行政院國家科學委員會專題研究計畫 成果報告
腦內啡 mu 型受體作用藥改善胰島素抗阻性效果的研究(3/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2320-B-006-010- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立成功大學藥理學科(所) 計畫主持人: 鄭瑞棠 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 9 月 2 日
Abstract
Aerobic exercise including treadmill running has long been used to successfully treat and/or prevent insulin resistance and Type-2 diabetes. Increase of plasma β-endorphin is observed with exercise. The present study was designed to clarify the role of endogenous β-endorphin in exercise-induced improvement of insulin resistance. We used a moderate exercise program consistingof treadmill running at 20 m.min-1 and 0 % grade for 1 h.day-1, 7 days.week-1, for 8 weeks. An increase of plasma
β-endorphin-like immunoreactivity (BER) in parallel with the reduction of plasma glucose was obtained in exercise-trained obese Zucker rats. Different to a marked reduction in sedentary obese rats, the value of insulin-stimulated glucose disposal rate (GDR), measured by the hyperinsulinemic euglycemic clamp technique obtained from the exercised obese rats was reversed to near that of the sedentary lean group eight weeks later of the last period of exercise. This effect of exercise was inhibited by naloxone or naloxonazine at doses sufficient to block opioid µ-receptors. Signaling-related defects in the soleus muscle of sedentary obese Zucker rats, which impaired glucose transporter subtype 4 (GLUT-4), included decreased phosphorylation of insulin receptor substrate (IRS)-1, as well as an attenuated p85 regulatory subunit of phosphatidylinositol 3-kinase (PI3 kinase) and Akt serine phosphorylation.. In contrast, exercise training failed to modify the levels of insulin receptor (IR), IRS-1, and IR tyrosine autophosphorylation in obese Zucker rats. Enhanced insulin sensitivity via exercise training might be mediated by endogenous β-endorphin through an increase of post-receptor insulin signaling related to the IRS-1-associated PI3-kinasestep that leads to the enhancement of GLUT 4 translocation and improved glucose disposal in obese Zucker rats.
Keywords: Akt; β-endorphin; glucose transporter subtype 4; insulin receptor; insulin receptor substrate-1; phosphatidylinositol 3-kinase
Introduction
Insulin resistance is a key feature of impaired glucose tolerance in type-2 diabetes [1] that can be characterized by a diminished ability of insulin sensitive tissues and a marked decrease of glucose metabolism in response to insulin. In target tissues of insulin, skeletal muscle that is the primary site of insulin-mediated glucose disposal shows a significant decrease of glucose uptake, as seen in insulin resistance [2]. Defects in the insulin signal cascade leading to impaired glucose utilization have been proposed to play a key role in the pathogenesis of insulin resistance [3].
A regular aerobic exerciseprogram is accepted as a positive factor in health and it is viewed as particularly useful in the management ofdiabetes, obesity and cardiovascular diseases [4]. It is established that both an acute bout of exercise and chronic endurance exercise trainingmayincrease muscle oxidative capacity to limit muscle insulin resistancein obese Zucker (fa/fa) rats and in insulin-resistant humans with impaired glucose tolerance or Type-2 diabetes [4]. However, the mechanisms responsible for the acquisition of tolerance afterexercise remain unclear.
It has been documented that exogenous β-endorphin induces an acute increase of circulating insulin in both non-diabetic people and in those with diabetes mellitus (DM) [5]. Recently, we reported that
β-endorphin may increase glucose utilization via opioid µ-receptors activation, which acts to lower plasma glucose in streptozotocin-induced diabetic rats [6]. Also, the induction of insulin resistance occurs more quickly in opioid µ-receptor knockout mice than in wild-type mice [7]. These findings implicate opioid
µ-receptor activation in the improvement of insulin action.
Exercise stimulates an increase of endogenous opioid [8]. It seems reasonable to suppose that
β-endorphin is involved in the improvement of insulin resistance by exercise training. The present study was carried out to clarify this contention.
Materials and Methods Animal models
(Animal Research Center, Tokyo Medical College, Tokyo, Japan) and were bred in the animal center of National Cheng Kung University Medical College. All animal procedures were performed according to the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health, as well as the guidelines of the Animal Welfare Act. All rats were fed with standard chow (Purina Mills, LLC, St Louis, MO). They were maintained in a temperature-controlled room (25 ± 1 ℃) under a 12:12 light-dark cycle (light on at 06:00 h). Food and water were available ad libitum.
Exercise program
The obese Zucker rats were randomly assigned to two groups: a group that remained sedentary andan exercise-trainedgroup. Animals in the exercise-trainedgroups were trained for every morning for 60 minutes for an eight-week period. Exercise consisted of running on a motorized treadmill. Rats were initially screened by determining whether they could run on a motor-driven treadmillat 15 m.min-1, 0 % grade, for 60 min. A minimum of one week later, those obese Zucker rats that completed the exercise screentest were included in the experiments described herein.
Forexercise experiments, the obese Zucker rats received one session of exercise, whichconsisted of running on a treadmill at 20 m.min-1, 0 % gradefor 60 min [9]. Then, the antagonists naloxone and naloxonazine (both from Research Biochemical Inc., Natick, MA) were used to identify the role of opioid
µ-receptors. The rats received repeated intraperitoneal (i.p.) injections of naloxone (1.0 mg.kg-1) or naloxonazine (1.0 mg.kg-1) three times daily (t.i.d.) during the exercise training regimen. Control rats were similarly injected with an equal volume of 0.9 % NaCl in distilled water. Lean and other obese animals in the sedentary groups were removed from their cages once a day each day and were placed on the stationary treadmill for 60 min toaccount for handling stress. Training began when the ratswere eight weeks old and finished at 16 weeks of age for investigation at the same age. After the hyperinsulinemic euglycemic clamp procedure, rats were sacrificedand the soleus muscle was immediately removed and washed withcold phosphate buffer. The tissueswere cut into 200–300 mg portions and stored separately at -70 °C for later immunoprecipitation and immunoblotting analyses.
Laboratory determinations
Blood samples were collected from the tail vein of rats for measurement of plasma glucose concentration and plasma level of insulin, or for β-endorphin-like immunoreactivity (BER). The concentration of plasma glucose was measured by the glucose oxidase method using an analyzer (Quik-Lab, Ames, Miles Inc., Elkhart, IA). The enzyme-linked immunosorbent assay (ELISA) was employed to quantify the plasma level of BER or insulin using a commercially available kit (Penisula Lab. Inc., Belmont, CA).
Hyperinsulinemic euglycemic clamp
Hyperinsulinemic euglycemic clamp was performed eight weeks after thelast exercise bout. Food was withheld beginning 12 hours before the procedure. Rats were then chronically cannulated under interperitoneal (i.p.) anesthesia with sodium pentobarbital (30.0 mg.kg-1) in the femoral vein for infusion of glucose, tracer, and insulin, and in the femoral artery for sampling. Animals were weighed and placed in a restrainer to which they were accustomed before the clamp. After the basal sample at -60 min, experiments were begun with a priming injection (2.5 µCi 0.5 ml-1) and constant infusion (0.167 µCi min-1) of D-[3-3H] glucose (New England Nuclear, Boston. MA). Tracer glucose was diluted in saline containing 5.5 mmol l-1 unlabeled D-glucose (Sigma-Aldrich, Inc., Saint Louis, MO) as carrier and 200 mg.dl-1 sodium benzoate (Sigma-Aldrich, Inc.) as preservative. Fifty minutes after the start of tracer infusion, blood samples were collected in heparinized microtubes at -10 min for determination of plasma glucose concentration and glucose specific activity. At 0 min, rats were randomly assigned to receive infusion of regular human insulin (Novo Industrias, Bagsvaerd, Denmark) at 40 mU kg-1 min-1. The infusate of insulin was diluted with saline containing 0.5 % human serum albumin (Baxter, Glendale, CA). Blood samples (10 µl) were drawn at 10-min intervals for the immediate determination of plasma glucose. Based on these values, 20 % dextrose (Abbott Laboratories, Chicago, IL) was variably infused to maintain the plasma glucose concentration at about 5.5 mmol.l-1. The steady state was generally achieved within 70-90 min, within which time a blood sample was collected for determination of glucose specific activity measured in duplicate after 5% zinc sulfate and 0.3 N barium hydroxide deproteinization. After termination blood
sampling at 120 min, animals were killed with a lethal dose of sodium pentobarbital (100.0 mg kg-1, i.p.). The glucose disposal rate (GDR) at steady state was calculated using Steele’s equation [10].
Muscle processing
Soleus muscle samples used for measuring insulin signaling were weighed while still frozen and were homogenized in ice-cold lysisbuffer (1:10 wt vol-1) containing 50 mmol.l-1 HEPES (pH 7.6), 150 mmol.l-1 NaCl,20 mmol.l-1 sodium pyrophosphate, 20 mmol.l-1 β-glycerophosphate, 10 mmol.l-1 sodiumfluoride, 2 mmol.l-1 sodium orthovanadate (Na3VO4), 2 mmol.l-1 EDTA (pH 8.0),1 % Nonidet P-40, 10 % glycerol, 1
mmol.l-1 phenylmethylsulfonyl fluoride,1 mmol.l-1 MgCl2, 1 mmol.l-1 CaCl2, 10 µg.ml-1 leupeptin, and 10
µg.ml-1 aprotinin.A Polytron homogenizer seton maximum speed for 30 s was used for homogenization. Homogenateswere incubated on ice for 20 min and then centrifuged at 15,000Xg for 20 min at 4 °C.
Muscle debris was removed, and protein concentrationsof crude extracts were estimated by BioRad protein dye binding assay (Bio-Rad Laboratories, Richmond, CA). Supernatantwas stored at 80 °C until used.
Immunoprecipitation and immunoblotting
Protein concentration in the tissue homogenates was determined by the Bio-Rad protein assay following the manufacturer's instructions(Bio-Rad Laboratories). A 1-mg sample of total protein was used for immunoprecipitation with anti-insulin receptor β-subunit antibody or anti-IRS-1 antibody at 4 °C overnight, followed by the addition of Protein A-Sepharose beads (Sigma-Aldrich, Inc.) for 1 h. The protein bead-antibodycomplexes were precipitated by briefcentrifugation. The pellets were washed three times in ice-cold buffer (0.5 % Triton X-100, 100 mmol.l-1 Tris, pH 7.4, 10 mmol.l-1 EDTA and 2 mmol.l-1 sodium vanadate), resuspended in Laemmli sample buffer, and boiled for 5 min before sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) in a 10 % acrylamide gel using the Bio-Rad Mini-Protein II system (55V and 130V during the stacking and separation gels, respectively). Protein was transferred to a polyvinylidene difluoride membrane using a Bio-Rad Trans-Blot system (2 h at 20 V in 25
mmol.l-1 Tris, 192 mmol.l-1 glycine and 20 % MeOH). Following transfer, the membrane was probed with anti-insulin receptor β-subunit antibody (NeoMarkers, Fremont, CA), anti-IRS-1 antibody (NeoMarkers) or anti-phosphotyrosine antibody (Santa Cruz Biotechnology, Inc.
,
Santa Cruz, CA) according to manufacturer’s instructions.For the detection of p85 regulatory subunit of PI3-kinase, Akt serine (Ser473) phosphorylation, and GLUT 4 content, equal amounts (50 µg) of proteinwere prepared from muscle homogenates and subjected to SDS-PAGE, transferred to polyvinylidene difluoride membrane as describedabove and blotted with anti-PI3-kinase p85 subunit antibody (NeoMarkers), anti-phosphoserine (Ser473) Akt antibody (Cell Signaling Technology, Inc., Beverly, MA) or anti-GLUT 4 antibody (Genzyme diagnostics, Cambridge, MA) according to manufacturer’s instructions. The intensity of the blots incubated with mouse monoclonal actin antibody (Santa Cruz Biotechnology, Inc.) were used as controls to ensure that the amount of protein loaded into each lane of the gel was constant.
After three 5-min washes in TBST (20 mmol.l-1 Tris-HCl (pH 7.5), 150mmol.l-1 NaCl, and 0.05 % Tween 20), membranes were then incubated with the appropriate peroxidase-conjugated secondary antibodies. The membranes were washed three times in TBST and visualizedon X-ray film using the enhanced chemiluminescence detectionsystem (Amersham Corp., Braunschweig, Germany) according to the protocol of manufacturer. Densities of the obtained immunoblots were quantified using a laser densitometer.
Statistical analyses
Data are expressed as the mean ± SEM for the number (n) of animals in the group as indicated in Tables and Figures. Statistical differences among groups were determined by using two-way repeated-measures ANOVA. The Dunnett range post-hoc comparisons were used to determine the source of significant differences where appropriate. A p-value < 0.05 was considered statistically significant.
Results
Animal characteristics
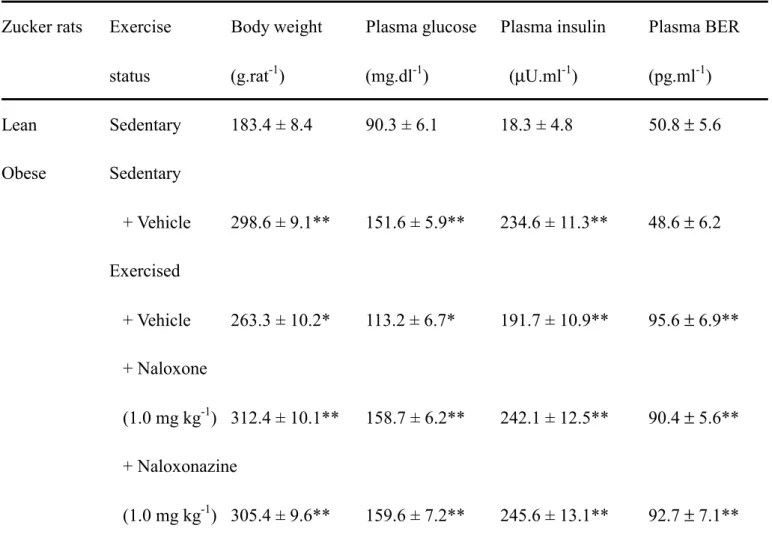
As shown in Table 1, untrained obeseZucker rats were significantly heavier than the lean littermates. Exercise trainingdid not decrease body weight markedly in these rats. But the body weights of exercised obese Zuckerrats were significantly increased in the presence of either naloxone or naloxonazine. Similar to the change of plasma insulin concentration, the fasting plasmaglucose concentrations were markedly raised in obese Zuckerrats as compared with the lean controls. However, the plasma BER of the obese rats was not statistically different with that of lean group. Eight weeks after thelast exercise bout, fasting plasma glucose concentration was significantly lower inobese Zucker rats than the sedentary counterparts (p<0.05), but the value was still markedly higher than that in the untrained lean Zucker rats (p<0.05). The plasma insulin concentration in fasting obese rats was reduced after the eight week exercise session although the level was still higher than that in fasting and sedentary lean littermates. An elevation of the plasma BER level was also observed in the same group of exercise-trained obese Zuckerrats, showing about 2-fold relative to the sedentary lean group (Table 1).
Increase of the plasma BER level was observed in exercised obese Zuckerrats who were pretreated with naloxone (Table 1). However, in the presence of naloxone plasma glucose concentration and plasma insulin level in fasting, exercise-trained obese Zuckerrats approached the values of untrained obese Zucker rats (Table 1). Moreover, the plasma glucose and insulin of the exercised obese rats were reversed to higher levels in the naloxonazine-treated group. Similar to the effect of naloxone, an elevation of the plasma BER in exercised obese Zuckerrats was also observed upon pretreatment with naloxonazine (Table 1).
Effect of exercise training on the glucose disposal rate of obese Zucker rats
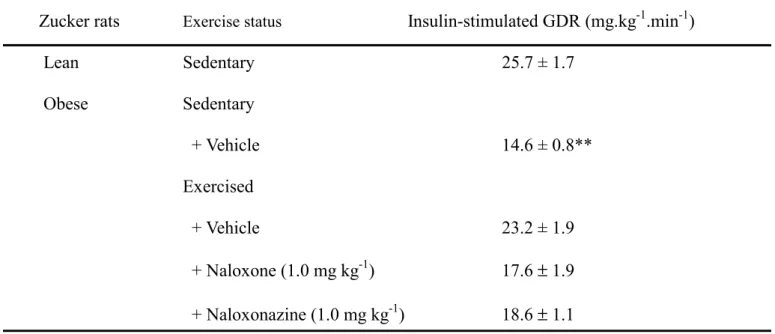
Plasma insulin was increased to 1,000–10,000 fold of the basal level during hyperinsulinemic euglycemic clamp. At the steady state of the hyperinsulinemic euglycemic clamp, a significant blunting in the
insulin-stimulated GDR (14.6 ± 0.8 mg.kg-1.min-1) was obtained from sedentary obese Zucker rats as compared to their sedentary lean littermates (25.7 ± 1.7 mg.kg-1. min-1) (Table 2). The value of insulin-stimulated GDR in the exercised obese rats became 23.2 ± 1.9 mg. kg-1.min-1, which was no different to that of sedentary lean rats (Table 2). In the presence of naloxone insulin-stimulated GDR in exercised obese Zucker rats was reversed to 15.1 ± 0.9 mg.kg-1.min-1, a value similar to that in untrained obese rats (Table 2). Similarly, naloxonazine treatment of obese Zucker rats reversed the insulin-stimulated GDR (15.4 ± 0.8 ng.kg-1.min-1) to near the value observed in the untrained animals (Table 2).
Effect of exercise training on the protein levels of insulin receptor in soleus muscle of Zucker rats
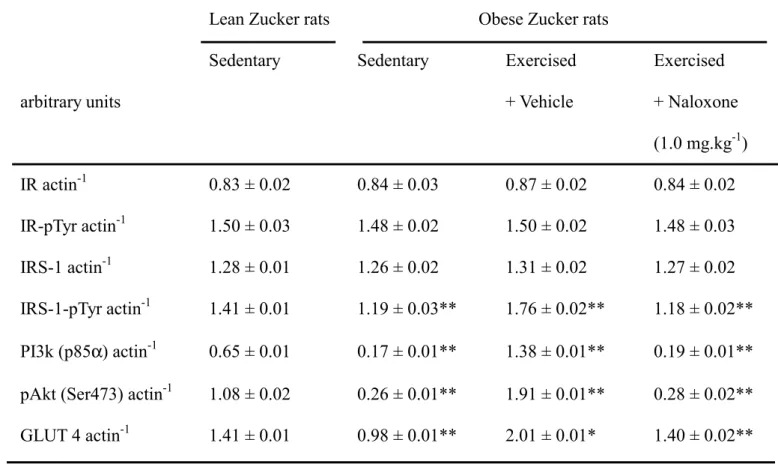
Expression of insulin receptor protein in soleus muscle was similar in obese Zucker rats and the lean littermates (Figure 1). Endurance exercise training did not modify these values. Similar results were observed in the tyrosine phosphorylation of the insulin receptor (Figure 1). The basal level oftyrosine phosphorylation of the insulin receptor was not significantly different from group to group, confirming prior observations [11]. Also, naloxone did not alter protein levels or the degree of the tyrosine phosphorylation of the insulin receptor in the soleus muscle of the exercised obese Zuckerrats (Figure 1, Table 3).
Effect of exercise training on the levels of IRS-1 in soleus muscle of Zucker rats
The expression of IRS-1 protein in soleus muscle of untrained obese Zucker rats was similar to that from the sedentary lean group (Figure 1). Endurance exercise training did not modify these values. In addition, naloxone did not affect the level of IRS-1 protein in exercise-trained obese rats (Figure 1). Consistent with previous observations [11], the basal value of IRS-1 tyrosine phosphorylation was not significantly different in any group. However, the degree of IRS-1 tyrosine phosphorylation in the soleus muscle of sedentary obese Zucker rats was about 80 % of that in sedentary lean animals (Figure 1). The degree of IRS-1 tyrosine phosphorylation in the soleus muscle of the obese rats was markedly elevated in the
exercise-trained group, which achieved a level near that of the sedentary lean group (Figure 1). However, exercise training failed to make similar changes of IRS-1 tyrosine phosphorylation in exercised obese rats pretreated with naloxone (Figure 1, Table 3).
Effect of exercise training on the level of p85 regulatory subunit of PI3-kinase in soleus muscle of Zucker rats
The expression of p85 regulatory subunit of PI3-kinase in the soleus muscle of sedentary obese Zucker rats was decreased to about 26 % of that in their sedentary lean littermates (Figure 1). But, the p85 regulatory subunit of PI3-kinase expression in soleus muscle of the exercise-trained obese rats was elevated approximately 2-fold as compared to their sedentary lean counterparts (Figure 1). A similar antagonism by naloxone on the enhancement of the level of p85 regulatory subunit of PI3-kinase in soleus muscle of exercise-trained obese Zucker rats was also obtained (Figure 1, Table 3).
Effect of exercise training on the degree of Akt serine phosphorylation in soleus muscle of Zucker rats
Similar to a previous study [11], the basal values ofAkt phosphorylation were not significantly different in any group. A marked reduction of Akt serine (Ser473) phosphorylation was observed in the soleus muscle of sedentary obese Zucker rats, showing only about 24 % of that from sedentary lean rats (Figure 1). However, the degree of insulin-stimulated Akt serine phosphorylation was significantly elevated in the obese rats after exercise training, being almost 1.8-fold higher than that of sedentary lean controls (Figure 1). Pretreatment with naloxone abolished this effect (Figure 1, Table 3).
Effect of exercise training on the protein levels of GLUT 4 in soleus muscle of Zucker rats
Before the exercise training, a marked (near 70%) decrease of GLUT 4 protein expression was observed in the soleus muscle of obese Zucker rats, compared to lean controls (Figure 1). After exercise training, the GLUT 4 protein expression was reversed to the level higher than that from the sedentary lean group
(Figure 1). The protein level of GLUT 4 was reduced upon the blockade of opioid µ-receptors by naloxone (Figure 1, Table 3).
Discussion
Obese Zucker rat exhibit a range of metabolic aberrations including hyperlipidemia, hyperglycemia, adipocytehypertrophy, and hyperinsulinemia. These symptoms are similar to thoseobserved in human Type-2 diabetes, making these rats useful as an animalmodel for the study of this malady [12]. Insulin resistance in this animal is responsive to exercise training[13]. Thus, these rats served as a suitable model system for the present study.
Due to the concerns of injury and high mortality rate [14], we employed a conservative regimen (20 m.min-1, 0 % grade) to elucidate the effects of moderate exercise training on insulin signaling in obese Zucker rats. Because chronic exercise training appears to induce a physiological adaptationthat augments insulin sensitivity, the exercise training was protracted (eight weeks).
Our resultsdemonstrate that this exercise regimen is effective in the reduction of plasma glucose accompanied by a relative improvement of plasmainsulin level. Insulin-mediated glucose utilization identified by therate of glucose disposal using hyperinsulinemic euglycemic clamp (the most widely used technique to characterize insulin sensitivity; [15]) is raised after endurance exercise training. Indeed, the ability of insulin to stimulate glucose disposal assessed by this method was shown to be markedly impaired in the obese Zucker rat, indicative of the presence of insulin resistance. Thus, our results demonstrate that the exercise training regimen improves insulin sensitivity.
This improvement was observed to be abolished by naloxone or naloxonazine at doses known to be sufficient to block opioid µ-receptors [16]. The endogenous β-endorphin has multiple physiological actions that are mediated by opioid µ-receptors [17]. In fact, an increase of plasma BER in parallel with the reduction of plasma glucose was presently obtained in exercise-trained obese Zucker rats. The amelioration of insulin-stimulated GDR by exercise mirrored the increase of plasma BER secretion, with both characteristics exhibiting about a 2-fold difference from the levels in sedentary rats. Therefore, it
appears that the recovery of insulin sensitivity in obese Zucker rats by aprogram of regular exercise might be associated with endogenous β-endorphin. This is consistent with prior observations that insulin resistance is more quickly developed in the absence of opioid µ-receptors [18]. Thus, the beneficial role of opioid µ-receptors activation can be considered in the improvement of insulin resistance.
Although β-endorphin is released with adrenocorticotrophic hormone from the pituitary gland [18], the pituitary gland-independent release of endogenous opioids occurs in other organs [19] such as the adrenal gland [19]. Various forms of polypeptides that cross-react with enkephalin antisera are stored together with catecholamine and soluble proteins in the granules of adrenal gland. We have previously demonstrated that activation of α1-adrenoceptors in the adrenal medulla that enhance β-endorphin
secretion is associated with a lowering of the plasma glucose concentration in diabetic rats [20]. Therefore, secretion of endogenous β-endorphin from the adrenal gland might be primarily responsible for the improvement of insulin sensitivity by moderate exercise training in obese Zucker rats.
In insulin resistance, certain intracellular signaling pathways are more resistant to insulin than others [3]. Skeletal muscle is recognized as the major tissue of insulin-stimulated glucose disposal, with impaired
in vivo glucose disposal being manifest in this local [2]. Insulin action is also greater in the red type I
fibers of the soleus muscle [21]. Exercise is known to increase insulinresponsiveness of skeletal muscle. Thus, the examination of soleus muscle samples from all animals who received the eight weeks endurance exercise training is an entirely appropriate means to investigate the insulinsignaling.
The final mediator of reduced glucose clearance in obese Zucker rats is the decrease of glucose transporter subtype-4 (GLUT 4) function in skeletal muscle cells [22]. Recent evidence suggests that this decrease is related to abnormalities in the insulin signal pathway including insulin receptor substrate (IRS)-1 and phosphatidylinositol 3-kinase (PI3-kinase) [23]. Similar to the protein expression of insulin receptor, insulin-stimulated tyrosine phosphorylation of theinsulin receptor β-subunit and the expression of IRS-1 were preserved in the obese Zucker rats, discordant with a reduction in IRS-1 tyrosine phosphorylation, the subsequent step in insulin signal cascade. Usually, IRS-1 tyrosine phosphorylation in response to insulin stimulation increases IRS-1 association with p85, the regulatory subunit of PI3-kinase,
which is the main molecule linking insulin signal to glucose transport [24]. We found that protein expression of p85 was markedly decreased in the soleus muscle of sedentary obese Zucker rats. Moreover, the Akt serine phosphorylation (Ser473) in the soleus muscle was also reduced as compared with that in sedentary lean controls. Although the signal intermediaries downstream from PI3-kinase remained to be delineated, current data indicates that serine/threonine kinase protein B (PKB) (also known as Akt) is a potential link between insulin-stimulated PI3-kinase and IRS-1 association [25]. Impairment of Akt serine phosphorylation in cells from animals exhibiting insulin resistance has been documented [23]. Indeed, Akt is mentioned as one of the potential mediators in insulin signals for glucose transportation, in which glucose uptake and GLUT 4 action could be stimulated by phosphorylated Akt in the target tissues [25]. Presently, we observed a decrease of GLUT 4 protein in the membrane fraction of soleus muscle from sedentary obese Zucker rats. It seems that proteins involved in the early stepsof insulin signaling were less impaired in this muscle. These results are consistentwith a previous study showing that insulin receptor signaling throughthe PI3-kinase pathway linking to glucose transport is reduced in this rodent model of insulinresistance [26]. We found that endurance exercise training reversed the IRS-1 phosphorylation in soleus muscle of obese Zucker rats. However, the protein expression of the insulin receptor or its autophosphorylation and the content of IRS-1 were not influenced by this exercisetraining. Furthermore, the expression of p85 and the insulin response for Akt serine phosphorylation as well as GLUT 4 translocation in sedentary obese Zucker rats were restored after eight weeks endurance exercise training. It is possible that the insulin sensitivity of obese Zuckerrats was improved by endogenous β-endorphin during endurance exercise training to increase the expressionof some genes involved in insulin signal pathway, specifically the IRS-1-associated PI3-kinasestep that leads the translocation of GLUT 4 to enhance glucose uptake and improved insulin action in exercised obese rats. Hence, the insulin-stimulated GDR would be significantly improved. However, insulin action was markedly improved while the plasma insulin level was reduced only slightly in exercised obese Zuckerrats. This basis of this contradiction is still unclear. Owing to the genetic changes and hyperlipidemia in obese mutant Zucker rats, it may be impossible to obtain the regular response of plasma glucose level reduction together with the improvement
of insulin action.
In genetically predisposed individuals, type 2 diabetes is pathogenically linked with progressive obesity [27]. The exercise training in the present study did not modify the body weight markedly in obese Zuckerrats. Indeed, the body weights of the exercised obese rats with a higher level of plasma BER were significantly increased in the presence of opioid µ-receptor blockers. It seems that increase of body weight may be one of the mechanisms for blockade of the opioid µ-receptor. Alternatively, a direct effect of opioid µ-receptor inhibitors could be operative. We could not discriminate between this two possibilities, since a control group in which rats received the inhibitors was not included.. Effects of exercise on GDR, body weight, plasma glucose and plasma insulin were blocked by opioid µ-receptor antagonist. Also, the exercised-induced improvement in IRS-1 tyrosine phosphorylation, p85/PI3K levels, and Akt phosphorylation were also reversed in the presence of opioid µ-receptor antagonist, indicating that the enhanced insulin sensitivity by exercise training is mediated by endogenous β-endorphin via opioid
µ-receptor activation.
A beneficial effect of this activation on the morphine-induced supraspinal antinociception linking the PI3-kinase and phospholipase C (PLC) pathways has been documented in the mouse [28]. The mechanism of the opioid tolerance involves a number of signals including the PLC-protein kinase C (PKC) pathway [29]. Opioid µ-receptors activation lowering plasma glucose concentration via the PLC-PKC pathway occurs in insulin deficient animals [30]. PLC and PKC participate in the insulin signals that link to the glucose transporter system [31]. In fact, diacylglycerol-sensitive PKCs do not appear to be required for insulin effects on glucose transport, whereas the atypical PKCs are essential for insulin stimulation of glucose transport in adipocytes and muscle tissue [32]. Although atypical PKCs are located downstream from IRS-1 and PI3 kinase in the established insulin signal pathways, IRS-1 is also a novel substrate for atypical PKCs [33]. Recently, it has also been documented that PKC-delta may be involved in the adaptive effects of exercise in skeletal muscles of type 2 DM [34]. Therefore, more studies to define the role of PKC isoforms in the signaling of β-endorphin linked to insulin signal pathways are required in the future.
β-endorphin to influence the postreceptorinsulin signaling, specifically at the IRS-1-associated PI3-kinase step in the cascade that leads to GLUT 4 translocation and glucose disposal in obese Zucker rats. These data strengthen the basis for recommending exerciseas a therapeutic intervention that can improve insulin action for patients with insulin resistanceand/or Type 2 diabetes.
References
1. Kruszynska YT, Olefsky JM. Cellular and molecular mechanisms of non-insulin dependent diabetes mellitus. J Invest Med 1996; 44: 413-428.
2. DeFronzo RA. Pathogenesis of type 2 (non-insulin dependent) diabetes mellitus: a balanced overview.
Diabetologia 1992; 35: 389-397.
3. Shulman GI. Cellular mechanisms of insulin resistance. J Clin Invest 2000; 106: 171-176.
4. Henriksen EJ. Invited review: Effects of acute exercise and exercise training on insulin resistance. J
Appl Physiol 2002; 93: 788-796.
5. Curry DL, Li CH. Stimulation of insulin secretion by beta-endorphin (1-27 and 1-31). Life Sci 1987;
40: 2053-2058.
6. Cheng JT, Liu IM, Tzeng TF, Tsai CC, Lai TY. Plasma glucose lowering effect of β-endorphin in streptozotocin-induced diabetic rats. Horm Meta Res 2002; 34: 570-576.
7. Cheng JT, Liu IM, Hsu CF. Rapid induction of insulin resistance in opioid mu-receptor knock-out mice. Neurosci lett 2003; 339: 139-142.
8. Sforzo GA, Seeger TF, Pert CB, Pert A, Dotson CO. In vivo opioid receptor occupation in the rat brain following exercise. Med Sci Sports Exerc 1986; 18: 380-384.
9. Kohzuki M, Kamimoto M, Wu XM, Xu HL, Kawamura T, Mori N, Nagasaka M, Kurosawa H, Minami N, Kanazawa M, Saito T, Yoshida K. Renal protective effects of chronic exercise and antihypertensive therapy in hypertensive rats with chronic renal failure. J Hypertens 2001; 19: 1877-1882.
Sci 1959; 82: 420-430.
11. Christ CY, Hunt D, Hancock J, Garcia-Macedo R, Mandarino LJ, Ivy JL. Exercise training improves muscle insulin resistance but not insulin receptor signaling in obese Zucker rats. J Appl Physiol 2002;
92: 736-744.
12. Kasiske BL, O'Donnell MP, Keane WF. The Zucker rat model of obesity, insulin resistance, hyperlipidemia, and renal injury. Hypertension 1992; 19: I110- I115.
13. Ivy JL, Sherman WM, Cutler CL, Katz A. Exercise and diet reduce muscle insulin resistance in obese Zucker rat. Am J Physiol Endocrinol Metab 1986; 251: E299-E305.
14. Smith SH, Kramer MF, Reis I, Bishop SP, Ingwall JS. Regional changes in creatine kinase and myocyte size in hypertensive and nonhypertensive cardiac hypertrophy. Circ Res 1990; 67: 1334-1344.
15. Bessesen DH. The role of carbohydrates in insulin resistance. J Nutr 2001; 131: S2782-S2786. 16. Martin WR. Opioid antagonists. Pharmacol Rev 1967; 19: 463-521.
17. Goldstein A. Binding selectivity profiles for ligands of multiple receptor types: focus on opioid receptors. Trends Pharmacol Sci 1987; 8: 456-459.
18. Vargo T, Rossier J, Minick S, Ling N, Rivier C, Vale W, Bloom F. β-endorphin and adrenal corticotropin are secreted concomitantly by the pituitary gland. Science 1977; 197: 1367-1369.
19. Arefolov VA, Dmitriev AD, Tennov AV, Val’dman AV. Detection of the pro-opiomelanocortin peptide fragments--beta-endorphin and ACTH-in the adrenals of rats and mice by immunohistochemistry.
Biull Eksp Biol Med 1986; 101: 445-447.
20. Cheng JT, Liu IM, Kuo DH, Lin MT. Stimulatory effect of phenylephrine on the secretion of
β-endorphin from rat adrenal medulla in vitro. Auton Neurosci Basic & Clinic 2001;93: 31-35.
21. James DE, Jenkins AB, Kraegen EW. Heterogeneity of insulin action in individual muscles in vivo: euglycemic clamp studies in rats. Am J Physiol 1985; 248: E567-E574.
22. King PA, Horton ED, Hirshman MF, Horton ES. Insulin resistance in obese Zucker rat (fa/fa) skeletal muscle is associated with a failure of glucose transporter translocation. J Clin Invest 1992; 90:
1568-1575.
23. Carvalho E, Rondinone C, Smith U. Insulin resistance in fat cells from obese Zucker rats--evidence for an impaired activation and translocation of protein kinase B and glucose transporter 4. Mol Cell
Biol 2000; 206: 7-16.
24. White MF. The insulin signalling system and the IRS proteins. Diabetologia 1997; 40: S2-S17.
25. Klippel A, Kavanaugh WM, Pot D, Williams LT. A specific product of phosphatidylinositol 3-kinase directly activates the protein kinase Akt through its pleckstrin homology domain. Mol Cell Biol 1997;
17: 338-344.
26. Anai, M, Funaki M, Ogihara T, Terasaki J, Inukai K, Katagiri H, Fukushima Y, Yazaki Y, Kikuchi M, Oka Y, Asano T. Altered expression levels and impaired steps in the pathway to phosphatidylinositol 3-kinase activation via insulin receptor substrates 1 and 2 in Zucker fatty rats. Diabetes 1998; 47: 13-23.
27. Goldstein BJ. Insulin resistance: from benign to type 2 diabetes mellitus. Rev Cardiovasc Med 2003;
4: S3-10.
28. Narita M, Ohnishi O, Nemoto M, Aoki T, Suzuki T. The involvement of phosphoinositide 3-kinase (PI3-Kinase) and phospholipase C gamma (PLC gamma) pathway in the morphine-induced supraspinal antinociception in the mouse. Nihon Shinkei Seishin Yakurigaku Zasshi 2001; 21: 7-14. 29. Freye E, Latasch L. Development of opioid tolerance-molecular mechanisms and clinical
consequences. Anasthesiol Intensivmed Notfallmed Schmerzther 2003; 38: 14-26.
30. Liu IM, Liou SS,Chen WC, Chen PF, Cheng JT. Signals in the activation of opioid µ-receptors by loperamide to enhance glucose uptake into cultured C2C12 cells. Horm Meta Res (in press).
31. Ishizuka T, Cooper DR, Hernandez H, Buckley D, Standaert M, Farese RV. Effects of insulin on diacylglycerol-protein kinase C signaling in rat diaphragm and soleus muscles and relationship to glucose transport. Diabetes 1990; 39: 181-190.
32. Vollenweider P, Menard B, Nicod P. Insulin resistance, defective insulin receptor substrate 2-associated phosphatidylinositol-3' kinase activation, and impaired atypical protein kinase C
(zeta/lambda) activation in myotubes from obese patients with impaired glucose tolerance. Diabetes 2002; 51: 1052-1059.
33. Ravichandran LV, Esposito DL, Chen J, Quon MJ. Protein kinase C-zeta phosphorylates insulin receptor substrate-1 and impairs its ability to activate phosphatidylinositol 3-kinase in response to insulin. J Biol Chem 2001; 276: 3543-3549.
34. Heled Y. Shapiro Y. Shani Y. Moran DS. Langzam L. Braiman L. Sampson SR. Meyerovitch J. Physical exercise enhances protein kinase C delta activity and insulin receptor tyrosine phosphorylation in diabetes-prone Psammomys obesus. Metab Clin Exp 2003; 52:1028-1033.
Table 1. General animal characteristics
Zucker rats Exercise Body weight Plasma glucose Plasma insulin Plasma BER status (g.rat-1) (mg.dl-1) (µU.ml-1) (pg.ml-1) Lean Sedentary 183.4 ± 8.4 90.3 ± 6.1 18.3 ± 4.8 50.8 ± 5.6 Obese Sedentary + Vehicle 298.6 ± 9.1** 151.6 ± 5.9** 234.6 ± 11.3** 48.6 ± 6.2 Exercised + Vehicle 263.3 ± 10.2* 113.2 ± 6.7* 191.7 ± 10.9** 95.6 ± 6.9** + Naloxone (1.0 mg kg-1) 312.4 ± 10.1** 158.7 ± 6.2** 242.1 ± 12.5** 90.4 ± 5.6** + Naloxonazine (1.0 mg kg-1) 305.4 ± 9.6** 159.6 ± 7.2** 245.6 ± 13.1** 92.7 ± 7.1** Data are means ± SEM. Assays were performed on tissue from eight different animals in each group. *p < 0.05 and *p < 0.01 compared to the values of sedentary lean rats, respectively.
Table 2. Effects of opioid µ-receptor antagonists on insulin-stimulated GDR at steady state of hyperinsulinemic euglycemic clamp in exercised obese Zucker rats
Zucker rats Exercise status Insulin-stimulated GDR (mg.kg-1.min-1)
Lean Sedentary 25.7 ± 1.7 Obese Sedentary + Vehicle 14.6 ± 0.8** Exercised + Vehicle 23.2 ± 1.9 + Naloxone (1.0 mg kg-1) 17.6 ± 1.9 + Naloxonazine (1.0 mg kg-1) 18.6 ± 1.1 Values (mean ± SEM) were obtained from each group of 8 animals.
Table 3. Quantification of the changes of protein expression and insulin-stimulated phosphorylation in isolated soleus muscle of trained and untrained obese Zucker rats
Lean Zucker rats Obese Zucker rats
Sedentary Sedentary Exercised Exercised
arbitrary units + Vehicle + Naloxone
(1.0 mg.kg-1) IR actin-1 0.83 ± 0.02 0.84 ± 0.03 0.87 ± 0.02 0.84 ± 0.02 IR-pTyr actin-1 1.50 ± 0.03 1.48 ± 0.02 1.50 ± 0.02 1.48 ± 0.03 IRS-1 actin-1 1.28 ± 0.01 1.26 ± 0.02 1.31 ± 0.02 1.27 ± 0.02 IRS-1-pTyr actin-1 1.41 ± 0.01 1.19 ± 0.03** 1.76 ± 0.02** 1.18 ± 0.02** PI3k (p85α) actin-1 0.65 ± 0.01 0.17 ± 0.01** 1.38 ± 0.01** 0.19 ± 0.01** pAkt (Ser473) actin-1 1.08 ± 0.02 0.26 ± 0.01** 1.91 ± 0.01** 0.28 ± 0.02** GLUT 4 actin-1 1.41 ± 0.01 0.98 ± 0.01** 2.01 ± 0.01* 1.40 ± 0.02** Values (mean ± SEM) were obtained from each group of 8 animals.
Figure legends
Figure 1. Representative immunoblot of protein expression and insulin-stimulated phosphorylation in
isolated soleus muscles of trained and untrained Zucker rats. Lanes: sedentary lean Zucker rats (lane 1), sedentary obese Zucker rats (lane 2), exercised obese Zucker rats (lane 3), naloxone (1.0 mg kg-1)-treated exercised obese Zucker rats (lane 4). Similar results were obtained in another seven determinations. Quantification of the data is showed in Table 3.