007
The local and global structural changes of cytochrome c induced by urea in aqueous solution have been studied using X-ray absorption spectroscopy (XAS) and small-angle X-ray scattering (SAXS). According to the XAS result, both the native (folded) protein and the unfolded protein exhibit the same preedge features taken at Fe K-edge, indicating that the Fe(III) in the heme group of the protein maintains a six-coordinated local structure in both the folded and unfolded states. Furthermore, the discernible differences in the X-ray absorption near-edge structure (XANES) of these two states are attributed to a possible spin transition of the Fe(III) from a low-spin state to a high-spin state during the unfolding process. The perseverance of six-coordination and the spin transition of the iron are reconciled by a proposed ligand exchange, with urea and water molecules replacing the methionine-80 and histidine-18 axial ligands, respectively. The SAXS result reveals a significant morphology change of cytochrome c from a globular shape of a radius of gyration Rg) 12.8 Å of the native protein to an elongated ellipsoid shape of Rg) 29.7 Å for the unfolded protein in the presence of concentrated urea. The extended X-ray absorption fine structure (EXAFS) data unveil the coordination geometries of Fe(III) in both the folded and unfolded state of cytochrome c. An initial spin transition of Fe(III) followed by an axial ligand exchange, accompanied by the change in the global envelope, is proposed for what happened in the protein unfolding process of cytochrome c.
1. Introduction
Since the folding concept of describing the polypeptide chains of a protein in its native structure was introduced, protein folding/unfolding has long been a subject of interest.1 Such interest is often revived by advances in technologies on structural probes such as absorption, fluorescence, and circular dichroism (CD), which are often used to monitor the local structure changes of proteins during an unfolding process through denaturants or by elevating the temperature in aqueous solu-tion.2,3 Structural information deduced from these optical spectroscopes is, however, somewhat less direct in comparison with that obtained from X-ray diffraction, scattering, and/or absorption, which give a direct probe of the electron density distribution or the molecular structure of a protein.
In a recent decade or so, the great advances in the synchrotron radiation source together with the measurement techniques and data analysis algorithms have made small-angle X-ray scattering (SAXS) a practical and increasingly popular probe for nano-structures. Although with a rather low spatial resolution for electron density in comparison with that of X-ray diffraction, SAXS is, however, very sensitive to the mean electron density
fluctuations on the subnanometer to sub-submicron scale either in the solid or in solution system and can provide adequate information for the study of morphology and interactions of biological macromolecules in solution, including equilibrium or dynamic aspects of protein folding/unfolding.3-6Boffi et al.7 and Soldatov et al.8 demonstrated that X-ray absorption spectroscopy (XAS) could be a direct probe for revealing the local coordination changes near the iron core of the heme group in cytochrome c or myoglobin when subjected to the changes in pH value of the solution. A combined study of SAXS and XAS for correlated global and local structural changes is expected to provide insights into the understanding of protein folding/unfolding and/or binding behaviors.3
In this work, XAS and SAXS are applied to study the structural changes of a model protein cytochrome c induced by a denaturant, urea. The oxidized cytochrome c from a horse heart is a member of the ubiquitous class of heme proteins; the protein contains 104 residues of amino acids, and a heme group consists of a six-coordinated Fe(III). The heme group is surrounded by three major helices and two cysteine side chains (Cys 14 and Cys 17), which provide the thioether bridges to the heme group. In a native (folded) state, the Fe of the heme is bonded to four nitrogen atoms of the porphyrin (Por) plane in a pseudo C4symmetry, with two axial ligands from histidine-18 (His-histidine-18) and methionine-80 (Met-80), as illustrated in Figure 1.9The changes in the coordination geometry of iron and the changes in the overall morphology are investigated using XAS and SAXS when the unfolding of cytochrome c takes place by introducing urea into the aqueous solution.
†Part of the “Sheng Hsien Lin Festschrift”.
* To whom correspondence should be addressed. E-mail: usjeng@ nsrrc.org.tw. Address: National Synchrotron Radiation Research Center, 101 Hsin-Ann Road, Hsinchu Science Park, Hsinchu 30076, Taiwan. Tel:
+886-3-578-0281, ext: 7108. Fax: +886-3-5783813.
‡National Taiwan University. §Academia Sinica.
£National Synchrotron Radiation Research Center. #Industrial Technology Research Institute.
10.1021/jp073031q CCC: $37.00 © 2007 American Chemical Society Published on Web 08/14/2007
2. Experimental Section
Cytochrome c (cyt c) from a horse heart in an oxidized form was purchased from the Sigma Chemical Company and used without further purification. Sample solutions containing 21 mg/mL (1.7 mM) of cytochrome c, in a buffer solution (pH∼ 7) of potassium phosphate (100 mM) and 0.1 M NaCl, were prepared with urea in the concentrations of 0, 5, 6, 6.5, 7, 8, and 10 M for both SAXS and XAS measurements at 25°C. For the sake of background subtraction, blank buffer solutions with urea in various concentrations were also prepared.
X-ray absorption spectra for the cytochrome c solutions were measured in a fluorescence mode at the wiggler beamline BL-17C1 of the National Synchrotron Radiation Research Center (NSRRC), Taiwan, with a double crystal monochromator of Si(111) of an energy resolution, ∆E/E, of 2× 10-4. The energy was scanned from 6.912 to 8.010 KeV using a Lytle detector filled with Ar gas or a 13 element solid-state detector for the near-edge region. A standard Fe foil was used for energy calibration. EXAFS data analysis was completed using the UWXAFS package,10with theoretical scattering paths generated by the FEFF7 program11based on model structures.9,15
SAXS data were collected using the small-angle X-ray scattering instrument at the beamline 17B3 of NSRRC, with a 0.5 mm diameter beam of 10 KeV. Details of the instrument description were reported previously.12Data measured were then corrected for electronic noise of the area detector, background scattering, incoming flux, sample thickness, and pixel sensitivity of the detector to yield the absolute scattering intensity (scat-tering cross section per unit sample volume).12Each spectrum of SAXS or XAS presented here was compiled by more than 10 data scans for better statistics. These data scans were quite reproducible, indicating that the radiation damage on the protein was negligible during the XAS or SAXS measurements.
3. Results and Discussion
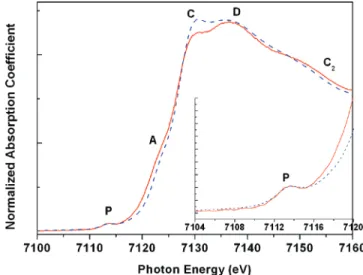
3.1. XAS for Local Structure Changes. Fe K-edge XANES spectra of cytochrome c solutions, both in a native state and in the unfolded state when the 10 M urea is added in the solution, are shown in Figure 2. The spectra are characterized in several regions of P (∼7113 eV), A (∼7122 eV), C (∼7130 eV), D (∼7140 eV), and C2(∼7153 eV), according to a previous XAS study8on a temperature-dependent spin transition of myoglobin. The preedge peak in the P region is often interpreted as the 1s
f 3d(t2g, eg) electron quadruple transition, which originates from a distortion of a perfect octahedral symmetry; the distortion increases the d-p mixing of Fe to make the quadruple transition possible. Therefore, the preedge peak is expected to be sensitive to local geometry changes in the coordination of Fe. Regions
A and C2are related to the motion of the iron relative to the porphyrin plane of the heme group, whereas C and D are indicative of the spin state of Fe as well as the coordination geometry.
As emphasized in the inset of Figure 2, both XANES spectra of the native state and the unfolded state give the same preedge transition in the P region, indicating that the iron of heme group in this protein maintains the same six-coordinated geometry in both states. Moreover, the significant enhancement in the C region for the protein in the unfolded state versus the folded state is consistent with a theoretical prediction8based on a spin transition model. In the model, Fe undergoes a spin transition from a low-spin state (LS, S ) 1/2) to a high-spin state (HS, S
) 5/2), accompanied by a lengthening of the Fe-N/O bond
distance which is responsible for such enhanced absorption in the C region; by lengthening the Fe-N distance, the ligand 2p orbitals participate less in the hybridization of the Fe(4s,4p) orbitals, leading to the enhancement of the 1s f 4p transition (C region).13
Furthermore, the spin transition and the accompanying geometrical change around Fe can also be interpreted by the coupled changes in the A, D, and C2regions shown in Figure 2, as indicated earlier by Soldatov et al.8and Della Longa et al.7Similar changes in the XANES spectra were also observed for spin-crossover Fe(II) complexes during spin transition.13
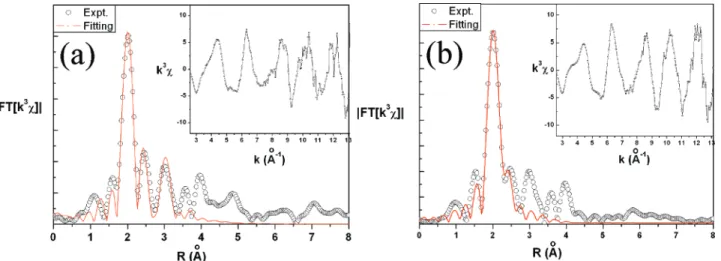
To extract more local structure information, we have per-formed EXAFS data analysis for the XAS spectra of the native and unfolded states (with 10 M urea), as shown in Figure 3a and b where the weighted EXAFS data (k3χ) are illustrated in the insets. The normalized EXAFS spectra are obtained from the absorption data based on the relation χ(k) ) [µ(k) -µ0(k)]/∆µ0(0), whereµ(k) is the measured absorption coefficient, µ0(k) the background, and∆µ0(0) the edge jump. The wave-number k ) [2m(E - E0)/p]1/2is defined by the photon energy E, the threshold energy E0, the plank constant p, and the mass of the electron m. The coordination geometry around Fe obtained from the Fourier transform of k3χ together with the fitted curve (solid line) for cytochrome c at the native state are displayed in Figure 3a; details of the fitting procedure used were given in a previous report.14The fitting result includes (1) the first coordination shell of the iron with bond distances, R, of 2.00 and 2.10 Å, which consists of the four N(Por) and one N(His-18) atoms, respectively; (2) the second shell with a char-acteristic bond distance of 2.41 Å from the S atom of Met-80;
Figure 1. Local structure of the heme group in a native cyto-chrome c.
Figure 2. XANES spectra of the cytochrome c both at the native (folded) state (solid line) and in the unfolded state (dashed line) (10 M urea). The inset is the enlarged preedge absorption region.
and (3) the third shell with a distance of 3.04 Å from the eight carbon atoms of the porphyrin. The fitted parameters are summarized in Table 1. These bond distances are consistent with those from a previous NMR result9for an average structure of cytochrome c in the native state, Fe-N(Por) ) 1.980-2.111 Å, Fe-N(His-18) ) 1.948 Å, and Fe-S(Met-80) ) 2.343 Å. Several possible models were tried for fitting the EXAFS data of the unfolded state; the information of a large global morphology change due to the unfolding of the protein deduced from the SAXS result (see below) does help to eliminate a few models. The final conclusion is a ligand exchange model, with urea and water molecules replacing the two axial ligands of the Met-80 and His-18, respectively. A similar ligand exchange model was also previously proposed by Yeh et al.16 in a resonance Raman scattering study on the unfolding of cyto-chrome c by a denaturing agent of guanidine chloride. The fitted curve using the ligand exchange model of the unfolding state is shown in Figure 3b. The fitted coordination geometry around Fe in the unfolding state is with four N(Por) at a bond distance of 2.00 Å, which is the same as that in the native state, indicating a relatively stable structure of the porphyrin plane of the heme group after unfolding of the protein. However, the two axial ligands of His-18 and Met-80 in the folded state are now replaced by the oxygen of water and urea, respectively, with a distance of 2.27 Å; an accompanying characteristic bond distance of R ) 2.60 Å corresponds to the distance from the Fe atom to the C atom of the urea carbonyl group. Of course, the current data cannot completely rule out the possibility of having His-18 remaining in one of the two axial positions in the unfolded state of the protein since there is only one electron difference in O and N. In this case, only one urea molecule replaces the Met-80 ligand, but with a somewhat lengthened Fe-N(His) bond length.
Overall, detailed coordination geometries of Fe in folded and unfolded states obtained from the EXAFS data analysis (Table
1) clearly indicate that during the folding/unfolding process, the local geometry of iron in the heme group changes dramati-cally along the axial position of the Por plane, namely, the breaking of the Met-80 ligation, and possibly the His-18, by urea and water molecules, whereas the XANES data reveal a small distortion in the Por plane accompanied by a spin transition of Fe(III).
3.2 SAXS for Global Structure Changes. Small-angle scattering profiles for colloidal particles of a monodisperse size can be modeled as
where P˜(Q) is the normalized form factor with P˜(0) ) 1, and S(Q) is the structure factor with S(Q)∼ 1 in a dilute colloidal solution.17 The zero angle scattering intensity I
0 ) CN(b -FsVdry)2depends on the concentration C, the aggregation number N, and the dry volume Vdry for the scattering particles. The scattering length for the particles and scattering length density for the solvent are denoted by b and Fs, respectively. Without protein aggregation, the expected I0for a cytochrome c solution of 21 mg/mL (1.7 mM), in the absolute scattering intensity unit, is 0.17 cm-1. With urea added in the aqueous solution, the result is Fs) (f1F1+ f2F2), where f1F1and f2F2are, respectively, the products of the volume fraction and scattering length density of water (F1) 9.43 × 10-6Å-2) and urea (F2) 12.0 × 10-6 Å-2). An empirical formula by Modig et al.18can be used to calculate the volume fractions of water and urea in the sample solutions prepared. Since the scattering length density of cytochrome c (F∼ 12.0 × 10-6 Å-2) is very close to that of urea, the increase of urea concentration in a cytochrome c solution reduces the scattering contrast of the protein and, thus, the I0 value (or the scattering intensity), as illustrated in the SAXS data shown below.
For homogeneous ellipsoids with major axis a and semi-minor axis b, the scattering form factor averaged for spatial orientation is given by
where V ) Q[A2µ2+ B2(1 -µ2)]1/2and j
1is the spherical Bessel function of the first order.17For macroion solutions, the structure factor S(Q) depends on the volume fractionη and the fractional ionization of the ionic aggregates.17 Nevertheless, in buffer solution with salt, the charge interaction between cytochrome
Figure 3. The Fourier transforms (open circle) of the k3χ data (inset) and the fitted curves (solid line) of (a) the native cytochrome c and (b) cytochrome c in 10 M urea.
TABLE 1: EXAFS Fitting Parameters for the Folded and Unfolded States of Cytochrome ca
folded state unfolded state
bond R (Å) σ2(Å2) bond R (Å) σ2(Å2) Fe-N(Por) 2.00(4) 0.0003(7) Fe-N(Por) 2.01(1) 0.0001(6) Fe-N(His) 2.10(4) 0.0003(7) Fe-O(water) 2.25(2) 0.004(3) Fe-S(Met) 2.41(2) 0.002(2) Fe-O(urea) 2.27(2) 0.004(3) Fe-C(urea) 2.60(9) 0.004(3)
aVariable: R, interatomic distance;σ2, mean squared deviation of R obtained from the Debye-Waller factor.
I(Q) ) I0P˜ (Q)S(Q) (1) P˜(Q) )
∫
0 1|
3j1(V) V|
2 dµ (2)c can be largely screened, and the structure factor S(Q) derived from the Percus-Yevick (PY) model of hard sphere interactions, based on the volume fractionη ) CV/N (∼1.6% volume fraction in our case) and an effective sphere diameterσ approximated using Vdry) 4/3πab2) 4/3π(σ/2)3, may be used to approximate the interactions between cytochrome c in a buffer solution.19 One merit of using the S(Q) from the PY model in a SAXS data fitting process lies in the fact that there is no additional parameter needed in the fitting process, other than that required by the form factor P(Q).
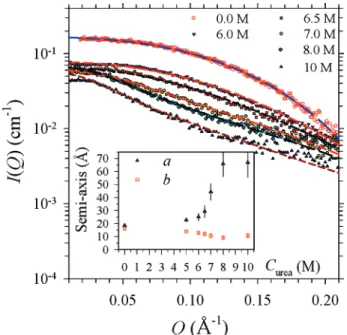
Selectively shown in Figure 4 are the SAXS data measured for the cytochrome c solutions with 0, 6.0, 6.5, 8.0, and 10 M urea. All of the data can be fitted systematically at the absolute intensity scale using the ellipsoid model for the form factor P(Q) and the Percus-Yevick model for the structure factor S(Q). The fitted semi-axis lengths a and b shown in the inset indicate that the native cytochrome c, of an aspect ratio (defined by a/b) of nearly unity, is essentially globular. The globular shape is stable up to a urea concentration of ∼5 M. Then, the envelope elongates significantly to an ellipsoid-like shape of an aspect ratio (a/b) of ∼6, at 10 M urea. From the fitted semi-axes lengths, we can deduce a radius of gyration Rg) 12.8 Å for the native protein and an Rgvalue of 29.7 Å for the unfolded protein with 10 M urea.17
4. Discussion
On the basis of the XAS and SAXS results, it is clear that cytochrome c undergoes a significant local and global structural transition during the unfolding process introduced by urea. Articulating the local and global structural changes observed, a possible unfolding route for the protein in urea solution is proposed: the approach of the urea molecule toward the heme group may induce a spin transition of the iron in the heme group from a LS to a HS state, thus weakening and breaking the bonds of Fe-S(Met-80) and/or Fe-N(His-18), leading to a ligand exchange of urea and/or H2O. Although we cannot completely rule out the possibility with water binding due to the weak effect of the second shell, the EXAFS fitting with urea, including the
C atom of urea to distinguish from water binding at the axial position, is better. A similar solvent effect on a spin transition of Fe(II) complexes was also observed in a previous report.20 In view of the molecular structure of cytochrome c, the heme can be a node for holding the globular structure of the protein, and the breaking of the axial ligands (observed by XAS) may serve as a key factor for unfolding the protein from a globular envelope to an elongated ellipsoid-like morphology (observed by SAXS). A systematic study, with combined XAS and SAXS, on the cytochrome c in various concentrations of urea is definitely useful for understanding the details of the unfolding process of the protein, for instance, the sequential unfolding process proposed recently by Maity et al.21
Furthermore, since the spin transition of iron from the LS to HS state could lead to a deformation of the porphyrin ring, it is possible to examine the spin transition due to the unfolding of cytochrome c by the changes of resonance Raman spectra in the region of ∼1000-1700 cm-1, a region that carries the characteristic C-N/C-C vibrations of the porphyrin ring. The two vibration bands at 1584 and 1566 cm-1are considered to be especially sensitive to the spin transition of Fe during the unfolding process of protein.22-23
5. Conclusions
The systematic study, based on combined XAS and SAXS, on the folding/unfolding of cytochrome c in the presence of urea leads to a ligand exchange model, with urea and/or water to replace the Met-80 and/or His-18 ligands of iron in the heme group. The XANES and EXAFS of native and unfolded cytochrome c measured at the Fe K-edge give direct evidence of spin transition in the Fe of the heme group together with significant changes in the ligand geometry, including the replacement of the Met-80 and/or His-18 ligand, for the unfolding process. The SAXS result reveals a drastic change in the protein morphology from a globular to an ellipsoidal shape during the unfolding process. A plausible mechanism for the unfolding of cytochrome c is that the spin transition of Fe takes place first in the presence of urea, followed by a ligand exchange from Met-80 and/or His-18 to urea and/or water molecules. Of course, the accompanied morphology change occurs when the Met-80 leaves the coordination of Fe. It is proven that an integrated study of XAS and SAXS can provide the necessary information of the unfolding process of a protein.
Acknowledgment. We thank Drs. H. S. Sheu and J. J. Lee for the discussion, Mr. C.-C. Tseng at the NSRRC for preparing part of the sample solutions, and Mr. C.-W. Pao for assisting with the XAS measurement. The work was partially supported by the NSC under Grant No. NSC-95-2112-M-213-002.
References and Notes
(1) Wu, H. Chin. J. Physiol. 1991, 5, 321-327.
(2) Ibarra-Molero, B.; Sanchez-Ruiz, J. M. Biochemistry 1997, 36, 9616-9624.
(3) (a) Doniach, S. Chem. ReV. 2001, 101, 1763-1778. (b) Cinelli, S.; Spinozzi, F.; Itri, R.; Finet, S.; Garsughi, F.; Onori, G.; Mariani, P.
Biophys. J. 2001, 81, 3522-3533.
(4) Koch, M. H. J.; Vachette, P.; Svergun, D. I. Q. ReV. Biophys. 2003,
36, 147-227.
(5) Uzawa, T.; Akiyama, S.; Kimura, T.; Takahashi, S.; Ishimori, K.; Morishima, I.; Fujisawa, T. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 1171-1176.
(6) Lipfert, J.; Doniach, S. Annu. ReV. Biophys. Biomol. Struct. 2007,
36, 307-327.
(7) Boffi, F.; Bonincontro, A.; Cinelli, S.; Castellano, A. C.; De Francesco, A.; Della Longa, S.; Girasole, M.; Onori, G. Biophys. J. 2001,
80, 1473-1479.
Figure 4. The SAXS data for the cytochrome c solutions, with several urea concentrations from 0 to 10 M, are fitted (dashed curves) using the ellipsoid form factor together with the PY structure factor. The inset shows the corresponding values for the semi-axes a and b extracted from the model fitting.
(8) (a) Soldatov, A. V.; Yalovega, G. E.; Smolentsev, G. Yu.; Kovtun, A. P.; Della Longa, S. Biophysics 2001, 46, 564-568. (b) Della Longa, S.; Pin, S.; Corte`s, R.; Soldatov, A. V.; Alpert, B. Biophys. J. 1998, 75, 3154-3162.
(9) Banci, L.; Bertini, I.; Gray, H. B.; Luchinat, C.; Reddig, T.; Rosato, A.; Turano, P. Biochemistry 1997, 36, 9867-9871.
(10) (a) Frenkle, A. I.; Stern, E. A.; Voronel, A.; Qian, M.; Newville, M. Phys. ReV. B 1994, 49, 11662-11665. (b) Newville, M.; Ravel, B.; Haskel, D.; Rehr, J. J.; Stern, E. A. Physica B 1995, 208, 154-159.
(11) Zabinsky, S. I.; Rehr, J. J.; Ankudinov, A.; Albers, R. C.; Eller, M. J. Phys. ReV. B 1995, 52, 2995-2999.
(12) Lai, Y. H.; Sun, Y. S.; Jeng, U.; Lin, J. M.; Lin, T.-L.; Sheu, H.-S.; Chuang, W.-T.; Huang, Y.-H.-S.; Hsu, C.-H.; Lee, M.-T.; Lee, H.-Y.; Liang, K. S.; Gabriel, A.; Koch, M. H. J. J. Appl. Crystallogr. 2006, 39, 871-877. (13) (a) Briois, V.; dit Moulin, Ch. C.; Sainctavit, Ph.; Brouder, Ch.; Flank, A.-M. J. Am. Chem. Soc. 1995, 117, 1019-1026. (b) Real, J. A.; Castro, I.; Bousseksou, A.; Verdaguer, M.; Burriel, R.; Miguel Castro, M.; Linares, J.; Varret, F. Inorg. Chem. 1997, 36, 455-464. (c) Lee, J. J.; Sheu, H. S.; Lee, C. R.; Chen, J. M.; Lee, J. F.; Wang, C. C.; Huang, C. H.; Wang, Y. J. Am. Chem. Soc. 2000, 122, 5742-5747. (d) Okamoto, K.; Nagai, K.; Miyawaki, J.; Kondoh, H.; Ohta, T. Chem. Phys. Lett. 2003,
371, 707-712. (e) Oyanagia, H.; Tayagakib, T.; Tanaka, K. J. Phys. Chem. Solids 2004, 65, 1485-1489. (f) Khalil, M.; Marcus, M. A.; Smeigh, A.
L.; McCusker, J. K.; Chong, H. H. W.; Schoenlein, R. W. J. Phys. Chem.
A 2006, 110, 38-44.
(14) Hsu, I. J.; Liu, R. S.; Chen, J. M.; Liu, R. G.; Jang, L. Y.; Lee, J. F.; Harris, K. D. M. Chem. Mater. 2000, 12, 1115-1121.
(15) The codes JAJJOU, MANJIU, and MITQEV in the Cambridge database are used to assist the building of the urea replacement model.
(16) (a) Yeh, S. R.; Takahashi, S.; Fan, B.; Rousseau, D. L. Nat. Struct.
Biol. 1997, 4, 51-56. (b) Takahashi, S.; Yeh, S. R.; Das, T. K.; Chan, C.
K.; Gottfried, D. S.; Rousseau, D. L. Nat. Struct. Biol. 1997, 4, 44-50. (c) Yeh, S. R.; Rousseau, D. L. J. Biol. Chem. 1999, 274, 17853-17859. (d) Yeh, S. R.; Han, S.; Rousseau, D. L. Acc. Chem. Res. 1998, 31, 727-736.
(17) Chen, S. H.; Lin, T. L. Methods of Experimental Physics. Neutron
Scattering in Condensed Matter Research; Sko¨d, K., Price, D. L., Eds.;
Academic Press: New York, 1987; Vol. 23B, Chapter 16.
(18) Modig, K.; Kurian, E.; Prendergast, F. G.; Halle, B. Protein Sci.
2003, 12, 2768-2771.
(19) Hayter, J.; Penfold, J. Mol. Phys. 1981, 42, 109-114.
(20) (a) Halder, G. J.; Kepert, C. J.; Moubaraki, B.; Murray, K. S.; Cashion, J. D. Science 2002, 298, 1762-1765. (b) Niel, V.; Thompson, A. L.; Carmen Mun˜oz, M.; Galet, A.; Goeta, A. E.; Real, J. A. Angew. Chem.,
Int. Ed. 2003, 42, 3763-3766. (c) Glaser, T. Angew. Chem., Int. Ed. 2003, 42, 5668-5670.
(21) Maity, H.; Maity, M.; Englander, S. W. J. Mol. Biol. 2004, 343, 223-233.
(22) (a) Spiro, T. G.; Strekas, T. C. J. Am. Chem. Soc. 1974, 96, 338-345. (b) Spiro, T. G.; Strekas, T. C. Proc. Natl. Acad. Sci. U.S.A. 1972, 69, 2622-2626. (c) Jordan, T.; Eads, K. C.; Spiro, T. G. Protein Sci. 1995, 4, 716-728.
(23) Yamamoto, T.; Palmer, G.; Gill, D.; Salmeen, T.; Rimai, L. J. Biol.