國立臺灣大學醫學院腫瘤醫學研究所 博士論文
Graduate Institute of Oncology College of Medicine
National Taiwan University Doctoral Dissertation
鈣網蛋白突變在骨髓增生性腫瘤的角色 The Roles of Calreticulin Mutations in
Myeloproliferative Neoplasms
林建鴻 Ken-Hong Lim
指導教授﹕郭遠燁博士、周文堅博士
Advisors: Yuan-Yeh Kuo, Ph.D., Wen-Chien Chou, M.D., Ph.D.
中華民國106年5月
May 2017
誌謝
新約聖經羅馬書八章 28 節題到“萬有都互相效力,叫愛神的人得益處。”這節經文所說 的相當符合我進修博士班的過程和經歷。我很感謝我所相信、我所愛、我所事奉並跟隨的救 主耶穌,帶領我在博士班進修這段過程裡面有很多的學習,讓我在基礎與臨床醫學的研究上,
立下了一個很好的根基,也在信仰上更多經歷祂這位又活又真的神。
我要感謝謝瑞坤院長鼓勵我報考腫瘤醫學研究所,並且在他的推薦之下,順利地進入博 士班進修。當初在找指導老師的時候,一開始我也不清楚要找那一位老師擔任指導老師,因 為印象中腫瘤所的老師們都是以固體腫瘤為研究主題,而我則想要以血液性腫瘤相關的題目 來作為研究主題。後來我就發現在眾多老師中就只有一位郭遠燁老師是專門做血液性癌症的 相關題目,剛好符合我的研究題目和方向。在臨床研究上的指導,我則是請求周文堅教授的 協助,而周教授也很樂意的答應了。我感謝他們兩位指導教授這幾年在研究方向、實驗規劃 和論文寫作上的指導。
我也要感謝擔任我論文指導委員的林亮音教授、徐志宏教授、蕭崇德副教授、陳功深助 理教授,在我的研究和論文上所給予的指導。2009 年 5 月我從美國進修回來的時候,馬偕醫 院很奇妙的開始積極推動使用斑馬魚動物模型來做研究,也請蕭崇德老師負責指導並成立了 斑馬魚房。很感謝蕭老師無私的分享和指導,他是我在斑馬魚研究上的啟蒙老師,一步步帶 領幫助我,讓我能夠接觸到斑馬魚的動物模型,並且嘗試使用它來做血液性腫瘤的研究。也 非常感謝蕭老師指導我如何撰寫研究計畫去申請研究經費,才能使我順利申請到科技部的多 年期計畫補助,和得到馬偕醫院醫研部的研究經費補助,讓我能夠持續進行研究並完成本論 文。
我要感謝馬偕醫院血液暨腫瘤科張義芳主任和許多的同事,在我研究過程中在病人收案 上給予的協助,也在工作上讓我有一些彈性去安排和協調。而陳功深醫師在研究和實驗上的 協助也是我能完成本論文的重要推手。我也要感謝在馬偕的團隊成員林煥超醫師、江翊豪醫 師和張育誠醫師,因為他們的參與和支持才使我的研究和實驗能夠持續的進行。在此特別的 感謝許多的研究助理,瑋婷、巧宜、靖淞、黃齡、和楨蔚,還有林宗毅博士和程俊嘉博士都 在整個研究和實驗過程中給予我幫助。
我特別要感謝我的太太琬緞,在這將近六年的時間裏扶持我,幫我照料、打理這個家和 小孩,讓我不用太多去掛心孩子和家人的事情。過程中一路有許多人的幫助,台北市召會四 會所和廿四會所許多的弟兄姊妹,常常為我提名代禱,希望我在博士班的過程中能夠順利畢 業。我也很感謝我的四個小孩,他們在這個過程中,帶給我很多的的鼓勵和歡樂。大女兒林
欣也能夠在過程中給我一些文字編輯上的協助。很感謝我的父母親給我從小到大在教育上的 栽培,願意把這樣的一篇論文獻給最親愛的父母親。
建鴻 於二 0 一七年四月四日
中文摘要
骨髓增生性腫瘤是克隆性造血幹細胞疾病,並且可以分類為“典型”和“非典型”骨髓 增生性腫瘤。典型骨髓增生性腫瘤通常會在周邊的血液中表現出終端骨髓細胞的擴增,包括 真性紅血血球增多症、原發性血小板增多症、原發性骨髓纖維化和慢性骨髓性白血病。近年
來,在大約 30%的 JAK2/MPL-未突變的骨髓增生性腫瘤中發現鈣網蛋白(Calreticulin, CALR)
突變。目前,鈣網蛋白突變已經成為診斷原發性血小板增多症和原發性骨髓纖維化的重要克 隆標誌物。
這項研究的第一個目的是開發一種快速和敏感的篩選工具,用於檢測鈣網蛋白突變。我 們使用 CFX Connect 即時系統成功開發了高分辨率熔解分析,以檢測原發性血小板增多症病 人的鈣網蛋白第 9 外顯子突變。我們的高分辨率熔解分析系統在識別病人基因組 DNA 中的 鈣網蛋白第 1 型和第 2 型突變體的最大敏感性為 2.5%。我們的高分辨率熔解分析系統假陽性 率為 3%,並且無假陰性出現。
本研究的第二個目的是評估台灣原發性血小板增多症病人鈣網蛋白突變和 JAK2 /CALR 共突變的臨床和預後意義。我們在 92 例成年原發性血小板增多症病人中利用高分辨率熔解分 析篩選了鈣網蛋白第 9 外顯子的改變,隨後並且進行了 TA 克隆。我們在 21 例(22.8%)病 人中鑑定出典型的鈣網蛋白插入/刪除型突變。鈣網蛋白突變與年齡較輕(p = 0.025),血小 板數較高(p <0.001)和較低的血紅素(p = 0.016)有相關。有趣的是,我們檢測到在 59 例 JAK2 突變的原發性血小板增多症病人中有 13 例(22%)的鈣網蛋白第 9 外顯子的改變。與 這些具有 JAK2 /CALR 共突變的原發性血小板增多症病人有相關的因子包括年齡較大(p = 0.025),診斷後發生血栓事件較多(p = 0.048),診斷後主要動脈血栓事件較多(p = 0.022),
且較多屬於血栓出血併發症高風險組病人(p = 0.023)。我們的研究顯示 JAK2 突變的原發性 血小板增多症病人可以出現頻繁的鈣網蛋白第 9 外顯子的改變,並且界定出一群具有血栓事 件風險增加的病人亞群。
本研究的第三個目的是要探討鈣網蛋白突變之原發性血小板增多症病人的 B 細胞免疫特 徵。我們篩選了 54 例台灣成年原發性血小板增多症病人的鈣網蛋白突變並評估了其 B 細胞 免疫特徵。在這 54 例原發性血小板增多症病人中有 19 例(35.2%)具有 8 種不同類型的鈣 網蛋白笫 9 外顯子突變,其中包括 4 例(7.4%)同時伴有 JAK2V617F 共突變的病人。經過
年齡、性別、追踨期和血液學參數校正後,我們進行的多變量分析證實,與健康成年人相比,
活化的 B 細胞在 JAK2 突變,鈣網蛋白突變和三陰性原發性血小板增多症病人中都有增加的 現象。因此,活化的 B 細胞增加在不同突變亞組的原發性血小板增多症病人中是一種普遍存 在的現象。
本研究的第四個目的是使用斑馬魚動物模型研究鈣網蛋白突變的分子發病機制。我們確 定了 3 種與人類鈣網蛋白直系同源的斑馬魚基因,稱為 calr,calr3a 和 calr3b。CALR-del52 和 CALR-ins5 突變體的表達使斑馬魚早期的造血幹/前驅細胞增加,並進而造成血小板球增多 但不影響正常的血管生成。我們發現使用 morpholino 降低 mpl 但不是 epor 或 csf3r 可以顯著 的減弱 CALR 突變體對血小板球增多的影響。此外,CALR 突變體的表達也活化 jak/stat 信息 傳遞路徑,而使用 JAK 抑製劑(ruxolitinib 和 fedratinib)可以抑制此活化現象。這些研究結 果說明 CALR 突變體通過 mpl 依賴機制活化 jak 信息傳遞路徑導致斑馬魚致病性之血小板球 生成。我們研究的結果也說明與突變型 CALR 腫瘤發生相關的訊息傳遞機制在人與斑馬魚之 間是保守的。
關鍵詞:
B 細胞、鈣網蛋白、原發性血小板增多症、高分辨率融合分析、免疫、突變、斑馬魚
Abstract
The myeloproliferative neoplasms (MPNs) are clonal hematopoietic stem cell disorders and can be classified into "classic" and "atypical" MPNs. Classic MPNs usually exhibit terminal myeloid cell expansion in the peripheral blood and include polycythemia vera, essential thrombocythemia (ET), primary myelofibrosis and chronic myeloid leukemia. Calreticulin (CALR) mutations have recently been discovered in about 30% JAK2/MPL-unmutated myeloproliferative neoplasms (MPN), and have become an important clonal marker for the diagnosis of essential thrombocythemia (ET) and primary myelofibrosis.
The first aim of this study is to develop a rapid and sensitive screening tool for the detection of CALR mutations. We successfully developed a high-resolution melting analysis (HRMA) with the CFX Connect real-time system to detect CALR exon 9 mutations in ET patients. The maximal sensitivity of our HRMA system in identifying both CALR type 1 and type 2 mutants from patients’
genomic DNA was 2.5%. Our HRMA has a false positive rate of 3% and no false negative.
The second aim of this study is to evaluate the clinical and prognostic significance of CALR mutations and JAK2/CALR co-mutations in Taiwanese ET patients. We screened for CALR exon 9 alterations with HRMA followed by TA-cloning in 92 adult ET patients. We identified classic CALR indel mutations in 21 (22.8%) patients. CALR mutations were associated with younger age (p=0.025), higher platelet count (p<0.001) and lower hemoglobin level (p=0.016). Interestingly, we detected various CALR exon 9 alterations in 13 (22%) of 59 JAK2-mutated ET patients.
JAK2-mutated ET patients with concomitant CALR alterations were associated with oldest age (p=0.025), higher thrombotic events after diagnosis (p=0.048), higher major arterial thrombotic events after diagnosis (p=0.022) and more patients being high risk group for thrombo-hemorrhagic complications (p=0.023). Frequent CALR exon 9 alterations in JAK2-mutated ET patients define a
specific subgroup of patients with increased risk of thrombotic events.
The third aim of this study is to determine the B cell immune profiles in CALR mutated ET patients.
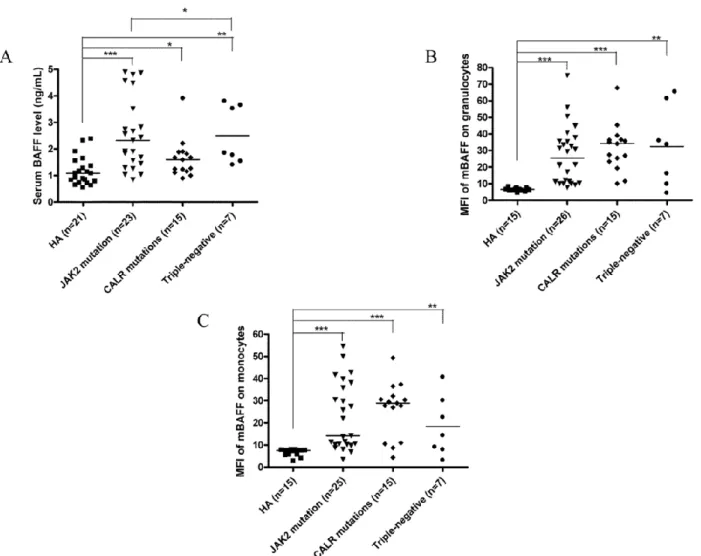
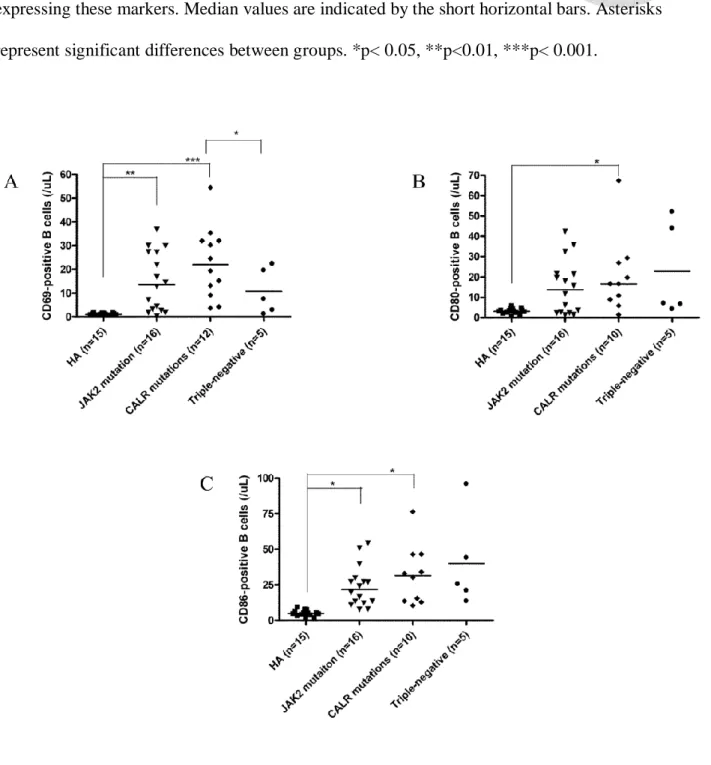
We screened for CALR mutations and evaluated B cell immune profiles in a cohort of 54 adult Taiwanese ET patients. 19 (35.2%) of 54 ET patients harbored 8 types of CALR exon 9 mutations including 4 (7.4%) patients with concomitant JAK2V617F mutations. Multivariate analysis adjusted for age, sex, follow-up period and hematological parameters confirmed that increased activated B cells were universally present in JAK2-mutated, CALR-mutated and triple-negative ET patients when compared to healthy adults. In conclusion, increased B cell activation is present in ET patients across different mutational subgroups.
The fourth aim of this study is to investigate the molecular pathogenesis of CALR mutations using zebrafish animal models. We identified 3 zebrafish genes orthologous to human CALR, referred to as calr, calr3a and calr3b. Expression of the CALR-del52 and CALR-ins5 mutants caused an increase in the hematopoietic stem/progenitor cells followed by thrombocytosis without affecting normal angiogenesis. The expression of CALR mutants also perturbed early developmental hematopoiesis in zebrafish. Importantly, morpholino knockdown of mpl but not epor or csf3r could significantly attenuate the effects of mutant CALR. Furthermore, expression of mutant CALR caused jak-stat signaling activation in zebrafish that could be blocked by JAK inhibitors (ruxolitinib and fedratinib). These findings showed that mutant CALR activates jak-stat signaling through an mpl-dependent mechanism to mediate pathogenic thrombopoiesis in zebrafish, and illustrated that the signaling machinery related to mutant CALR tumorigenesis are conserved between human and zebrafish.
Keywords: B cell, CALR, essential thrombocythemia, high-resolution melting analysis, immune,
mutation, zebrafish
Contents
誌謝...i
中文摘要...iii
Abstract ...v
Contents ...viii
List of Figures ...xi
List of Tables ...xiii
Chapter 1. Classic BCR-ABL-negative myeloproliferative neoplasms (MPNs) ……..……..…1
1. MPNs ...1
1.1 Introduction of MPNs ...1
1.2 Driver mutations in MPNs ………...1
2. CALR mutations in MPNs ...2
2.1 Calreticulin ...2
2.2 The function of CALR ……….……..2
2.3 CALR mutations in JAK2/MPL-unmutated essential thrombocythemia and primary myelofibrosis patients ………....3
2.4 Screening for CALR mutations in MPNs ...………....4
2.5 Clinical and and prognostic significance of CALR mutations ...…..………..4
3. Molecular pathogenesis of CALR mutations in MPNs ...5
3.1 Mutated CALR is an initiating event in MPNs ...5
3.2 Mutant CALR causes overactivation of JAK-STAT signaling ………..5
3.3 In vitro and in vivo models of CALR mutations ………...…..6
4. Zebrafish animal models ………...………..….7
4.1 Zebrafish is a valuable model to study hematopoiesis and leukemogenesis ...…..7
5. B cell immune profiles in essential thrombocythemia patients ....……..………….8
5.1 Increased B cells activation in essential thrombocythemia patients ....…………..8
6. Aims of the study ...9
Chapter 2. High-resolution melting analysis as a rapid and sensitive screening tool for the detection of CALR mutations ...11
1. Summary ...11
2. Introduction ...11
3. Patients and Methods ...13
4. Results ...16
5. Discussion ...17
Chapter 3. The clinical and prognostic significance of CALR mutations and JAK2/CALR co-mutations in Taiwanese essential thrombocythemia patients ………...21
1. Summary ...21
2. Introduction ...22
3. Patients and Methods ...23
4. Results ...24
5. Discussion ...26
Chapter 4. B cell immune profiles in CALR mutated essential thrombocythemia patients...29
1. Summary ...29
2. Introduction ...30
3. Patients and Methods ...31
4. Results ...35
5. Discussion ...39
Chapter 5. The molecular pathogenesis of CALR mutations using zebrafish animal models ...47
1. Summary ...47
2. Introduction ...47
3. Materials and Methods ...49
4. Results ...52
5. Discussion ...56
Chapter 6. Conclusions and future work ...61
Figures ...63
Tables ...82
References ...106
Appendices ...124
List of Figures
Figure 1. CALR exon 9 mutations generate a novel peptide sequence in the C-terminus with the loss of acidic domain and the KDEL ER retention sequence. ………...……..63 Figure 2. Results of the dilution series of CALR mutants from plasmid DNA in a background of
plasmid CALR wild-type DNA detected by HRMA in triplicate samples. …………...…64 Figure 3. Results of the dilution series of CALR mutants from patients’ genomic DNA in a background of wild-type DNA from healthy control detected by HRMA in triplicate samples. ……….…65 Figure 4. Representative normalized melting curves and difference curves from the 22 patient samples. ……….66 Figure 5. The comparison of HRMA with Sanger sequencing in 6 patient samples with discordant results between HRMA and Sanger sequencing. ……….….67 Figure 6. Normalized difference curves of 16 JAK2 V617F-mutated essential thrombocythemia patient samples showing distinct melting curves from CALR exon 9 wild-type samples. ……….……68 Figure 7. Variants of CALR exon 9 alterations in essential thrombocythemia patients detected by high-resolution melting analysis followed by TA-cloning. ………..69 Figure 8. Elevated serum BAFF levels and higher membrane-bound BAFF expression in peripheral
granulocytes and monocytes of ET patients. ……….….……..70 Figure 9. Fractions of activated B cells with TLR4 expression, and IL-1β and IL-6 production are
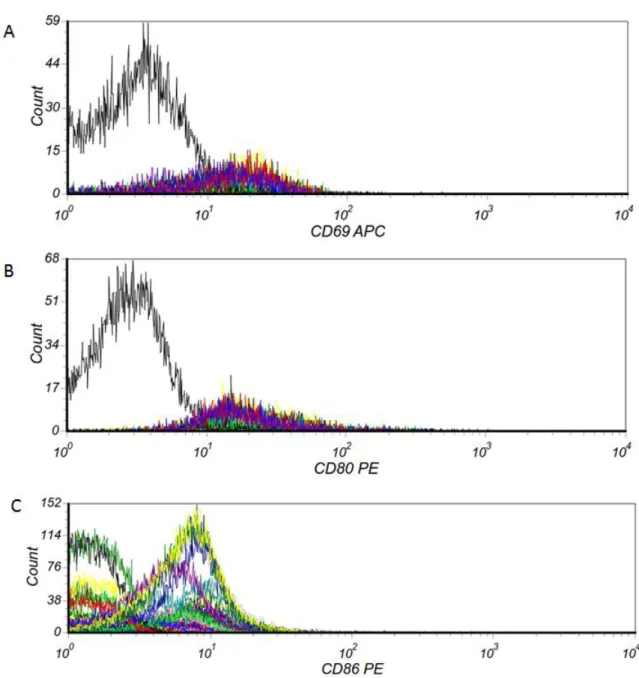
higher in ET patients. ………71 Figure 10. CALR mutations are associated with activated B cells in patients with ET. ………...….72 Figure 11. Gated CD19+ B lymphocytes in whole blood cells using flow cytometry. ………...…..73 Figure 12. Expression of CD69, CD80 and CD86 on CD19+ B lymphocytes using flow cytometry was showed in histograms. ………...….74
Figure 13. The map for pSYC-102-CALR vectors. ………..………75 Figure 14. Identification of 3 zebrafish calr genes. ………..76 Figure 15. Effects of the expression of mutant CALR on the number of hematopoietic
stem/progenitor cells and angiogenesis. ………77 Figure 16. Mutant CALR requires mpl to cause thrombocytosis in zebrafish. ……….…79 Figure 17. The expression of mutant CALR activates jak-stat signaling in zebrafish. ……….80
List of Tables
Table 1. Clinical and laboratory characteristics at diagnosis or referral in 21 essential thrombocythemia patients with 6 types of CALR mutations. ………82 Table 2. CALR exon 9 alterations and single nucleotide polymorphism in 14 JAK2 V617F-mutated essential thrombocythemia patients detected by high-resolution melting analysis. …….83 Table 3. Clinical and laboratory characteristics at diagnosis or referral of 91 essential
thrombocythemia patients stratified by mutation profiles. …………...………85 Table 4. Clinical and laboratory characteristics in healthy adults and patients with essential
thrombocythemia. ……….………..…….87 Table 5. Univariate analysis of B cell immune profiles in healthy adults and patients with essential
thrombocythemia. ……….………..….89 Table 6. Laboratory characteristics and B cell immune profiles in essential thrombocythemia
patients with and without treatment with hydroxyurea. ……...………...92 Table 7. Correlation of platelet count at testing, serum BAFF levels, and B cell immune profiles in
this study. ………...………,...94 Table 8. Comparison of B cell immune profiles between healthy adults with patients with essential
thrombocythemia using linear regression model adjusted for hematological parameters. ………..……….95 Table 9. Comparison of B cell immune profiles among patients with essential thrombocythemia
using linear regression model adjusted for age, sex, follow-up period and hematological parameters. ………..…….97 Table 10. Characteristics and the frequency of CALR and JAK2V617F co-mutations in patients with
essential thrombocythemia. ………...…...98 Table 11. Univariate analysis of B cell immune profiles in healthy adults, reactive thrombocytosis,
polycythemia vera and essential thrombocythemia. ………...100
Table 12. Morpholino sequences for mpl, epor and csf3r knockdown. ………....…………..……101 Table 13. Real-time quantitative polymerase chain reaction primer sequences for zebrafish
hematopoietic lineage-specific, thrombopoiesis, cytokine and cytokine receptor
genes. ………..…102 Table 14. Effects of CALR mutant mRNA injection on the expression of genes in zebrafish
embryo. ………..….104
Chapter 1
Classic BCR-ABL-negative myeloproliferative neoplasms 1. Myeloproliferative neoplasms
1.1 Introduction of myeloproliferative neoplasms
The myeloproliferative neoplasms (MPNs) are clonal hematopoietic stem cell disorders.
According to the 2008 World Health Organization (WHO) classification, the MPNs are classified into "classic" and "atypical" MPNs.1 The classic MPNs usually exhibit terminal myeloid cell expansion in the peripheral blood and include polycythemia vera (PV), essential thrombocythemia (ET), primary myelofibrosis (PMF) and BCR-ABL-positive chronic myeloid leukemia (CML). The classic BCR-ABL-negative MPNs are heterogenous diseases with overlapping clinical and molecular characteristics.2 PV is characterized by increased hematocrit/red blood cell mass accompanied by the presence of a JAK2 mutation along with a decreased erythropoietin level. PMF is characterized by the presence of bone marrow fibrosis that cannot be attributed to another myeloid disorder such as CML, ET or myelodysplastic syndrome. ET is characterized by the presence of a clonal or autonomous thrombocytosis and the exclusion of other myeloid diseases. The chronic-phase PV and ET are associated with an increased risk of arterial and venous thrombosis, hemorrhagic complication, and sometimes with constitutional symptoms.2,3 Myelofibrosis may be primary or may follow PV/ET, and may be considered as an accelerated-phase MPN with an increased risk of leukemic transformation and decreased survival.4
1.2 Driver mutations in MPNs
The JAK2 V617F mutation was discovered in 2005, and has provided important diagnostic, therapeutic, and prognostic implications in MPNs. The frequency of JAK2 V617F mutation is over 90% in PV, and about 60% in ET and PMF.5-7 Besides, MPL mutations are identified in
about 4-5% of JAK2-unmutated ET and PMF patients.8 Following these seminal reports, other somatic mutations such as LNK, TET2 and DNMT3A have also been detected in patients with MPN.9 However, they are not mutually exclusive with JAK2 and MPL mutations and also not specific to patients with MPN.9,10 Despite many somatic mutations have been identified in patients with ET, clonal molecular marker is still not identified in ~40% of ET patients.
2. CALR mutations in MPNs 2.1 Calreticulin
Calreticulin (CALR) is a 46-kDa developmentally highly conserved, multicompartmental and multifunctional protein. Its gene is located on chromosome 19p13.2 and contains nine exons.
Mature CALR consists of three structurally and functionally distinct domains:11
(i) Residues 1-180: the globular N-terminal domain is lectin binding and consists of a signal sequence for targeting to the ER.
(ii) Residues 181–290: the middle proline-rich or P-domain contains high affinity, low capacity, binding sites for Ca2+.
(iii) Residues 291–400: the highly acidic C-terminal domain contains a number of high capacity, low-affinity Ca2+ binding sites, and terminates in a KDEL ER retention sequence, which can be used by KDEL receptors to retrieve CALR from the cis-Golgi back to the ER.
The N- and P-domains are mainly responsible for the protein’s chaperone function, whereas the C-domain is principally involved in calcium regulation in the ER.
2.2 The function of CALR
CALR is best known for its role as a Ca2+ binding chaperone protein located primarily in the endoplasmic reticulum (ER).12 Inside the ER, CALR plays an integral role in calcium
homeostasis and protein folding. CALR can also be found in the nucleus, localize to cell surface and accumulate in extracellular compartments. Outside the ER, CALR can regulate integrin-mediated cell adhesion, gene nuclear transport, programmed cell removal, and immunogenic cell death.12-14 CALR was also found to involve in the immune response to pre-apoptotic cancer cells, and early cell surface exposure of CALR is followed by expression and release of heat-shock proteins (e.g. HSP70), and high-mobility group I (HMGB1) protein.15 Recombinant CALR fragment was shown to exhibit potent stimulatory activities against B cells.16,17
2.3 CALR mutations in JAK2/MPL-unmutated ET and PMF patients
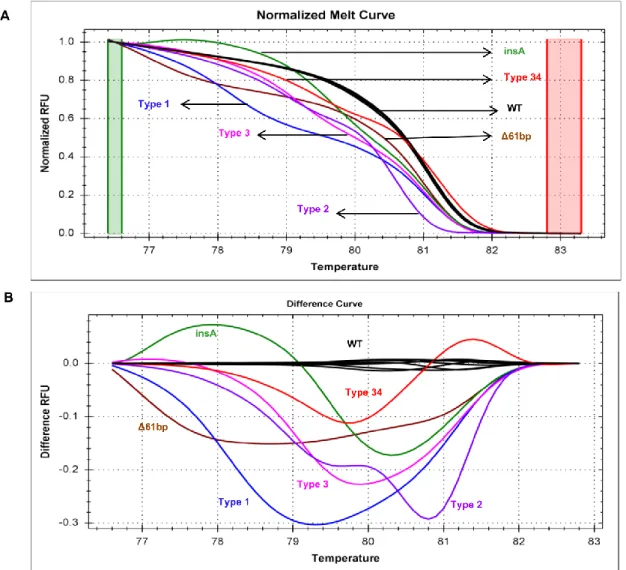
In 2013, two research groups discovered a high frequency of somatic CALR mutations in 60–88% patients with JAK2/MPL-unmutated ET and PMF patients, but none in patients with PV.18,19 The pattern of most CALR mutations in MPN is heterozygous base pair insertions or deletions (indels) in exon 9 causing one base pair (bp) reading frameshift. As a consequence, CALR exon 9 mutations generate a novel peptide sequence in the C-terminus with the loss of acidic domain and the KDEL ER retention sequence (Figure 1).20
More than 50 different mutational types in CALR have been detected. The most common types of CALR mutations are a 52 bp deletion (L367fs*46, type 1 mutation) and a 5 bp insertion of TTGTC (K385fs*47, type 2 mutation) accounting for more than 80% of all pat ients with mutant CALR. Rarely, CALR exon 9 point mutations have been reported in follicular lymphoma (E403X and E405Q) 21, PMF (E379D) 18 and chronic neutrophilic leukemia (E398D).22 Importantly, CALR mutations are not only mutually exclusive with JAK2 and MPL mutations, but they are also infrequently detected in other myeloid neoplasms such as myelodysplastic syndrome, chronic myelomonocytic leukemia and atypical chronic myeloid
leukemia.18,19 In addition, CALR mutations have not been found in other myeloid or lymphoid malignancies, solid tumors, or healthy controls. These findings indicate that CALR mutations are selective for ET and PMF. Based on these discoveries, CALR mutations have been included in the 2016 WHO classification system for the molecular diagnosis of ET and PMF.23,24 The JAK2 46/1 haplotype does not predispose to CALR-mutated MPNs in one study.25
2.4 Screening for CALR mutations in MPNs
Several methods have been used to detect CALR exon 9 mutations including direct DNA sequencing, PCR followed by fragment analysis and immunostaining.18,19,26,27
Although fragment analysis has a relatively high sensitivity for CALR mutations detection, it cannot discriminate point mutation from wild-type sequence. High-resolution melting analysis has also been used for the screening of CALR mutations.28
2.5 Clinical and and prognostic significance of CALR mutations
CALR mutations have been shown to have important clinical and prognostic significance in ET and PMF patients.18,19,29 In ET patients, CALR mutations are associated with younger age, more frequently male, higher platelet counts, lower hemoglobin and leukocyte counts, and a lower risk of thrombosis than those with the JAK2 mutation, with no difference in the rate of transformation to post-ET MF.19,29-31 In PMF patients, CALR mutations are associated with younger age, higher platelet count, and lower DIPSS-plus score.32 CALR-mutated PMF patients are also less likely to be anemic, require transfusions, or display leukocytosis. In an international study of 570 PMF patients, CALR+ASXL1- patients had the longest survival (median 10.4 years) and CALR-ASXL1+ patients had shortest survival (median 2.3 years).33 In another study demonstrating the additional value of the number of prognostically detrimental mutations in PMF, CALR mutations favorably affected survival, independently of both number
of mutations and IPSS/DIPSS-plus.34 The prognostic advantage of CALR mutations in PMF patients might be confined to type 1 or type 1-like CALR variants.35
3. Molecular pathogenesis of CALR mutations in MPNs 3.1 Mutated CALR is an initiating event in MPNs
CALR mutations were acquired at the level of the hematopoietic stem cell (HSC) and clonal characterization of MPN samples has shown that mutated CALR is present in the earliest clone, which is consistent with it being an initiating event in MPNs.18,19 Mutant CALR was shown to be highly expressed in the megakaryocyte lineage in bone marrow trephines immunostained with a polyclonal antibody against a 17-amino-acid peptide derived from mutant CALR’s novel C-terminus.26 In this study, wild-type CALR was also found to have a megakaryocyte-restricted expression pattern, which may explain why the mutant form is associated with MPNs characterized predominantly by abnormal megakaryopoiesis.26
3.2 Mutant CALR causes overactivation of JAK-STAT signaling
Although the C-terminus Golgi-to-ER retention signaling motif (KDEL) is lost in the CALR mutant protein, it appears that mutant CALR remains largely within the ER.18,19 The characteristic basic and positively charged novel C-terminal peptide sequence has been predicted to alter the calcium-binding capacity of the protein.36 How the novel mutant CALR C-terminus result in myeloid proliferation with a specificity for abnormal megakaryopoiesis remain unclear. Preliminary evidence suggests that mutant CALR may also lead to overactivation of JAK-STAT signaling. By using transcriptional studies, Rampal et al. have also shown that activated JAK2 signaling is seen in all MPN patients, including those with CALR mutations.37 Besides, overexpression of the most frequent CALR deletion, but not wild-type CALR caused cytokine-independent growth in cell lines owing to the activation of
signal transducer and activator of transcription 5 (STAT5) by means of an unknown mechanism.18 Nevertheless, the MARIMO cell line which harbors a CALR mutation has been found to not dependent on JAK/STAT signaling and was more resistant to JAK inhibitors, in marked contrast to JAK2-mutated cell lines.38 Upon addition of thapsigargin (which blocks ER Ca2+-ATPase channels resulting in ER calcium depletion and increased cytosolic calcium levels), MARIMO cells also showed the slowest rate of increase of cytoplasmic calcium levels consistent with the concept that mutant CALR alters ER dependent calcium homeostasis.38 This study raised the possibility that mutations of CALR may have activation of pathways other than the STATs.
3.3 In vitro and in vivo models of CALR mutations
Recent studies have focused on the underlying mechanism of CALR mutations in the pathophysiology of MPNs. With the use of in vitro cell lines and retroviral mouse models, CALR mutants were found to activate the JAK-STAT signaling in an MPL-dependent manner.39-43 Araki and co-workers used the TPO-dependent megakaryocytic cell line UT-7/TPO to demonstrate that mutant CALR, but not wild-type, activates MPL and downstream signaling molecules including JAK2, STAT5, and extracellular signal-regulated kinase 1/2, and subsequently promotes the TPO‐ independent growth of UT‐ 7/TPO cells.39 Besides, they also showed that mutant CALR preferentially binds to MPL, and that the mutant‐ specific domain of CALR is required for this interaction. Marty et al. engrafted lethally irradiated recipient mice with bone marrow cells transduced with retroviruses expressing CALR type 1 and type 2 mutants.40 In contrast to wild-type CALR, CALR type 1 and, to a lesser extent, CALR type 2 induced thrombocytosis due to a megakaryocyte hyperplasia. It is noteworthy that CALR type 1 mutation-expressing mice rapidly developed marked thrombocytosis and then progressed to a condition similar to human myelofibrosis. By contrast, CALR type 2
mutation-expressing mice had a mild ET phenotype with low propensity to disease progression.
In the report by Chachoua et al., they found that pathogenic CALR mutants specifically activate the TPO receptor by a mechanism dependent on the presence of the extracellular N-glycosylation residues of MPL and the glycan-binding site at the new C-terminal tail of the mutant CALR.41 They also found that CALR mutants can activate JAK2, and downstream STAT5/3/1, mitogen-activated protein kinase, and phosphatidylinositol-3 kinase signaling via MPL. Finally, Elf et al. demonstrated that expression of mutant CALR alone is sufficient to engender MPN in mice and recapitulates the disease phenotype of patients with CALR-mutant MPN.42 They further showed that the thrombopoietin receptor MPL is required for mutant CALR-driven transformation through JAK-STAT pathway activation. Interestingly, Elf et al.
also showed that the oncogenicity of mutant CALR is dependent on the positive electrostatic charge of the C-terminus of the mutant protein, which is necessary for physical interaction between mutant CALR and MPL.
4. Zebrafish animal models
4.1 Zebrafish as a valuable model to study hematopoiesis and leukemogenesis
Zebrafish has proven to be a useful vertebrate model in which to elucidate the molecular mechanisms of hematologic malignancies based on the high degree of genetic and morphological similarity in hematopoiesis between the zebrafish and human.44,45 Over the last decade, studies using the zebrafish model have contributed to our understanding of vertebrate hematopoiesis, myelopoiesis, and leukemogenesis.46-48 A high degree of similarity in the gene signatures of specific types of tumor cells in fish and humans have been demonstrated after comparisons of cancer-associated gene expression profiles indicating that the contributing genetic pathways leading to cancer are evolutionarily conserved.49 Therefore, zebrafish can provide valuable knowledge about the mechanisms behind pathogenesis of leukemia.
Furthermore, zebrafish animal model offers obvious advantages as a result of its rapid external development, small size and optical transparency of the embryos, which are permeable to small molecules and drugs. As a result, chemical screenings have been successfully conducted by using appropriated zebrafish lines.50 The unique advantage of in vivo imaging in zebrafish also helps to dissect the molecular pathways underlying tumor initiation, progression and metastasis.
4.2 Genome editing tools in zebrafish
The zebrafish is an affordable, efficient, and genetically modifiable vertebrate model for studying hematopoiesis and leukemogenesis when compared to the traditional mammalian models. In addition, many molecular methods and models have been established to facilitate both forward and reverse genetic studies in zebrafish. For example, the expression of proteins can be “knocked down” transiently in the embryos by morpholino antisense oligonucleotides (MO) and microRNA or permanently “knockouted” by using the recently developed transcription activator-like effector nucleases (TALEN) technology or clustered regularly interspaced short palindromic repeat (CRISPR)/Cas9 system.51-59 Highly efficient Tol2 transgenic tool is also available for use in the study of hematopoiesis and leukemogenesis in zebrafish.46,60 The advance in technology has significantly improved the genetic tractability in zebrafish. One of the main limitations in zebrafish model is that antibody markers for hematopoiesis are mostly not available. However, this lack of markers can be compensated in some cases by the use of fluorescent transgenic blood cell reporter lines to identify hematopoietic cell types in zebrafish.61
5. B cell immune profiles in ET patients
5.1 Increased B cells activation in ET patients
We have reported that activated B cells are increased in ET patients, and can facilitate platelet production mediated by cytokines, such as interleukin (IL)-1beta and IL-6 regardless JAK2V617F mutational status.62 We found that increased production of B cell-activating factor (BAFF) by granulocytes and monocytes up-regulates toll-like receptor 4 (TLR4) expression on B cells of ET patients and promotes B cell activation, which play a pathogenic role augmenting thrombocytosis in ET by producing IL-1beta and IL-6. However, whether CALR mutations are also associated with activated B cells in ET patients requires further study.
6. Aims of the study
6.1 To develop a rapid and sensitive screening tool for the detection of CALR mutations
CALR mutations have become an important clonal maker for the diagnosis of MPNs especially in JAK2/MPL-unmutated ET and PMF. Our first aim is to develop a rapid and sensitive screening tool using HRMA for the detection of CALR exon 9 mutations.6.2 To evaluate the clinical and prognostic significance of CALR mutations and JAK2/CALR co-mutations in Taiwanese ET patients
Although the clinical and prognostic significance of CALR mutations in Caucasian ET patients have been studied,63,64 there is still a need to evaluate the clinical and prognostic significance of CALR mutations in Taiwanese ET patients. Besides, JAK2 and CALR co-mutations have been reported in a few MPN patients in several studies. With the use of a highly sensitive HRMA, we have identified higher frequency of JAK2 and CALR co-mutations in ET patients. We therefore want to evaluate the clinical and prognostic significance of JAK2/CALR co-mutations in Taiwanese ET patients.
6.3 To determine the B cell immune profiles in CALR mutated ET patients
We have reported that activated B cells are increased in ET patients, and can facilitate platelet production mediated by cytokines, such as interleukin (IL)-1β and IL-6 regardless JAK2V617F mutational status.62 The discovery of CALR mutations in JAK2/MPL-unmutated ET patients in December 2013 have prompted us to ask the question that whether increased B cell activation can also be found in ET with CALR mutations similar to that in JAK2V617F-mutated ET.18,19,65
6.4 To investigate the molecular pathogenesis of CALR mutations using zebrafish animal models
Although the expression of CALR mutants resulted in pathogenic thrombocytosis in adult mice, whether CALR mutants may disrupt normal hematopoiesis during early development remains unknown. We aim to evaluate the pathophysiologic effects of mutant CALR during embryonic hematopoietic development and to test the therapeutic effects of JAK inhibitors on mutant CALR using the in vivo zebrafish model.
Chapter 2
High-resolution melting analysis as a rapid and sensitive screening tool for the detection of CALR mutations
1. Summary
Somatic CALR exon 9 mutations have recently been identified in patients with JAK2/MPL-unmutated myeloproliferative neoplasm, and have become an important clonal marker for the diagnosis of essential thrombocythemia (ET) and primary myelofibrosis. In the present study, we sought to use high-resolution melting analysis (HRMA) as a screening method for the detection of CALR mutations. 32 JAK2/MPL-unmutated ET patients were retrospectively enrolled and 8 healthy adults were used as wild-type control.
CALR exon 9 mutation was independently screened by HRMA with the CFX Connect real-time system and Sanger sequencing. TA-cloning was used to detect CALR exon 9 mutations in patients suspected to have low mutant allele burden. The maximal sensitivity of HRMA in identifying both CALR type 1 and type 2 mutants from patients’ genomic DNA was 2.5%. Twenty-two samples were found to have distinct melting curves from wild-type. The presence of CALR mutations in 16 of these 22 samples were confirmed by Sanger sequencing, while the other 6 samples were wild-type by sequencing. After TA-cloning, CALR mutations were detected in 5 of 6 patients from 1 (6%) of 16 clones to 1 (2%) of 50 clones. Therefore, HRMA identified CALR mutations in 21 (65.6%) of 32 ET patients compared to 16 (50%) patients by Sanger sequencing, with a false positive rate of 3% and no false negative. In conclusion, the HRMA developed in our system is a rapid and sensitive technique for the detection of CALR exon 9 mutations.
2. Introduction
hematopoietic stem cell disorder and includes polycythemia vera (PV), essential thrombocythemia (ET) and primary myelofibrosis (PMF).2 ET is characterized by increased number of mature megakaryocytes in the bone marrow and sustained thrombocytosis in the peripheral blood. Although most ET patients have a normal life expectancy, some may encounter serious events such as thrombotic and hemorrhagic complications and leukemic transformation during their clinical course.3 The JAK2 V617F mutation was discovered in 2005, and has provided important diagnostic, therapeutic, and prognostic implications in MPNs. The frequency of JAK2 V617F mutation is over 90% in polycythemia vera (PV), and about 60% in ET and PMF.5-7 Besides, MPL mutations are identified in about 4-5% of JAK2-unmutated ET and PMF patients.8 Following these seminal reports, other somatic mutations such as LNK, TET2 and DNMT3A have also been detected in patients with MPN.9 However, they are not mutually exclusive with JAK2 and MPL mutations and also not specific to patients with MPN.9,10 Despite many somatic mutations have been identified in patients with ET, clonal molecular marker is still not identified in ~40% of ET patients.
Recently, a high frequency (around 49-88%) of somatic calreticulin (CALR) mutations was identified in patients with JAK2/MPL-unmutated patients with ET and PMF.18,19,32,66,67
Most CALR mutations in MPNs are heterozygous indels in exon 9 causing one base pair reading frameshift and resulted in the generation of a novel CALR protein C-terminus. The majority of the CALR exon 9 mutants were a 52 bp deletion of nt1172 to nt1223 (L367fs*46, type 1 mutation) and a 5 bp insertion of TTGTC (K385fs*47, type 2 mutation).
Rarely, CALR exon 9 point mutations have been reported in follicular lymphoma (E403X and E405Q),21 PMF (E379D)18 and chronic neutrophilic leukemia (E398D).22 Importantly, CALR mutations are not only mutually exclusive with JAK2 and MPL mutations, but they
are also infrequently detected in other myeloid neoplasms such as myelodysplastic syndrome, chronic myelomonocytic leukemia and atypical chronic myeloid leukemia.18,19 These findings indicate that CALR mutations are quite specific for ET and PMF. Based on these discoveries, CALR mutations have been proposed to be included in the World Health Organization classification system for the molecular diagnosis of ET and PMF.23 Therefore, the detection of CALR mutations with reliable and cost-effective methods in patients suspected to have ET or PMF is very important.
Several methods have been used to detect CALR exon 9 mutations including direct DNA sequencing, PCR followed by fragment analysis and immunostaining.18,19,26,27
Although fragment analysis has a relatively high sensitivity for CALR mutations detection, it cannot discriminate point mutation from wild-type sequence. High-resolution melting analysis (HRMA) is a closed-tube and PCR-based technique for the detection of gene polymorphism and mutations by measuring changes in the melting of a DNA duplex.68 HRMA is a well-established method for the detection of or prescreening for mutations both in a routine molecular laboratory and in a research setting. For example, HRMA has shown high sensitivity and specificity for the detection of JAK2 V617F and JAK2 exon 12 mutations in patients with MPN.69-71 Recently, the feasibility of using HRMA for the detection of CALR mutations in ET and persistent thrombocytosis has been reported using the LightCycler 480 platform (Roche Diagnostics).28 In this study, we sought to assess HRMA for rapid and sensitive detection of CALR exon 9 mutations in ET using the CFX Connect real-time system (Bio-Rad Laboratories, Hercules, CA, USA).
3. Patients and Methods
3.1 Patient samples and DNA extraction
The screening for mutations in patients with hematologic neoplasms was approved by the Institutional Review Board of Mackay Memorial Hospital. 32 adult patients with JAK2/MPL-unmutated ET were retrospectively enrolled based on the 2008 World Health Organization classification and 8 healthy adults were used as wild-type control. Written informed consent was obtained from all patients. Patient genomic DNA was derived from bone marrow or peripheral blood by using EasyPure Genomic DNA Spin Kit (Bioman, Taipei, Taiwan).
3.2 Assay design and the HRMA technique
Oligonucleotide primers were designed by Primer3 software to flank all CALR exon 9 variants reported in MPN. The primers were used to amplify a 134 bp amplicon [GenBank:
NM_004343]: forward 5’ - GAAACAAATGAAGGACAAACAGG -3’, and reverse 5’ - CCTCATCCTCCTCATCCTCA -3’. PCR was performed in a 20 μl reaction volume containing precision melt supermix (Bio-Rad Laboratories, Hercules, CA, USA), 100 nM of each primer, and 25 ng genomic DNA. The 134 bp amplicon was run according to the following conditions: an initial denaturation step of 95˚C for 2 min, followed by 35 cycles of 95˚C for 10 sec, 58˚C for 30 sec, and 72˚C for 30 sec. After completion of amplification, DNA was heated at 95˚C for 30 sec, kept at 60˚C for 1 min, and then melted from 70 to 95˚C (increment 0.2˚C, dwell time 10 sec). The results were analyzed using the Bio-Rad Precision Melt Analysis software. Melting profiles were normalized, grouped and displayed as fluorescence-versus-temperature plots or subtractive difference plots (-df/dt vs T). All samples with distinguished melting curves from wild-type were confirmed by duplicate study. Both type 1 and type 2 CALR exon 9 mutant cDNA were obtained by direct DNA synthesis, and CALR wild-type cDNA was cloned from patient sample.
3.3 Sanger sequencing
All patients were also independently screened for CALR exon 9 mutations spanning codons 352–417 by Sanger sequencing on an ABI 3730 sequencer based on previously described method.19 All identified sequence variants were subjected to repeated bi-directional sequencing for confirmation. Mutations were identified using DNA Dynamo sequence analysis software (Blue Tractor Software Ltd, Conwy, UK). All patients had been screened for JAK2V617F and MPL exon 10 mutations as previously described.7,9
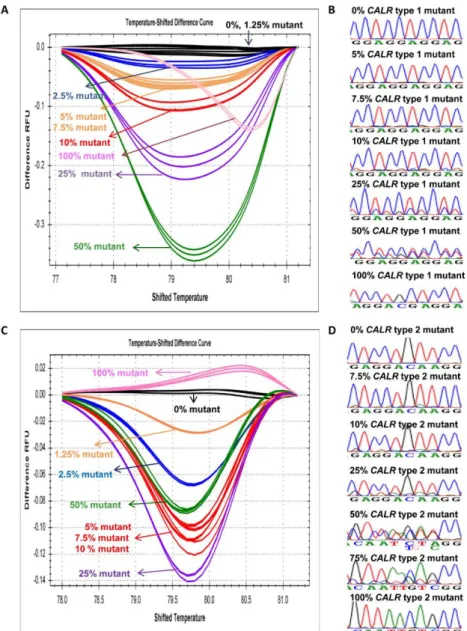
3.4 Sensitivity of HRMA in detecting CALR type 1 and type 2 mutations
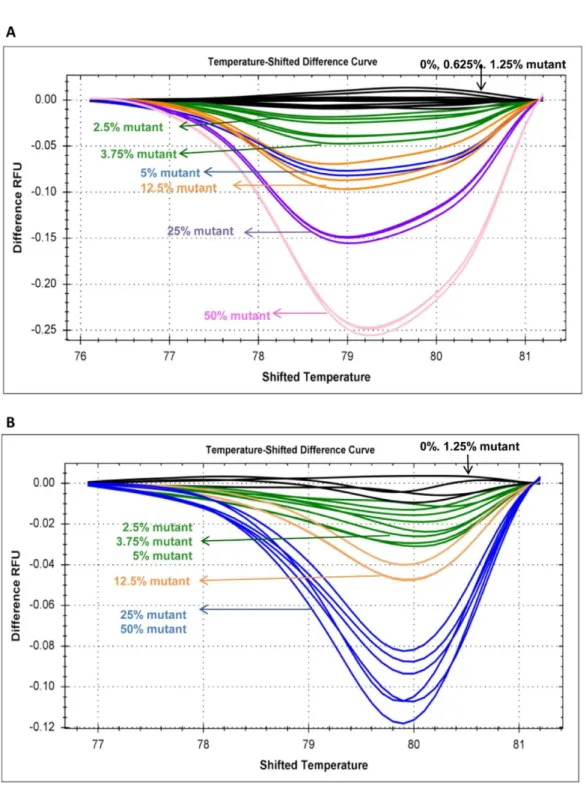
To study the sensitivity of the methodology, we serially diluted two plasmids carrying CALR type 1 and type 2 mutations with wild-type plasmid DNA in different concentrations (100% mutant, 50% mutant, 25% mutant, 10% mutant, 7.5% mutant, 5% mutant, 2.5%
mutant, 1.25% mutant, and 0% mutant). The sensitivity tests were carried out in triplicate samples. The sensitivity of HRMA was validated by serially diluting two patient samples carrying CALR type 1 and type 2 mutations with control DNA. Based on the relative peak areas of the mutant and wild-type PCR products, the mutant allele burden of these 2 patient samples was estimated to be ~50%. CALR type 1 and type 2 patients’ DNA were also serially diluted by wild-type DNA in different concentrations (50% mutant, 25% mutant, 12.5% mutant, 5% mutant, 3.75% mutant, 2.5% mutant, 1.25% mutant, 0.625% mutant, and 0% mutant). We did not evaluate the sensitivity of HRMA for other types of CALR mutations because they are less frequently detected.
3.5 TA-cloning
The PCR products of CALR exon 9 of 6 ET patients suspected to have a low allele burden mutant were purified using a EasyPure High Pure PCR clean-up Kit (Bioman, Taipei,
Taiwan) and cloned into a pGEMT-easy vector (Promega, Madison, CA, USA). We obtained at least 16 clones in each individual. The PCR product of each clone was checked on a 2% agarose gel by electrophoresis for the presence of mutant band. All selected clones were then sent for Sanger sequencing regardless the presence or absence of mutant band.
4. Results
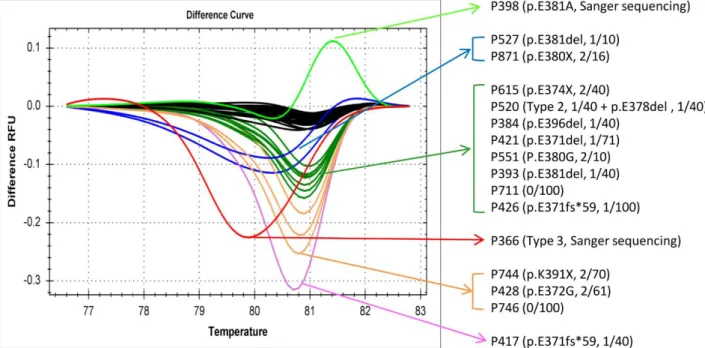
4.1 Sensitivity of HRMA in identifying the CALR type 1 and type 2 mutants
We first evaluated the sensitivity of HRMA in detecting the CALR type 1 and type 2 mutant plasmid DNA with different concentrations of mutant DNA serially diluted by wild-type plasmid DNA. HRMA could distinguish CALR type 1 and type 2 mutants with the maximal sensitivity of 2.5% and 1.25%, respectively (Figure 2A and C). Whereas, the maximal sensitivity of Sanger sequencing for the detection of both CALR type 1 and type 2 mutants was at least 10% or higher (Figure 2B and D). Besides, the maximal sensitivity of HRMA was validated with 2 patient samples and was found to be 2.5% for both CALR type 1 and type 2 mutants (Figure 3A and B).
4.2 Detection of CALR exon 9 mutations in JAK2/MPL-unmutated ET patients
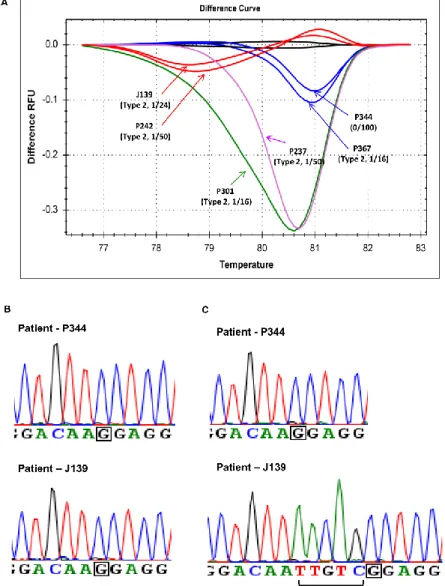
In this cohort of 32 ET patients, the normalized melting curves of 22 (68.8%) patient samples clearly showed a distinctive difference from that of wild-type group, and the representative normalized melting curves from 6 of the 22 patient samples were shown in Figure 4A. When the data were represented in difference plots, the individual nature of the mutant melting curves became more apparent as illustrated in Figure 4B. To determine the concordance between HRMA and Sanger sequencing, all 32 ET patients were also screening for CALR exon 9 mutations by Sanger sequencing. In the 22 ET patients with distinctive melting curves, Sanger sequencing could only detect CALR exon 9 mutations in
16 patients. All the remaining 6 patients with distinctive melting curves as shown on Figure 5A had wild-type CALR exon 9 sequences by Sanger sequencing. The other 10 patients were determined to have wild-type CALR by both HRMA and Sanger sequencing.
In the 6 patients with discordant results between HRMA and Sanger sequencing, we then performed TA-cloning to determine whether these 6 patients had low allele burden CALR mutations not detected by Sanger sequencing (Figure 5B and C).
After TA-cloning, CALR type 2 mutations were detected in 5 of 6 patients from 1 (6%) of 16 clones to 1 (2%) of 50 clones, and only 1 clone from each patient was tested positive for the CALR mutation (Figure 5). We did not identify CALR mutation in the last patient after screening for 100 clones. Therefore, HRMA identified CALR mutations in 21 (65.6%) of 32 ET patients compared to 16 (50%) by Sanger sequencing. The possible 3% false positive rate is low and no false negative was detected in our HRMA system.
In this study, 21 JAK2/MPL-unmutated ET patients were found to harbor 6 types of CALR exon 9 mutations: 5 type 1 (p.L367fs*46), 11 type 2 (p.K385fs*47), 1 type 3 (p.L367fs*48), 2 type 34 (p.K385fs*47), and 2 other types (p.L367fs*43 and p.E369fs*50).
All CALR exon 9 mutations are indels causing +1 base-pair reading frameshift, with type 2 (11/21, 52.4%) being the most prevalent mutational type. In these 21 patients with CALR mutations, the number of female patients was slightly higher than male patients (57% vs 43%) (Table 1).
5. Discussion
The identification of CALR mutations is important in the molecular diagnosis of MPN especially in JAK2/MPL-unmutated patients. In addition, CALR mutational status was
found to be one of the most significant risk factor for survival in PMF.33 Sanger sequencing has been used to detect CALR exon 9 mutations in many studies, but it is rarely sensitive below a 10% mutant allele burden as illustrated in Figure 2B and D. Fragment analysis assay was also used and the sensitivity was estimated to be 5% or less for CALR exon 9 mutations.27 Although fragment analysis assay is able to detect most indel mutations in CALR, it cannot discriminate point mutation from wild-type sequence.
Recently, Bilbao-Sieyro et al. showed that HRMA is a feasible method for the detection of CALR mutations using the LightCycler 480 platform.28 The amplicon size of their primer sequences was 265 bp, and the limit of detection for CALR type 2 (K385fs*47) mutant was of 3%. However, the ideal amplicon length for HRMA is usually less than 250 bp. In this study, the HRMA primers with an amplicon size of 134 bp were designed and are capable of detecting common CALR exon 9 mutations in myeloid neoplasms with satisfactory sensitivity.
Based on the dilution studies using patients’ genomic DNA, the maximal sensitivity of our HRMA using CFX Connect real-time system for both CALR type 1 (L367fs*46) and type 2 (K385fs*47) mutants was of 2.5%. In addition to 16 CALR mutated samples that could be detected by both HRMA and Sanger sequencing, we were able to identify another 5 patients with low CALR mutant allele burden only by HRMA. In this situation, we used TA-cloning followed by Sanger sequencing to confirm the mutation suspected.
Alternatively, fragmented analysis may be used for mutation detection because it also has a better sensitivity than Sanger sequencing. We were not able to detect CALR mutation in 1 of the 6 patients after screening for 100 clones. It is likely that this patient might still have low allele burden CALR mutation which, by chance was missed by random selection of clones (Figure 5A). However, we counted the result as a possible 3% false positive rate to
avoid overestimation of our data. Importantly, no false negative was found in our HRMA system and this is critical in regard to its role as a screening tool.
HRMA developed in this study can be utilized for rapid, sensitive and reliable detection of CALR mutations. Although a total of 5 SNPs (rs201971744, rs143880510, rs370029737, rs374121178 and rs150264068) are reported in the region covered by our amplicon, the minor allele frequency of 3 of them is reported to be less than 0.01%. Therefore, the influence of these 5 SNPs to our HRMA system will likely be very small. Nevertheless, one limitation to this HRMA methodology is that it will not be able to identify the 2 CALR exon 9 point mutations reported in follicular lymphoma (E403X and E405Q) because they are not covered by our 134 bp amplicon. The frequency and significance of these 2 CALR point mutations in follicular lymphoma are currently not yet clear. Therefore, our HRMA methodology is suitable for use in patients suspected to have myeloid neoplasms especially ET and PMF. By using HRMA, we detected a total of 6 different types of CALR mutations in ET patients. All the CALR mutations detected in this study resulted in a +1 base-pair shifting in the reading frame and generated the characteristic novel peptide sequence in the C-terminus. All the CALR exon 9 indel mutations likely contribute to a similar, yet not clearly understood molecular pathogenesis in ET and PMF. In addition, the number of female patients was slightly higher than male patients (57% vs 43%) in our 21 ET patients with CALR mutations, and this has also been observed in other study.72
HRMA, a close-tube method, is not only rapid as it is conducted immediately after PCR amplification, but is also cost effective because it can reduce the use of Sanger sequencing.
By using HRMA, a medium-throughput screening for CALR mutations is also possible.
Based on these advantages, our results clearly illustrated that HRMA is a more suitable and
sensitive method over Sanger sequencing for the screening of CALR mutations in both clinical and research settings. Nevertheless, in samples with distinct melting curves, complimentary Sanger sequencing is still required to determine their exact genotypes because the pattern of melting curves does not correlate with specific CALR mutational types.
In conclusion, we have shown that HRMA is a rapid, sensitive, reliable and cost effective method for the detection of CALR mutations. Because CALR mutations have important diagnostic and prognostic significance in ET and PMF, HRMA can be a useful screening method for the identification of common CALR mutations.
Chapter 3
The clinical and prognostic significance of CALR mutations and JAK2/CALR co-mutations in Taiwanese ET patients
1. Summary
Frequent CALR mutations have been discovered in patients with JAK2/MPL-unmutated essential thrombocythemia (ET) and primary myelofibrosis. We sought to screen for CALR exon 9 alterations with high-resolution melting analysis (HRMA) in 92 adult ET patients, and to determine the clinical and molecular correlates. In this cohort, 59 (64%) patients harbored JAK2 V617F mutation and one (1%) harbored MPL W515K mutation. By HRMA followed by TA-cloning, we identified classic CALR indel mutations in 21 (22.8%) patients. Eleven (12%) patients were triple-negative. The 59 JAK2-mutated patients were also screened for CALR exon 9 alterations by HRMA, and 16 (27.1%) samples were found to have distinct melting curves from wild-type. In 2 of these 16 samples, one CALR type 3 mutation and one single nucleotide polymorphism (rs143880510) were detected by Sanger sequencing. Although the remaining 14 patients were wild-type by Sanger sequencing, CALR alterations were detected in 12 (85.7%) patients after TA-cloning: 3 harbored classic CALR indel mutations, 5 (8.5%) harbored 4 types of 3 bp inframe deletions, and 5 (8.5%) harbored 5 types of point mutations. Overall, various CALR exon 9 alterations were detected in 13 (22%) of 59 JAK2-mutated ET patients. In comparable to previous reports, CALR mutations were associated with younger age (p=0.025), higher platelet count (p<0.001) and lower hemoglobin level (p=0.016). JAK2-mutated ET patients with concomitant CALR alterations were associated with oldest age (p=0.025), higher thrombotic events after diagnosis (p=0.048), higher major arterial thrombotic events after diagnosis (p=0.022) and more patients being high risk group for thrombo-hemorrhagic
complications (p=0.023). Frequent CALR exon 9 alterations in JAK2-mutated ET patients define a specific subgroup of patients with increased risk of thrombotic events.
2. Introduction
Essential thrombocythemia (ET) is a clonal hematopoietic stem cell neoplasm and one of the classic BCL-ABL1-negative chronic myeloproliferative neoplasm (MPN), which also includes polycythemia vera (PV) and primary myelofibrosis (PMF).2 JAK2 V617F mutation can be detected in more than 95% PV patients, and 50% to 60% of ET and PMF patients. MPL mutations at codon 515 are found in 3% to 5% of JAK2-unmutated ET and PMF patients. The 2008 World Health Organization (WHO) classification has incorporated JAK2 V617F and MPL mutations into the diagnostic criteria of MPN. Both JAK2 V617F and MPL mutations cause activation of the Janus kinase/signal transducer and activator of transcription (STAT) signaling pathway leading to the development of JAK inhibitor therapy in MPN.
Recently, two seminal studies discovered a high frequency of somatic calreticulin (CALR) mutations in patients with JAK2/MPL-unmutated ET and PMF.18,19 The pattern of most CALR mutations in MPN is heterozygous indels in exon 9 causing one base pair (bp) reading frameshift. CALR mutations have been shown to have important diagnostic and prognostic significance in ET and PMF patients,18,19,29 and will likely be incorporated into the WHO diagnostic criteria for MPN. In vitro studies on the molecular pathogenesis of CALR mutations in MPN have shown controversial results in regard to the involvement and/or activation of the JAK/STAT signaling pathway,18,19,38 and the exact pathogenesis of
Several techniques such as Sanger sequencing and polymerase chain reaction (PCR) followed by fragment analysis have been used to detect CALR mutations.10,18,19,27
High-resolution melting analysis (HRMA) is a well-established method for the screening of mutations, and we have developed a rapid and sensitive HRMA for the detection of CALR exon 9 mutations.73 In this study, we sought to screen a cohort of 92 Taiwanese ET patients for CALR exon 9 mutations with HRMA and Sanger sequencing independently, and to determine the clinical and molecular correlates.
3. Patients and Methods 3.1 Patients
The institutional review board of Mackay Memorial Hospital has approved the screening for mutations. All patients provided written informed consent. Diagnosis of ET was established based on the 2008 WHO criteria. The clinical and laboratory characteristics at the time of diagnosis or referral were collected. Genomic DNA derived from bone marrow, peripheral blood, and peripheral blood granulocytes and/or mononuclear cells were used for mutation screening.
3.2 Screening for CALR mutations
CALR mutations were screened by Sanger sequencing on an ABI 3730 sequencer as preciously described.19 CALR exon 9 mutations were independently screened by HRMA using a CFX96 real-time PCR detection system (Bio-Rad Laboratories, Hercules, CA) as previously described with a maximal sensitivity of 2.5% for both CALR type 1 and type 2 mutants.73 Briefly, a pair of oligonucleotide primers were used to amplify a 134 bp amplicon [GenBank: NM_004343] which flanked all CALR exon 9 variants reported in
MPN. All samples with distinguished melting curves from wild-type were confirmed by duplicate studies. Peripheral blood samples from 78 healthy adults were also used to validate the specificity of our HRMA. JAK2 V617F mutation was screened by allele-specific PCR with an analytic sensitivity of 5% and MPL exon 10 mutation by Sanger sequencing as previously described.7,9
3.3 TA-cloning
TA-cloning was performed by pGEM-T easy vector system (Promega, Madison, CA, USA) as previously described.73 At least 10 clones in each individual were randomly selected for the screening of CALR exon 9 alterations by Sanger sequencing. All novel single nucleotide variant that was only detected once was treated as artifact and excluded.
3.4 Statistical analysis
The correlation between clinical characteristics and mutational status was calculated by the chi-square test or Fisher’s exact test. The comparison between continuous and categorical variables was performed by the Mann-Whitney U test or Kruskal-Wallis H test. SPSS Statistics software (IBM, New York, USA) was used for all calculations. P values <0.05 were considered significant.
4. Results
4.1 CALR exon 9 mutations
Among the 92 ET patients (median age 53 years; 58% females), 59 (64%) patients harbored JAK2 V617F mutation and one (1%) patient harbored MPL W515K mutation. 32 JAK2/MPL-unmutated ET patients were utilized for the development of our HRMA
wild-type. In 16 of these 22 samples, Sanger sequencing confirmed the presence of 6 types of CALR mutations: 5 type 1 (p.L367fs*46), 6 type 2 (p.K385fs*47), 1 type 3 (p.L367fs*48), 2 type 34 (p.K385fs*47), and 2 other types (p.L367fs*43 and p.E369fs*50).
The other 6 samples were wild-type by sequencing, and CALR type 2 mutations were detected in 5 of 6 patients after TA-cloning indicating the presence of low allele burden CALR mutants in them. By using our HRMA platform, we identified CALR mutations in 21 (22.8% overall and 65.6% in JAK2/MPL-unmutated) ET patients and this frequency is comparable to other studies.18,19,29 11 (12%) ET patients were negative for JAK2, CALR and MPL mutations. In the 78 healthy adults, 2 samples were found by HRMA to have distinct melting curves from wild-type. One single nucleotide polymorphism (SNP, rs143880510) and one wild-type were found after Sanger sequencing in these 2 samples.
Therefore, the false positive rate of our HRMA system was 1.3%.
4.2 CALR exon 9 alterations
After screening the 59 JAK2 V617F-mutated ET patients for CALR alterations by HRMA, 16 (27.1%) samples were found to have distinct melting curves from wild-type (Figure 6).
In 2 of these 16 samples, one CALR type 3 mutation (p.L367fs*48) and one SNP (rs143880510) were detected by Sanger sequencing. All the other 14 samples were wild-type by sequencing. Surprisingly, we detected a high frequency of CALR exon 9 alterations in 12 (85.7%) of these 14 patients after TA-cloning (Table 2). Three patients harbored the classic CALR indel mutations: one each of type 2 p.K385fs*47, p.E370fs*60 and p.E371fs*59. Hence, 4 (6.8%) ET patients had classic CALR indel and JAK2 V617F co-mutations in this cohort. Five patients (8.5%) including the aforementioned patient (P520) with type 2 CALR mutation harbored 4 types of 3 bp inframe deletions all resulted in the deletion of a single amino acid of glutamic acid: two p.E381del, and one each of