Effects of dietary Glutamine on Antioxidant Enzyme Activity and Immune Response in Burned Mice
Sung-Ling Yeh, PhD1, Huey-Fang Shang, PhD2, Ming-Tsan Lin, MD3, Chiu-Li Yeh, MS1, Wei-Jao Chen, MD3
1Institute of Nutrition and Health Science, 2Department of Microbiology and Immunology, Taipei Medical University, and 3Department of Surgery, College of Medicine, National Taiwan University, Taipei, Taiwan, Republic of China.
Running title: Effect of glutamine on immune response in burns
Corresponding author: Wei-Jao Chen, MD Department of Surgery
National Taiwan University Hospital 7 Chung-Shan S. Road
Taipei, Taiwan 100 Republic of China
Tel: 8862-23970800, ext. 2122 Fax: 8862-23412969
E-mail: [email protected]
ABSTRACT
OBJECTIVES: To investigate the effect of dietary glutamine (Gln) on specific antibody production and antioxidant enzyme activities in burned mice vaccinated with detoxified Pseudomonas exotoxin A linked with the outer membrane proteins I and F (PEIF). Also, the survival rate of vaccinated and non-vaccinated burned mice
infected with Pseudomonas aeruginosa (P. aeruginosa) was evaluated.
METHODS: There were 3 consecutive experiments. Experiment 1: Thirty BALB/c mice were assigned to 2 groups. A control group was fed casein as the protein source;
the Gln group provided 4% Gln (w/w) to replace part of the casein. Mice were immunized twice with PEIF, and the production of specific antibodies against PEIF was measured every week. Eight weeks after immunization, all mice received a 30%
body surface area burn injury. Mice were sacrificed 24 h after the burn. The antioxidant enzyme activities and lipid peroxides in the tissues as well as specific antibody production were analyzed. Experiment 2: Twelve mice were divided into a control and Gln groups, and fed with one of 2 experimental diets for 4 weeks. Then burn injury was induced, and mice were sacrificed 24 h later. In vitro, splenocytes were cultured, and interleukin (IL)-4,IL-10 was measured after mitogen stimulation.
Experiment 3: Survival rates of vaccinated and non-vaccinated burned mice complicated with P. aeruginosa infection were evaluated. The survival rate was observed for 8 days after the burn.
RESULTS: Antioxidant enzyme activities and lipid peroxides in tissues tended to be lower in the Gln group than in the control group after the burn. Specific antibody production against P. aeruginosa increased significantly in the Gln group at 4 and 7 weeks after immunization, and at 24 h after the burn. IL-4 concentrations in mitogen-
stimulated splenocytes were significantly higher in the Gln group than in the control group. Survival rates of non-vaccinated burned mice in the Gln group were
significantly higher than that of the control group, whereas there was no difference in the survival of vaccinated burned mice after bacterial infection.
CONCLUSIONS: These results suggest that vaccinated mice receiving a Gln- enriched diet may have enhanced humoral immunity and attenuated oxidative stress induced by burn injury. Also, Gln supplementation improves the survival of burned mice complicated with P. aeruginosa infection.
Key words: burns, glutamine, antioxidant enzyme activity, antibody, vaccination, Pseudomonas aeruginosa
INTRODUCTION
Burn injury is a post-traumatic inflammatory disease. Severe injury, particularly thermal injury leads to a profound depression of both humoral and cell-mediated immunities.1-4 The immunosuppression which occurs after burn injury causes increased susceptibility to infection.5 In addition, the response to the initial burn injury and the presence of the burn are often associated with secondary damage to tissues distant from the injured site.6-9 This response appears to be mediated by both reactive oxygen metabolites and activated neutrophils.10,11 Nishigaki et al.12 reported that lipid peroxide levels increase in burned rat skin, and lipid peroxide generated in the burn wound may accumulate in the liver, lung, kidney, and gut of injured animals.
Glutamine (Gln) has traditionally been thought of as a nonessential amino acid, but laboratory and clinical data suggest that it may be essential during certain inflammatory
conditions, such as infection and injury.13,14 A number of studies have demonstrated beneficial effects of supplying Gln or its precursors (ornithin -ketoglutarate and - ketoglutarate) for metabolic-stressed conditions by increasing nitrogen retention,
preserving the integrity of the intestinal mucosa and intestinal permeability, maintaining immunologic function, and reducing infections.13-19 Parry-Billings et al.14 reported that plasma Gln levels decreased and remained depressed for 3 weeks after injury in burn patients. Their experiments showed that the depressed Gln concentrations were associated with reduced proliferation of lymphocytes from healthy volunteers and depressed phagocytosis by peritoneal macrophages from normal mice. Ogle et al.20 also reported that Gln improved the bactericidal ability of abnormal neutrophils from pediatric patients after burns. Although Parry-Billings et al. and Ogle et al. suggested the efficacy of Gln supplementation, they did not supply Gln to their patients. Very few studies of Gln supplementation on immune responses have been performed in burn injury. To our knowledge, there is no study investigating the effects of Gln on humoral immunity and antioxidant status in burn injury.
Pseudomonas bacteremia presents another medical problem because of its high mortality and strong antibiotic resistance.21 Patients with extensive thermal injury are at high risk of infection with Pseudomonas aeruginosa (P. aeruginosa).22 Chen et al.23 designed a novel vaccine, PEIF, against P. aeruginosa, which can enhance specific antibody production and effectively block P. aeruginosa challenge in burned mice. In this study, we immunized mice with this novel vaccine against P.
aeruginosa before burn injury to investigate whether Gln has beneficial effects on attenuating oxidative stress induced by burn injury and the specific antibody
production against PEIF. In addition, in vitro cytokine secretion and the survival rates in vaccinated and non-vaccinated burned mice complicated with a lethal dose of P.
aeruginosa were also evaluated.
MATERIALS AND METHODS Animals
Male BALB/c mice aged 4 wk and weighing 10-15 g were purchased from Animal Center, Taiwan University Hospital, Taipei, Taiwan. All mice were housed in temperature- and humidity-controlled rooms, and allowed free access to standard chow (TAMA Feeds Inc. Richmond, Indiana) and water for 1 week prior to the experiment. Animals included in this study were kept according to standard
experimental animal care protocols. All procedures involving animals were approved by Taipei Medical University Standing Committee on Animal Research.
Study protocol
Experiment 1: Thirty mice were randomly assigned to 2 groups, 15 in each group.
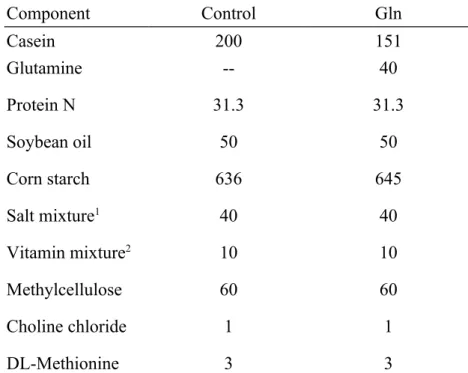
One group was fed a control diet (control), in which all amino acids were provided by casein. The other group was fed a Gln (Gln)-enriched diet, by which 4% Gln (w/w, Sigma, St Louis, MO) was used to replace part of the casein, corresponding to 40% of the dietary nitrogen source. The 2 diets were isonitrogenous (Table I). Mice were anesthesized with ether, and blood was taken from the retrobulbar vessel before immunization with the novel PEIF vaccine against P. aeruginosa. Production and purification of the recombinant PEIF protein followed procedures described
previously.23 Theemulsified vaccine was prepared by mixing the purified recombinant PEIF protein with an equal volume of complete Freund’s adjuvant, and then each mouse was vaccinated subcutaneously at a dose of 2 g/mouse on day 1. A booster
injection was given at a dose of 4g/mouse of PEIF emulsified with an equal volume of incomplete Freund’s adjuvant on day 28. Before the burn, immunized mice were bled from the retrobulbar vessel on days 21, 28, 35, 42, 49, and 56. The sera were isolated and stored at –70 ℃ until being assayed. After 8 weeks, a modification of the burned mouse procedure was used.24,25 Mice were anesthetized with sodium pentobarbitol (0.71 g/g body wt) and shaved dorsally prior to burning. A Teflon template with a precisely cut window (2.5 x 3 cm2) was pressed firmly against the shaved back. Ethanol (95% v/v, 0.5 ml) was evenly spread over the area of the back outlined by the window, ignited, and allowed to burn for 15 s.24,25 Animals were immediately resuscitated with an intraperitoneal injection of sterile 0.9% saline (10 ml/100 g body wt).15 This procedure produced a full-thickness burn injury on
approximately 30% of the total body surface area. All mice were deprived of food for 24 h with only free access to water, in order to induce a hypermetabolic state
simulated in burn patients.16,26 Mice were anesthetized and sacrificed by cardiac puncture 24 h after the burn. Blood samples were collected in tubes containing heparin for analysis of T lymphocyte subpopulations. The remaining blood samples were centrifuged to isolate the sera. Amino acid was analyzed by the standard ninhydrin technology (Beckman Instrument, model 6300, Palo Alto, CA), after deproteinization of the plasma with salicylic acid.27 Tissues including the liver and kidneys were rapidly excised. All samples were stored at –70 oC until being assayed for antioxidant enzyme activities and lipid peroxide levels.
Experiment 2: Twelve mice were divided into the control and Gln groups, and fed either the control or Gln-enriched diet for 4 weeks. After that, burn injury was induced, and mice were sacrificed 24 h after the burn. Spleens of the mice were removed aseptically, and were placed in phosphate-buffered saline (PBS) for
pretreatment of in vitro splenocyte cultures.
Experiment 3: The survival rates of non-vaccinated and vaccinated burned mice complicated with P. aeruginosa were evaluated. Thirty mice were allocated to the non-vaccinated group and were further divided into 2 experimental groups, with each group containing 15 mice. Mice were fed the control or Gln diets for 8 weeks. After that, burn injury was induced, and immediately after the burn, 3.2 x 105 CFU (1x LD50) P. aeruginosa strain PAO1 (ATCC 15692) was injected into the burned skin of the mice. Additional 30 mice were immunized twice with the novel PEIF vaccine against P. aeruginosa and were fed with control or Gln diets for 8 wks as described in Exp 1. After that, burn was induced and 2x LD50 P. aeruginosa was subcutaneously injected into the burn area. Survival of the burned mice was recorded every 6 h in the first 3 days, and then every 12 h until the end of 8 days. The mice were also deprived of food except water for 24 h, then the experimental diets were given for the
remaining days.
Measurements of antioxidant enzymes and TBARS
Fifteen-percent tissue homogenates were prepared at 4 oC in a 250-mM sucrose solution, containing 10 mM-Hepes (pH 7.4) using a homogenizer. The homogenates were centrifuged to remove cell debris and mitochondria.28 Thesupernatant was used for analysis of superoxide dismutase (SOD) and glutathione peroxidase (GSHPx) activities (enzyme kits from Randox, Antrim, Northern Ireland). Protein
concentrations of supernatants were measured by Lowry's method. The production of thiobarbituric acid-reactive substances (TBARS, assumed to be mainly
malondialdehyde and its precursors) in mouse liver and kidney homogenates was determined by the method of Uchiyama and Mihara.29 Briefly, the homogenates were
precipitated with trichloroacetic acid and mixed. The mixture was centrifuged and TBA was added to the supernatant. After boiling for 1 h, the reactants were cooled, and TBARS were measured at 532nm. The molar extinction coefficient of
malondialdehyde was assumed to be 156,000.30 Analysis of specific antibody production against PEIF
The specific antibody production of vaccinated mice was measured by ELISA as described previously.23 Briefly, purified recombinant PEIF protein was coated onto polyvinylchloride, flat-bottom, 96-well Falcon microtiter plates overnight at 4 ℃ with a protein concentration of 3 g/ml in the coating buffer (pH 9.6 carbonate buffer). The coated plates were then blocked with 0.5% BSA-PBS. Mouse sera from each group were diluted 1000-fold with 0.5% BSA-PBS, and 50 l of diluted sera was added to the coated well and incubated for 1 h at 37 ℃. Bound specific antibodies were detected using
peroxidase-conjugated goat anti-mouse secondary antibody (Sigma, St. Louis, MO). After 3 washings, 100 l of substrate solution (0.54 mg/ml 2,2’-azinobis-3-ethylbenzthiazoline- 6-sulfonic acid, ABTS, and 0.03% H2O2 in 0.1 M citric acid) was added to each well, and the absorbance was read after 15 min using a microplate reader at 405 nm. Normal mouse serum was used as the negative control.
Analysis of T lymphocyte subpopulations
Flow cytometry was used to determine the proportions of CD4+, CD8+, and CD3 T lymphocytes in fresh blood. One hundred microliters of blood was incubated for 15 min at 4 oC with 10 ul of fluorescein-conjugated (FITC) rat monoclonal anti-mouse CD4+, phycoerythrin-conjugated (PE) rat anti-mouse CD8+, and FITC rat anti-mouse CD3 (Serotec, Oxford, UK). After that, red blood cells (RBC) were lysed with lysing buffer (Serotec). Fluorescence data were collected on 5 x 104 viable cells and
analyzed by flow cytometry (Coulter, Miami, FL).
In vitro cultures of splenocytes and cytokine assay
Splenocytes were obtained by mechanical disruption of the spleen using the homogenizer. Cell suspensions were passed through a sterile nylon mesh to remove debris. RBCs were lysed by sterile distilled water for 15 s, and immediately
neutralized to isotonic cell suspensions. After washing with PBS 3 times (300 xg for 5 min), splenocytes were resuspended in RPMI-1640 with antibiotics and fetal calf serum. The number of isolated splenocytes was determined by hemacytometer count using the trypan blue dye exclusion method. Phytohemagglutinin (PHA, 10 ng/mL;
Sigma) was used to stimulate cytokine production by isolated splenocytes in culture.
Triplicate wells of 96-well flat-bottomed microtiter plates (Falcon, Becton Dickinson, Oxford, CA.) were seeded with 100 ul of splenocytes (2.5 x 106 cells/ml in RPMI- 1640) and 100 ul of mitogen. Control wells contained cells plus 100 ul of medium.
The final volume in all wells was 200 ul. Splenocytes were incubated in the presence or absence of PHA for 24 h at 37 oC in an atmosphere of 95% air and 5% CO2. Supernatants were centrifuged and stored at –70 oC until the analysis for cytokine.
Interleukin (IL)-4 and IL-10 concentrations in splenocyte supernatants were determined by a commercially available enzyme-linked immunosorbent assay
(ELISA) in microtiter plates, antibodies specific for mice cytokines were coated onto the wells of the microtiter strips provided (Amersham Pharmacia Biotech,
Buckinghamshire, UK).
Statistics
Data are expressed as the mean SD. Differences between groups were analyzed by student’s t-test. The survival rate was determined by Kaplan-Meier survival analysis. A p
value < 0.05 was considered statistically significant.
RESULTS
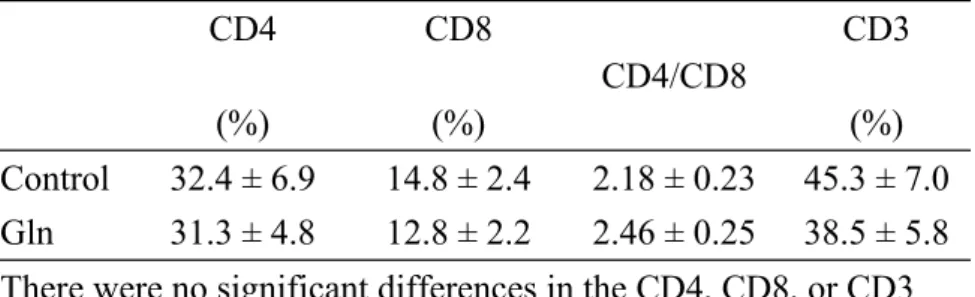
There were no differences in initial body weights and weights after experimental diets between the 2 experimental groups in either experiments 1, 2, or 3 (data not shown). The results in experiment 1 showed that the Gln group had significantly higher plasma Gln levels than did the control group (462.2 82.6 vs. 332.5 37.7 nmol/mL, p < 0.05). No significant differences in the percentages of CD4, CD8, CD3 T cells or the CD4/CD8 T cell ratio were observed between the Gln and control groups after the burn (Table II). Antibody production increased logarithmically after the second booster and reached a plateau after 7 weeks. The specific antibody production in the Gln group was significantly higher than that in the control group at various time schedules (Fig. 1). SOD activities and lipid peroxide concentrations in the kidney and GSHPx activities in the liver were significantly lower in the Gln group than in the control group after the burn of vaccinated mice (Table III).
In experiment 2, stimulation of splenocytes with PHA significantly increased the production of IL-4 in the Gln group. However, the production of IL-10 did not differ between the 2 groups when the same concentration of mitogen was administered (Fig.
2A,B).
In experiment 3, the survival rate of non-vaccinated burned mice in the Gln group was significantly higher than that of the control group after bacterial infection (p <
0.001). However, no statistically significant difference in survival was observed between the 2 groups in the vaccinated burned mice challenged with 2x LD50 of P.
aeruginosa (p = 0.178) (Fig. 3A, B).
DISCUSSION
In this study, we administered Gln prior to burn injury to investigate the effect of Gln on burned mice, because rodents are known to be relatively resistant to trauma and to recover rapidly after a burn.16 With dietary supplementation in a very short period of time after a burn, the influence of Gln on burns might not be observed.
Since very few studies have investigated the effect of Gln on burn injury, the results of this study may provide basic information on the possible roles of Gln in burns. We did not include a non-burn control group in this study, because numerous studies have already shown that burn injury results in oxidative stress and impaired immune function.1-4, 6-9
Gln is of major importance for muscle metabolism and is a preferred energy source for cells of the intestinal mucosa and of the immune system, particularly macrophages and lymphocytes.20 An in vitro study by Rohde et al.31 showed that Gln influences the production of T cell-derived cytokines and is important for optimal T lymphocyte proliferation. Yaqoob and Calder32 revealed that IL-2 production and IL-2 receptor expression depend on the concentration of Gln present in the culture medium.
Gismondo et al.33 also showed that orally administered Gln to nude mice significantly increased intestinal CD3, CD4, and CD8 lymphocytes when compared to the group without Gln supplementation. In this study, blood CD4, CD8 T cells, and the CD4/CD8 ratio did not differ between the 2 groups. Also, there were no differences in CD3 populations between the 2 groups. Our findings are inconsistent with those of the reports mentioned above. There are 2 possible explanations for this: 1) the result
of in vitro studies may not actually reflect the in vivo situations; and 2) samples used for evaluating the effect of Gln on lymphocyte functions were derived from healthy volunteers or normal rodents. This may differ from the stressed metabolic condition observed in this study, and may consequently lead to different immune responses.
The findings in this study showed that the production of specific antibodies against P. aeruginosa was significantly higher in the Gln group than in the control group after immunization and after the burn. In order to understand the possible role of cytokine secretion on antibody production, IL-4 and IL-10 productions in mitogen-stimulated splenocyte cultures were analyzed. IL-4 and IL-10 are produced by Th2 lymphocytes.
A predominant Th2 effect results in activation of B lymphocyte, up-regulation of antibody production and is anti-inflammatory. The Th2 cytokines, particularly IL-4 promote differentiation of Th2 cells.34 Although IL-10 did not differ between the 2 groups, IL-4 production was significantly higher in the Gln group than the control group when the same concentration of mitogen was administered. This result suggests that orally supplemented Gln may enhance IL-4 secretion which may consequently promote antibody production in burned mice.
Burn injury is a trauma with high oxidative stress.35 After burn injury, generalized tissue inflammation is present in uninjured organs within hours.36 Organ injury remote from the region of thermal injury has been shown to be due to the intravascular action of complements, resulting in stimulation of intravascular neutrophils, leading to the formation of toxic oxygen products.8 Lipid peroxide is thought to be one of the most harmful substances produced after burns.37 SOD and GSHPx are enzymes which protect tissues from the effects of free radicals and lipid peroxides, and the activities of both SOD and GSHPx increase after free-radical-
mediated injury and lipid peroxidation.38 Saitoh et al.37 demonstrated that Mn-SOD activities in lung and kidney were significantly higher than in the control group after a burn. The results of this study revealed that SOD activities in kidney and GSHPx activities in liver were significantly lower in the Gln group when compared with the control group. Also, lipid peroxide concentrations in the kidney were lower in the Gln group than in the control group after the burn of vaccinated mice. This finding may indicate that Gln supplementation tends to attenuate oxidative stress induced by burn injury. A study by Morlion et al.39 showed that generation of cysteinyl-
leukotrienes was enhanced after Gln supplementation in surgical patients. In an animal model of preinfusion with Gln and hepatic injury, Hong et al.40 also revealed that the availability of Gln is critical in the generation of hepatic glutathione. Because glutathione is a potent antioxidant that protects tissue from free radical injury, and cysteinyl-leukotrienes also contain glutathione, it is possible that Gln supplementation enhances antioxidant protection after a burn.
An animal study by Gianotti et al.41 showed that Gln supplementation decreased bacterial translocation and increased survival rates in septic rats. Wischmeyer et al.42 demonstrated that the incidence of gram-negative bacteremia was significantly reduced in the intravenously Gln-administered burn patients. In this study, we also observed higher survival rates in the Gln group than in the control group in burned mice complicated with P. aeruginosa infection. The survival rates of the Gln and control groups were 23% and 0% at 8 days after the burn. The trend was also observed in PEIF vaccinated burned mice with 2x LD50 P. aeruginosa infection, although the difference between the 2 groups was not statistically significant. The survival rates of the Gln and control group were 69% and 40%, respectively. This finding indicate that PEIF vaccination effectively reduced the mortality of burned
mice after P. aeruginosa infection. However, the vaccination effect of PEIF against P. aeruginosa may be too strong to observe the beneficial effect of Gln on the survial in burned mice. Since the defense mechanism against bacteria is dependent on the neutralization and opsonization of antibodies,24 the results of this study may indicate that Gln augments host defense by enhancing antibody production, and consequently improves survival in burned infected mice.
In conclusion, the findings of this study suggest that Gln supplementation did not influence T cell populations; however, humoral immunity was enhanced and
oxidative stress induced by burn injury was attenuated in vaccinated burned mice.
Also, Gln administration before burn injury significantly increased survival in burned mice complicated with P. aeruginosa infection.
Acknowledgements
This study was supported by research grant NSC 90-2314-B-002-305 from the National Science Council, ROC.
REFERENCES
1. Farrell MF, Day NK, Tsakraklides V, et al. Study of lymphocyte depletion and serum complement perturbations following acute burn trauma. Surgery
1973;73:697
2. Goldman AS, Rudloff HB, McNamee R, et al. Deficiency of plasma humoral recognition factor activity following burn injury. J Reticuloendothel Soc 1974;15:193
3. Miller CL, Baker CC. Changes in lymphocyte activity after thermal injury: the role of suppressor cells. J Clin Invest 1979;63:202
4. Ninnemann JL. Immunosuppression following thermal injury through B cell activation of suppressor T cells. J Trauma 1980;20:206
5. O’Sullivan ST, O’Connor TPF. Immunosuppression following thermal injury: the pathogenesis of immunodysfunction. Br J Plastic Surg 1997;50:615
6. Till GO, Beauchamp C, Menapace D, et al. Oxygen radical dependent lung damage following thermal injury of rat skin. J Trauma 1983;23:269
7. Till GO, Hatherill JR, Tourtellotte WW, et al. Lipid peroxidation and acute lung injury after thermal trauma to skin. Am J Pathol 1985;119:376
8. Cetinkale O, Konukoglu D, Senel O, et al. Modulating the functions of neutrophils and lipid peroxidation by FK506 in a rat model of thermal injury. Burns
1999;25:105
9. Gurbur V, Corak A, Yegen BC, et al. Oxidative organ damage in a rat model of thermal injury: the effect of cyclosporin A. Burns 1997;23:37
10. Simpson R, Alon R, Kobzik L et al. Neutrophil and neutrophil-mediated injury in intestinal ischemia-reperfusion. Ann Surg 1993;218:444
11. Hernandez LA, Grisham MB, Twohig B et al. Role of neutrophils in ischemia- reperfusion induced microvascular injury. Am J Physiol 1987;253:H699
12. Nishigaki I, Hagihara M, Hiramatsu M, et al. Effect of thermal injury on lipid peroxide levels of rat. Biochem Med 1980;24:185
13. Willmore DW. The effects of glutamine supplementation in patients following
elective surgery and accidental injury. J Nutr 2001;131:2543S
14. Parry-Billings M, Evans J, Calder PC, et al. Does glutamine contribute to immunosuppression after major burns? Lancet 1990;336:523
15. Boucher JL, Obled C, Farges MC, et al. Ornithine -ketoglutarate modulates tissue protein metabolism in burn-injured rats. Am J Physiol 1997;273:E557 16. Vaubourdolle M, Coudray-Lucas C, Jardel A, et al. Action of enterally
administered ornithine -ketoglutarate on protein breakdown in skeletal muscle and liver of the burned rats. J Parenter Enter Nutr 1991;15:517
17. Stehle P, Wurste N, Puchestein C, et al. Effect of parneteral glutamine peptide supplements on muscle glutamine loss and nitrogen balance after major surgery.
Lancet 1989;1:231
18. van der Hulst, RRWJ, van Kreel BK, von Meyenfeldt MF, et al. Glutamine and the preservation of gut integrity. Lancet 1993;341:1363
19. Calder PC. Glutamine and the immune system. Clin Nutr 1994;13:2
20. Ogle CK, Ogle JD, Mao JX, et al. Effect of glutamine on phagocytosis and bacterial killing in normal and pediatric burn patient neutrophils. J Parenter Enter Nutr 1994;18:128
21. Hancock REW. Intrinsic antibiotic resistance of Pseudomonas aeruginosa. J Antimicrob Chemother 1986;18:653
22. Liberman MM, Walker HL, Ayala E, et al. Active and passive immunization with Pseudomonas aeruginosa ribosomal vaccines and antisera in the burned rat model. J Surg Res 1986;40:138
23. Chen TY, Shang HF, Chen TL, et al. Recombinant protein composed of Pseudomonas exotoxin A, outer membrane protein I and F as vaccine against P.
aeruginosa infection. Appl Microbiol Biotechnol 1999;52:524
24. Holder IA. The pathogenesis of infection owing to Pseudomonas aeruginosa using the burned mouse model: experimental studies from the Shriners Burns Institute, Cincinnatti. Can J Microbiol 1985;31:393
25. Holder IA, Jogan M. Enhanced survival in burned mice treated with antiserum prepared against normal and burned skin. J Trauma 1971;11:1041
26. Cynober L. Amino acid metabolism in the thermal burns. J Parenter Enter Nutr 1989;13:196
27. Smith RJ, Panico K. Automated analysis of o-phthalaldehyde derivatives of amino acids in physiological fluids of reverse phase high performance liquid chromatography. J Liq Chromatogr 1985;8:1783
28. Yeh SL, Chang KY, Huang PC, et al. Effects of n-3 and n-6 fatty acids on plasma eicosanoids and liver antioxidant enzymes in rats receiving toal parenteral nutrition. Nutrition 1997;13:32
29. Uchiyama M, Mihara M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal Chem 1978;86:271
30. Hu ML, Frankel EN, Leibovitz BE, et al. Effect of dietary lipids and vitamin E on in vitro lipid peroxidation in rat liver and kidney homogenates. J Nutr
1989;119:1574
31. Rohde T, Maclean DA, Klarlund Pedersen B. Glutamine, lymphocyte proliferation and cytokine production. Scand J Immunol 1996;44:648
32. Yaqoob P, Calder PC. Glutamine requirement of proliferating T lymphocytes.
Nutrition 1997;13:646
33. Gismondo MR, Drago L, Fassina MC, et al. Immunostimulating effect of oral glutamine. Dig Dis Sci 1998;43:1752
34. Dipiro JT. Cytokine networks with infection: mycobacterial infections, leishmaniasis, human immunodeficiency virus infection, and sepsis.
Pharmacotherapy 1997;17:205
35. Tredget EE, Yu YM. The metabolic effects of thermal injury. World J Surg 1992;16: 68
36. Youn YK, LaLonde C, Demling R. The role of mediators in the response to thermal injury. World J Surg 1992;16:30
37. Saitoh D, Okada Y, Ookawara T, et al. Prevention of ongoing lipid peroxidation by wound excision and superoxide dismutase treatment in the burned rat. Am J Emerg Med 1994;12:142
38. Gonzales R, Auclair C, Voisin E, et al. Superoxide dismultase, catalase, and
glutathione peroxidase in red blood cells from patients with malignant disease.
Cancer Res 1984;44:4137
39. Morlion BJ, Stehle P, Wachtler P, et al. Total parenteral nutrition with glutamine dipeptide after major abdominal surgery. Ann Surg 1998;227:302
40. Hong RW, Rounds JD, Helton WS, et al. Glutamine preserves liver glutathione after lethal hepatic injury. Ann Surg 1992;215:114
41. Gianotti L, Alexander JW, Gennari R, et al. Oral glutamine decreases bacterial translocation and improves survival in experimental gut-origin sepsis. J Parenter Enter Nutr 1995;19:69
42. Wischmeyer PE, Lynch J, Liedel J, et al. Glutamine administration reduces gram-negative bacteria in severely burned patients: a prospective, radomized, double blind trial verus isonitrogenous control. Crit Care Med 2001;29:2075
FIGURE LEGENDS
Fig. 1. Production of PEIF-specific antibodies in the control and Gln groups. Mice were immunized twice with recombinant PEIF protein on days 1 and 28, and sera antibody titers were measured by ELISA at weeks 0, 3, 4, 5, 6, 7, and 8. The dilution of mice antiserum was 1:1000. * Significant difference between the 2 groups (p <
0.05).
Fig. 2. A) Interleukin (IL)-4 and B) IL-10 concentrations in PHA-stimulated
splenocyte cultures. * p < 0.05 by Student’s t test. There was no difference in IL-10 concentrations between the 2 groups after mitogen stimulation.
Fig. 3. A) Survival curves of non-vaccinated burned mice challenged with 1x LD50 P.
aeruginos Survival rates in the Gln group were significantly higher than those of the control group (p < 0.001). B) Survival curves of vaccinated burned mice challenged with 2x LD50 P. aeruginos. There was no significant difference in the survival between the 2 groups (p = 0.178).
Table I. Composition of the experimental diets (g/kg)
Component Control Gln
Casein 200 151
Glutamine Protein N Soybean oil Corn starch Salt mixture1 Vitamin mixture2 Methylcellulose Choline chloride DL-Methionine
-- 31.3
50 636
40 10 60 1 3
40 31.3
50 645
40 10 60 1 3
1Salt mixture contains the following (mg/g): calcium phosphate diabasic 500mg, sodium chloride 74mg, potassium sulfate 52mg, potassium citrate monohydrate 220mg, magnesium oxide 24mg, manganese carbonate 3.5mg, ferric citrate 6mg, zinc carbonate 1.6mg, curpric carbonate 0.3mg, potassium iodate 0.01mg, sodium selenite 0.01mg, and chromium potassium sulfate 0.55mg.
2Vitamin mixture contains the following (mg/g): thiamin hydrochloride 0.6mg, riboflavin 0.6mg, pyridoxine
hydrochloride 0.7mg, nicotinic acid 3mg, calcium pantothenate 1.6mg, D-biotin 0.02mg, cyanocobalamin0.001mg, retinyl palmitate 1.6mg, DL-α-tocopherol acetate 20mg, cholecalciferol 0.25mg, menaquinone 0.005mg.
Table II. Superoxide dismutase (SOD), and glutathione peroxidase (GSHPx) activities and thiobarbituric acid-reactive substances (TBARS) in liver and kidney homogenates after the burn
SOD (U/g tissue) GSH-Px (U/g tissue) TBARS (nmol/g tissue)
Control Gln Control Gln Control Gln
Liver 6.85 ± 1.34 10.38 ± 8.99 1.91 ± 0.32 1.64 ± 0.28* 2.91 ± 0.90 2.55 ± 0.58 Kidney 7.84 ± 1.45 5.32 ± 3.14* 1.11 ± 0.31 1.20 ± 0.3 2.34 ± 0.74 1.85 ± 0.61* Data are expressed as the mean SD.
*Significantly different from the control group at p < 0.05.
Table III. Blood CD4, CD8, and CD3 cells and the CD4/CD8 ratio between the 2 groups after the burn
CD4 (%)
CD8 (%)
CD4/CD8
CD3 (%) Control 32.4 ± 6.9 14.8 ± 2.4 2.18 ± 0.23 45.3 ± 7.0 Gln 31.3 ± 4.8 12.8 ± 2.2 2.46 ± 0.25 38.5 ± 5.8 There were no significant differences in the CD4, CD8, or CD3 populations or the CD4/CD8 ratio between the 2 groups.
0 0.2 0.4 0.6 0.8 1
0 week 4 weeks 7 weeks 24 h after the burn
O D
405Control
*
Gln
* *
0 50 100 150 200 250
0 10
PHA(ng/mL)
IL-4(pg/mL)
Control Gln
0 100 200 300 400 500 600 700
0 10
PHA(ng/mL)
IL-10(pg/mL)
Control Gln
*
0 20 40 60 80 100
0 1 2 3 4 5 6 7 8
Time after burn (day)
Survival rate (%)
GLN ALA
GLN Control
0 20 40 60 80 100
0 1 2 3 4 5 6 7 8 天
GLN Control
Days after the burn
Survival rate (%)
Days after the burn
*
0 20 40 60 80 100
0 1 2 3 4 5 6 7 8
GLN Control Time after the burn (d)
Survival rate (%)
*
0 20 40 60 80 100
0 1 2 3 4 5 6 7 8
Day after burn (day)
Survival rate (%)
GLN ALA
GLN Control