海產食品的品質與其貯藏中腐敗的起因 The Quality of Seafoods and the Causes for
Their Spoilage during Storage
邱思魁 Tze-Kuei Chiou
國立臺灣海洋大學生命科學院食品科學系名譽教授
Received 09 May 2019; revised 16 July 2019; accepted 08 August 2019; available online 09 September 2019
摘 要
本文介紹海產食品利用上最重要的三項問題,第一項為海產食品的品質與其影響 因素,重點包括:食品品質的屬性、海產食品原料的感官性質、可食用和不可食用部 位比率、主要組成分含量與其營養價值和生物活性、過敏原/毒素和寄生蟲的影響、生 物與環境因素對海產食品資源加工價值的影響等。第二項為捕撈後的變化,重點包 括:能量儲備耗盡的影響、含氮化合物的變化、脂質水解和氧化、影響變化速率的因 素等。第三項為魚類貯藏中的腐敗,重點包括:自家消化的變化、酵素性的變化、微 生物的作用(分為微生物的變化、棲息地不同水生動物的微生物菌群、微生物的腐敗、
特定腐敗生物等)、化學性腐敗、生鮮魚的感官變化等。
關鍵字:海產食品、品質、捕撈後的變化、貯藏、腐敗、微生物作用。
一、前言
食品品質(food quality)有兩個面向,
其 一 為 那 些 能 夠 使 用 心 理 物 理 學 方 法 (psychophysical methology)品評的,這反 映消費或嚐到平常條件下製備和提供的 食品產品所感知的特徵印象(characteristic impression),另一對品質的認識為許多物
*通訊作者電子信箱:[email protected]
理 、 生 物 或 化 學 可 測 得 的 性 質( 屬 性 attributes) 而 決 定 了 優 良 程 度 (degree of excellence)的總和,這些屬性表明產品到 底符合最終貢獻於消費者滿意度的各種 要求和標準的程度高低。針對海產食品 (seafood)原料,本文所提的品質此術語只 要 了 解 是 可 測 量 性 質 (measurable properties)的總和,影響食品品質的特徵
屬 性 會 因 商 品 的 不 同 而 異(Sikorski and
Kołakowski, 2011)。
海產食品品質的保證是整個產品供 應鏈中生產者和參與者的責任,這乃基 於了解遺傳因素和水生棲息地對魚類和 海洋無脊椎動物的建構、組成、代謝活 性、寄生蟲感染和污染的影響,還有漁 法及漁獲後處置和捕撈後貯藏對腐敗發 生速率的影響。有很多技術可用來測定 和紀錄環境與操作上的重要參數,以及 原料的各種性質,紀錄的數據用於各種 品質管理系統,例如良好作業規範(Good Manufacturing Practice; GMP)和危害分析 重要管制點(Hazard Analysis and Critical Control Points; HACCP),在全面品質管 理(Total Quality Management; TQM)計畫 內實施(Schiefer, 2007)。
海產食品包括各種魚類、甲殼類動 物(crustaceans)、軟體類動物(mollusks)和 棘 皮 類 動 物(echinoderms)是蛋白質、脂 肪、維生素和礦物質的優良來源,且兼 具 美 味 又 營 養 價 值 高 而 廣 受 喜 好 。 然 而,由於各種水產原料營養成分的含量 高、中性 pH 值和水分含量高,海產食品 的貯藏壽命(shelf life)就有所受限(Viji et
al ., 2017)。在死後不久的海產食品原料
中,微生物和生化反應很快就發生而導 致感官和營養特性的變化,從而縮短貯 藏壽命(Olatunde and Benjakul, 2018a)。通 常,海產食品原料含有豐富的多元不飽 和脂肪酸(polyunsatured fatty acids),這使 之更易發生脂質氧化,如令人不快的氣
味和風味的形成、營養損失、不健康分 子的產生和顏色變化都主要是脂質氧化 所造成(Secci and Parisi, 2016)。
微生物、化學和物理變化共同貢獻 於海產食品腐敗的複雜性(Oladunde and
Benjakul, 2018b
)。魚類鮮度在初期的下 降歸因於生物體內原有的酵素和化學的 反 應 , 而 魚 類 的 完 全 腐 敗(complete spoilage)是微生物代謝活性的作用(Sriket,2014)。收穫或漁獲地點與加工設施之間
的距離、儲存溫度和加工方法對於品質 和變質(deterioration)都至關重要。外在和 內在的因素以及漁獲法對於海產食品的 品質和貯藏壽命可能是正面或負面的影 響(DeWitt and Oliveira, 2016)。由於對新 鮮冷藏食品,特別優質海產食品的需求 高,採用多樣的保存技術和非加熱技術 延 長 食 品 的 貯 藏 壽 命 也 愈 受 注 目 (Olatunde and Benjakul, 2018b; Sallam,2007)。
針對影響海產食品的穩定性和貯藏 壽命,
Toldra and Reig (2016)
指出:腐敗 是一種自然的過程,在其中海產食品經 歷了變質,從顏色和滋味的損失開始,接著質地和顏色的改變以及不良風味(off- flavors)的生成。腐敗主要是由於微生物 和內源性酵素的作用,與貯藏壽命密切 相關。海洋中捕獲的魚類因激烈運動,
通常會耗盡肝醣(glycogen),因此死後肌 肉中不能產生乳酸,使得肌肉 pH 接近中 性而有利於微生物的生長,而一些加速 腐敗速率之物理性損傷的產生則取決於
原料如何處置。一般,大型魚類的腐敗 比小型魚類更慢,寡脂魚(lean fish)比多 脂 魚(fatty fish) 更 易 腐 敗 (Hyldig et al.,
2007)。皮較厚比較薄者對微生物而言提
供更好的保護,另一方面,死後肌肉中 許 多 生 化 反 應 如 蛋 白 質 水 解 (proteolysis)、脂質水解(lipolysis)、醣解 (glycolysis) 和 氧 化 的 發 生 (Nielsen andNielsen, 2006),依據速率和強度而定,也
造成貯藏壽命長短不同。二、海產食品的品質與影響因素(Sikorski
and Kołakowski, 2011)
(一) 食品品質的屬性
論及食品品質時,最普遍應用的屬 性如下所述:
層 面(dimensions) 、 形 狀 、 顏 色 、 光 澤 、 香 氣 、 氣 味 、 風 味、滋味、質地和應力下的變 形(deformation under stress) 。
物種身分(species identity) 。
符合官方限制的安全性面向,
包括毒素、殺蟲劑、重金屬、
礦物質、病原性微生物、寄生 蟲和過敏原。
影 響 組 成 分 的 消 化 性 (digestibility) 和 營 養 可 利 用 性 (nutritional availability),例如特 定酵素的活性和酵素抑制劑。
營 養 和 健 康 促 進 組 成 分 的 含 量,主要是蛋白質和必需胺基 酸、脂質和必需脂肪酸、可消
化醣類、纖維、礦物質成分、
維 生 素 、 益 生 ( 菌 ) 元 (prebiotics) 、 益 生 菌 (probiotics)、色素、抗氧化物和 自由基清除劑。
在加工中會影響原料表現特定 技術上的功能性的適合程度之 那些特性,例如某些酵素的活 性、介面活性劑或食物基質中 參與各種交互作用的其它組成 分有否存在。
來 自 特 定 的 大 小 、 形 狀 、 質 地、化學組成、代謝率、腐敗 微生物菌群體的特徵,以及在 特定儲存條件下會影響商品貯 藏壽命之不當處理等的性質。
與主要針對收穫中和收穫後處 置(handling)的條件、各種組成 分含量、添加物、污染物、基 因 改 造 生 物 及 其 新 陳 代 謝 產 物,以及衛生、包裝和標示需 求等各種食品法規和標準的相 容性。
關於罐頭、軟袋(pouch)或包裝 盒大小和開封、再封蓋和處置 的尺寸和方便性方面,不同烹 煮(cooking)和烹飪處理(curlinary treatments)的適用性,產品輸送 和儲存期間特定處理的需求,
建 議 產 品 如 何 使 用 之 內 附 資 料。
就海產食品品質,在特別有特徵的 (characteristic)特點(features)之中,乃那些 與鮮度(freshness)有關的,即表示和剛捕 獲魚類、軟體類動物和甲殼類動物的那 些性質的相似度,這些可利用感官分析 或化學/儀器技術來測量。鮮度的好壞程 度可以魚冰藏時仍保留與剛捕獲時同樣 的感官屬性之日數表示,這通常決定了 原料作為食品烹煮和加工製作食品等各 種用途的適用性,也大大影響海產食品 的市場價格。其它特徵屬性包括捕撈的 可 追 溯 性(traceability) 、 季 節 和 漁 撈 技 術、物種身分和收穫後的處置。
(二) 海產食品原料的感官性質(sensory properties)
1. 顏色。剛捕獲魚類和海洋無脊椎 動 物 的 色 澤 主 要 來 自 類 胡 蘿 蔔 素 (carotenoids),以蝦紅素(astaxanthin)、角 黃素(canthaxanthin)、鮪黃素(tunaxanthin) 及 其 衍 生 物 為 主 , 與 蛋 白 質 、 醣 蛋 白 (glycoproteins) 、 磷 酸 化 醣 蛋 白 (phosphorylated glycoproteins)、醣脂蛋白 (glycolipoproteins) 及 脂 蛋 白 (lipoproteins) 形成非共價結合的複合物而存在皮膚和 甲殼,這些色素顯現為黃色、橙色、紅 色、紫色、藍色、銀白色或綠色。魚直 接在明亮光線下儲存,複合物會解離而 褪色。其它參與形成皮膚顏色的色素包 括褐黑色的黑色素類(melanins)和黑色素 蛋白類(melanoproteins)、藍紫色的靛藍素 類(indogoins)、四吡咯類(tetrapyrroles)和 黃物類(flavins)。生鮮龍蝦的藍色或藍灰
色甲殼是來自水溶性胡蘿蔔素的甲殼黃 素(crustaxanthin) , 甲 殼 類 在 水 中 煮 熟 後,蛋白質複合物變性而釋出游離態蝦 紅素,使得煮熟成品變成鮮紅色。貯藏 中魚體表面色澤的變化是由於這些色素 的降解進程不同,因此可作為漁獲物鮮 度的指標之一。
魚 肉 的 顏 色 取 決 於 紅 色 肌 肉(red muscles)的含量和肌肉中的血質蛋白類 (hemoproteins)―肌紅素(myoglobin)、血 紅 素 (hemoglobin) 與 細 胞 色 素 類 (cytochromes)的濃度及氧化性變化,鮭科 (salmonids)的肉色是來自類胡蘿蔔素的色 素類。
2. 香氣。很新鮮魚的氣味是讓人可 以接受的,類似於香瓜或海藻氣味,由 組織中存在濃度很低、氣味閾值低的許 多揮發性有機組成分所引起,主要是羰 基化合物類(carbonyl compounds)、醇類 (alcohols) 、 三 甲 胺 (trimethylamine;
TMA)、溴酚類(bromophenols)、甲基硫 酸 酯(methyl sulfate)和 其它的含硫化合 物。這些揮發性氣味化合物是脂質和含 氮組成分(nitrogenous components)在酵素 性降解的過程中,以及透過來自浮游生 物的食物鏈所蓄積的各種前驅物質所產 生的。之後,由於微生物腐敗進展,剛 捕獲魚類和貝類的最初令人愉快的香氣 經過長時間的貯藏,逐漸轉變為不新鮮 (stale)至最後成為腐敗(putrid)、不能接受 (offensive)、不良氣味(off-odor)。
3. 質地。魚類和甲殼類動物的肉具
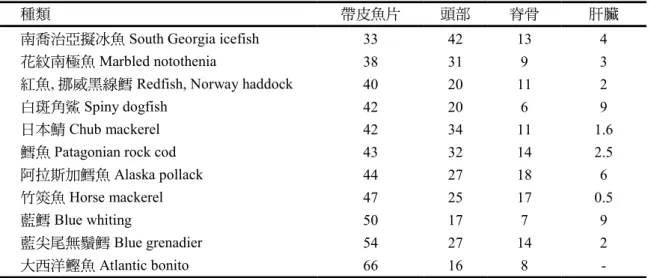
表1. 魚類主要魚體部位的一般比率(g/100 g)
種類 帶皮魚片 頭部 脊骨 肝臟
南喬治亞擬冰魚 South Georgia icefish 33 42 13 4
花紋南極魚Marbled notothenia 38 31 9 3
紅魚, 挪威黑線鱈 Redfish, Norway haddock 40 20 11 2
白斑角鯊 Spiny dogfish 42 20 6 9
日本鯖 Chub mackerel 42 34 11 1.6
鱈魚Patagonian rock cod 43 32 14 2.5
阿拉斯加鱈魚 Alaska pollack 44 27 18 6
竹筴魚 Horse mackerel 47 25 17 0.5
藍鱈 Blue whiting 50 17 7 9
藍尖尾無鬚鱈 Blue grenadier 54 27 14 2
大西洋鰹魚Atlantic bonito 66 16 8 -
來源:Sikorski and Kołakowski (2011)。
有纖維狀質地,類似於牛肉或禽肉,但 通常韌性(toughness)差甚多。另一方面,
魷魚胴肉(mantle)少有纖維狀,較類似橡 膠般的。然而這兩例,負責質地的肌肉 成分都是蛋白質的肌原纖維(myofibrils)和 膠原蛋白(collagens),很大程度上也取決 於 纖 維 ( f i b e r s ) 和 膠 原 蛋 白 片 層 (collagenous sheets)的立體排列。魚類和 頭足類動物(cephalopods)兩者的韌性程度 是物種依存性的(species-dependent),受 組織中的膠原蛋白和脂肪含量所影響。
在一些漁產品,質地主要受到脂質的影 響,而一些多脂魚如大西洋鯖魚(Atlantic mackerel)和波羅地海鯡魚(Baltic sprat),
如脂肪含量會因季節性而降至某限度以 下,就不適合加工成高品質的煙燻或罐 頭產品。低鹽漬鯡魚(maatjes)的柔軟質地 是由於未成熟多脂鯡魚組織中的脂質含 量高。測試者用手指觸壓生鮮肉所引起 的回應,通常被認為是反映魚的新鮮程
度的一項特徵性質。
(三) 可食用和不可食用部位的比例
魚類和海洋無脊椎動物的可食用器 官的比率,主要取決於種類、年齡、雄 雌、性腺成熟度和動物的營養狀況。理 解不可食用這用語可能有些模糊,因為 不同社會或種族人群也會將不同的魚體 部位視為適合於食用。雖然對大多數消 費者而言,常見的主要可食用部分是魚 片(fillet),尤其剝皮的,但世界上仍有許 多原住民群落也使用魚頭和內臟來製備 菜餚,甚者整尾小型魚鹽醃或燻煙後,
與皮、內臟和骨骼一起食用。
帶皮魚片約佔魚體總重的 30~65%。
表 1
顯示一些不同體型魚的數據,每種 魚的數值大多落在數個百分點的差異範 圍內,這可從表2
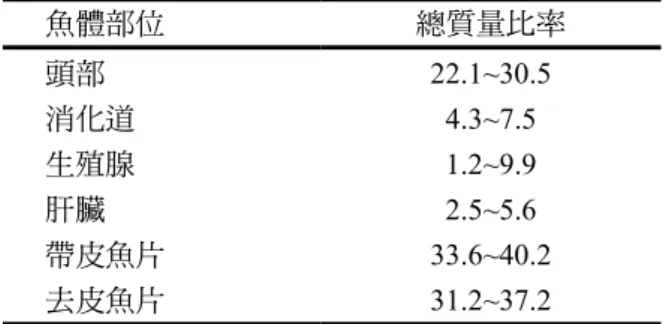
的數據得到說明。多 種小型魚由於在勞力密集作業中切割和 去皮的產能低,故經濟上常不可行,在 此情況,有幾種魚可改用自動機器同時表2. 波羅的海鱈魚主要魚體部位的比率(g/100 g)
魚體部位 總質量比率
頭部 22.1~30.5
消化道 4.3~7.5
生殖腺 1.2~9.9
肝臟 2.5~5.6
帶皮魚片 33.6~40.2
去皮魚片 31.2~37.2
來源:Cieglewicz (1954)。
去頭、去鰓和切除腹部皮瓣(belly flaps) (Knyszewski, 1996)。海洋無脊椎動物的 可食部位的製成率從12%至 72% (
表 3)。
(四) 主要組成分的含量
魚類、軟體類和貝類的可食用部分 富含蛋白質、非蛋白態含氮化合物(non- ptotein nitrogenous compounds)、脂質、
礦物質成分和維生素等,含量上主要取 決 於 種 類 、 捕 撈 季 節 和 魚 的 狀 況 。 水 分 、 含 氮 化 合 物 和 脂 質 合 計 占 魚 肉 的 98%左右(
表 4)。影響海產食品的脂質含
量及組成的因素,可參考Kołakowska et
al . (2003)
的詳細報告。醣類只占海洋動物肌肉的很小部分:肝醣量 0.6%至 1.5%
(濕重,剛捕獲魚),核苷酸(nucleotides)和
表3. 軟體類及甲殼類的可食用部位(一般含量,g/100 g)
軟體類 可食用部位 甲殼類 可食用部位
牡蠣 12 南極蝦 30
扇貝 18 蟹 40
貽貝 28 龍蝦 44
鮑魚 50 蝦 45
烏賊 Loligo spp 72 淡水螯蝦 16
來源:Sikorski and Kołakowski (2011)。
表4. 魚類及海洋無脊椎動物類肉中的主要組成分含量(一般範圍,g/100 g 濕重)
種類 水分 粗蛋白* 脂質
無鬚鱈Hake 74~81 16~19 0.8~4.3
藍鱈Blue whiting 79~82 14~17 0.5~3
狹鱈Walleye pollock 79~84 15~20 0.3~2.9
黑鮪Bluefin tuna 67.5~72.5 23.3~27.5 1.2~8
狹鱗庸鰈Pacific halibut 77~79 20~22 0.6~3.6
紅魚Redfish 72.5~78.5 20~26.5 0.6~8.4
粉紅鮭Pink salmon 69~78 17~20.5 2~9.5
白斑角鯊Spiny dogfish 67~77 18~24 1.4~13
秋刀魚Saury pike 65~72 22~25 1.5~11
眼斑雪冰魚Antarctic icefish 79~81 17~19 1.1~1.2
牡蠣 oyster 77~83 8~13 1.1~2.5
扇貝Scallop 74.5~85.5 15~20 0.3~1.6
*蛋白質和非蛋白態含氮化合物 (N × 6.25)。
來源:Sikorski and Kołakowski (2011)。
核酸(nucleic acids)中的戊糖(pentoses),
以及膠原蛋白和糖蛋白中的各種糖基部 分,特別在貝類。海產食品中的礦物質 組成分總量一般為濕重的 1.0~1.5%,但 隨著海洋生物群的營養鏈的生物蓄積,
一 些 元 素 可 能 以 異 常 高 的 濃 度 存 在 魚 類,尤其大型(高齡)捕食者及牡蠣組織。
另可參考
Szefer and Nriagu (2007)
專書中 關於食品的礦物質組成分之綜述。(五) 海產食品組成分的營養價值和生物活 性
1. 蛋白質。海產食品的可食用部分 中 , 肌 肉 纖 維 的 結 構 蛋 白 質(structural proteins)占粗蛋白(crude protein)總量的最 大比率,參與肌肉的收縮,營養價值最 高,且主要負責肉的質地、保水力(water- holding capacity) 、 成 膠 力 (gel-forming ability)、乳化性質(emulsifying properties) 和起泡力(forming ability)等。魚類肌肉比 起牛肉和豬肉通常其膠原蛋白含量低甚
多(
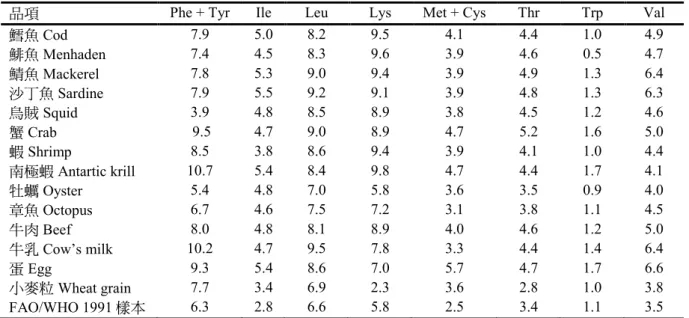
表 5),因此營養上作為必需胺基酸來
源 的 實 用 價 值 高 , 除 了 色 胺 酸
(tryptophan)濃度略低於 FAO/WHO 評分
值(
表 6)。魚肉片中不同蛋白質的胺基酸
組成是不同的,因此略微影響蛋白質的 生物價(biological value)。在人類,魚和 肉 蛋 白 質 的 消 化 率 約 94%,全麥穀物 (whole wheat grain)僅約 86%。此外由於 肉質較嫩,魚類、軟體類動物和甲殼類 動物通常烹飪製作時只輕度地加熱,這 可避免由於加熱變化而造成蛋白質生物 價的顯著降低。肌肉蛋白質的水溶性區 分含有原血紅素蛋白和大量的酵素及酵 素抑制劑(Haard and Simpson, 2000),它 們參與生物體的代謝過程,並在組織的 死後變化中發揮關鍵性的作用。
2. 脂質。海產食品可提供具有營養 價值的脂質,但可能也是提供低脂膳食 但美味組成分也豐富的來源,因不同魚 類和海洋無脊椎動物的脂肪含量變動歧 異 大( 見
表 4) 。 魚 類 脂 質 中 的 飽 和
(saturated)、單元不飽和(monoenoic)和多 元不飽和(polyenoic)脂肪酸含量分別介於 24~38%、21~42%和 26~45%。魚肉所含表5. 魚類、貝類和牛肉組織中膠原蛋白的一般含量
膠原蛋白
來源 g/100 g 粗蛋白 g/100 g 濕重
魚類肌肉,一般Fish muscles generally 1~12 0.2~2.3
常見硬骨魚肌肉Common teleost fish muscle 2~7 0.3~1.5
海鰻和鮑魚肌肉Conger eel and abalone muscle 5~12 1.0~2.0
軟骨魚肌肉Elasmobranch fish muscle 7~12 1.4~2.3
魚皮Fish skin 40~85 14~25
巴塔哥尼亞鎖管Loligo patagonica 3.0 0.36
阿根廷魷魚Illex argentinus 11 1.43
牛肉,最高等級Beef meat, highest grade 3.8 0.83
牛肉,最低等級Beef meat, lowest grade 60 12
來源:Sikorski and Kołakowski (2011)。
表6. 魚類肌肉和其它食物蛋白質的必需胺基酸含量
品項 Phe + Tyr Ile Leu Lys Met + Cys Thr Trp Val
鱈魚Cod 7.9 5.0 8.2 9.5 4.1 4.4 1.0 4.9
鯡魚Menhaden 7.4 4.5 8.3 9.6 3.9 4.6 0.5 4.7
鯖魚Mackerel 7.8 5.3 9.0 9.4 3.9 4.9 1.3 6.4
沙丁魚Sardine 7.9 5.5 9.2 9.1 3.9 4.8 1.3 6.3
烏賊Squid 3.9 4.8 8.5 8.9 3.8 4.5 1.2 4.6
蟹Crab 9.5 4.7 9.0 8.9 4.7 5.2 1.6 5.0
蝦Shrimp 8.5 3.8 8.6 9.4 3.9 4.1 1.0 4.4
南極蝦Antartic krill 10.7 5.4 8.4 9.8 4.7 4.4 1.7 4.1
牡蠣Oyster 5.4 4.8 7.0 5.8 3.6 3.5 0.9 4.0
章魚Octopus 6.7 4.6 7.5 7.2 3.1 3.8 1.1 4.5
牛肉 Beef 8.0 4.8 8.1 8.9 4.0 4.6 1.2 5.0
牛乳Cow’s milk 10.2 4.7 9.5 7.8 3.3 4.4 1.4 6.4
蛋Egg 9.3 5.4 8.6 7.0 5.7 4.7 1.7 6.6
小麥粒Wheat grain 7.7 3.4 6.9 2.3 3.6 2.8 1.0 3.8
FAO/WHO 1991 樣本 6.3 2.8 6.6 5.8 2.5 3.4 1.1 3.5
來源:Sikorski and Kołakowski (2011)。
註:苯丙胺酸 phenylalanine、酪胺酸 tyrosine、異白胺酸 isoleucine、白胺酸 leucine、離胺酸 lysine、甲 硫胺酸methionine、半胱胺酸 cysteine、蘇胺酸 threonine、色胺酸 tryptophan、纈胺酸 valine。
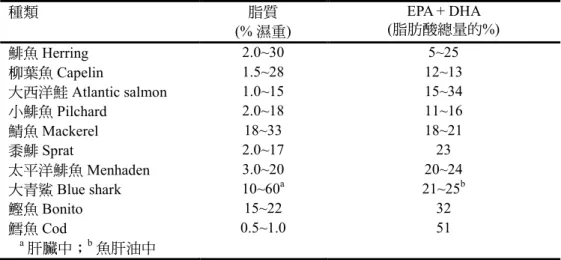
的和肝臟所提煉的海產魚油(marine fish oils)屬多元不飽和必需 n-3 脂肪酸的最豐 富 天 然 來 源 : 二 十 碳 五 烯 酸 (eicosapentaenoic acid; 20:5; EPA)、二十 二 碳 五烯 酸(docosapentaenoic acid; 22:5;
DPA)和二十二碳六烯酸(docosahexaenoic acid; 22:6; DHA) (
表 7)。多元不飽和 n-3
和 n-6 脂肪酸的生物活性與海產脂質在飲 食上的重要性,可參考Kołakowska et al.
(2003)
和Shahidi (2008)
報告。作為多元不 飽和脂肪酸來源之各種水產品的價值,不僅要考慮這些脂肪酸在脂肪酸總量中 的 比 率 , 還 應 考 慮 魚 料 理 中 的 脂 質 含 量。
3. 礦物質組成分。食物礦物質組成 分在膳食上的價值是基於它們參與人體 的建構和在許多生化過程中的作用。然 而,有幾種陽離子會促進食物中脂質的
氧化。礦物組成分的功能、以及巨量元 素 (macroelements) 和 微 量 元 素 (microelements)在各種水產品中含量,請 參考
Nabrzyski (2007)
報告。4. 維生素。魚類和海洋無脊椎動物 的肉通常比屠宰動物的肉含有更多的維 生素,魚油更是特別富含維生素 A、D 和 E。魚類脂質尤其魚肝油長久以來都作為 脂溶性維生素的豐富來源。海洋魚類魚 油中的維生素 A 群,主要是以視黃醇 (retinol 7,9,11,13-trans) 和 新 視 黃 醇 (neoretinol 13-cis 及 7,9,11-trans)構成的維 生 素 A1, 大 部 分 是 為 棕 櫚 酸 視 黃 酯 (retinyl palmitate)的酯化型式。一些海洋 魚類的肝臟中,有高達維生素 A 總活性 的 40%是來自維生素 A2,例如全反式 3- 脫氫視黃醇(all-trans dehydroretinol),在 淡水魚魚油中居優勢的成分。各種水產
表7. 魚類的 n-3 脂肪酸二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)的一般含量範圍
種類 脂質
(% 濕重) EPA + DHA
(脂肪酸總量的%)
鯡魚Herring 2.0~30 5~25
柳葉魚Capelin 1.5~28 12~13
大西洋鮭Atlantic salmon 1.0~15 15~34
小鯡魚Pilchard 2.0~18 11~16
鯖魚Mackerel 18~33 18~21
黍鯡Sprat 2.0~17 23
太平洋鯡魚Menhaden 3.0~20 20~24
大青鯊Blue shark 10~60a 21~25b
鰹魚Bonito 15~22 32
鱈魚Cod 0.5~1.0 51
a 肝臟中;b 魚肝油中
來源:Sikorski and Kołakowski (2011)。
品當作維生素 A 來源的價值,主要取決 於魚的種類和器官、產卵週期、氣候條 件、以及飼料可利用性。魚肉中維生素 A 的一般活性(依存於脂肪含量)從約 0.5 至約10 IU/g,魚肝油中從 200 至 250,000 IU/g。維生素 D 在魚類的含量與維生素 A 相似,肝臟中的含量也遠高於肉中。
由浮游植物合成的生育酚(tocopherols)存 在魚和海洋無脊椎動物的可食用部位,
含量約從0.002 至 3 mg/g 濕重。
魚和貝類的可食用部分是維生素 B 群的最豐富來源。通常,一份水產品菜 餚就含足夠滿足日常人類所需的維生素B 群。魚肉中的維生素 C 含量比起含量多 的漿果和水果約低 100 倍,但有些魚卵 和肝臟中維生素 C 濃度可能達數百 µg/g 溼重。水產品菜餚中水溶性維生素的含 量很大程度上取決於烹飪製備過程中汁 液和烹煮的流失。
5. 具有健康促進功能的組成分。海 產食品還含有許多的組成分,具有營養
功能以外的各種健康促進活性,可為消 費者帶來一些生理或醫學上的益處。各 種研究和開發的工作仍進行中,大多聚 焦於製備功能性食品成分或作為保健食 品,亦即尋找具有應用價值的各類生物 活性材料分離物,其中如膠原蛋白和明 膠 , 以 及 生 物 活 性 胜 肽 的 蛋 白 質 水 解 物 , 這 些 具 有 血 管 收 縮 素 Ⅰ 轉 換 酶 (angiotensin I converting enzyme)抑制活 性 、 鈣 吸 收 促 進 特 性 和 低 膽 固 醇 (hypocholesterolemic)、抗氧化及 α-葡萄 糖苷酶(α-glucosidase)抑制活性(Fujita and
Yoshikawa, 2008; Kim et al., 2008)。此
外,幾丁質和幾丁聚糖以及幾丁聚寡糖 可用作營養保健品,因具有作為膳食纖 維 補 充 劑 的 功 能 , 也 報 導 可 作 為 抗 菌 的、抗發炎的、抗氧化的和抗癌的製劑 (Kim et al., 2008; Rasmussen and Morrisey,2008)。魚骨也可利用於生產生物活性鈣
物質。(六) 過敏原、毒素和寄生蟲對海產食品品 質的影響
有些魚類和貝類會引起食物過敏。
IgE-依賴型食物過敏的全部病例中,約 90%可追溯及花生、堅果、牛奶、雞蛋、
大豆、魚類、甲殼類和小麥(Wróblewska,
2007)。鱈魚擬清蛋白(parvalbumin)已被
認定是過敏原 Gad c 1―這樣的蛋白質也 存在於其它種類的肌肉。此外,蝦類、淡水螯蝦(crayfish)和龍蝦的原肌凝蛋白 (tropomyosin)、從蟹肉淬取的其它一些蛋 白質也具有過敏性質。
魚體表面的腺體(glands)和魚貝類可 食用部分或內臟存在的各種毒素可能會 引起一些害處,從漁獲物處理者的皮膚 受到刺激疼痛一直至嚴重的病症,甚至 導致海鮮消費者的死亡。水產食品的毒 素所牽連的危害,可透過遵守地方和國 際有關漁業的規定而減低,例如某些資 源的開採季節加以限制,漁獲物的利用 例如透過資格認證人士去除危害成分。
Botana (2009)
專書介紹水產食品和淡水魚毒素的實況與知識。
水產食品中的幾種礦物質成分,尤 其汞、砷和鎘,如果存在濃度過高,可 能會對人體產生毒性作用。一些元素的 毒性也取決於其物種(Nabrzyski, 2007)。
(七) 生物和環境因素對海產食品資源加工 價值的影響
1. 水 鹽 度 。 透 過 細 胞 內 溶 質 (intracellular solutes) 如 游 離 胺 基 酸 (free amino acids)、無機離子(inorganic ions)和
四 級 胺 化 合 物 (quaternary ammonium compounds)等濃度的改變,魚類讓其細 胞體積去順應水的鹽度(salinity)。這些變 化 的 特 徵 取 決 於 肉 中 滲 透 活 性 化 合 物 (osmoactive compounds)的含量。紅肉魚 (dark-fleshed fish)如鮪魚、鯖魚和鯡魚的 肌 肉 富 含 游 離 胺 基 酸 , 主 要 是 組 胺 酸 (histidine) 、 丙 胺 酸 (alanine) 和 甘 胺 酸 (glycine)等。洄游性魚類骨骼肌中的非蛋 白 態 含 氮 化 合 物 庫(pool of non-protein nitrogenous compounds),主要包含組胺 酸、肌肽(carnosine)、鵝肌肽(anserine)、
肌 酸 (creatine) 和 肌 酸 酐 (creatinine) 等 (Suyama and Yoshizawa, 1973)。扁魚(flat fish)除外的白肉魚(white-fleshed fish)中,
肌酸為主要成分,組胺酸含量極少;在 軟骨魚類(elasmobranches),主要成分是 尿素(urea) (Sakaguchi and Simidu, 1964)。
肌肉中的非蛋白態含氮化合物總量是環 境鹽度影響魚類生物的一項好指標,但 對魚的加工適性的影響複雜。
魚 肉 的 重 要 滲 透 壓 調 節 成 分 (osmoregulative component)是氧化三甲胺 (trimethylamine oxide; TMAO),淡水魚的 含量低於 50 mg/100 g 或零值,最高的濃 度在海洋軟骨魚類(1000~1500 mg/100 g) 和鱈科(Gadidae; 300~1000 mg/100 g),其 它海水魚的肌肉含 100 至 250 mg/100 g 濕重。洄游性魚類從海洋攝食洄游至溯 河產卵,TMAO 含量逐漸減少至幾乎為 零值。
海水鹽度增高所引起肉中游離胺基
酸濃度的增加,對魚的營養價值並無明 顯的影響,因增加的對象以非必需胺基 酸為主,但可能會影響魚肉的滋味。
一些胺基酸尤其是組胺酸和離胺酸 (lysine),可透過降低蛋白質分子間的靜 電排斥力(electrostatic repulsion forces)而 促進肌肉蛋白質的冷凍變性,因其淨正 電荷分別為7.59 和 9.74,而肉的 pH 值通 常為 5.8~6.8。另一方面,中性胺基酸可 提高冷凍貯藏期間魚類蛋白質的穩定性 (Jiang and Lee, 1985)。
在冷藏魚,許多的革蘭氏陰性嗜冷 性(psychrotropic)細菌能夠利用 TMAO 作 為氧的來源,厭氧條件下死後的數日,
在 肉 中 即 成 為 優 勢 菌 。 在 冷 凍 魚 , TMAO 被內源性脫甲基酶(demethylase)作 用 而 產 生 的 甲 醛 可 能 參 與 蛋 白 質 交 聯 (cross-linking),尤其鱈科魚類,是其蛋白 質進行冷凍變性的反應之一。
環境的鹽度也會影響魚類脂質的組 成。多元不飽和 C:18 脂肪酸含量一直都 高,淡水魚的 C:20 和 C:22 脂肪酸含量低 於海水魚。在洄游魚類,脂肪酸組成在 產卵洄游過程中變化明顯(Love, 1970)。
2. 溫度。將魚移至溫度不同的環境 中會導致新陳代謝的改變,暖水域和冷 水 域 馴 化 魚 的 ATP (adenosine triphosphate; 腺苷三磷酸)消耗速率會有所 不同,但魚種類和貯藏溫度也是影響要 因。死後貯藏期間 ATP 和肝醣的減少速 率及乳酸蓄積的增加速率,暖水域馴化 鯉 魚 肌 肉 高 於 冷 水 域 馴 化 者(Abe and
Okuma, 1991) 。 將 冷 水 域 馴 化 帝 王 鮭
(chikoon salmon) (10.7oC 和 12.4oC)貯存於−1.2oC 至 6oC,回應溫度的升高所測定的 pH 值和 ATP/AMP 濃度變化,比馴化至 18.8oC 組高出 2.2 倍(Jerrett et al., 2000)。
馴化也影響膜磷脂質的脂肪酸組成,熱 帶 水 域 魚 的 粒 線 體 膜 (mitochondrial membrane)磷脂質的脂肪酸組成中,比起 冷水域魚含有更多的飽和脂肪酸(Irving
and Watson, 1976)。
3. 魚棲息深度。通常,每公克骨骼 肌中的酵素活性隨著魚的最小棲息深度 的增加而呈指數關係地下降,儘管種類 之間的酵素活性變動相當大。活躍的表 層 游 泳 型 魚 類 的 乳 酸 脫 氫 酶(lactate dehydrogenase) 和 丙 酮 酸 激 酶 (pyruvate kinase) 活 性 遠 高 於 深 層 生 存 的 坐 著 等 (deep-living sit-and-wait)魚類。在深層生 存魚種中,底棲性表層魚類(bentopelagic fishes)像鼠尾鱈(rattails)和銀鱈(sable fish) 的活性最高,這表明與大多數其它深層 魚類相比,這些種類的索餌行為的運動 性更大。深層生存魚類骨骼肌的蛋白質 含量顯著低於淺層生存(shallow-living)魚 的含量,而水分含量則略高,以平均而 言,乳酸脫氫酶活性約 5 倍高於在淺層 生存魚(Sullivan and Somero, 1980)。
4. 飼養狀況和性成熟度。密集飼養 期間,尤其產卵後,魚消化系統的活性 非 常 高 , 酵 素 易 擴 散 至 腹 部 皮 瓣 的 肌 肉,導致小型魚冷藏期間更多的自家消 化(autolysis) 而 易 使 腹 部 裂 開 , 尤 其 鯡
魚。先前曾提及的魚類肌肉主要組成分 含量的波動,這與生殖腺成熟的年周期 (annual cycle)有關。多脂魚種如鯡魚和鯖 魚的魚肉,脂肪含量從剛產卵後的 1~3%
(濕重)經密集飼養後增至 20~30%,這同 時 影 響 了 營 養 價 值 和 貯 藏 時 的 變 質 變 化。寡脂魚的性成熟度與蛋白質濃度的 降低、肌肉水分含量的增加有關,在產 卵後肌漿蛋白質(sarcoplasmic proteins)減 少 的 幅 度 大 於 肌 原 纖 維 蛋 白 質 (myofibrillar proteins),而產卵後的密集 飼養期間,以肌原纖維蛋白質的再生較 快速。雄性魚生殖腺的成熟會導致較多 的脂質下降,相對於雌性魚則以蛋白質 的減少更明顯。用於製造生殖腺的組織 蛋白(histones)的材料需求增加,這會造 成魚肌肉蛋白質的鹼性胺基酸的部分損 失。酵素水解後剩餘的其它胺基酸,例 如甘胺酸、丙胺酸和脯胺酸(proline)可利 用於合成膠原蛋白,使得剛產卵後它們 在魚肌肉中有較高的含量。
5. 漁撈技術和去頭(decaptiation)。決 定 漁 獲 物 的 性 質 之 最 重 要 因 素 為 漁 具 (fishing gear) 的 機 械 效 應 (mechanical effects)、捕獲至拖上船之間的時間和船 上的魚處置。根據漁具的不同,捕獲的 魚可能停留水中多至數日,如在刺網中 會受緊迫或耗竭(exhausted)、或沿海底拖 網數小時而使漁獲物損傷與受到腸道內 容物及底泥的污染。定置網捕獲的魚在 定置網內可自由游動數小時,而手釣捕 獲的魚通常幾分鐘內就拉上船,這些不
同的條件對肌肉中代謝過程的速率都會 影響。
通常,捕撈後迅速宰殺和放血可延 長僵直(rigor)發生的時間,因而延長冷藏 魚的貯藏壽命,對一次網捕獲大量的小 型 魚 實 際 上 是 不 可 行 , 但 建 議 用 於 底 棲、大型表層和養殖魚類。已有各種處 理技術採用,例如棒打頭部、頭蓋穿刺 (cranial spiking)、電擊(electrocution)和暴 露於二氧化碳。根據魚的種類和狀況,
各種處理對於最終下降後的肌肉 pH、以 及僵直發生的延遲和貯藏壽命的延長等 的影響都有所不同。高品質貯藏壽命的 延長可能從數小時至數日不等,這通常 適用於冷水域魚種,即捕撈上船時不會 已是耗竭狀態。但一些溫水域魚類,若 很快就穿刺和冷卻反而僵直提早發生,
以 及 可 能 使 肉 質 產 生 冷 休 克 僵 硬(cold- shockening stiffening)。
三 、 捕 撈 後 的 變 化 (Sikorski and
Kołakowski, 2011)
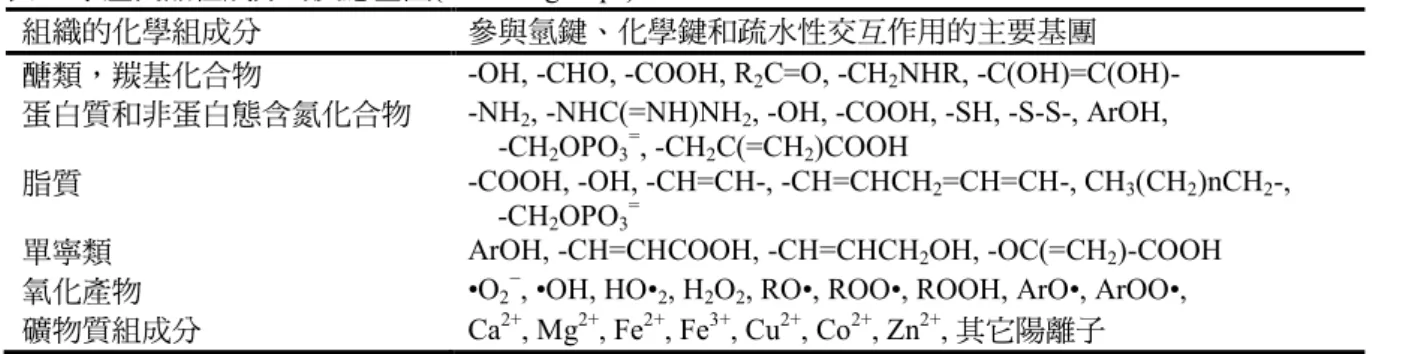
魚貯藏時,其組織的組成分易生變 化,此乃存在各種化學活性基團且當處 於有利的條件下,可參與許多的反應(
表 8)。再者於窒息(asphyxia)後,在活生物
體中原本活性的生物控制系統也停止,故而受到各種的內源性酵素(endogenous enzymes)的水解作用,隨著捕撈後的時間 增長,微生物酵素也參與分解組織。
剛 離 水 的 魚 肉 狀 態 取 決 於 漁 撈 技 術 。 如 果 魚 鉤 上 或 漁 網 內 魚 掙 扎 很 激 烈,隨後迅速宰殺和放血,肌肉中能量
供 應 化 合 物 的 儲 備 已 經 耗 盡 而 無 法 回 復 , 這結 果 是 肉的 高能 量 磷酸 酯 (high- energy phosphates)和肝醣含量很低。另一 方面,充分靜止(rested)的魚立即宰殺,
魚肉中這些化合物的含量仍多,這樣的 魚若立即冷藏,可看出其特徵的現象變 化過程(表 9)是不同的。
(一) 能量儲備耗盡的影響
核苷酸的降解:ATP → ADP + Pi → AMP + Pi → IMP + NH3 → Ino + Pi →Hx + ribose 或 ribose-1-P,其中 ATP (腺苷三 磷酸)、ADP (adenosine diphosphate)為腺 苷二磷酸、IMP (inosine mononucleotide) 為腺苷單磷酸、Ino (inosine)為肌苷、Hx (hypoxanthine)為次黃嘌呤、Pi (inorganic phosphate)為無機磷酸鹽。由於內源性酵 素的活性,醣類在死後早期即出現(Gill,
2000),IMP 逐漸減少,很大程度上 IMP
減少可能造成新鮮魚所需鮮美風味的損 失。ATP 及其降解產物的濃度可作為一 項測試,表示魚類鮮度從早期開始的下 降情形,即 K 值變化:K 值(%) = 100 (Hx + Ino)/ATP + ADP + AMP + IMP + Ino + Hx,反映魚類鮮度的一項指標。一 般而言,K 值<10%表示初始鮮度(prime freshness),K = 60%時產品已不適合消 費。然而由於冷藏期間不同種類和狀態 的魚,其核苷酸的異化作用(catabolism) 速率也有所差異,故這些數值必須與感 官分析的結果一起判斷。死後僵直(rigor mortis)現象是在肌漿 (sarcoplasm)中鈣離子(Ca2+)存在下,肌肉
纖 維 的 肌 節 (sarcomeres) 中 肌 凝 蛋 白 (myosin) 和 肌 動 蛋 白 (actin) 微 原 纖 維 (microfibrils)之間相互作用而引起。對於 魚類有其重要的實質意義,即魚體變僵 硬且常變形,因此不適於採用機械剖取 魚片。另外,剛捕獲就立即剖取魚片,
可能造成魚片長度的收縮最大達 40%。
然而,只要保持在僵直狀態,漁獲物就 處 於 最 佳 鮮 度 的 狀 態 。 在 組 織 和 魚 體 中,微生物菌群不斷成長的同時,也有 更多的酵素變化發生而最終僵硬消失,
新鮮度逐漸降低,最後變成不新鮮的和 腐敗的,如表 9 所示。雖然冷藏牛肉在 死後僵直階段之後可能費時數週達到所 要的嫩化程度,海產食品通過僵直階段 之後,通常數日內吸引人消費的屬性都 消失。
(二) 含氮化合物的變化
魚肉蛋白質的水解(hydrolysis)最初是 由 內 源 性 肌 肉 蛋 白 酶 (proteinases) 催 化 (Kołodziejska and Sikorski, 1995; 1997),
而未去內臟的小型魚受到腎臟和肝臟的 組織蛋白酶(cathepsins)以及消化道內的消 化酵素(digestive enzymes)催化。貯藏時 間延長後,微生物酵素的作用也參與貢 獻蛋白質的變化。通常,蛋白質和肌肉 結構的部分斷片化(partial fragmentation) 導致肉流變性質緩慢地變質,如
表 9
所 示。製造各種鹽漬和發酵海產食品時,蛋白質水解的變化(proteolytic changes)扮 演 關 鍵 作 用 (Kołakowski and Sikorski,
2000)。
表8. 水產食品組成分的反應基團(reactive groups)
組織的化學組成分 參與氫鍵、化學鍵和疏水性交互作用的主要基團
醣類,羰基化合物 -OH, -CHO, -COOH, R2C=O, -CH2NHR, -C(OH)=C(OH)- 蛋白質和非蛋白態含氮化合物 -NH2, -NHC(=NH)NH2, -OH, -COOH, -SH, -S-S-, ArOH,
-CH2OPO3=, -CH2C(=CH2)COOH
脂質 -COOH, -OH, -CH=CH-, -CH=CHCH2=CH=CH-, CH3(CH2)nCH2-, -CH2OPO3=
單寧類 ArOH, -CH=CHCOOH, -CH=CHCH2OH, -OC(=CH2)-COOH
氧化產物 •O2−, •OH, HO•2, H2O2, RO•, ROO•, ROOH, ArO•, ArOO•, 礦物質組成分 Ca2+, Mg2+, Fe2+, Fe3+, Cu2+, Co2+, Zn2+, 其它陽離子
表9. 在肌肉中的變化過程與其對捕獲魚的影響
捕撈後的 階段
高能量磷酸酯、
醣類
蛋白質和非蛋白態 含氮化合物
脂質 感官的影響
在 漁 網 、 魚 鉤 或 船
板上掙扎;
窒息
肌 酸 磷 酸 的 去 磷 酸 化 、ATP 降 解 為 ADP 、 AMP 、 IMP、Ino、Hx、 磷
酸根、 氨和核醣,
肝醣降解為乳酸
氧合血質蛋白損失 變化不明顯 魚 皮 顏 色 鮮 明 , 外
觀 亮 麗 , 鰓 鮮 紅 , 新 鮮 海 藻 般 香 氣 , 魚體有彈性
最初鮮度 貯 存 物 受 內 源 性 酵
素 催 化 而 進 一 步 消
耗,乳酸蓄積,pH
下降
在 軟 骨 鯊 目 (Selachii) 尿 素 水 解 , 一 些 蛋 白 酶 活 化
輕 度 酵 素 催 化 氧 化 生 成 揮 發 性 物 質 醇 類和羰基物類
魚 皮 顏 色 鮮 明 , 外 觀 亮 麗 , 鰓 鮮 紅 , 海 藻 般 香 氣 , 在 軟 骨 鯊 目 有 輕 微 的 氨 味,魚體有彈性
死後僵直 缺少 ATP 而導致從
肌漿網滲漏出Ca2+
肌 肉 收 縮 系 統 的 交 互作用
內 源 性 酵 素 催 化 的 初 期 水 解 和 氧 化 性 變化
魚 皮 顏 色 鮮 明 , 外 觀 亮 麗 , 鰓 鮮 紅 , 海 藻 般 香 氣 , 在 軟 骨 鯊 目 可 感 知 氨 味,魚體堅硬 最 初 鮮 度
下降
降 解 產 物 被 微 生 物
利用和反應,pH 上
升
結 構 蛋 白 質 被 鈣 蛋 白 酶 、 組 織 蛋 白 酶 及 蛋 白 酶 體 (proteasomes)等部分 水 解 , 非 蛋 白 態 含 氮化合物變化
內 源 性 和 細 菌 酵 素 催化的水解和氧化
魚 皮 變 色 失 光 澤 , 略 帶 乳 白 色 粘 液 , 鰓 色 變 淡 , 海 藻 香 氣 消 失 , 略 帶 魚 腥 味,魚體可彎曲
細 菌 成 長 快速
降 解 產 物 被 微 生 物 利用,pH 上升
酵 素 性 和 細 菌 性 降 解 , 揮 發 性 氣 味 化 合物形成
更 持 續 的 水 解 , 氣 味性降解產物生成
變 色 魚 皮 , 鰓 灰 褐 色 並 略 帶 乳 白 色 的 粘 液 , 魚 腥 味 和 氨 不 良 氣 味 , 魚 體 軟 化
細 菌 性 腐 敗
許 多 蛋 白 質 降 解 為 胜 肽 及 低 分 子 量 化 合 物 , 揮 發 性 氣 味 產物蓄積
更 持 續 的 水 解 , 有 氣 味 性 降 解 產 物 生 成
皮 膚 和 眼 睛 有 粘 稠 的 變 色 粘 液 , 肉 變 黃 , 腐 敗 氣 味 , 魚 體變形
內源性和微生物酵素所催化非蛋白 態含氮化合物的降解(degradation),顯著 影響海產食品的氣味。最典型的是氧化 三 甲 胺(TMAO)還原 為 三甲胺(TMA):
(CH3)3NO + NADH + H+ → (CH3)3N + NAD+ + H2O , 以 及 被 去 甲 基 化 (demethylation) 成 為 二 甲 胺 (dimethylamine; DMA) 及 甲 醛 (formaldehyde):(CH3)3NO → (CH3)2N + HCHO。
鯊魚(sharks)和魟魚(rays)肉在捕撈後 不久就聞到一股氨氣味,乃內源性尿素 酶(urease)催化尿素的水解:(CH3)3NO + H2O → 2NH3 + CO2。
不同揮發性化合物是經由胺基酸的 降 解 而 形 成 , 包 括 胺 類(amines)、醛類 (aldehydes)、硫化物類(sulfides)、硫醇類 (mercaptans)和短鏈脂肪酸類(short-chain fatty acids),具有明顯另人不快的氣味。
組胺酸、離胺酸、麩胺醯胺(glutamine)和 精胺酸(arginine)是一些非揮發生物二胺 類(biogenic diamines)的前驅物質,主要 由微生物酵素的作用所生成。龍蝦、蝦 和蟹中存在的內源性多酚氧化酶複合物 (polyphenoloxidase complex) 催 化 酪 胺 酸 (tyrosine)的氧化而生成二羥基苯丙胺酸 (dihydroxyphenylalnine; DOPA)和 DOPA 醌(DOPA chinone),並繼續聚合形成甲殼 類動物表層的高分子黑色色素,蛋白質 中的半胱胺酸(cysteine)和離胺酸也涉入 聚合反應。
(三) 脂質水解和氧化
魚 類 脂 質 的 水 解 變 化(hydrolytic changes)是內源性脂酶(lipases)和磷脂酶 (phospholipases)所催化,捕撈後很快即開 始。磷脂質(phospholipids)最易被水解,
初期這些反應對海產食品的感官特性影 響 小 , 在 脂 質 氧 化 的 初 期 階 段 也 是 如 此。之後,在冷藏期間透過微生物作用 所 生 成 脂 質 的 水 解 產 物 和 氧 化 二 次 產 物,使不新鮮海產食品引起不良氣味的 化合物持續增加。有關海產食品貯藏中 的脂質變化,可參考
Kołakowska (2010)
報告。(四) 影響變化速率的因素
魚類和貝 類 收穫 後變 化(postharvest changes)的速率,取決於種類特徵(species characteristics)和動物的狀態、來自捕撈 技術的緊迫、捕撈船上的處置條件、放 血和去內臟,並高度受到漁獲迅速冷卻 的 效 率 的 影 響 。 所 有 造 成 能 量 儲 備 (energy reserves)耗盡的條件都會縮短僵直 發生的時間和僵直狀態的持續時間,從 而也縮短了貯藏壽命。捕撈後立即宰殺 或腦穿刺可延遲僵直的發生。處理過程 中產生的瘀傷最易使微生物侵入,暴露 陽光下有害皮膚呈色。於冰中迅速冷卻 至 0℃附近的溫度可降低所有生化的和微 生物的變質過程的速率,並阻止有害化 合 物 例 如 組 織 胺(histamine)或細菌毒素 (bacterial toxins)的產生。
四、海產食品腐敗的起因
海產食品其初始狀態的任何變化都 會造成氣味、滋味、外觀和質地等的接 受程度的下降,這稱為腐敗(spoilage),
可歸因於海產食品中的酵素、化學或微 生 物 活 性 所 致(Ghaly et al., 2010) 。
Adebowale et al. (2008)
指出死後僵直是魚 類肌肉在死後立即發生的生化變化,導 致肌肉的揉屈性(flexibility)喪失。內源性 蛋白酶和脂酶活性、腐敗微生物和脂質 氧化被認為是造成死後僵直後的貯藏期 間腐敗或變質的原因(Ghaly et al, 2010;Grant et al., 2003)。
新鮮魚易腐敗,貯藏壽命短。一些 因素會影響魚類作為加工原料或消費食 品的品質,新鮮魚的腐敗是微生物活性 所主導引起的,儘管不全如此,這取決 於微生物種類。由於不當處置的物理性 損傷可能影響魚體的完整性,導致品質 降低或整條魚損失;在後續的處理操作 時,也會更易使得新鮮魚加速水分流失 及 自 家 消 化 活 性 、 條 件 性 感 染 (opportunistic infection)與氧化反應。魚類 脂質的反應會導致品質變質,特別在冷 凍魚和乾燥魚。不論原因如何,腐敗所 造成的感官變化,很大程度上決定了消 費者對產品品質的感知。
( 一 ) 自 家 消 化 的 變 化 (Sikorski and
Kołakowski, 2011)
很早就知道魚類的腐敗至少有兩種 類型:細菌的和酵素的。酵素活性造成 魚類死後最初數日所發生自家消化的變 化。
Uchiyama and Ehira (1974)
指出對於鱈魚(cod)和黃鰭鮪(yellowfin tuna),酵素 性變化先行影響鮮度,而無關由細菌引 起的變化。在其它種類如魷魚和鯡魚,
酵素性變化先行並主導冷藏魚的腐敗樣 式,至於其餘魚類整體品質的損失,除 了微生物介入的影響,自家消化的貢獻 程度不一。
1. 醣解:腺苷三磷酸(ATP)的降解。
魚死後的醣解導致乳酸的蓄積,使肌肉 的 pH 下降,並促使肌肉蛋白質上的淨表 面 電 荷 數 減 少 , 引 起 部 分 變 性 (denaturation)及失去部分的保水力。當魚 死亡,死後僵直開始,肌肉中 ATP 含量 降至約 1.0 μmoles/g,這最終導致肌肉縮 短,變成硬直(stiff)不能伸展。死後僵直 中的魚通常不能夠切割魚片或加工,因 魚體太硬而操作困難,且經常又扭曲變 形,機械操作更不可行。死後僵直解除 後,肌肉組織隨之變軟(鬆弛),這與自家 消化的變化併行一致。魚死後而被探討 的最早自家消化變化之一的是 ATP 相關 化合物的降解,也就是 K 值(前述),K 值 或鮮度指標主要是基於魚類貯藏期間肌 肉中發生自家消化的變化而給予相對的 鮮度評級(rating)。然而,採用感官品評 技術並以訓練型品評員判定的貯藏壽命 在根本未達最後的期限之前,有些魚類 如大西洋鱈魚(Atlantic cod)就已達到最高 點 K 值,因此,K 值不適合作為所有海 洋 有 鰭 魚 類 的 鮮 度 指 標 。 從 日 本 魷 (Todarodes pacificus)的研究,ATP 及其相 關化合物的降解而增加的核糖含量,是
煮 熟 、 乾 燥 和 調 味 魷 魚 產 品 褐 變 的 原 因 , 因 此 保 持 魷 魚 原 料 的 鮮 度 很 重 要 (Omura et al., 2007)。
2. 自 家 消化 涉及 的蛋 白 質水 解 酵 素。從魚肉中已分離出多種蛋白酶,這 些蛋白質的水解斷裂所造成的影響常和 組織的軟化(softening)有關。或許最受注 目的舉例之一為自家消化的蛋白質水解 (autolytic proteolysis),在表層(多脂魚)魚 種例如鯡魚和柳葉魚(capelin)發生的腹部 爆裂(belly-bursting),這類型的組織軟化 主要發生在夏季月份,當大量攝食尤其 是橈足類(copepods)和磷蝦(euphausiids)構 成的「紅色餌料red feed」。
3. 組織蛋白酶(cathepsins)。屬酸性的 蛋白酶,封存於稱為溶酶體(lysosomes)的 微小胞器中,組織蛋白酶 L 導致肌肉的 軟 化 ,主 要 作用發 生 於 冷 凍/解凍的組 織。
4. 鈣蛋白酶(calpains)。是細胞內的 內切胜肽酶(intracellular endopeptidases),
鈣蛋白酶主要負責肉的死後自家消化,
透 過 消 化 肌 原 纖 維 的 z-線(z-line)蛋白 質。大多數鈣蛋白酶的活性位於生理 pH 值範圍,因此可合理地懷疑冷藏期間對 肌肉軟化的重要性。適應較冷環境溫度 的魚類比熱帶水域者更易受到鈣蛋白酶 自家消化的影響。
5. 膠原蛋白酶(collagenases)。冷藏鱒 魚(trout)肌肉的質地以儀器測量,結果顯 示質地的劣化隨著膠原蛋白 V 型的溶解 量而增加,可能由於自家消化的膠原蛋
白酶的作用(Sato et al., 1991)。這些酵素 很可能在長時間冰藏的魚或高溫下短期 儲存時,肌節(myotome)裂開或崩解的起 因。冷藏大蝦(prawn)由於組織的軟化而 貯藏壽命較短,也是膠原蛋白酶所引起 (Nip et al., 1985),膠原蛋白酶存在大蝦的 肝胰臟(消化器官)。
6. 冷凍貯藏期間自家消化的變化。
氧化三甲胺(TMAO,存在許多海洋硬骨 魚的滲透調節化合物)的還原(reduction)通 常是細菌的作用,但有些魚種的肌肉組 織 中 存 在 能 將 TMAO 分 解 成 二 甲 胺 (DMA)和甲醛的酵素,甲醛會引起肌肉 蛋白質的交聯,使肌肉韌化(toughening) 與保水力下降,該酵素稱為氧化三甲胺 酶(TMAO-ase) 或 氧 化 三 甲 胺 脫 甲 基 酶 (TMAO demethylase),最常見於鱈科魚類 (gadoid fishes) (鱈魚家族 cod family)。大 多數的氧化三甲胺脫甲基酶都是膜結合 型,當因冷凍或加入界面劑的溶解而使 組織細胞膜破壞時,活性就變成最高。
在深色(紅色)肌肉的活性高於在白色肌 肉,其它如魚腎臟、脾臟和膽囊等組織 中的酵素量特別高。因此,冷凍貯藏魚 要完全避免韌化,魚漿(minced fish)中一 定要完全不含器官組織,例如鱈魚科的 腎臟。
防止冷凍魚的自家消化產生甲醛的 最實用方法是−30°C 以下儲存魚類,盡量 減少溫度的波動,避免不當處置或凍結 前魚體受擠壓。影響新鮮魚和冷凍魚的 可食用性之自家消化的變化整理列於表
10
。通常,造成自家消化之最重要單一 因素是肌肉細胞的物理性破壞,許多自 家 消 化 酵 素 原 位 於 分 散 的 膜 結 合 處 (membrane-bound packages) 而 與 基 質 隔 離,當受到物理性的不當作業會破裂釋 出 而 接 觸 基 質 。 即 使 細 菌 汙 染 量 相 當 低,冰或其它方式造成魚體的損傷也會 嚴重影響可食用性和魚片製成率。(二) 酵素性變質(enzymatic deterioration) (Olatunde and Benjakul, 2018b)
受內源性酵素降解蛋白質的過程即 自家消化,在死後僵直結束後就立即開 始,這過程為細菌生長創造了有利的環 境(Ghaly et al., 2010)。海產食品感官性質 的改變可歸因於蛋白酶和脂酶(Engvang
and Nielsen, 2001),即使冷藏和冷凍貯
藏,海產食品的自家消化仍以緩慢的速 率進行(FAO, 2005),但若貯藏不當,透 過 內 源 性 和 微 生 物 的 蛋 白 酶 介 導 的 過程 , 蛋 白 質 迅 速 降 解 。 胰 凝 乳 蛋 白 酶 (chymotrypsin)、組織蛋白酶、胰蛋白酶 (trypsin)、脂酶和磷脂酶存在水產品的肝 胰 臟(hepatopancreas)、脾臟(spleen)和幽 門盲腸(pyloric ceca),胃蛋白酶(pepsin)則 位於胃(Odedeyi and Fagbenro, 2010)。常 發生的魚腹部爆裂,就是魚內臟酵素作 用而使蛋白質快速分解。海產食品貯藏 和加工過程中,和甲醛產生一起的質地 變化(肉韌化)也是酵素活性所致(Ghaly et
al ., 2010)。有些魚類存在氧化三甲胺脫甲
基酶將 TMAO 去甲基化而生成二甲胺 (DMA) 和 甲 醛 , 誘 發 甲 醛 的 形 成 (Goncalves and de Oliveira, 2016;
Leelapongwattana et al., 2005)。甲醛經由
亞甲基橋接(methylene bridging)而與蛋白 質交聯,造成魚類肌肉的韌化且保水力 降低(Immaculate and Jamila, 2018)。蝦類 的黑變(黑色色素形成)也是蝦中的酪胺酸表10. 冷藏魚中自家消化的變化整理
酵素 基質 變化 預防/抑制
醣解酵素 肝醣 肌肉中,乳酸生成、組織pH
下降、保水力降低;
高溫僵直可能造成肉質裂開
可能的話,應讓魚在0oC 附
近通過僵直階段 要避免僵直前的緊迫
自家消化酵素 (涉及核
苷酸降解) ATP, ADP,
AMP, IMP 新鮮魚風味消失、次黃嘌呤
帶來苦味(較後階段)
同上述
處置不當或擠壓會加速裂解
組織蛋白酶 蛋白質
胜肽
組織軟化、造成加工上困難 或不可行
貯藏和卸貨過程中處置粗暴
胰凝乳蛋白酶、胰蛋白 酶、羧基肽酶
蛋白質 胜肽
在表層魚類(腹部爆裂)內臟腔
的自家消化
凍結/解凍重複或長時間冷藏
產生的問題
鈣蛋白酶 肌原纖維蛋白
質
軟化、甲殼類脫殼引起的軟 化
移除鈣因而預防活化?
膠原蛋白酶 結締組織 魚片裂開、軟化 結締組織降解與冷藏時間及
溫度有關
氧化三甲胺脫甲基酶 TMAO 冷凍鱈魚科魚類中甲醛引起
的肉質韌化
魚貯藏在溫度≦ −30°C
物理性損傷和冷凍/解凍促進
甲醛引起的肉質韌化
酶(tyrosinase) 或 多 酚 氧 化 酶 (polyphenol oxidase)作用所致(Sae-Leaw et al., 2017)。
蛋白質水解的產物(游離胺基酸和胜 肽)可作為微生物成長的營養物質,導致 於腐敗以及生物胺(biogenic amines)的形 成(Fraser and Sumar, 1998; Ghaly et al.,
2010)。溫度和 pH 是影響蛋白酶活性的
因素,大多數蛋白酶的最適 pH 值落在鹼 性和中性範圍內(Ghaly et al., 2010)。透過 內 源 性 或 微 生 物 的 酵 素 作 用 和 代 謝 , TMAO 被還原為 TMA 和其它鹼性揮發物 而 提 高 貯 藏 海 產 食 品 的 pH (Leelapongwattana et al., 2005; 2008)。海產食品中的脂肪可被脂酶或磷脂 酶水解(Aryee et al., 2007; Kaneniwa et al.,
2004)。單酸甘油酯、雙酸甘油酯和三酸
甘油酯(mono-, di- and triglycerides)的水 解,是水存在時由三醯基甘油醯基水解 酶 (triacylglycerol acylhydrolase; EC 3.1.1.3)所誘導(Fernandes, 2016)。海產食 品中釋出的游離脂肪酸易氧化而生成不 良氣味,特別是魚腥味(fishy odor),魚腥 味與醛類有關,主要來自多元不飽和醛 (polyunsaturated aldehydes) (Maqsood etal ., 2014)。放血的魚其魚腥味強度比未放
血者弱(Maqsood et al., 2014)。此外,主 要 存 在 於 鰓 或 皮 膚 的 脂 氧 合 酶 (lipoxygenase)也會誘導貯藏魚的氧化,
特 別 貯 藏 時 間 增 長 時(Sae-leaw et al.,
2013)。
(三) 微生物的作用(Loreal and Dalgaard,
2014; Olatunde and Benjakul, 2018b)
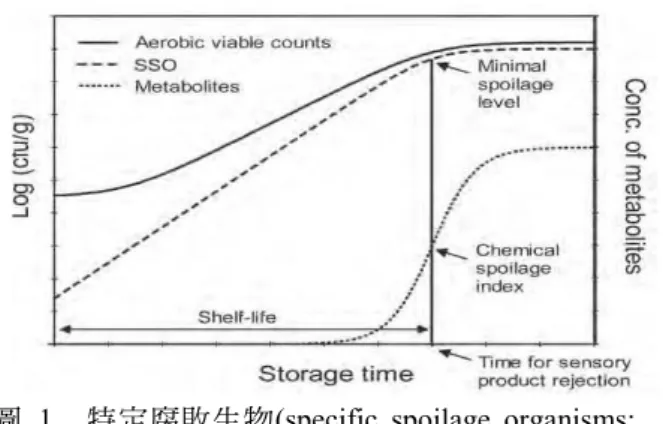
1. 微生物變化。在有鰭魚類、甲殼類 動物和軟體貝類等的原料和產品中,微 生物的存在、生長和活性都會影響其腐 敗和貯藏壽命,為避免造成缺陷和貯藏 壽命短的問題,相關微生物的評估和管 理就非常重要。水產品中的眾多微生物 之中,僅一些對腐敗是重要的,貯藏期 間 常 觀 察 到 稱 為 特 定 腐 敗 生 物(specific spoilage organisms; SSO)概念之微生物的 生長和活性模式。在剛加工、生鮮或輕 度保藏(lightly preserved)魚肉,通常 SSO 存 在 濃 度 非 常 低 , 僅 及 總 微 生 物 菌 群 (total microflora)的一小部分。但其後的貯 藏中,SSO 比其餘微生物菌群的生長更 快,產生具有不良風味的代謝物,最後 導致產品被拒受(
圖 1)。SSO 通常由單一
或一些微生物菌種組成,在感官評定達 腐敗的該時間點的產品中,所發現的微 生物菌群通常包括數種微生物群,而當 感官評定的腐敗程度已達明顯階段時,SSO 存 在 的 一 般 濃 度 約 107 cell/g 或 colony/g。
圖 1. 特定腐敗生物(specific spoilage organisms;
SSO)概念―新鮮魚貯藏期間的總生菌數、
SSOs 和透過 SSOs 所產生代謝物的典型變 化。
2. 棲息地不同水生動物的微生物菌 群 。 捕 撈 或 漁 獲 水 生 動 物 的 微 生 物 菌 群,大程度上取決於所生活水中的微生 物菌群(microflora)。微生物都存在於外部 表面(如魚皮、鰓部和腸道),健康水生動 物的肌肉組織是無菌的。可培養微生物 的 濃 度 是 變 動 的 , 一 般 而 言 , 皮 膚 102~105 cfu/cm2,鰓部 103~107 cfu/g,腸 道 10 至>108 cfu/g,腸道微生物的濃度變 動大,與水生動物的食物攝取有關。棲 息溫度較高,在水中和水生動物中通常 可培養的微生物濃度較高,但水的鹽度 對微生物的總濃度幾乎沒有影響(Cahill,
1990; Horsley, 1977; Liston, 1980;
Okuzumi and Awano, 1983; Yoguchi et al., 1990; Yoshimizu and Kimura, 1976)。至於
捕撈方法的影響,拖網捕獲的有鰭魚類 比起延繩釣捕獲的同類魚,魚皮和鰓部 上 的 微 生 物 濃 度 可 高 達 10~100 倍 (Shewan, 1961)。剛捕獲或漁獲的有鰭魚類、甲殼類 和貝類在魚皮、外殼和鰓部存在的微生 物屬或群都已被廣泛探討。主要的革蘭 氏 陰 性 菌 群 為 : (1) 不 動 桿 菌 屬 (Acinetobacter)和莫拉氏菌屬 /冷桿菌屬 (Moraxella/Psychrobacter) , (2) 假 單胞 菌 屬 (Pseudomonas) 和 希 瓦 氏 菌 屬 (Shewanella) , (3) 黃 桿 菌 屬 (Flavobacterium) 和 嗜 胞 菌 屬 (Cytophaga),(4)弧菌屬(Vibrio)和發光桿 菌 屬(Photobacterium) , (5) 氣 單 胞 菌 屬 (Aeromonas) , (6) 腸 桿 菌 科
(Enterobacteriaceae)。優勢的革蘭氏陽性 菌群為球菌類(cocci),主要是微球菌屬 (Micrococcus)、棒狀桿菌屬(coryneforms) 和 包 含 桿 菌 屬 (Bacillus) 、 梭 菌 屬 (Clostridium)和乳酸菌之桿菌(rods)。綜合 許多研究數據,有關水溫和鹽度的影響 或如底層(dermersal)或表層(pelagic)動物 類型對於微生物的屬及/或群的分佈百分 比的影響很少。但在淡水中的動物,鈉 需要種類的弧菌屬和發光桿菌屬極罕見 存在,反而氣單胞菌屬和腸桿菌科相對 更 重 要 。 此 外 , 黃 桿 菌 屬- 嗜 胞 菌 屬 (Flavobacterium-Cytophaga)菌群似在海洋 動物中居主導者較少見,在熱帶水域的 桿菌屬、微球菌屬和腸桿菌科所占的百 分比往往高於在溫帶水域者。從棲息地 不同水生動物的微生物菌群之間的差異 並不明顯,這有可能是由於採用了只根 據 項 目 不 多 的 表 型 特 徵 (phenotypic characteristics)而過於簡單的菌種鑑識方 案所致(Cahill, 1990; Gram and Huss, 2000;
Horsley, 1977; Karunasagar and Karunasagar, 1991; Liston, 1980; Shewan, 1962)。
由於水生動物腸道中的微生物濃度 高,加工過程中產品的污染就顯得重要 且難以避免。消化道發達的海洋魚類含 有特定的腸道微生物菌群,由包括螢光 發光菌(Photobacterium phosphoreum)的海 洋弧菌屬(vibrios)組成,而簡單消化道的 魚類如未成熟魚則含有更複雜的腸道菌 群,這反映出水中和餌料中的微生物菌
群(Sera and Ishida, 1972;
Yoshimizu and Kimura, 1976)。一些研究數據顯示,弧菌
屬/發光桿菌屬、假單胞菌屬和腸桿菌科 在海洋魚類的腸道菌群中占主導地位,反之腸桿菌科、氣單胞菌屬和假單胞菌 屬在淡水種類的腸內含量中占多數,但 是,不動桿菌屬/莫拉氏菌屬、乳酸菌、
酵母和包括擬桿菌屬(Bacterioides)和梭菌 屬 的 完 全 厭 氧 微 生 物(strictly anaerobic microorganisms)等也可能高濃度存在。
3. 微 生 物 的 腐 敗 (microbial spoilage)。剛捕獲魚類和貝類常帶有種特 異的風味(species-specific flavor),冷藏數 日後就消失。當繼續貯藏一段時間後不 良 風 味 就 逐 漸 產 生 , 一 般 是 氨 味 般 (ammonia-like)、硫味的(sulphurous)、麥 芽般(malt-like)或酸敗的(rancid)。比較魚 肉的不良風味,可了解海產食品腐敗時 微生物活性的重要性:(1)滅菌,(2)接種 特定的微生物,或(3)天然污染。這些研 究顯示許多產品的貯藏壽命短,可透過 其微生物菌群和化學特性來解釋。
許多有其加工重要性的魚類和其它 水生動物都生活在冷水域,它們的天然 微生物菌群包括在溫度−2oC 至 0oC 以上 的冷藏產品中能快速生長的耐冷性菌種 (psychrotoleant species)。這解釋許多冷水 域魚種在冰藏中的貯藏壽命(12~18 日)較 短,相較於熱帶的白肉魚貯藏 0oC 時達 18~35 天(Dalgaard and Huss, 1997; Gram,
1989)。此外,一些魚類、甲殼類動物和
軟體類動物肉含的氧化三甲胺(TMAO),可 刺 激 微 生 物 的 生 長 和 活 性 。 一 般 而 言,淡水產比海水產動物的 TMAO 含量 更少,但兩棲息地都存在的種類彼此的 含量差異變動很大(Hebard et al., 1982)。
氣 單 胞 菌 屬 、 交 替 單 胞 菌 屬 (Alteromonas) 、 多 數 的 腸 桿 菌 科 (Enterobacteriaceae) 、 希 瓦 氏 菌 屬 (Shewanella)和弧菌屬(Vibrio)、以及包括 所 有 海 洋 發 光 細 菌 的 發 光 桿 菌 屬 (Photobacterium)都可將 TMAO 還原成三 甲胺(TMA),這樣的厭氧呼吸(anaerobic respiration)有助於它們在氧氣受限狀況下 的生長,例如真空包裝或調氣(modified- atmosphere) 包 裝 產 品 (Barret and Kwan,
1985; Proctor and Gunaslus, 2000)。TMA
貢獻腐敗海產食品(spoilaged seafoods)的 典型氨味般(ammonia-like)和魚腥味(fishy) 之不良氣味,特別在 pH 值高於約 6.5 的 產品(Castell and Triggs, 1955)。且同牛肉 和豬肉相比,有鰭魚類、甲殼類動物和 軟體類動物在僵直後的 pH 值較高,這也 關係於較短的貯藏壽命。白肉底層有鰭 魚類和甲殼類的 pH 值約 6.5 至 7 以上,而 表層 紅肉魚如鮪魚、鯕鰍(鬼頭刀,
mahi-mahi)、鯖魚和長嘴硬鱗魚(garfish) 的 pH 值低至~5.8。軟體動物的 pH 值與 白 肉 有 鰭 魚 相 似 , 但 其 碳 水 化 合 物 (2.5~5.0%)較豐富,相對的有鰭魚類和甲 殼類的含量在 0.5%以下。其結果,在降 低的 pH 下發酵類型的腐敗,對軟體動物 而 言 是 常 態 的 , 但 其 它 海 產 食 品 很 罕 見,除非也加入碳水化合物(Bremner and
Statham, 1983; He et al., 2002; ICMSF, 1998; López-Caballero et al., 2000;
Vasakou et al., 2003)。最後,海產食品中
高濃度存在的游離胺基酸會被腐敗微生 物代謝,例如蝦中的精胺酸、及紅色肉 海 洋 有 鰭 魚 中 的 組 胺 酸(Abe, 1983;Chinivasagam et al., 1998)。
利用來自海產食品的許多微生物生 產胞外蛋白質水解酵素已有許多的商業 化 (Kobatake et al., 1992; Venugopal,
1990)。不過,海產食品腐敗的微生物通
常 產 生 的 不 良 風 味 是 從 肌 肉 萃 取 物 (muscle extractives)中的基質,蛋白質水 解活性對生鮮海產食品的腐敗並不重要 (Karnop, 1982; Lerke et al., 1967)。微生物 蛋白質分解酵素的評定主要針對生鮮海 產食品,故也有必要繼續研究包括輕度 保 藏 和 半 保 藏(semi-preserved)的海產食 品。海 產 食 品 很 容 易 受 到 條 件 性 (opportunistic)和病原 性 (pathogenic)微生 物 的 侵 入 。 棲 息 地 屬 微 生 物 豐 富 的 環 境,即主要決定了海產食品中的微生物 含 量(Ghaly et al., 2010; Kuley et al.,
2017)。通常,海產食品的腐敗大部分是
由 和 產 生 生 物 胺(biogenic amines) 、 醇 類、組織胺、腐胺(putrescine)、硫化物、有機酸、醛類和酮類等關連的微生物的 生長和代謝引起(Kuley et al., 2017)。嗜冷 菌(psychrophilic bacteria)是造成冷藏或冷 凍 海 產 食 品 的 腐 敗 的 主 要 微 生 物 群 。
Sivertsvik et al. (2002)
發現好氧(aerobic)或兼性厭氧(facultative anaerobic)嗜冷菌的 革 蘭 氏 陰 性 菌 , 例 如 莫 拉 氏 菌 屬 (Moraxella) 、 腐 敗 希 瓦 氏 菌 (Shewanella
putrefaciens
) 、 不 動 桿 菌 屬(Acinetobacter) 、 假 單 胞 菌 屬 (Pseudomonas) 、 發 光 桿 菌 屬 (Photobacterium)、氣單胞菌屬、黃桿菌 屬(Flavobacterium)和弧菌屬(Vibrio)是海 產食品中的主要腐敗菌,而特定腐敗生 物如希瓦氏菌屬(Shewanella)、磷光發光 菌(Photobacterium phosphoreum)和假單胞 菌 等 是 造 成 腐 敗 的 主 因 (Gram and
Dalgaard, 2002)。
革蘭氏陰性菌是海產食品腐敗的主 要貢獻者,但連續加工或貯藏/運輸時間 的延長也提供革蘭氏陽性菌有機會變成 優 勢 菌 進 而 導致 腐 敗(Al Bulushi et al.,
2010)。Dalgaard (2000)
指出革蘭氏陰性菌 的 磷 光 發 光 菌 和 乳 酸 菌 (lactic acid bacteria)兩者都是魚類的主要腐敗菌。革 蘭 氏 陽 性 菌 如 微 球 菌 屬 、 棒 狀 桿 菌 屬 (Corynebacterium)、桿菌屬、葡萄球菌屬 (Staphylococcus) 、 梭 菌 屬 、 鏈 球 菌 屬 (Streptococcus) (Al Bulushi et al., 2010)和 熱 死 環 絲 菌(Brochothrix thermosphacta) (Fall et al., 2010; Lalitha et al., 2005)也確 認是海產食品中的腐敗微生物。因而可 以推斷革蘭氏陰性和革蘭氏陽性菌兩者 都 負 責 海 產 食 品 的 腐 敗 , 可 是 採 樣 地 點、地理位置和捕撈方法才是決定微生 物類型和數量的因素(Ghaly et al., 2010)。組 織 中 的 低 分 子 量 物 質 例 如 小 分 子 胜
肽、碳水化合物和游離胺基酸,被微生 物利用作為生長的能量來源和一些副產 物 的 生 成 , 包 括 生 物 胺(Masniyom,
2011)、組織胺(Visciano et al., 2012)、含
硫化合物(Varlet and Fernandez, 2010)和其 它組成分。一些其餘細菌的酵素活性,例如耐 冷 腸 桿 菌 (psychrotolerant Enterobacteria)、弧菌屬(Vibrio spp)、氣 單胞菌屬和腐敗希瓦氏菌(S. putrefaciens) 等可將海產食品中的氧化三甲胺(TMAO) 還 原 為 會 產 生 魚 腥 味 的 三 甲 胺(TMA) (Arfat et al., 2015;
Lidbury et al., 2014)。
TMA 的產生伴隨次黃嘌呤的增加,後者 使海產食品帶有苦味(Tikk et al., 2006)。
次黃嘌呤的產生由內源性酵素誘導或者 更快地透過細菌誘導核苷酸 (肌苷或肌苷 酸) 的 分 解 (Masniyom, 2011; Varlet and
Fernandez, 2010; Visciano et al., 2012)。
4. 特定腐敗生物。生鮮冷藏和好氧 貯藏(aerobically stored)海產食品的腐敗,
主要是由硫化氫(H2S)產生的希瓦氏菌屬 細菌和假單胞菌屬引起的(
表 11),但這些
革蘭氏陰性和非發酵性桿菌的分類正改 變中。在漁產品,硫化氫產生的希瓦氏 菌屬細菌在鐵洋菜(iron agar)培養基使用 注入塗佈時,常被當作黑色菌落分離,然 後 以數量不多的表型試驗(phenotypic tests)鑑定為腐敗希瓦式菌。最近已經得 知這些硫化氫產生菌是由一些希瓦氏菌 屬組成,常出現於幾種漁產品的是波羅 的 海 希 瓦 式 菌 (Shewanella baltica)
(Fonnesbech Vogel et al., 2005;
Satomi et al ., 2006; Stenström and Molin, 1990;
Ziemke, 1998)。
通常,假單胞菌屬(Pseudomonas)都 未被鑑定至菌種層次(species level),但菌 株類似於海產食品中常見的莓實假單胞 菌 (Ps. fragi) 、 螢 光 假 單 胞 菌 (Ps.
fluorescens
)和戀臭假單胞菌(Ps. putida),而隆德假單胞菌(Ps. lundensis)是希臘產 好 氧 冷 藏 海 水 魚 的 主 導 腐 敗 菌 群 (Gillespie, 1981; Stenström and Molin,
1990)。假單胞菌屬(Pseudomonas spp.)無
法還原 TMAO,且在氧氣受限的條件下 生長明顯減緩。其它的細菌也可能影響 生鮮冷藏和好氧貯藏海產食品的腐敗,因而磷光發光菌(P. phosphoreum)可能負 責 TMA 生成,以及貢獻於好氧貯藏各種 冷藏魚的腐敗。參見
Dalgaard (1998)
綜述 和針對不同魚類的研究(Dalgaard et al.,2006; Olafsdottir et al., 2006a; 2006b)。另
外,具有脂質水解(lipolytic)能力的靜止 嗜冷桿菌(Psychrobacter immobilis)是海水 魚和淡水魚的主導腐敗菌群,儘管腐敗 潛在性低,但可能促進沙丁魚的酸敗型 腐敗(rancid spoilage)(Gennari et al., 1999;González et al., 2000)。
儲存在高 CO2濃度調氣包裝的新鮮 海產食品,磷光發光菌(P. phosphoreum) 不論發光和非發光變種都是重要的腐敗
微生物(
表 11)。磷光發光菌的菌種群是異
質性的(heterogeneous),自海洋食品分離 的菌種可能含有數種,包括魚腸發光桿