國立臺灣大學生命科學院生化科學研究所 碩士論文
Graduate Institute of Biochemical Sciences College of Life Science
National Taiwan University Master Thesis
岩藻糖基化之膜蛋白與癌症轉移之間關係
The relationship between fucosylated membrane
proteins and cancer metastasis
白昕永 Hsin-Yung Pai
指導教授﹕陳水田 博士 Advisor:Shui-Tein Chen, Ph.D.
中華民國 101 年 7 月 July, 2012
ii
中文摘要
根據世界衛生組織統計,在人類眾多癌症中,十大死因排名第三名的疾病。在 臨床檢體的分析中,惡性的肺癌細胞周邊組織及血清會表現較大量的第四型岩藻 糖轉酶,其功能為將岩藻糖以 α-1,3/4 的方式接上 N-乙醯葡萄醣胺
(N-acetylglucosamine)為核心的 N 端連結糖蛋白。岩藻糖基化是屬於轉錄後修飾 中糖基化的一部分,由於岩藻糖基化的增加,導致細胞表面特定糖結構大量表現,
會影響細胞內外的訊息傳遞、細胞貼附能力以及細胞與細胞間的辨識功能。在先 前的研究中,利用已建立穩定表現第四型岩藻糖轉酶的肺腺癌細胞株(A549-FutIV) 與轉入空質體的肺腺癌細胞株(A549-Mock)比較,發現當中有十九種 N 端連結的膜 蛋白上,有較多或較少的鹽藻糖基化現象。在這十九種 N 端連結糖蛋白中,大部 份蛋白的功能並未明確被研究出。
在本篇研究裡,我們想要去探討關於這些岩藻糖基化改變的膜蛋白與癌症的惡 性轉移之間的關係,利用小干擾核醣核酸將特定基因的信使核醣核酸分解,達到 蛋白質表現下降,來探討 SLC3A2, CD166 以及 CD44 對於癌症轉移的影響。由於 CD166 與 CD44 的表現量對於癌症惡性程度是有相關性,當 CD166,CD44 基因敲除後,對 於癌症轉移能力有顯著性的減少,CD166 本身與細胞外基質的結合能力透過實驗得 知其具有貼附能力。SLC3A2 本身為氨基酸轉運子重鏈,在纖維連接蛋白的聚集上 扮演調控的角色,同時,在腎細胞癌中扮演著生物標記。由實驗結果得知,SLC3A2 對於癌症的轉移能力並無顯著性的差異,但確實對於纖維連接蛋白擁有貼附能力。
CD44 基因敲除後,雖然會抑制惡性癌症的轉移,但並非因細胞外基質的貼附能力 所致。結合侵犯實驗及貼附實驗發現,SLC3A2 的貼附能力對於易轉移之癌細胞有 抑制功效,CD166 及 CD44 所造成的抑制癌症轉移效果並非經由貼附能力所致。
關鍵字:
肺癌/第四型岩藻糖轉酶/癌症轉移/膜蛋白/轉錄後修飾/N 端連結糖基化/細胞侵 犯實驗
iii
Abstract
Lung cancer is a disease and ranked as third among the ten leading causes of human deaths according to statistics reports of the world health organization. In clinical studies, lung cancer of a more malignant state may express more fucosyltransferase IV(Fut IV), which is an enzyme that transfers a fucose to a core N-acetylglucosamine sugar of N-linked glycoprotein via α-1,3/4 linkage. Fucosylation is a common glycosylation process in posttranscriptional modification that can affect signal transduction, cell adhesion and cell-cell recognition. In previous studies, the A549-FutIV cell line was established, which expresses relatively high amounts of Fut IV.
A549-FutIV, consists of nineteen membrane proteins with more or less higher degree of fucosylation of their N-glycoproteins when compared with A549-Mock. The functions of most of these membrane proteins remain unknown.
In this study, we focus on the relationship between these membrane proteins and cancer metastasis in A549-FutIV. The protein (SLC3A2, CD166 or CD44) was knockdown by siRNA. CD166 and CD44 expressions correlated with the degree of cancer malignancy. After CD166 and CD44 knockdown, the cancer invasion ability had significantly decreased compared to the control and CD166 could also bind to the extracellular matrix(ECM). SLC3A2 was an amino acid transporter heavy chain that played an important roles as a mediator of fibronectin assembling and also as a
iv
biomarker in renal cell cancer. According to our experiments, SLC3A2 did not cause any significantly changed to cancer metastasis, but it could bind to fibronectin.
Although CD44 knockdown could inhibit cancer invasion, it did not result from its adhesion ability. By combining invasion assay and adhesion assay results, we found that SLC3A2 knockdown could inhibit cancer invasion through its adhesion ability.
However, the inhibition of cancer invasion after CD166 and CD44 knockdown did not occur via their adhesion abilities.
Key words:
Lung cancer / Fucosyltransferase VI / Metastasis / membrane protein / PTM / N-linked glycosylation / invasion assay
v
Content
Acknowledgement ... i
Abstract (Chinese) ... ii
Abstract (English) ... iii
Abbreviations ... 1
1. Introduction ... 3
1.1 Lung cancer ... 3
1.2 Membrane protein ... 4
1.3 Posttranscriptional modification ... 6
1.3.1 N-linked Glycoprotein ... 7
1.3.2 O-linked Glycoprotein ... 8
1.3.3 Fucosylation ... 9
1.4 Metastasis ... 10
1.5 Nineteen membrane proteins in study ... 12
1.6 Target genes: SLC3A2, CD166, CD44 ... 16
1.6.1 SLC3A2 ... 16
1.6.2 CD166 ... 16
1.6.3 CD44 ... 17
vi
2. Materials and Methods ... 18
2.1 Cell line and cell culture ... 18
2.2 siRNA knockdown ... 18
2.3 RNA preparation, reverse transcription polymerase chain reaction (RT-PCR) and quantitative PCR (qPCR) ... 19
2.3.1 RNA preparation ... 19
2.3.2 Reverse transcription polymerase chain reaction (RT-PCR) ... 20
2.3.3 Quantitative PCR (qPCR) ... 21
2.4 Migration/Invasion assay ... 21
2.4.1 Migration assay ... 21
2.4.2 Invasion assay ... 22
2.5 Adhesion assay ... 22
2.6 Protein extraction ... 23
2.7 Western blot ... 24
3. Results ... 26
3.1 Characteristics of A549-Mock and A549-FutIV ... 26
3.2 Functional network analysis of the nineteen genes ... 26
3.3 Analysis of the siRNA knockdown effect in A549-Mock and A549-FutIV 27 3.4 Functional analysis of the target genes after the genes knockdown ... 27
vii
3.4.1 The effect of target proteins invasion ability ... 27
3.4.2 The effect of target proteins adhesion ability ... 28
3.4.3 The effect of target proteins migration ability ... 28
4. Discussion ... 30
4.1 Fucosyltransferase IV over-expression in A549 might make cancer more malignant ... 30
4.2 The relationship between membrane proteins and cancer metastasis ... 30
4.2.1 SLC3A2 ... 30
4.2.2 CD166 ... 31
4.2.3 CD44 ... 32
4.3 Characteristics of the three proteins regarding cell adhesion to ECMs .... 32
4.4 The protein functions as related to invasion and adhesion ability ... 33
5. Reference ... 34
6. Figures ... 46
viii
Lists of Figures
Figure 1. A549-mock and A549-FutIV cell shapes ... 45
Figure 2. Fucosyltransferase IV mRNA level ... 46
Figure 3. Protein expression of fucosyltransferase IV... 46
Figure 4. AAL blot between A549-Mock and A549-FutIV ... 46
Figure 5. The invasion ability between A549-Mock and A549-FutIV... 47
Figure 6. 12 membrane proteins network ... 48
Figure 7. Protein function classification ... 49
Figure 8. mRNA expression of the target genes ... .50
Figure 9. During the experiments, cell viability after siRNA transfection. ... 51
Figure 10. The protein level of these genes had turnover after 72h ... 52
Figure 10.1. SLC3A2 expression after siRNA knockdown ... 53
Figure 10.2. CD166 expression after siRNA knockdown ... 54
Figure 10.3. CD44 expression after siRNA knockdown ... 55
Figure 11. The invasion ability after the target knockdown ... 56
Figure 12. The percentage of transfected-cell invasion ability ... 57
Figure 13. The percentage of transfected-cell invasion ability ... 59
Figure 14. The adhesion ability of each siRNA knockdown cell lines ... 61
Figure 15. The migration ability of each target proteins after knockdown ... 62
ix
Lists of Tables
Table 1. 19 membrane proteins identified from A549-mock and A549-FutIV cell
lines ... 14
Tab le 2. The relative folds of invasion ability ... 58
Tab le 3. The relative folds of invasion ability ... 60
1
BSA CTCs DC ECM FUT GalNAc GPI HRP HA IgG IMPs MT1-MMP MMP NSCLC O-GalNAc O-GlcNAc PBST PVDF
Abbreviations
Bovine serum albumin Circulating tumor cells Dendritic cell
Extracellular matrix Fucosyltransferase Galactosamine
Glycosylphophatidylinositol Horseradish peroxidase Hyaluronic acid
Immunoglobin G
Integral membrane proteins
Membrane-type 1 matrix metalloproteinase Metalloproteinase
Non-small-cell lung carcinoma O-N-acetylgalactosamine O-N-acetylglucosamine
Phosphate buffer saline tween-20 Polyvinylidene fluoride
2
PTM qPCR RT-PCR SCLC siRNA
Posttranscriptional modification Quantitative PCR
Reverse transcription polymerase chain reaction Small-cell lung carcinoma
Small interfering ribonucleic acid
3
1. Introduction
1.1 Lung cancer
Lung cancer is one of the most common and deadliest forms of cancer around the world. According to estimations of the International Agency for Research on Cancer (IARC) and cancer statistics from the National Cancer Institute (NCI), lung cancer
patients composed approximately 1.61 million new cancer cases in 2008, representing
12.7% of all new cancers (Ferlay et al, 2010). In this year, it was also found to be the
most common cause of death from cancer, with 1.38 million deaths (18.2% of the total),
ranking as the third most common cause of human deaths in high income countries by the statistical reports of the World Health Organization (WHO) (WHO:http://gamapserver.who.int/gho/interactive_charts/mbd/cod_2008/graph.html).
Although long-term exposure to tobacco smoke is the most common cause of lung cancer, there are still other factors, such as genetic predispositions and exposure to asbestos or air-pollution, including secondhand smoke and lampblack, can affect cancer formation.
There are two main types of lung cancers based on histological type, namely small-cell lung carcinoma (SCLC) and non-small-cell lung carcinoma (NSCLC).
Small-cell lung carcinoma (SCLC) displays high aggression and usually occurs in smokers. It grows rapidly, and about 60% of patients show widespread metastasis. The
4
non-small-cell lung carcinoma (NSCLC) can be categorized into three sub-types:
squamous cell lung carcinoma, adenocarcinoma, and large-cell lung carcinoma. There are about 30% of non-small-cell lung cancer patients that classify as having squamous cell lung carcinoma (or epidermoid carcinoma), a cancer that looks like fish scales under a microscope. This type of lung cancer usually occurs in the central bronchi and often grows more slowly compared with other cancer because of its well-differentiation.
Adenocarcinoma is observed in the cases of NSCLC involving non-smokers, although a significant percentage of adenocarcinoma cases are still associated with smoking, Large cell carcinoma is often found in the central part of the lung and usually discovered at a later stage of lung cancer (Gorlova et al, 2007; Hong & Tsao, 2008; NCI, 2012) and may spread into lymph nodes and to distant organs.
1.2 Membrane protein
The biological membrane consists primarily of a lipid bilayer, that is not only separates the interior of cells from the extracellular environment but also allows division of biologic functions into separate organelles, such as the mitochondria, Golgi, lysosome, Endoplasmic Reticulum and so on (Lodish et al, 2000). A variety of proteins, called membranes, accomplish vital biologic functions on/in the membrane, as structural proteins, cell adhesion proteins, enzymes, receptors and transporters.
The membrane proteins can be divided into three categories based on how they
5
make contact with the membrane: (1) integral membrane protein, (2) peripheral membrane protein and (3) lipid-anchored protein. Integral membrane proteins (IMPs) are permanently attached to the membrane because they contain many hydrophobic
regions which allow them to pass through the membrane once or multiple times forming α-helices or multiple β-strands for membrane-spanning domains. They can be released
from the membrane using detergents, nonpolar solvents, or denaturing agents. There are many different functional proteins directly or indirectly related to IMPs. For example, Na+/K+ ion transporters maintain normal potential voltage in nerves or receptor tyrosine kinases that receive stimulation from the extracullular environment, are involved in self-phosphorylation or signal transduction. The most important IMPs for cell adhesion are of the cadherin family, and desmoglein family.
Peripheral membrane proteins bind to the membrane indirectly by interacting with integral proteins through hydrogen bonding to their hydrophilic portions or directly by interacting with membrane surfaces through electrostatic forces or polar head groups of membrane phospholipids. The protein kinase C, plays a role in signal transduction and also belongs to the peripheral membrane protein group. There are still other peripheral membrane proteins, located in the ECM.
The last category of membrane proteins includes lipid-anchored proteins which are located on the surface of the lipid bilayer. They are covalently bound to the lipids
6
via a prenyl group. There are many lipid-anchored proteins covalently linked to glycosylphosphatidylinositol (GPI) on the external plasma membrane, called GPI-anchored membrane proteins (Becker et al, 2004).
Although malignant tumors can be the result of mutations in a number of genes, the direct factors affecting tumor metastasis may be uncontrolled proliferation, cancer immortality and the expression of membrane proteins (Christofori & Semb, 1999).
Membrane proteins expression may increase cancer malignancy. In breast cancer, for example, thirteen membrane proteins showed abnormally high expression levels and three membrane proteins were under-expressed in metastatic cells, as compared to the non-metastatic cell line (Leth-Larsen et al, 2009). In cancer metastasis, loss of the epithelial adhesion molecules, E-cadherin , is thought to enable metastasis (Kowalski et al, 2003). When E-cadherin loss, the induction of multiple transcription factors, such as Twist, is necessary for E-cadherin loss–induced metastasis. This suggests that loss of E-cadherin may contribute to metastatic dissemination in tumor. Also relevant is that in different tumors, the role of E-cadherin may also be different (Onder et al, 2008).
1.3 Posttranscriptional modification
Posttranscriptional modification(PTM) is a mechanism that happens after gene transcription and mRNA translation that is important for proper protein folding, signal transduction, cell adhesion, molecule trafficking and so on. The entire human genome
7
has been sequenced, which is estimated to contain roughly 20,000 to 25,000 genes. It is estimated that there are over 1 million proteins that are derived from these genes.
Although this seems to be quite a lot, it is still not enough to support a complex organism, such as humans (Consortium, 2004). PTMs provide a substantial increase in molecular diversity for the different kinds of cells to interact with. There are many different kinds of PTM in the cell, including glycosylation, acetylation, alkylation, methylation, biotinylation, glutamylation, isoprenylation, phosphorylation and sulfation.
All of them are essential for cell functions (Beck-Sickinger & Mörl, 2006).
1.3.1 N-linked Glycoprotein
N-glycosylation is a modification in which a sugar molecule is transferred to the nitrogen atom of an asparagine in the polypeptide chain of a protein. Typically, there are two common sites for N-glycosylation: Asp-x-Ser and Asp-x-Thr, where X can be any amino acid residue except for proline. There is a universal pentasaccharide core common to all N-glycans, Man3GlcNAc2, and further modification processes allow classification into three main classes, a high-mannose type, a complex type, and a hybrid type, which all undergo process reaction in the Golgi complex. However, many functions of proteins are affected by N-linked glycoproteins. Take SLC26A3 for example, which is a Cl-/HCO3-
exchanger that plays a major role in Cl- absorption from the intestine. When SLC26A3 has been deglycosylated, the transporter activity
8
depresses compared to the wild-type and its mature N-linked glycoprotein also can protect SLC26A3 from trypsin digestion (Hayashi & Yamashita, 2012). The N-glycosylation of membrane proteins are also related to cancer malignancy (Lau &
Dennis, 2008). For example, in E-cadherin, it has been found that the more β1,6-branched expression on the N-glycans, the more malignant the tumor they reside in
(Pinho et al, 2009).
1.3.2 O-linked Glycoprotein
O-linked glycosylation of secreted and membrane bound proteins take place in the cis-Golgi compartment after N-glycosylation and protein folding. The O-glycans are commonly fixed to serine and threonine, and also rarely to hydroxylysine and hydroxyproline amino acid residues. This type of glycoylation plays an important role in protein trafficking and in cellular communication during development. There are many O-glycan forms, such as O-N-acetylgalactosamine (O-GalNAc), O-fucose, O-glucose, O-N-acetylglucosamine (O-GlcNAc) and O-mannose, which are also referred to as “mucin-type” glycan (Hanisch, 2001). O-GalNAc can be further categorized by core type, as determined by eight distinct cores, designated core1-core8 (Varki et al, 2009). Although they do not have a universal core like N-glycans, the structural complexity of the chains initiated by O-linked GalNAc exceeds that of N-linked chains by far. In cancer cells, the cell-surface levels of α2β1 are reduced,
9
which is a result of core3 O-glycans causing inhibition of the normal interaction between the β1 subunit and the α2 subunit. This impairs FAK-mediated signal transduction and cell migration. Consequently, core3 O-glycan may cause suppression of cancer metastasis (Tsuboi et al, 2012).
1.3.3 Fucosylation
Fucosylation is one of the most common modifications for glycoproteins and glycolipids. This modification involves transfer of a fucose residue to N-glycans, O-glycans and glycolipids. In the fucosyltransferase family, there are nine fucosyltransferases, which are distributed into three main groups, depending on their type of fucosylation linkage: (1) α1,2, (2) α1,3/4 and (3) α1,6 fucosyltransferases. FUTI and FUTII are the specific members belonging to the α1,2-linked fucosyltransferase group and which catalyze the transfer of a fucose residue to the terminal Gal residue.
They are important for the ABO blood group system. There are six types of α 1,3/4-fucosyltransferases, including FUTIII, FUTIV, FUTV, FUTVI, FUTVII and FUTIX (Javaud et al, 2003). All of them except for FUTIII produce Lex, Ley and sLex epitopes which are essential for cell migration and adhesion. They attach a fucose to GlcNAc which belong to Gal-GlcNAc structures. It plays an important role in cancer metastasis because sLex can bind to its ligand, E-selectin, which expresses in endothelial cells, leukocytes and platelets (Borsig et al, 2002). When tumors invade the
10
blood vessels, they can persist in circulation by forming circulating tumor cells (CTCs), which interact with platelets through P-selectin and xLex. Moreover, the CTCs disseminate to appropriate sites to extravasate the blood vessels via interaction with endothelial cells that express E-selectins. Therefore, the influence of fucosylation on cancer metastasis depends on what epitopes forming (Homeister et al, 2001; Malý et al, 1996; Miyoshi et al, 2008).
1.4 Metastasis
Malignant tumors usually have some characteristics that are life-threatening, such as uncontrolled proliferation and immortality. Progression toward metastasis is one of the key factors that affects patient recovery after surgery or chemotherapy. It represents all of the end products of a multistep cell–biological process termed the invasion-metastasis cascade. In general, the invasion-metastasis cascade can be distributed into seven steps, (1) local invasion, (2) intravasation, (3) forming circulating tumor cell, (4) arrest at distant organ sites, (5) extravasation, (6) forming micrometastases, (7) metastatic colonization (Friedl & Wolf, 2003). Each steps is important for cancer metastasis.
During local invasion, the malignant cancer cells may secret some proteases, metalloprotease and/or cathepsin, which can digest the basement membranes and ECM surrounding epithelial cells and make cancer cells touching stroma cells, which secrete
11
many growth factors or stimulate tumor-associated macrophages activity. Therefore, they establish a potentially self-amplifying positive feedback loop. Most types of carcinoma can invade as cohesive multicellular units, jointly called a “collective invasion.” However, individual tumor cells may invade through two ways: (1) mesenchymal invasion or (2) amoeboid invasion (Friedl & Wolf, 2003). Cancer cells invade their neighbor cells via “epithelial-mesenchymal transition”, which decreases the E-cadherin expression and increase a set of pleiotropically acting transcription factors, including Snail, Twist and ZEB1, making them more stem-like (Thiery et al, 2009).
Even though there are many drugs that inhibit the protease activity, cancer cells also can invade, via amoeboid invasion, a mode of invasion that depends on diffusion of non-clustered integrins(Wang et al, 2004).
All the factors in the ECM can make cancer cells more malignant and may even lead to poor diagnosis. After cancer cells intravasate into blood vessels, they can form relatively large emboli via interaction with blood platelets through L-/P-selectins (Joyce
& Pollard, 2009). They shield themselves from shear forces and evade immune detection. When the CTCs disseminate to appropriate sites where they may make contact through ligand-receptor interactions or physical trapping, they can extravasate into the tissue. During the cancer cells extravasation, the primary tumors are capable of secreting some factors to perturb microvessel permeability, such as MMP-1, MMP-2,
12
and COX-2 to make them pass easily (Gupta et al, 2007; Padua et al, 2008). In order to form micrometastasis, the cancer cells change their gene expression and receive stimulation by some growth factors from stroma cells to maintain their life. They change their state via “mesenchymal-epithelial transition”. Finally, the primary cancer cells may now thrive at the secondary site and become more malignant.
1.5 The nineteen membrane proteins in the present study
Two different A549 cell lines, A549-mock and A549-Fut IV, were derived by Dr. Yu of the Department of Internal Medicine, National Taiwan University Hospital and National Taiwan University College of Medicine. The A549-FutIV cell line is transfected with gene fut4 for over-expression of FucT IV while A549-mock is transfected with empty plasmid as control. The only difference between these two cell lines is that the former can attach more fucose residues on the membrane proteins and affect cancer biology.
In a cite, comparative glycoproteomic approached were used to study the differences in the fucosylated membrane proteins when comparing the two A549 cell lines.
Nineteen membrane proteins were identified (Table1) that had high fucosylation. While the functions of some of these proteins remains unknown, for example, TM9SF3, TMEM206 and GOLM1, some of them seem to participate in cell adhesion, such as CAMD4, DSG2 and CNTN1. In normal cells, PLXNB2 is a cell surface receptor for
13
SEMA4C, SEMA4 and SEMA4G and plays an important role in cell-cell signaling.
Moreover, PLXNB2 affects neuron migration and is associated with RhoA activation (Conrotto et al, 2004; Perrot et al, 2002). EphA2 is a receptor tyrosine kinase which binds to ephrin-A family ligands on adjacent cells (Wykosky & Debinski, 2008).
Moreover, it can regulate cell migration by ephrin-A1/EFNA1 or promote cell adhesion by DSG1/desmoglein-1 (Lin et al, 2010; Ogawa et al, 2000; Wykosky & Debinski, 2008). It also may affect cancer metastasis through RhoA GTPase activation and make tumors become more malignant.
14
Gene entry name Protein name Sequence of identified
peptide
F/M
IMPAD1
Q9NX62
Inositol monophosphatase 3 QVALQTFGN259QTTIIPAG 1.683 GAGYK
SLC3A2
P08195
4F2 cell-surface antigen heavy chain
SLVTQYLN424ATGNR 6.89
CNTN1 Q12860 Isoform1 of Contactin-1 GTEWLVN457SSR 16.084 AN494STGTLVITDPTR 16.392 GKAN494STGTLVITDPTR 7.356 TM9SF3
Q9HD45
Transmembrane 9 superfamily member 3
IVDVN174LTSEGK 5.164
ALCAM Q13740 CD166 antigen N361ATVVWMK 10.118
ALGDCISEDSYPDGN167IT 11.844 WYR
TMEM20 6
Q9H813
Transmembrane protein206 IN155YTDPFSN162QTVK 2.186
GOLM1
Q8NBJ4
Isoform 2 of Golgi membrane protein 1
AVLVNN109ITTGER 6.27 Table 1. 19 membrane proteins identified from A549-mock and A549-FutIV
cell lines.
15
DSG2 Q14126 Desmoglein-2 YVQN462GTYTVK 11.169
PTPRJ
Q12913
Receptor-type
tyrosine-protein phosphatase eta precursor
VSDN391ESSSNYTYK 0.762
PRNP
P04156
Isoform 1 of Major prion protein
GEN197FTETDVK 2.685
EPHA2 P29317 Ephrin type-A receptor 2 TASVSIN435QTEPPK 5.79 GLG1
Q92896
Isoform 2 of Golgi apparatus protein 1
LN165LTTDPK 1.745
ITGA3
P26006
Isoform Alpha-3A of Integrin alpha-3
ELAVPDGYTN86R(T) 9.555
PLXNB2 O15031 Plexin B2 protein ALSN127ISLR 9999 LAMP1
P11279
LAMP1 protein variant (Fragment)
N62MTFDLPSDATVVLN76R 11.398 (S)
CD44 P16070 Isoform 12 of CD44 antigen AFN57STLPTMAQMEK N/A
ODZ3 Q9P273 Teneurin-3 IGPFAN2124TTK 4.213
CPD O75976 Carboxypeptidase D FANEYPN522ITR 9999 CADM4 Q8NFZ8 Cell adhesion molecule 4 QTLFFN67GTR 9999
16
1.6 Target genes: SLC3A2, ALCAM, CD44
1.6.1 SLC3A2
The 4F2 cell-surface antigen heavy chain is the protein encoded by the SLC3A2 gene. It is a multifunctional protein involved in cell transformation, integrin signaling, cell fusion and amino acid transport. 4F2hc is a type II glycoprotein, which forms heterodimers with different light subunits. When it associate with different light chains, it can transport different amino acids involved in gastric acid secretion (Drummond et al, 2010; Kirchhoff et al, 2006; Pfeiffer et al, 1999; Verrey et al, 2009). For example, SLC3A2 associates with SLC7A6 or SLC7A7, acting as an arginine/glutamine exchanger that follows an antiport mechanism for amino acid transport(Nel et al, 2012;
Verrey et al, 2009). Another function of the SLC3A2 is to make cell exert force on the matrix to interact with integrins to support downstream signals that lead to activation of RhoA small GTPase (Feral et al, 2007).
1.6.2 CD166
The CD166 antigen is a type I transmembrane glycoprotein, encoded by the ALCAM gene. It mediates both heterophilic (ALCAM-CD6) and homophilic
(ALCAM-ALCAM) cell-cell interactions. When it binds to CD6, recruited the antigen-induced dendritic cell (DC) to the T cell contact zone and sustains DC-induced T-cell proliferation after initial contact (Skonier et al, 1996b; Zimmerman et al, 2006a).
17
Although CD166 attracts lymphocytes through CD6 interaction, it also can help cell migration and make cancer cell more malignant. In colorectal carcinoma, ALCAM is frequently up-regulated and is a new independent prognostic marker (Weichert et al, 2004). Truncation of CD166 was found to diminish primary tumor growth and enhance melanoma metastasis (van Kempen et al, 2004). ALCAM in breast cancer is a tumor suppressor because lower ALCAM expression makes tumors more aggressive (Jezierska et al, 2006). It may have different effects in different tumors.
1.6.3 CD44
CD44 is a multi-structural and multi-functional cell surface molecule. The variety of CD44 isoforms can be divided into three groups: (1) CD44s the standard isoform, (2) CD44v which contains variable exons, and (3) CD44E which includes exons v8-10 (Goodison et al, 1999). CD44 is a receptor for hyaluronic acid (Aruffo et al, 1990). It also mediates cell-cell and cell-matrix interaction through its affinity for HA, collagens, osteopontin and matrix metalloproteinases (MMPs). CD44 may provide a platform for tumor invasion because CD44 forms hyaluronan-induced aggregates that make MMP9 work easily (Yu & Stamenkovic, 1999). In addition, CD44 can be cleaved by membrane-type 1 matrix metalloproteinase (MT1-MMP) and promotes cell migration (Kajita et al, 2001).
18
2. Materials and Methods
2.1 Cell lines and cell culture
The human lung adenocarcinoma cell lines, A549-Mock and fucosyltranferase IV-transfected A549 (A549-Fut4) were obtained from the Department of Internal Medicine, College of Medicine, National Taiwan University. Both of these cell lines were grown in RPMI1640 (Hyclone, Thermo Scientific,) supplemented with 10%(v/v) Fetal bovine serum (Biological industries), 100 unit/mL penicillin, 0.1 mg/mL streptomycin, 0.25 μg/mL amphotericin(Biological industries), 0.15 %(w/v) sodium bicarbonate (Sigma), 10mM HEPES (Sigma), 0.25% (w/v) glucose, and 1mM sodium pyruvate(Caissonlabs, North Logan, US). They were incubated in a humidified atmosphere with 5% CO2 at 37℃. The cells were subcultured every 2-3 times per week.
2.2 siRNA knockdown
The siRNA of target genes (SLC3A2, CD166, CD44) were ordered from Thermo Scientific Dharmacon. All siRNA of these genes were dissolved in RNase-free water to produce 20 μM as stock, then stored in a -80℃ refrigerator. Before the siRNA transfection, both cell lines (A549-Mock and A549-FutIV) were seeding in 24-well plate and 5*104/mL/well. The culture medium was replaced with antibiotic-free complete medium and incubated overnight at 37℃. Then, 50μL of the siRNA in opti-medium (invitrogen) was prepared by adding 2.5μL of 5μM siRNA to 47.5μL of
19
opti-medium in an 1.5mL eppendorf and gently mixed by pipetting carefully up and down, then incubated for 5 minutes at room temperature. This preparation was labeled as tube 1.
DharmaFECT transfection reagent in opti-medium was prepared by adding 1μL DharmaFECT reagent and 49μL opti-medium gently mixed and incubated for 5 minutes at room temperature, then labeled as tube 2. The content of tube 1 was then added to tube 2, mixed by pipetting carefully up and down and incubated for 20 minutes at room temperature. Then, 400 μL of antibiotic-free complete medium was added to get 500μL transfection medium with a final concentration of 25nM siRNA. This tranfection medium was prepared fresh before use. The antibiotic-free medium in the 24-well cultured plate was removed and 500 μL of fresh transfection medium added to each well.
The cell were incubated for 24 hours.
2.3 RNA preparation, reverse transcription polymerase chain reaction (RT-PCR) and
quantitative PCR(qPCR)
2.3.1 RNA preparation
The total RNA of A549-Mock and A549-Fut4 in the 24-well plate were extracted using a TRIzol Reagent (Invitrogen) following the protocol provided by the manufacturer. Briefly, the medium was removed, the cells were washed by PBS twice and then 1 mL TRIzol reagent was added to each well. The cell lysate was incubated 5
20
minutes to homogenize the RNA and then transferred to a new tube. Two hundred microliters of chloroform was added into 1 mL of homogenized mixture, which was shaken vigorously, and then centrifuged at 12000g for 15 minutes at 4℃ to separate the RNA into an aqueous phase, DNA into an internal phase and protein into an organic phase. For RNA precipitation, the aqueous phase was transferred into a new tube and the tube was inverted several times after adding isopropyl alcohol. Then, the aqueous-isopropyl alcohol mixture was centrifuged at 12000g for 10 minutes at 4℃, producing an RNA pellet at the bottom of tube. The supernatant was discarded and 75%
ethyl alcohol added twice to wash the pellet. Finally, the RNA pellet was dried out and dissolved in 50 μL RNase-free water..
2.3.2 Reverse transcription polymerase chain reaction (RT-PCR)
To obtain 1μg cDNA, the total RNA concentration obtained by the previous procedures was measured using NanoDrop(Thermo scientific) and appropriate volume of total RNA was used as template in a sterile PCR tube. One hundred pmol of oligodeoxythymidine primer (oligo(dT)18) was mixed with template RNA and filled with RNase-free water to 12.5 μL. The mixture would chill on ice after incubating at 65℃ for 5 minutes. Meanwhile, pre-mix was prepared, which contained 4μL of 5X reaction buffer, 0.5μL of RNaseOUT, 2μL of 10mM dNTP mix and 1μL of RevertAid reverse transcriptase (Fermentas). These two mixtures were mixed to a total volume of
21
20μL and the mixture incubated at 42℃ for 60 minutes. Finally, the reaction was
terminated by heating at 70℃ for 10 minutes and used in further experiments.
2.3.3 Quantitative PCR(qPCR)
The qPCR probe for these genes (Fut4, SLC3A2, CD166, CD44) were designed by LightCycler Probe Design Software (Roche) and the sequence of these genes were obtained from NCBI(accession number: NM_002033.3, AB018010.1, NM_001627.3, and NM_000610.3, respectively). The replicon of qPCR length was generally about 160-180 base pairs and the melting temperature was about 60℃. To avoid primer dimer, the probes used did not have ΔG less than -2000, a threshold value estimated by the software. The probes used in this study are shown in the following table.
2.4 Migration/Invasion assay
2.4.1 Wound healing assay
A549-mock and A549-FutIV cell lines were seeded at a density of 2 × 105cells primer name F primer 5'->3' R primer 5'->3'
Fut4 CCCAgACCgTgCCAACTA ggAggTgATgTggACAgC SLC3A2 CCAgAAggATgATgTCgCT CAACCTgAgTggAgAACC
ALCAM ggCAgTggAAgCgTCATA AgCAgAgACATTCAAggAgT
CD44 TCAACAgTggCAATggAgC gCAggTTCCTTgTCTCATCA
22
per well in a 24-well plate and grown overnight to confluence in complete medium. The monolayer was scratched with a pipette tip and washed with PBS to remove floating cells. The scrape was monitored and photographed after 4, 8, 12, 24 h incubation.
2.4.2 Invasion assay
A cell invasion assay was performed using a Boyden chamber (Millpore, Co., USA) with 8 μm pore polycarbonate filters that were coated with 40μg geltrex (Invitrogen)
which was diluted by serum-free RPMI-1640. After 72 hrs of transfection, 2*104 cells were resuspended by 200μL serum-free medium and seeded to the upper compartment
of the Boyden chamber. The lower chamber was filled with 1 mL of RPMI-1640 complete medium. The cells were allowed to migrate for 48 hrs, incubated at 37℃, 5%
CO2. The cells were then removed from the upper chamber using a cotton swab. The cells on the lower surface of the chamber were stained with Liu’s stain and counted.
Data representing the average number of cells per pixel in five fields were compared between the siRNA groups, NC groups, and blank control groups.
2.5 Adhesion assay
Ninety-six-well plates were coated with Collagen (Cohesion; Vitrogen), Fibronectin
(Sigma-Aldrich), Gelatin (Sigma-Aldrich) and BSA for 12 hours at 4°C. Each coating protein was dissolved in PBS (pH: 7.4) to yield a final concentration of 60 μg/mL, and a volume of 100 μL was added to each individual wells. The plates were then blocked
23
with 10 mg/mL Bovine serum albumin (Sigma-Aldrich) which was heated at 85℃ for 10 minutes in PBS for 1 hour at 37°C and washed with PBS twice.
Targeted knockdown cells were isolated by trypsinization and washed once in RPMI1640 with 10% FBS to stop trypsin activity. Then the cells were resolved in serum-free RPMI1640 to remove serum components. Suspensions of 104 cells/mL viable targeted knockdown cells were then added to each well and allowed to attach for 1 hour at 37°C, 5% CO2. To determine the cell adhesion ability, plates were then carefully washed three times with PBS and fixed by 5% paraformaldehyde for 15 minutes. And then they were washed by ddH2O three times and stained by crystal violet for 30 minutes. Then, they were washed by ddH2O three times and resolved crystal violet by 10 % acetic acid. Finally, the optical density was measure at 570 nm.
2.6 Protein extraction
The organic phase from the RNA extraction which contained protein was added to
300 μL of 100% ethanol per 1 mL of TRIzol reagent and then mixed by inverting the
sample. The mixture was stored for 3 minutes at room temperature and centrifuged for 2000 g for 5 minutes at 4℃. The supernatant was transferred to a new eppendorf and then 1.5 mL of isopropanol was added for homogenization. The mixture was stored at room temperature for 15 minutes and centrifuged at 12000 g for 10 minutes at 4℃. The supernatant was removed and the protein pellet was washed by a solution containing
24
0.3M guanindine hydrochloride in 95% ethanol three times. During each wash step, the protein pellet was stored in wash solution for 20 minutes at room temperature and centrifuged at 7500 g for 5 minutes at 4℃. After the final washing, the protein pellet was vortexed in 2 ml absolute ethanol and allowed to sit for 20 minutes at room temperature. In the final step, it was centrifuged at 7500g for 5 minutes at 4℃ and the supernatant was discarded. The protein pellet was resolved by the solution with 9.5 M Urea and 2% CHAPS and the sample was stored at -20 ℃ for further use.
2.7 Western blot
Protein samples (15 μg) were separated by SDS-PAGE and transferred to a PVDF membrane The membrane was blocked with 5% (w/v) BSA (sigma) in PBST for 1hr at room temperature, incubated with (1) 200 ng/mL rabbit polyclonal IgG to SLC3A2 (santa cruz, CA. U.S.A.), (2) 200 ng/mL mouse monoclonal IgG to CD166 (santa cruz, CA. U.S.A.), or (3) 400 ng/mL rabbit polyclonal IgG to CD44 (santa cruz, CA. U.S.A.)
in PBST involving 5% (w/v) BSA overnight at 4 ℃. The membrane was washed with
PBST for 30 mins twice and incubated with (1) 80 ng/mL Goat polyclonal secondary antibody to rabbit IgG HRP (Abcam, Cambridge, UK), or (2) 200 ng/mL rabbit polyclonal secondary antibody to Mouse IgG HRP (Abcam, Cambridge, UK) in PBST involving 5% BSA (v/v) for 1hr. The membrane was washed with PBST for 30 minutes twice. The immunoreactive bands were visualized by film exposure (GE Healthcare,
25
Uppsala, Sweden) through the ECL enhanced chemiluminescence detection system.
26
3. Results
3.1 Characteristics of A549-Mock and A549-FutIV
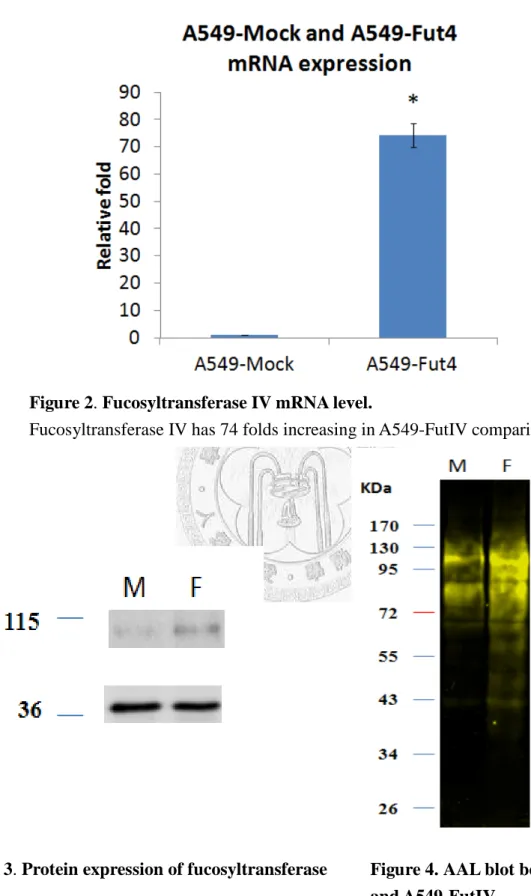
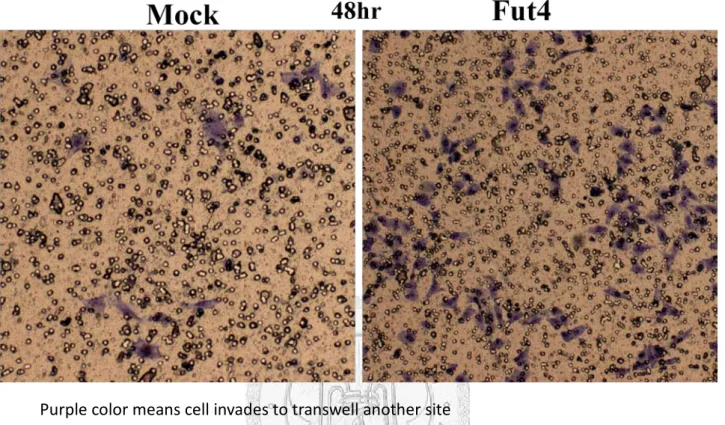
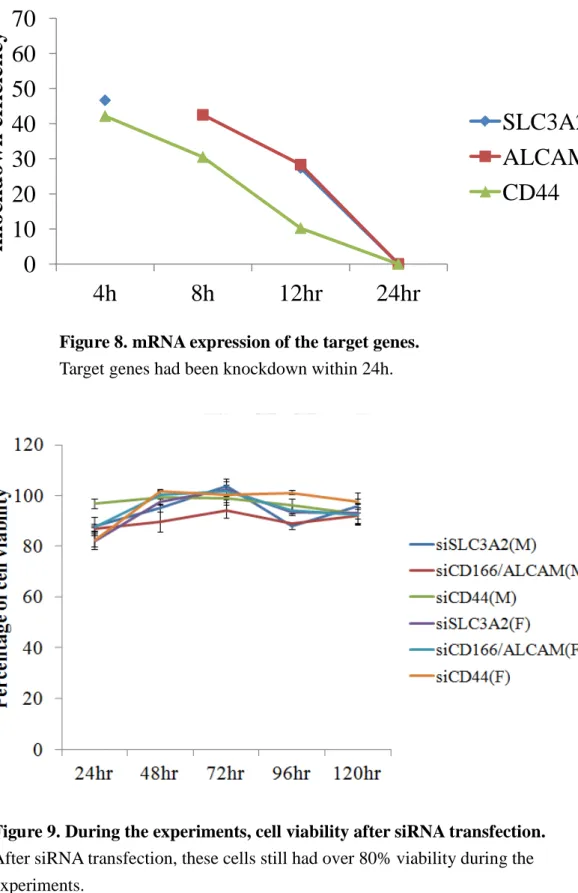
Under microscopic observation, the morphology of A549-Mock and A549-FutIV appeared different. The former had a thinner spindle shape and the latter had a round shape (Fig.1). To confirm the FUT IV expression, real-time PCR and western blot had been employed. The total RNA extractions from the two cell lines were extracted and reversely transcribed into cDNA. The FUT IV primers had been designed and it had about seventy-four folds change in real-time PCR examination (Fig.2). The FUT IV protein expression and fucosylation also had a significant change in A549-FutIV (Fig.3-4). In order to identify which cell lines were more malignant, we analyzed invasion ability to investigate their metastasis in vitro. The result showed that A549-FutIV had a higher degree of cell invasion to another transwell site than A549-Mock (Fig.5).
3.2 Functional network analysis of the nineteen genes
The 19 membrane proteins that were more or less saturated with fucose were analyzed by Ingenuity Pathways Analysis. There were 12 membrane proteins that could be linked to form a network which were associated with cell morphology, cell movement and cell assembly (Fig.6). Six of them were relative to tumorigenesis, metastasis and cell migration, namely PRNP, ITGA3, CD44, EPHA2, DSG2 and
27
CD166. There was one protein associated with amino acid transport, SLC3A2 (Fig.7).
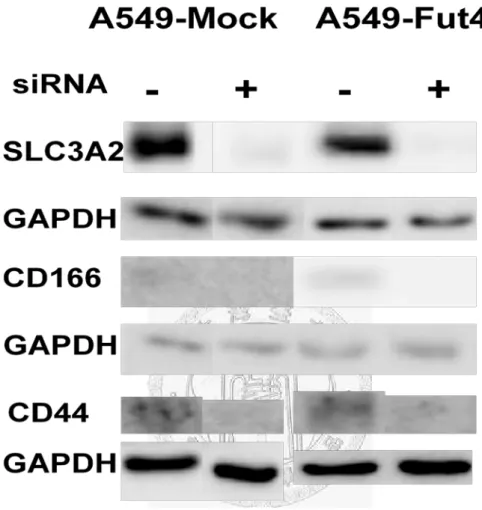
3.3 Analysis of the siRNA knockdown effect in A549-Mock and A549-FutIV
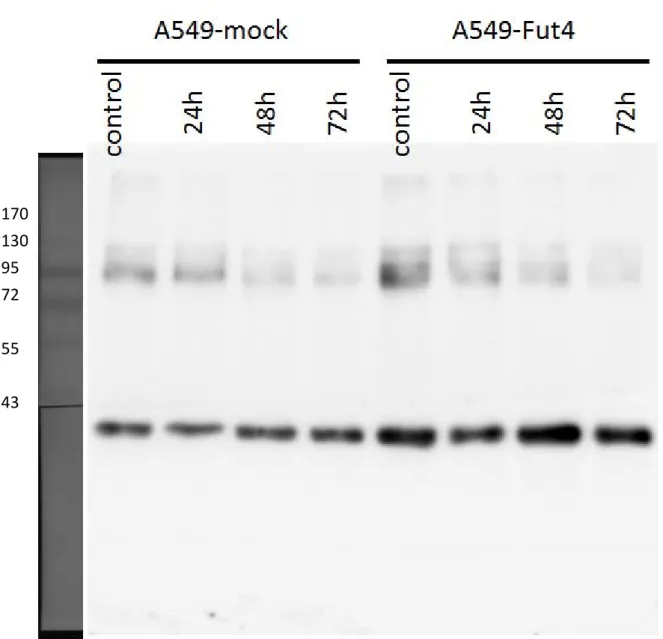
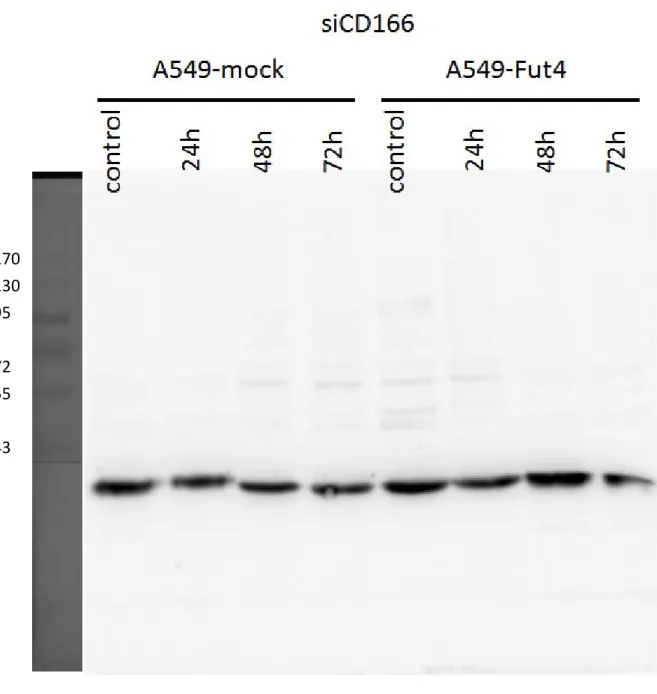
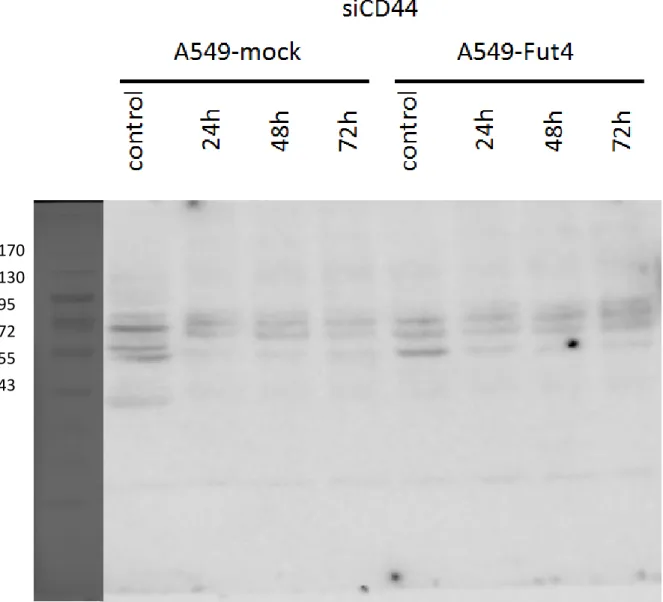
siRNA had been employed to decrease the target genes expression level for SLC3A2, CD166 and CD44. Under the rules of siRNA knockdown, the knockdown efficiency of target genes must be over 90%. In other words, the target gene expression must lower than 10% compared to the control. From the quantitative polymerase chain reaction examination, all the genes’ expressions decreased below 0.06 compared to the control after 24h transfection (Fig.8). Transfected-cell lines had no cytotoxicity from siRNA or transfection reagent and their viabilities also still had over 80% during the experiments (Fig.9). After siRNA transfection 3 days, the target proteins decreased(Fig.10-10.3).
3.4 Functional analysis of the target genes after genes knockdown
3.4.1 The effect of target proteins invasion ability
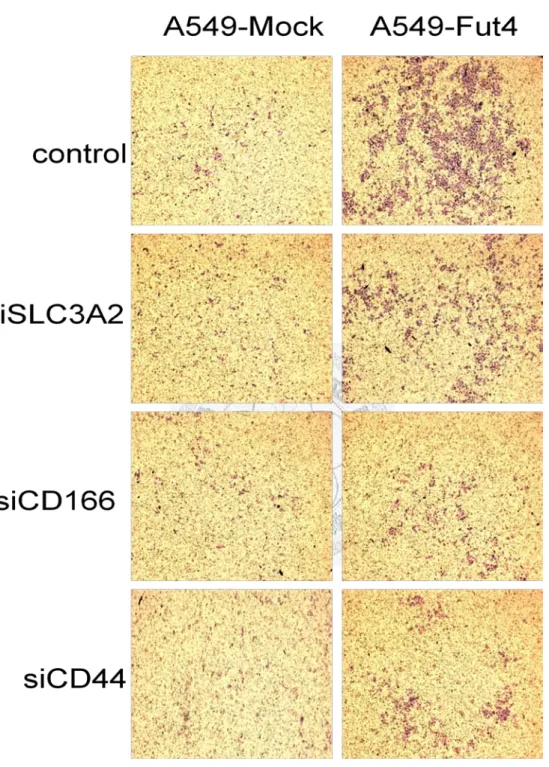
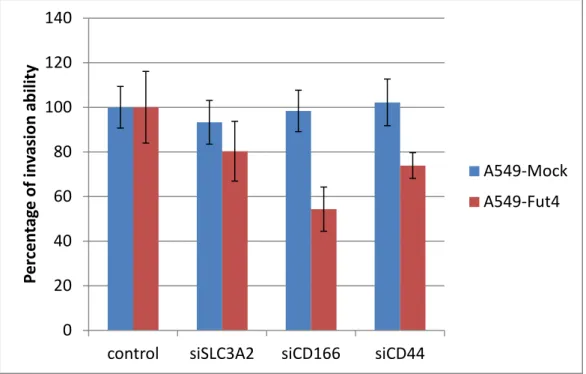
Since membrane proteins could affect cancer metastasis, we investigated whether the membrane proteins influenced it and made cancer more malignant. In an in vitro study, cell invasion was assessed using Boyden chambers. While cancer cells invaded another side of the transwell, they would be stained by Liu’s stain, causing them to displayed a purple color(Fig.11). Knockdown for all the three genes showed different invasion ability compared to the control (Fig.12) and had different fold changes in cell counting (Table2). From the results, CD166 had the greatest ability to inhibit cancer cell
28
invasion, especially in the more malignant cell line, A549-FutIV. The second strongest invasion inhibitor was CD44 and the weakest was SLC3A2. All transfected A549-mock cell lines had no significant difference regarding cancer cells invasion inhibition. When all the transfected-cell lines were compared to A549-Mock, the invasion ability of A549-FutIV that was transfected with CD166 was closest in similarity to A549-mock (Fig.13 Table3).
3.4.2 The effect of target proteins adhesion ability
In order to investigate how these fucosylated membrane proteins after knockdown could affect cell adhesion, the cell adhesion ability was studied with different ECMs.
We employed fibronectin, collagen, gelatin and BSA as negative controls. With different ECM coatings, the transfected cell lines showed different adhesion abilities to different ECMs (Fig.14). CD166 knockdown of the A549-FutIV cell line had significantly decreased its adhesion ability to collagen, gelatin and fibronectin. SLC3A2 had slightly reduced its binding ability to fibronectin and CD44 had slightly reduced its binding ability to collagen.
3.4.3 The effect of target proteins migration ability
To investigate how these proteins after knockdown could affect cell migration. We
used would healing assay to study their migration ability and found that when SLC3A2 was knockdown, the migration ability of the cells increased comparing to control after
29
8h. CD166 and CD44 did not have any significantly changed.(Fig.15)
30
4. Discussion
4.1 Fucosyltransferase IV over-expression in A549 might make cancer more
malignant
In our observation, the fucosyltranferaseIV over-expressed A549 lung carcinoma showed relatively more malignancy compared with others. Regarding invasion ability, A549-FutIV was more invasive than A549-Mock and it was thought that invasion ability might relate to MMP activities. In the zymography assay, A549-FutIV showed that it had more MMP2/9 proteins expression than A549-mock. This meant that the A549-FutIV cell line could reach metastasis and make cancer more malignant.
4.2 The relationship between membrane proteins and cancer metastasis
4.2.1 SLC3A2
SLC3A2 is an amino acid transporter heavy chain that is able to form heterodimers with other light chains, such as L-type amino acid transporter 1(Yanagida et al, 2001), LAT2(Broer et al, 2000), y+LAT2(Broer et al, 2001) and others. In addition, it can stabilize the activity of the glucose transporter GLUT1(Ohno et al, 2011). During cell growth, SLC3A2(CH98hc) also plays a role in mediating integrin signaling. Because of integrin mediation, SLC3A2 might influence the cell shape. SLC3A2 over-expression would lead to anchorage-independent cell growth and tumorigenesis in 3T3 fibroblast and activation of certain integrin-regulated signaling(Feral et al, 2005). As a tumor
31
marker, SLC3A2 has been observed to express in renal cell cancer and it was correlated directly with grade of malignancy(Prager et al, 2009). In our study, although SLC3A2 knockdown could decrease cell viability to 80%, the transfected-A549 cell lines did not show a significant change in their invasion ability nor cell shape. This means that SLC3A2 might not affect cancer metastasis even though it mediates integrin signaling.
4.2.2 CD166
CD166 was classified as a cell adhesion protein and that could create neurite extension via heterophilic and homophilic interactions(Swart, 2002; Wade et al, 2012).
In immunoreactions, CD166 binds to CD6, which is a cell surface receptor expressed on immature thymocytes and mature T and B1a lymphocytes and form a central supramolecular activation cluster. So, it plays an important role in intra- and intercellular molecular interactions mediated by CD6 during T cell activation and proliferation processes(Gimferrer et al, 2004; Hassan et al, 2004; Skonier et al, 1996a;
Zimmerman et al, 2006b). But in cancer studies, CD166 expression levels were higher in primary tumors and lower in metastase or in higher-grade lesions, such as found in prostate cancer(Kristiansen et al, 2003) and breast cancer(King et al, 2004). Further, in a clinical analysis, CD166 over-expressed in colorectal carcinoma and shortened patients survival(Weichert et al, 2004). In our study, we used siRNA to knockdown CD166 expression and found that it could inhibit cancer invasion ability in lung
32
adenocarcinoma. From a recent study, we know that CD166 can control matrix metalloproteinase activity in melanoma cell lines(Lunter et al, 2005). Cancer invasion ability might decrease because CD166 controlled MMPs activities. But the reason that primary tumors showed higher CD166 expression while lower during metastases is still unknown(Ofori-Acquah & King, 2008).
4.2.3 CD44
Previous research suggests that CD44 has high expression on primary tumors of malignant melanoma, causing patients to have poor survival(Dietrich et al, 1997) and in prostate cancer, they found that primary tumors were expressing CD44 at increased levels while lower in metastases (Noordzij et al, 1999). In our study, the CD44 function was found to be associated with cancer metastasis from the in vitro study, and further support from previous works is that CD44 expression was found related to lymph node metastasis in non-small cell lung cancer by a clinical statistical analysis(Miyoshi et al, 1997). It might mediate cancer invasion via cyclooxygenase-2-dependent pathway (Dohadwala et al, 2001).
4.3 Characteristics of the three proteins regarding cell adhesion to ECMs
In our observation, although SLC3A2 is an amino acid transporter, it still could bind to ECMs and provide weak cell adhesion. Furthermore, we found that CD166 could also bind to collagen, gelatin and fibronectin and it influenced cancer adhesion ability. It
33
might provide cancer migration when cancer invades the ECM. CD44 is an adhesion protein and a hyaluronate receptor, which could bind to some ECM, like collagen, fibronectin, laminin and so on (Aruffo et al, 1990; Ishii et al, 1993; Jalkanen & Jalkanen, 1992; Lokeshwar & Bourguignon, 1991). But in our study, CD44 knockdown cell lines did not have any significantly change in cell adhesion, which might result from CD44 being expressed at a very low level (Fig.3c) and thus unable to make strong adhesion(Fig.4k).
4.4 The protein functions as related to invasion and adhesion ability
Membrane proteins are cell surface proteins that can provide cell-cell adhesion, cell migration, signal transduction and so on. Cancer cell metastasis must attach to the ECM or basement membrane and secrete some proteinase, like MMPs, to digest the ECM and ease migration. CD166 provided a binding ability to ECMs, whcih meant that cancer cells might migrate to the other site due to CD166. When CD166 was knockdown, there had be a significant change, not only in invasion ability, but also cell adhesion ability in malignant cancer cells. SLC3A2 had a slight adhesion ability to the ECM, so it might increase cell adhesiveness. CD44 antigen presented a positive control to cancer invasion and cell adhesion when it was knockdown.
34
5. References
1. Aruffo A, Stamenkovic I, Melnick M, Underhill CB, Seed B (1990) Cd44 Is the Principal Cell-Surface Receptor for Hyaluronate. Cell 61: 1303-1313
2. Beck-Sickinger AG, Mörl K (2006) Posttranslational modification of proteins : Expanding nature's inventory, Vol. 45: Roberts and Co.
3. Becker WM, Kleinsmith LJ, Hardin J (2004) The world of the cell, sixth edn.:
Pearson.
4. Borsig L, Wong R, Hynes RO, Varki NM, Varki A (2002) Synergistic effects of L- and P-selectin in facilitating tumor metastasis can involve non-mucin ligands and implicate leukocytes as enhancers of metastasis. Proceedings of the National Academy of Sciences of the United States of America 99: 2193-2198
5. Broer A, Friedrich B, Wagner CA, Fillon S, Ganapathy V, Lang F, Broer S (2001) Association of 4F2hc with light chains LAT1, LAT2 or y(+)LAT2 requires different domains. Biochem J 355: 725-731
6. Broer A, Wagner CA, Lang F, Broer S (2000) The heterodimeric amino acid transporter 4F2hc/y+LAT2 mediates arginine efflux in exchange with glutamine.
Biochem J 349 Pt 3: 787-795
7. Christofori G, Semb H (1999) The role of the cell-adhesion molecule E-cadherin as a tumour-suppressor gene. Trends in Biochemical Sciences 24: 73-76
35
8. Conrotto P, Corso S, Gamberini S, Comoglio PM, Giordano S (2004) Interplay between scatter factor receptors and B plexins controls invasive growth. Oncogene 23: 5131-5137
9. Consortium IHGS (2004) Finishing the euchromatic sequence of the human genome. Nature 431: 15
10. Dietrich A, Tanczos E, Vanscheidt W, Schopf E, Simon JC (1997) High CD44 surface expression on primary tumours of malignant melanoma correlates with increased metastatic risk and reduced survival. European journal of Cancer 33:
926-930
11. Dohadwala M, Luo J, Zhu L, Lin Y, Dougherty GJ, Sharma S, Huang M, Pold M, Batra RK, Dubinett SM (2001) Non-small cell lung cancer cyclooxygenase-2-dependent invasion is mediated by CD44. The Journal of biological chemistry 276: 20809-20812
12. Drummond MJ, Glynn EL, Fry CS, Timmerman KL, Volpi E, Rasmussen BB (2010) An increase in essential amino acid availability upregulates amino acid transporter expression in human skeletal muscle. American journal of physiology Endocrinology and metabolism 298: E1011-1018
13. Feral CC, Nishiya N, Fenczik CA, Stuhlmann H, Slepak M, Ginsberg MH (2005) CD98hc (SLC3A2) mediates integrin signaling. Proceedings of the National
36
Academy of Sciences of the United States of America 102: 355-360
14. Feral CC, Zijlstra A, Tkachenko E, Prager G, Gardel ML, Slepak M, Ginsberg MH (2007) CD98hc (SLC3A2) participates in fibronectin matrix assembly by mediating integrin signaling. The Journal of cell biology 178: 701-711
15. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM (2010) Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. International journal of cancer Journal international du cancer 127: 2893-2917
16. Friedl P, Wolf K (2003) Tumour-cell invasion and migration: diversity and escape mechanisms. Nature reviews Cancer 3: 362-374
17. Gimferrer I, Calvo M, Mittelbrunn M, Farnos M, Sarrias MR, Enrich C, Vives J, Sanchez-Madrid F, Lozano F (2004) Relevance of CD6-mediated interactions in T cell activation and proliferation. J Immunol 173: 2262-2270
18. Goodison S, Urquidi V, Tarin D (1999) CD44 cell adhesion molecules. J Clin Pathol-Mol Pa 52: 189-196
19. Gorlova OY, Weng SF, Zhang Y, Amos CI, Spitz MR (2007) Aggregation of cancer among relatives of never-smoking lung cancer patients. International journal of cancer Journal international du cancer 121: 111-118
20. Gupta GP, Nguyen DX, Chiang AC, Bos PD, Kim JY, Nadal C, Gomis RR, Manova-Todorova K, Massague J (2007) Mediators of vascular remodelling
37
co-opted for sequential steps in lung metastasis. Nature 446: 765-770
21. Hanisch FG (2001) O-glycosylation of the mucin type. Biological Chemistry 382:
143-149
22. Hassan NJ, Barclay AN, Brown MH (2004) Frontline: Optimal T cell activation requires the engagement of CD6 and CD166. Eur J Immunol 34: 930-940
23. Hayashi H, Yamashita Y (2012) Role of N-glycosylation in cell surface expression and protection against proteolysis of the intestinal anion exchanger SLC26A3.
American journal of physiology Cell physiology 302: C781-795
24. Herbst RS, Heymach JV, Lippman SM (2008) Molecular origins of cancer: Lung cancer. New Engl J Med 359: 1367-1380
25. Homeister JW, Thall AD, Petryniak B, Maly P, Rogers CE, Smith PL, Kelly RJ, Gersten KM, Askari SW, Cheng G, Smithson G, Marks RM, Misra AK, Hindsgaul O, von Andrian UH, Lowe JB (2001) The alpha(1,3)fucosyltransferases FucT-IV and FucT-VII exert collaborative control over selectin-dependent leukocyte recruitment and lymphocyte homing. Immunity 15: 115-126
26. Hong WK, Tsao AS. (2008) Lung Carcinoma.
27. Ishii S, Ford R, Thomas P, Nachman A, Steele G, Jessup JM (1993) Cd44 Participates in the Adhesion of Human Colorectal-Carcinoma Cells to Laminin and Type-Iv Collagen. Surg Oncol 2: 255-264
38
28. Jalkanen S, Jalkanen M (1992) Lymphocyte CD44 binds the COOH-terminal heparin-binding domain of fibronectin. The Journal of cell biology 116: 817-825 29. Javaud C, Dupuy F, Maftah A, Julien R, Petit JM (2003) The fucosyltransferase
gene family: an amazing summary of the underlying mechanisms of gene evolution.
Genetica 118: 157-170
30. Jezierska A, Olszewski WP, Pietruszkiewicz J, Olszewski W, Matysiak W, Motyl T (2006) Activated Leukocyte Cell Adhesion Molecule (ALCAM) is associated with supression of breast cancer cells invasion. Medical Science Monitor 12: 12
31. Joyce JA, Pollard JW (2009) Microenvironmental regulation of metastasis. Nature reviews Cancer 9: 239-252
32. Kajita M, Itoh Y, Chiba T, Mori H, Okada A, Kinoh H, Seiki M (2001) Membrane-type 1 matrix metalloproteinase cleaves CD44 and promotes cell migration. Journal of Cell Biology 153: 893-904
33. King JA, Ofori-Acquah SF, Stevens T, Al-Mehdi AB, Fodstad O, Jiang WG (2004) Activated leukocyte cell adhesion molecule in breast cancer: prognostic indicator.
Breast cancer research : BCR 6: R478-487
34. Kirchhoff P, Dave MH, Remy C, Kosiek O, Busque SM, Dufner M, Geibel JP, Verrey F, Wagner CA (2006) An amino acid transporter involved in gastric acid secretion. Pflugers Archiv : European journal of physiology 451: 738-748
39
35. Kowalski PJ, Rubin MA, Kleer CG (2003) E-cadherin expression in primary carcinomas of the breast and its distant metastases. Breast cancer research : BCR 5:
R217-222
36. Kristiansen G, Pilarsky C, Wissmann C, Stephan C, Weissbach L, Loy V, Loening S, Dietel M, Rosenthal A (2003) ALCAM/CD166 is up-regulated in low-grade prostate cancer and progressively lost in high-grade lesions. The Prostate 54:
34-43
37. Lau KS, Dennis JW (2008) N-Glycans in cancer progression. Glycobiology 18:
750-760
38. Leth-Larsen R, Lund R, Hansen HV, Laenkholm AV, Tarin D, Jensen ON, Ditzel HJ (2009) Metastasis-related plasma membrane proteins of human breast cancer cells identified by comparative quantitative mass spectrometry. Molecular &
cellular proteomics : MCP 8: 1436-1449
39. Lin S, Gordon K, Kaplan N, Getsios S (2010) Ligand targeting of EphA2 enhances keratinocyte adhesion and differentiation via desmoglein 1. Molecular biology of the cell 21: 3902-3914
40. Lodish H, Berk A, L. ZS (2000) Molecular Cell Biology: W. H. Freeman.
41. Lokeshwar VB, Bourguignon LY (1991) Post-translational protein modification and expression of ankyrin-binding site(s) in GP85 (Pgp-1/CD44) and its
40
biosynthetic precursors during T-lymphoma membrane biosynthesis. The Journal of biological chemistry 266: 17983-17989
42. Lunter PC, van Kilsdonk JW, van Beek H, Cornelissen IM, Bergers M, Willems PH, van Muijen GN, Swart GW (2005) Activated leukocyte cell adhesion molecule (ALCAM/CD166/MEMD), a novel actor in invasive growth, controls matrix metalloproteinase activity. Cancer research 65: 8801-8808
43. Malý P, Thall A, Petryniak B, Rogers CE, Smith PL, Marks RM, Kelly RJ, Gersten KM, Cheng G, Saunders TL, Camper SA, Camphausen RT, Sullivan FX, Isogai Y, Hindsgaul O, von Andrian UH, Lowe JB (1996) The alpha(1,3)fucosyltransferase Fuc-TVII controls leukocyte trafficking through an essential role in L-, E-, and P-selectin ligand biosynthesis. Cell 86: 643-653
44. Miyoshi E, Moriwaki K, Nakagawa T (2008) Biological function of fucosylation in cancer biology. Journal of biochemistry 143: 725-729
45. Miyoshi T, Kondo K, Hino N, Uyama T, Monden Y (1997) The expression of the CD44 variant exon 6 is associated with lymph node metastasis in non-small cell lung cancer. Clinical Cancer Research 3: 1289-1297
46. NCI. (2012) General information about non-small cell lung cancer.
47. Nel MJ, Woodiwiss AJ, Candy GP (2012) Modeling of cellular arginine uptake by more than one transporter. The Journal of membrane biology 245: 1-13
41
48. Noordzij MA, van Steenbrugge GJ, Schroder FH, Van der Kwast TH (1999) Decreased expression of CD44 in metastatic prostate cancer. International Journal of Cancer 84: 478-483
49. Ofori-Acquah SF, King JA (2008) Activated leukocyte cell adhesion molecule: a new paradox in cancer. Translational research : the journal of laboratory and clinical medicine 151: 122-128
50. Ogawa K, Pasqualini R, Lindberg RA, Kain R, Freeman AL, Pasquale EB (2000) The ephrin-A1 ligand and its receptor, EphA2, are expressed during tumor neovascularization. Oncogene 19: 6043-6052
51. Ohno H, Nakatsu Y, Sakoda H, Kushiyama A, Ono H, Fujishiro M, Otani Y, Okubo H, Yoneda M, Fukushima T, Tsuchiya Y, Kamata H, Nishimura F, Kurihara H, Katagiri H, Oka Y, Asano T (2011) 4F2hc stabilizes GLUT1 protein and increases glucose transport activity. Am J Physiol-Cell Ph 300: C1047-C1054
52. Onder TT, Gupta PB, Mani SA, Yang J, Lander ES, Weinberg RA (2008) Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways.
Cancer research 68: 3645-3654
53. Padua D, Zhang XH, Wang Q, Nadal C, Gerald WL, Gomis RR, Massague J (2008) TGFbeta primes breast tumors for lung metastasis seeding through angiopoietin-like 4. Cell 133: 66-77
42
54. Perrot V, Vazquez-Prado J, Gutkind JS (2002) Plexin B regulates Rho through the guanine nucleotide exchange factors leukemia-associated Rho GEF (LARG) and PDZ-RhoGEF. The Journal of biological chemistry 277: 43115-43120
55. Pfeiffer R, Rossier G, Spindler B, Meier C, Kuhn L, Verrey F (1999) Amino acid transport of y(+)L-type by heterodimers of 4F2hc/CD98 and members of the glycoprotein-associated amino acid transporter family. Embo J 18: 49-57
56. Pinho SS, Osorio H, Nita-Lazar M, Gomes J, Lopes C, Gartner F, Reis CA (2009) Role of E-cadherin N-glycosylation profile in a mammary tumor model.
Biochemical and biophysical research communications 379: 1091-1096
57. Prager GW, Poettler M, Schmidinger M, Mazal PR, Susani M, Zielinski CC, Haitel A (2009) CD98hc (SLC3A2), a novel marker in renal cell cancer. European journal of clinical investigation 39: 304-310
58. Skonier JE, Bowen MA, Emswiler J, Aruffo A, Bajorath J (1996a) Recognition of diverse proteins by members of the immunoglobulin superfamily: Delineation of the receptor binding site in the human CD6 ligand ALCAM. Biochemistry 35:
12287-12291
59. Skonier JE, Bowen MA, Emswiler J, Aruffo A, Bajorath J (1996b) Recognition of Diverse Proteins by Members of the Immunoglobulin Superfamily: Delineation of the Receptor Binding Site in the Human CD6 Ligand ALCAM. Biochemistry 35
43
60. Swart GW (2002) Activated leukocyte cell adhesion molecule (CD166/ALCAM):
developmental and mechanistic aspects of cell clustering and cell migration.
European Journal of Cell Biology 81: 313-321
61. Thiery JP, Acloque H, Huang RY, Nieto MA (2009) Epithelial-mesenchymal transitions in development and disease. Cell 139: 871-890
62. Tsuboi S, Hatakeyama S, Ohyama C, Fukuda M (2012) Two opposing roles of O-glycans in tumor metastasis. Trends in molecular medicine 18: 224-232
63. van Kempen LC, Meier F, Egeblad M, Kersten-Niessen MJ, Garbe C, Weidle UH, Van Muijen GN, Herlyn M, Bloemers HP, Swart GW (2004) Truncation of Activated Leukocyte Cell Adhesion Molecule: A Gateway to Melanoma Metastasis.
The Journal of Investigative Dermatology 122
64. Varki A, Cummings RD, Esko JD, Freeze HH, Stanley P, Bertozzi CR, Hart GW, Etzler ME (2009) Essentials of Glycobiology, second edn.: Cold Spring Harbor Laboratory Press.
65. Verrey F, Singer D, Ramadan T, Vuille-dit-Bille RN, Mariotta L, Camargo SM (2009) Kidney amino acid transport. Pflugers Archiv : European journal of physiology 458: 53-60
66. Wade A, Thomas C, Kalmar B, Terenzio M, Garin J, Greensmith L, Schiavo G (2012) Activated leukocyte cell adhesion molecule modulates neurotrophin
44
signaling. Journal of neurochemistry 121: 575-586
67. Wang W, Goswami S, Lapidus K, Wells AL, Wyckoff JB, Sahai E, Singer RH, Segall JE, Condeelis JS (2004) Identification and testing of a gene expression signature of invasive carcinoma cells within primary mammary tumors. Cancer research 64: 8585-8594
68. Weichert W, Knosel T, Bellach J, Dietel M, Kristiansen G (2004) ALCAM/CD166 is overexpressed in colorectal carcinoma and correlates with shortened patient survival. Journal of clinical pathology 57: 1160-1164
69. Wykosky J, Debinski W (2008) The EphA2 receptor and ephrinA1 ligand in solid tumors: function and therapeutic targeting. Molecular cancer research : MCR 6:
1795-1806
70. Yanagida O, Kanai Y, Chairoungdua A, Kim DK, Segawa H, Nii T, Cha SH, Matsuo H, Fukushima J, Fukasawa Y, Tani Y, Taketani Y, Uchino H, Kim JY, Inatomi J, Okayasu I, Miyamoto K, Takeda E, Goya T, Endou H (2001) Human L-type amino acid transporter 1 (LAT1): characterization of function and expression in tumor cell lines. Biochimica et Biophysica Acta (BBA) - Biomembranes 1514: 291-302
71. Yu Q, Stamenkovic I (1999) Localization of matrix metalloproteinase 9 to the cell surface provides a mechanism for CD44-mediated tumor invasion. Genes Dev 13:
45
35-48
72. Zimmerman AW, Joosten B, Torensma R, Parnes JR, van Leeuwen FN, Figdor CG (2006a) Long-term engagement of CD6 and ALCAM is essential for T-cell proliferation induced by dendritic cells. Blood 107: 3212-3220
73. Zimmerman AW, Joosten B, Torensma R, Parnes JR, van Leeuwen FN, Figdor CG (2006b) Long-term engagement of CD6 andALCAM is essential for T-cell proliferation induced by dendritic cells. Blood 107: 3212-3220
46
6.Figure
Figure1. A549-mock and A549-FutIV cell shapes A549-mock had a spindle shape and
A549-FutIV had a round shape.
47
Figure 2. Fucosyltransferase IV mRNA level.
Fucosyltransferase IV has 74 folds increasing in A549-FutIV comparing to A549-Mock.
Figure 3. Protein expression of fucosyltransferase IV.
In protein expression, fucosyltransferase IV also showed higher expression in A549-FutIV cell line.
Figure 4. AAL blot between A549-Mock and A549-FutIV.
A549-Fut4 proteins had been more decorated fucose than A549-Mock.
48
Purple color means cell invades to transwell another site
Figure 5. The invasion ability between A549-Mock and A549-FutIV A549-FutIV showed more invaded than A549-Mock after 48h treating.
49
Figure 6. 12 membrane proteins network.
There are 12 membrane proteins which are associated with cell adhesion, cell migration can belinked to a network.
50
Figure 7. Protein function classification.
Most of them could be classified to cell adhesion or cell migration. One of them is related to amino acid transport.