doi:10.6342/NTU201701930
國立台灣大學醫學院解剖學暨細胞生物學研究所 博士論文
Graduate Institute of Anatomy and Cell Biology Cillege of Medicine
National Taiwan University Doctoral Dissertation
研究 Eupafolin 和山苦瓜對腫瘤壞死因子刺激肺臟發 炎之影響及相關機轉
To study the protective effects of eupafolin and wild bitter gourd on TNF-α-induced lung inflammation and
the related mechanisms
宋欣錦
Hsin-Ching Sung
指導教授:陳玉怜 博士 Advisor: Yuh-Lien Chen, Ph.D.
中華民國 106 年 7 月
July 2017
doi:10.6342/NTU201701930
I
doi:10.6342/NTU201701930
II
誌 謝 (Acknowledgment)
首先我要由衷地感謝我的博士班指導老師─陳玉怜教授,在她的耐心指導
下,讓我在研究的這條道路上能順利的往前邁進。老師不僅指引我研究的方向,
也不斷的協助我突破實驗上的困境,同時嚴格要求論文的撰寫,使其精益求精。
這段求學時間,老師的支持和鼓勵,讓我有勇氣堅持到最後,逐步完成我的夢想。
誠摯的感謝梁展榮博士及劉振偉博士在實驗過程中,給予諸多的建議與提 點,減少我可能嘗試錯誤的時間,同時也感謝這些年來一起在實驗室互相支持的 夥伴─顏郁秀醫師、蒲啟明醫師、沈紋君博士、明憲、又溱、雅君、柏志、俊威、
哲宇、紫琳及奕潔等,也因為有您們的陪伴,讓我的研究生活多采多姿。
此外,感謝長庚的同事在工作上的體諒與協助,讓我無後顧之憂。更重要的 是要感謝我的家人在這段時間包容,尤其在工作和求學同時要兼顧的這段重要時 刻,父親和先生對家庭事務的分擔和關懷,以及兩位可愛小孩對忙碌媽媽的體諒,
都是讓我放心完成學業重要的動力和支持。
承蒙吳建春教授、江美治教授、陳永祥教授與林豐彥教授於百忙中撥冗參與 論文及口試指導,在此致上最誠摯的謝意。
最後還是感謝所有協助過我的朋友,讓我能夠順利完成學業。未來希望在這 條學術研究的道路上,可以跟指導教授和許多前輩看齊,期許自己在這浩瀚的醫 學領域中,不斷地讓研究有所精進和突破。
宋 欣 錦 謹誌於 國立台灣大學 解剖暨細胞生物學研究所 中華民國一零六年七月
doi:10.6342/NTU201701930
III
中文摘要
在肺臟相關的發炎疾病中,白血球會經由與呼吸道上皮細胞的細胞黏附因子 黏著而移動,因此細胞黏附因子對發炎疾病具有重要的功能。Eupafolin 是一種類 黃酮,也是鴨舌癀(Phyla nodiflora)中的主要活性物質,具有抗發炎的能力。另外,
山苦瓜萃取物也具有許多藥理上的活性。本文主要目的在探討,在經過 TNF-α 刺 激肺泡上皮細胞、C57BL/6 小鼠以及 miRNA-221/222 基因剔除小鼠中,eupafolin 及山苦瓜萃取物對細胞黏附因子表現的影響。研究中首先利用西方點墨法及免疫 螢光染色法,觀察 eupafolin 及山苦瓜萃取物對 TNF-α 刺激 A549 細胞後 ICAM-1 及相關蛋白表現的影響。另外小鼠以腹腔注射 eupafolin 3 天(第一部分),或是給予 C57BL/6 小鼠及 miRNA-221/222 基因剔除小鼠口服 5 天山苦瓜萃取物(第二部分) 後, 再以插管方式給予 TNF-α 1 天後取出肺臟。接著再以西方點墨法及組織免疫染 色觀察 ICAM-1 表現的改變。第一部分實驗結果顯示,eupafolin 確實可降低因 TNF-α 刺激引起的 ICAM-1 表現,而此作用是經由抑制 ERK1/2、JNK、p38 和 AKT/PI3K 的磷酸化。然而,加入 p38 和 PI3K 的抑制劑並不會改變 ICAM-1 的表 現。再者,eupafolin 同時降低了 TNF-α 所引起之 NF-κB 的活化及核轉移。在小鼠 肺臟組織中,受 TNF-α 刺激而表現量增加的 ICAM-1 會受到 eupafolin 的抑制。第 二部分實驗結果顯示,在 A549 細胞中,山苦瓜萃取物確實可經由抑制
PI3K/AKT/NF-κB/IκB 的磷酸化作用而減緩因 TNF-α 引起的 ICAM-1 表現,並且減 少了白血球的黏附作用。除此之外,山苦瓜萃取物也會降低內生性的 ICAM-1 表 現且會增加 miRNA -221/-222 的表現。讓細胞過度表現 miRNA 222 也可降低 PI3K/AKT/NF-κB/IκB 的磷酸化及 ICAM-1 表現量與白血球的黏附作用。另外,在 小鼠肺臟組織中,山苦瓜萃取物可抑制 TNF-α 刺激或沒刺激而表現的 ICAM-1 以 及增加 miRNA -221/-222 的表現;但並不影響 miRNA-221/222 基因剔除小鼠之 miRNA-221/-222 但些微影響 ICAM-1 的表現。此結果顯示,eupafolin 及山苦瓜萃 取物在細胞及動物實驗中皆可降低 ICAM-1 的表現。Eupafolin 可降低因 TNF-α 引 起的 ICAM-1 表現以及白血球的黏附作用,且是經由抑制 AKT/ERK1/2/JNK 的磷 酸化作用以及 NF-κB 的核轉移所致;而山苦瓜萃取物則是經由
miR-221/-222/PI3K/AKT/NF-κB 這條路徑來控制。因此,eupafolin 及山苦瓜萃取物 也提供了另一種治療肺臟發炎相關疾病的藥物的另一種新選擇。
doi:10.6342/NTU201701930
IV
英文摘要
The deregulation of cell adhesion molecules associated with the
epithelium-leukocyte interaction plays the important role in the pathogenesis of lung airway inflammatory disorders. Eupafolin, a major bioactive compound found in Phyla nodiflora, has been reported to have the anti-inflammatory property. In addition, the
extracts from wild bitter gourd fruit (WBGE) also possess numerous pharmacological activities. The purpose of this study was to investigate the effects of eupafolin or WBGE on intercellular adhesion molecule-1 (ICAM-1) expression in epithelial cells, C57BL/6 wild-type (WT) mice or microRNA (miR)-221/-222 knockout (KO) mice with or without tumor necrosis factor-α treatment and the related mechanisms. At first, the effects of eupafolin and WBGE on ICAM-1 expression and the related signals in A549 cells were examined by western blot and immunofluorescent staining. The part of the manuscript is the mice were injected intraperitoneally with or without eupafolin and then left untreated or injected intratracheally with TNF-α. The part section of them, WT mice and miR-221/-222 KO mice were orally treated with or without WBGE and then left untreated or injected intratracheally with TNF-α. The expression levels and patterns of ICAM-1 in the lung tissues were examined by western blot and
immunohistochemical staining. In part one, eupafolin pretreatment reduced the TNF-α-induced ICAM-1 expression and also the ERK, JNK, p38, and AKT/ PI3K phosphorylation. However, the increase in ICAM-1 expression with TNF-α treatment was unaffected by p38 and PI3K inhibitors. Moreover, eupafolin decreased the TNF-α-induced NF-κB p65 activation and its nuclear translocation. Furthermore,
eupafolin reduced ICAM-1 expression in the lung tissues of TNF-α-treated mice. In part two, WBGE significantly decreased the TNF-α-induced ICAM-1 expression in A549 cells through the inhibition of PI3K/ AKT/ NF-κB /IκB phosphorylation and decreased
doi:10.6342/NTU201701930
V
leukocyte adhesion. In addition, WBGE reduced endogenous ICAM-1 expression and upregulated miR-221/-222 expression. The overexpression of miR-222 decreased PI3K/AKT/NF-κB/IκB phosphorylation and ICAM-1 expression, which resulted in reducing monocyte adhesion. Moreover, WBGE reduced ICAM-1 expression in lung tissues of WT mice with or without TNF-α treatment and upregulated miR-221/222.
WBGE did not affect the miR-221/-222 level and had little effect on ICAM-1
expression in miR-221/-222 KO mice. These results suggest that eupafolin and WBGE reduced ICAM-1 expression both under in vitro and in vivo conditions. The protective effects of eupafolin were mediated via AKT/ERK1/2/JNK phosphorylation and nuclear translocation of NF-κB. Furthermore, WBGE were mediated partly through the
miR-221/-222/PI3K/AKT/NF-κB pathway. Therefore, eupafolin and WBGE may represent novel therapeutic agents targeting epithelial activation in lung inflammation.
doi:10.6342/NTU201701930
VI
簡介
The dysregulated, chronic inflammation is a critical risk factor in the initiation and development of pulmonary diseases, such as asthma and chronic obstructive pulmonary disease (Barnes, 2011; Lee and Yang, 2013). The expression of adhesion molecules on airway epithelial cells was important for selective recruitment of effector cells onto epithelial cells (Chen et al., 2001). Intercellular adhesion molecule-1 (ICAM-1) which is an adhesion molecule, has long been known for its importance in mediating cell-cell interactions. Its expression is increased by the stimulation of inflammatory cytokines and enhances adhered leukocytes that migrated across the vascular endothelial cells and then interacted with epithelial cells at sites of inflammatory airways (Lee and Yang, 2013). The clinical study has also showed that the ICAM-1 level was much higher in plasma of patients with pulmonary diseases (Demir et al., 2002). ICAM-1 was shown to be upregulated in epithelial cells by the stimulation of several pro-inflammatory
cytokines such as tumor necrosis factor- (TNF-), interleukin-1, and interferon-
(IFN-) (Kim et al., 2008; Wang et al., 2014). Currently available therapies for inflammation include corticosteroids and CXC chemokine receptor antagonists
(Durham et al., 2015). However, many of these agents have a number of serious adverse effects. Modulation of the ICAM-1 expression provides a rationale for the development of therapeutic agents against a variety of inflammatory lung diseases.
Herbs have been extensively used in foods and in traditional medicines in oriental countries for centuries. The active components of herbs may provide a useful platform for the development of effective pharmacological agents. Phyla nodiflora, which belongs to the Verbenaceae family, is a common ingredient of herbal tea for the treatment of inflammation, menstrual disorders, and infectious diseases (Yang et al.,
doi:10.6342/NTU201701930
VII
1998). Eupafolin (6-methoxy-5, 7, 3′, 4′-tetrahydroxyflavone), the major bioactive flavonoid isolated from the dried aerial parts of P. nodiflora, inhibits the NO release in LPS-stimulated macrophages (Maas et al., 2011). These results indicate that eupafolin has antioxidative and anti-inflammatory effects on macrophages (Ko et al., 2014; Lai et al., 2011; Maas et al., 2011).
Wild bitter gourds (WBG, Momordica charantia L.) are consumed as a vegetable and have been used as a traditional herbal medicine in Asia (Chao et al., 2014). The noticeable pharmacological properties of WBG fruit extract (WBGE) are anti-diabetic (Chaturvedi, 2012), anti-inflammatory (Chao et al., 2014, Lii et al., 2009), anti-tumor (Bai et al., 2016; Somasagara et al., 2015) and anti-oxidative actions (Lu et al., 2014).
However, the anti-inflammatory effects of both eupafolin and WBGE on human lung epithelial cells and the underlying mechanisms have not been investigated.
The intracellular signaling pathways by which TNF- causes ICAM-1 expression are, in the main, not well understood, but certain have been proposed, including
mitogen-activated protein kinases (MAPKs), PI3K/AKT and transcriptional factors (Binion et al., 2009; Chen et al., 2001). In addition, evidence also suggested that
miRNA was involved in lung inflammation (Alipoor et al., 2016). MiRNAs, which are a new class of non-coding small RNAs, are 19-25 nucleotides in length (Alipoor et
al.2016). They target mRNAs through complementarity between the miRNAs and the 3'-untranslated regions (3'UTRs) of target mRNAs, which causes either mRNA cleavage or translational suppression and results in gene silencing (Hu et al., 2010).
Additionally, recent studies have shown that miR-221 or miR-222 can suppress
ICAM-1 translation and regulate ICAM-1 expression (Hu et al., 2010; Gong et al., 2011;
Jansen et al., 2015). However, the relationship between miRNAs and ICAM-1 in lung inflammation is still unclear.
doi:10.6342/NTU201701930
VIII
Therefore, a better understanding of the effects of eupafolin and WBGE on ICAM-1 expression and the mechanisms might provide important insights into the prevention of airway inflammation. Accordingly, this study is the initial study to
elucidate the anti-inflammatory effects of both eupafolin (a bioactive component of herb) and WBGE (a common vegetable in Asia), and the related mechanisms on
TNF-α-treated A549 cells and mice.
doi:10.6342/NTU201701930
i
目錄
口試委員會審定書...I 誌 謝 ...II 中文摘要 ... III 英文摘要 ... IV 簡介 ... VI
PART I... 1
中文摘要 ... 1
英文摘要 ... 2
第一章 簡介 ... 3
第二章 材料與方法 ... 5
2.1. Extraction and purification of eupafolin ... 5
2.2. Cell culture ... 5
2.3. Preparation of cell lysates and Western blot analysis ... 6
2.4. Immunofluorescent staining of ICAM-1 and NF-κB p65 ... 7
2.5. siRNA knockdown of ERK, JNK, and p65 ... 8
2.6. Preparation of nuclear extracts and electrophoretic mobility-shift assay (EMSA) ... 8
2.7. Epithelial cell–leukocyte adhesion assay ... 9
2.8. Animal care and experimental procedures ... 9
2.9. Immunohistochemistry ... 10
2.10. Statistical analysis ... 10
第三章 結果 ... 12
3.1. Eupafolin reduces TNF-α-induced upregulation of ICAM-1 in A549 cells ... 12
3.2. The inhibition of ERK1/2 and JNK phosphorylation mediates eupafolin-increased reduction in TNF-α-induced ICAM-1 expression ... 12
3.3. The inhibition of AKT phosphorylation mediates eupafolin-increased reduction in TNF-α-induced ICAM-1 expression ... 13
3.4. The inhibition of NF-κB activation and NF-κB p65 translocation mediates eupafolin-reduced ICAM-1 expression in TNF-α-treated A549 cells ... 14
3.5. TNF-α-induced ICAM-1 expression was mediated by AKT/ERK1/2/JNK/NF-B signaling pathway ... 15
3.6. Eupafolin suppressed the adhesion of monocytes to TNF-α-stimulated A549 cells . 16 3.7. Eupafolin reduces ICAM-1 expression in lung tissues in TNF-α-treated mice ... 17
doi:10.6342/NTU201701930
ii
第四章 討論 ... 18
參考資料 ... 23
附圖 ... 29
Figure 1: Chemical structure of eupafolin. ... 29
Figure 2: The effects of TNF-α-induced ICAM-1 expression in A549 cells. ... 30
Figure 3: The effects of eupafolin on ICAM-1 expression in TNF-α-treated A549 cells. .. 31
Figure 4: The role of MAPKs activation on eupafolin-reduced ICAM-1 expression in TNF-α-treated A549 cells. ... 33
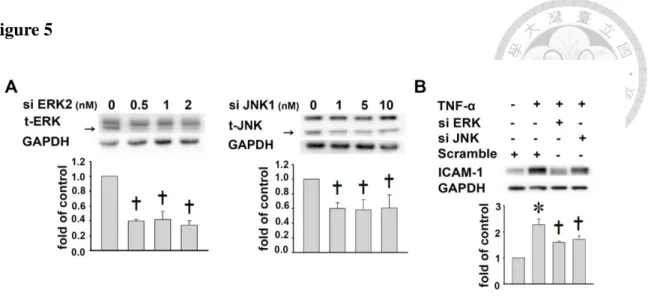
Figure 5: The effects of MAPKs phosphorylation on ICAM-1 expression in TNF-α-treated A549 cells. ... 35
Figure 6: The roles of AKT/PI3K activation on eupafolin-reduced ICAM-1 expression in TNF-α-treated A549 cells. ... 36
Figure 7: The effects of eupafolin on NF-κB and IκB phosphorylation in TNF-α-treated A549 cells. ... 38
Figure 8: The effects of eupafolin on NF-κB expression in TNF-α-stimulated A549 cells.39 Figure 9: The effects of NF-κB on ICAM-1 expression in TNF-α-stimulated A549 cells. 40 Figure 10: The effects of eupafolin on the nuclear activation of NF-κB in TNF-α-stimulated A549 cells. ... 41
Figure 11: The crosstalk among AKT, ERK1/2, JNK and NF-B signaling pathways in TNF-α-treated A549 cells. ... 42
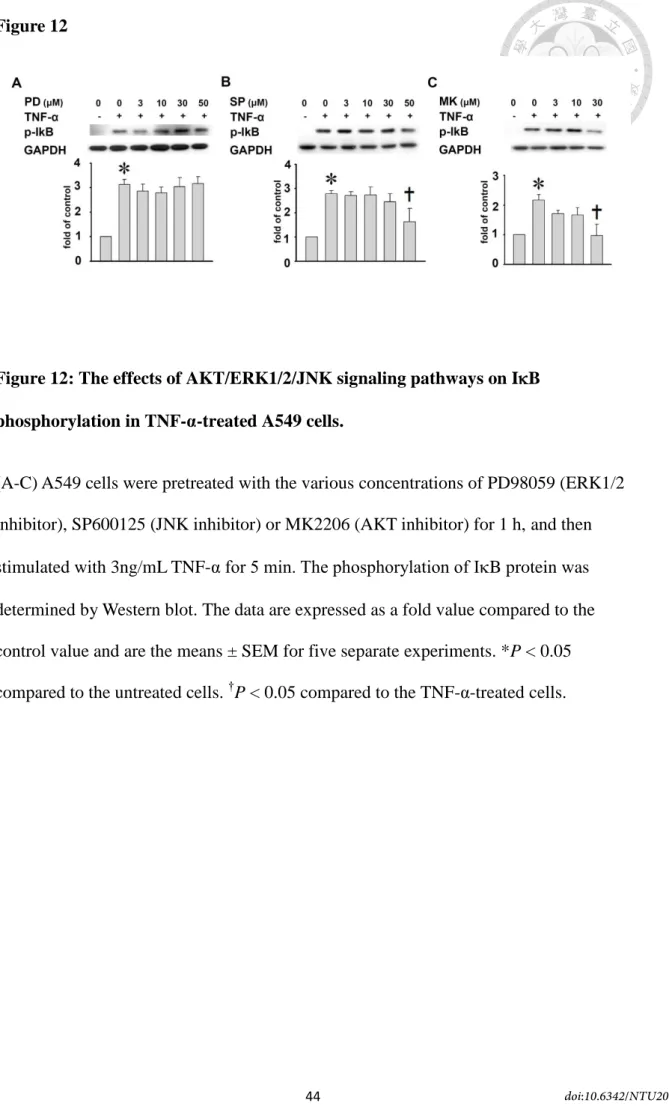
Figure 12: The effects of AKT/ERK1/2/JNK signaling pathways on IB phosphorylation in TNF-α-treated A549 cells. ... 44
Figure 13: The effects of eupafolin on the adhesion of U937 cells to TNF-α-treated A549 cells. ... 45
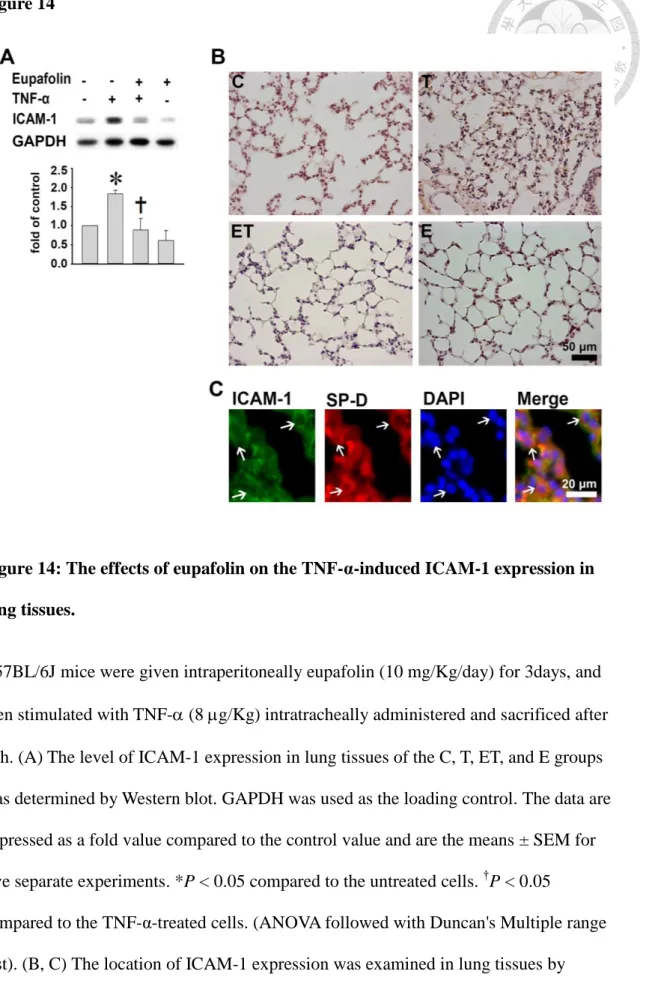
Figure 14: The effects of eupafolin on the TNF-α-induced ICAM-1 expression in lung tissues. ... 47
Figure 15: A summary diagram showing that eupafolin reduced ICAM-1 expression in TNF-α-treated A549 cells through the inhibition of the phosphorylation of AKT, ERK, and JNK as well as the inactivation of transcription factor NF-B. ... 49
PART II ... 50
中文摘要 ... 50
英文摘要 ... 51
第一章 簡介 ... 53
第二章 材料與方法 ... 56
2.1 Extraction of WBG (WBGE) ... 56
2.2 Analysis of WBGE by HPLC ... 56
2.3 Cell culture ... 57
doi:10.6342/NTU201701930
iii
2.4 Preparation of cell lysates and Western blot analysis ... 57
2.5 Immunocytochemical localization of ICAM-1 and NF-κB p65 ... 58
2.6 Preparation of cytoplasmic and nuclear extracts for Western blotting ... 58
2.7 Chromatin immunoprecipitation assay ... 58
2.8 RNA preparation and real-time PCR ... 59
2.9 Overexpression of miR-221/-222 ... 60
2.10 Cell adhesion assay ... 60
2.11 Luciferase reporter assay ... 61
2.12 Mouse model, diets, and experimental procedures ... 61
2.13 Immunohistochemistry ... 62
2.14 Statistical analysis ... 62
第三章 結果 ... 63
3.1 Characterization of WBGE ... 63
3.2 WBGE decreased the TNF-α-induced ICAM-1 expression in A549 cells ... 63
3.3 The inhibition of PI3K/AKT phosphorylation mediates the reduction in ICAM-1 expression by WBGE in TNF-α-treated A549 cells ... 64
3.4 The inhibition of NF-κB p65 activation and translocation mediates WBGE-reduced ICAM-1 expression in TNF-α-treated A549 cells ... 65
3.5 WBGE reductions in endogenous ICAM-1 expression in A549 cells involves miR-222 upregulation ... 66
3.6 WBGE reduces ICAM-1 expression in lung tissues of TNF-α-treated WT mice ... 68
第四章 討論 ... 69
參考資料 ... 74
附圖 ... 78
Figure 1: Characterization of WBGE ... 78
Figure 2: The effects of WBGE and charantin on ICAM-1 expression in TNF-α-treated A549 cells ... 79
Figure 3: The effects of WBGE and charantin on ICAM-1 expression patterns in A549 cells ... 80
Figure 4: The effects of WBGE on ICAM-1 promoter activity in A549 cells. ... 81
Figure 5: The effects of WBGE on the adhesion of U937 cells to TNF-α-treated A549 cells ... 82
Figure 6: The effects of WBGE on MAPKs phosphorylation in TNF-α-treated A549 cells. ... 84
Figure 7: The effects of PI3K/AKT phosphorylation on WBGE-reduced ICAM-1 expression in TNF-α-treated A549 cells ... 85
doi:10.6342/NTU201701930
iv
Figure 8: The effects of PI3K/AKT on the adhesion of U937 cells to TNF-α-treated A549 cells ... 86 Figure 9: The effects of WBGE on NF-κB and IκB phosphorylation in TNF-α-treated A549 cells. ... 87 Figure 10: The effects of WBGE on NF-κB activation in TNF-α-treated A549 cells ... 88 Figure 11: The effects of NF-κB on WBGE-reduced ICAM-1 expression in TNF-α-treated A549 cells ... 90 Figure 12: The effects of NF-κB on the adhesion of U937 cells to TNF-α-treated A549 cells ... 91 Figure 13: The levels of miRNA-221 and -222 expression in TNF-α-treated A549 cells .. 92 Figure 14: The effects of pre-miRNA-221/-222 on the TNF-α-treated A549 cells. ... 93 Figure 15: The effects of pre-miRNA-221/-222 on the adhesion of U937 cells to the TNF-α-treated A549 cells ... 95 Figure 16: The effects of pre-miRNA-221/-222 on the A549 cells ... 97 Figure 17: The effects of pre-miRNA-221/-222 on the adhesion of U937 cells to the A549 cells ... 98 Figure 18: The effects of p-AKT, p-PI3K, p-p65 on endogenous expression of ICAM-1 in A549 cells ... 99 Figure 19: The effects of WBGE on the TNF-α-induced miRNA-221/-222 expression in lung tissues ... 100 Figure 20: The effects of WBGE on the TNF-α-induced ICAM-1 expression in lung tissues ... 101 結論 ... 103 未來展望 ... 105
doi:10.6342/NTU201701930
1
PART I
中文摘要
Eupafolin 是一種類黃酮,也是鴨舌癀(Phyla nodiflora.)中的主要活性物質。在 肺臟相關的發炎疾病中,白血球會經由與呼吸道上皮細胞的細胞黏附因子黏著而 移動,因此細胞黏附因子對發炎疾病具有重要的功能。本文研究主要目的在探討,
eupafolin 對肺臟上皮細胞被腫瘤壞死因子(tumor necrosis factor-α, TNF-α)刺激後所 引起之細胞黏附因子(intercellular cell adhesion molecule-1, ICAM-1)的表現量影響 及其作用機轉。研究中利用西方點墨法及免疫螢光染色實驗,觀察 TNF-α 刺激人 類呼吸道上皮細胞(A549 cells)後 ICAM-1 的表現以及影響的相關蛋白表現。在動物 實驗部分,小鼠以腹腔注射 eupafolin 3 天,再以插管方式給予 TNF-α 1 天後取出 肺臟。然後利用西方點墨法及組織免疫染色觀察 eupafolin 對 ICAM-1 表現的影 響。細胞實驗結果顯示,eupafolin 可降低因 TNF-α 引起的 ICAM-1 表現,而此作 用是經過抑制 ERK1/2、JNK、p38 和 AKT/PI3K 的磷酸化作用。然而,加入 p38 和 PI3K 的抑制劑並不會改變 ICAM-1 的表現。再者,eupafolin 同時降低了 TNF-α 所 引起之 NF-κB 的活化及核轉移。動物的實驗結果顯示,在小鼠肺臟組織中,受 TNF-α 刺激而表現量增加的 ICAM-1 會受到 eupafolin 的抑制。Eupafolin 可降低因 TNF-α 引起的 ICAM-1 表現以及白血球的黏附作用,且是經由抑制 AKT/ERK1/2/JNK 的 磷酸化作用以及 NF-κB 的核轉移所致。因此,eupafolin 對呼吸道上皮細胞具有抗 發炎的能力;而此實驗結果也提供了另一種治療肺臟發炎相關疾病的藥物的新選 擇。
關鍵字: eupafolin; 細胞黏附因子-1; MAPKs; AKT; 發炎
doi:10.6342/NTU201701930
2
英文摘要
Eupafolin, a major bioactive compound found in Phyla nodiflora, has the
anti-inflammatory property. Upregulation of cell adhesion molecules in the lung airway epithelium is associated with the epithelium-leukocyte interaction and plays a critical role in the pathogenesis of lung airway inflammatory disorders. To investigate the effects of eupafolin on tumor necrosis factor-α (TNF-α)-induced intercellular cell adhesion molecule-1 (ICAM-1) expression in A549 human lung airway epithelial cells and the underlying mechanisms. The effect of eupafolin on ICAM-1 expression in A549 cells were examined by Western blotting and immunofluorescent staining. The mice were injected intraperitoneally with or without eupafolin and then were left untreated or were injected intratracheally with TNF-α. To detect the effect of eupafolin on ICAM-1 expression, the lung tissues were also examined by Western blotting and
immunohistochemical staining. Eupafolin pretreatment reduced the TNF-α-induced ICAM-1 expression and also the ERK1/2, JNK, p38, and AKT/PI3K phosphorylation.
However, the increase in ICAM-1 expression with TNF-α treatment was unaffected by p38 and PI3K inhibitors. Eupafolin decreased the TNF-α-induced NF-κB p65 activation and its nuclear translocation. Furthermore, eupafolin reduced ICAM-1 expression in the lung tissues of TNF-α-treated mice. Eupafolin exerts its anti-inflammatory activity by suppressing the TNF-α-induced ICAM-1 expression and subsequent monocyte adhesion via AKT/ERK1/2/JNK phosphorylation and nuclear translocation of NF-κB p65. These results suggest that eupafolin may represent a novel therapeutic agent targeting
epithelial activation in lung inflammation.
Keywords: eupafolin; intercellular cell adhesion molecule-1; MAPKs; AKT;
inflammation
doi:10.6342/NTU201701930
3
第一章 簡介
The dysregulated, chronic inflammation is a critical risk factor in the initiation and development of pulmonary diseases, such as asthma and chronic obstructive pulmonary disease (Barnes, 2011; Lee and Yang, 2013). The increased adhesion molecules and inflammatory chemokines on airway epithelial cells would be important in the selective recruitment of effector cells onto epithelial cells (Rosseau et al., 2000). Among the cell adhesion molecules, intercellular cell adhesion molecule-1 (ICAM-1), a cell surface glycoprotein that belongs to the immunoglobulin supergene family, is overexpressed on airway epithelial cells during inflammation, which enhances the adherence and
infiltration of leukocytes across the endothelial cells and epithelial cells at sites of inflammatory airways. The clinical study has also showed that the ICAM-1 level was much higher in plasma of patients with pulmonary diseases (Demir et al., 2002).
ICAM-1 was shown to be upregulated in epithelial cells by the stimulation of several pro-inflammatory cytokines such as tumor necrosis factor- (TNF-), interleukin-1, and interferon- (IFN-) (Kim et al., 2008; Wang et al., 2014). Modulation of the ICAM-1 expression provides a rationale for the development of therapeutic agents against a variety of inflammatory lung diseases.
Herbs have been extensively used in foods and in traditional medicines in oriental countries for centuries. The active components of herbs may provide a useful platform for the development of effective pharmacological agents. Phyla nodiflora, which belongs to the Verbenaceae family, is a common ingredient of herbal tea for the treatment of inflammation, menstrual disorders, and infectious diseases (Yang et al., 1998). Eupafolin (6-methoxy-5, 7, 3′, 4′-tetrahydroxyflavone; Fig. 1), the major bioactive flavonoid isolated from the dried aerial parts of P. nodiflora, inhibits the NO
doi:10.6342/NTU201701930
4
release in LPS-stimulated macrophages (Maas et al., 2011). Eupafolin also reduced pro-inflammatory inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) expression in these cells (Lai et al., 2011). In addition, eupafolin exerts anti-tumor activity by the inhibition of cancer cell proliferation (Abe et al., 2002; Liu et al., 2014). These results indicate that eupafolin has antioxidative and anti-inflammatory effects on macrophages (Ko et al., 2014; Lai et al., 2011; Maas et al., 2011). However, the anti-inflammatory effects of eupafolin on human lung epithelial cells and the related mechanisms have not been investigated. The intracellular signaling pathways by which TNF- causes ICAM-1 expression are, in the main, not well understood, but certain have been proposed, including mitogen-activated protein kinases (MAPKs), PI3K/AKT and transcriptional factors (Binion et al., 2009; Chen et al., 2001). Little is known about the effects of eupafolin on ICAM-1 expression and the mechanisms of these effects, and a better understanding of this might provide important insights into the prevention of airway inflammation. In the present study, we showed that eupafolin reduced
AKT/ERK1/2/JNK phosphorylation in TNF-α-treated A549 cells (a human airway type II alveolar epithelial cells), resulting in the reduced activation of NF-κB in the ICAM-1 promoter, which is followed by the reduction of ICAM-1 expression and monocyte adhesion. Moreover, we found that TNF--induced ICAM-1 expression was also suppressed in lung tissues of C57BL/6J mice by eupafolin treatment.
doi:10.6342/NTU201701930
5
第二章 材料與方法
2.1. Extraction and purification of eupafolin
P. nodiflora was collected from a local farm (Tainan, Taiwan). The authenticity of the plant species was identified by a pharmacognosist, Professor Ih-sheng Chen, and stored as a voucher specimen (2007-02-PNM) in the Herbarium of the Department of Fragrance and Cosmetic Science, Kaohsiung Medical University, Kaohsiung, Taiwan.
Eupafolin was purified from the plant as described previously (Ko et al., 2014). In brief, dried aerial part of P. nodiflora was chopped and immersed in methanol at room
temperature. The methanol extract was filtered and concentrated by rotary vacuum evaporation. The crude methanolic extract was suspended in water and successively portioned with an equivalent volume of n-hexane, chloroform, and ethyl acetate. The partition was subjected to silica gel column chromatography, purified with a mixture of n-hexane and ethyl acetate, and followed by purification with methanol. The purified fraction was recrystallized to obtain eupafolin. Eupafolin was stored at −20°C until further use. The chemical structure and purity of eupafolin (Fig. 1) was identified by
1H-NMR. Eupafolin was dissolved in dimethylsulfoxide (DMSO) and then further diluted with the assay medium (1-50 µM). Solvent controls with DMSO were carried out in each assay.
2.2. Cell culture
Human lung epithelial alveolar cells (A549 cells) and human monocytic leukemia cells (U937 cells) were obtained from American Type Culture Collection (Manassas, VA, USA). A549 cells were cultured in Dulbecoo’s Modified Eagle Medium (DMEM)
(Gibco, Life Technology, Grand Island, NY, USA) supplemented with 10% fetal bovine
doi:10.6342/NTU201701930
6
serum(FBS) and 1% penicillin-streptomycin. U937 cells were cultured in RPMI 1640 medium (M.A. Bioproducts, Walkersville, MD, USA) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin. These cells were grown at 37℃ in a humidified atmosphere of 95% air and 5% CO2.The
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay was used to measure cell viability. The principle of this assay is that mitochondrial
dehydrogenase in viable cells reduces MTT to a blue formazan. Briefly, cells were grown in 96-well plates and incubated with various concentrations of TNF-α (0.1-10 ng/mL) or eupafolin (1-50 μM) for 24 h, then 100 μL of MTT (0.5 mg/mL) was added to each well and incubation continued at 37°C for an additional 4 h. The medium was then carefully removed, so as not to disturb the formazan crystals formed. Dimethyl sulphoxide (DMSO; 100 μL), which solubilizes the formazan crystals, was added to each well and the absorbance of the solubilized blue formazan read at 530 nm (reaction) and 690 nm (background) using a DIAS Microplate Reader (Dynex Technologies, U.S.A.). The reduction in optical density caused by cytokine and eupafolin was used as a measurement of cell viability, normalized to cells incubated in control medium, which were considered 100% viable. In the present study, A549 cells treated with 0.1-10 ng/mL of TNF-α or 1-50 μM of eupafolin for 24 h did not cause the cytotoxicity.
2.3. Preparation of cell lysates and Western blot analysis
Western blot analyses were performed as described previously (Liang et al., 2013).
A549 cells were incubated with various concentrations (0.1, 0.3, 1, 3, or 10 ng/mL) of TNF-α at different time intervals (4, 8, 16, or 24 h). Cell lysates were prepared by incubating cells in a lysis buffer (20 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton X-100, 1 mM phenylmethylsulfonyl fluoride, pH 7.4) for 1 h at 4 °C.
doi:10.6342/NTU201701930
7
The cell lysate was centrifuged at 4000 g for 30 min at 4 °C and the supernatant retained.
Equal amounts of the supernatant (20 μg of protein) were separated on a 10% sodium dodecyl sulfate (SDS)-PAGE and blotted on to PVDF membranes. The membranes were blocked nonspecific binding of antibody in 3% nonfat milk in 0.1 M phosphate buffer for 1 h at room temperature (RT). Immunoreaction was performed with the following primary antibodies: rabbit antibodies against anti-ICAM-1(1:5000, Santa Cruz Biotechnology, Dallas, TX, USA), anti-phospho ERK1/2 (1:10000, Cell Signaling, Danvers, MA, USA), anti-phospho p38 (1:1000, Santa Cruz Biotechnology),
anti-phospho JNK (1:1000, Cell Signaling), anti-phospho PI3K (1:1000,
EnoGene Biotech, New York, NY, USA), anti-phospho AKT (1:1000, EnoGene Biotech), anti-phospho p65 (1:1000, Epitomics, Burlingame, CA, USA), anti-phospho IB (1:1000, Cell Signaling), anti-total p65 (1:1000, Epitomics), anti-total ERK1/2 (1:10000, Cell signaling), and anti-total JNK (1:1000, OriGene, Rockville, MD, USA) at 4 °C for overnight. The membranes were then incubated for 1 h at room temperature with HRP-conjugated goat anti-rabbit IgG antibodies (1:2000, Santa Cruz
Biotechnology). The immunoreactive bands were detected using the
Chemiluminescence Reagent Plus (NEN, Boston, MA, USA). The intensity of each band was quantified using a densitometer. Antibodies against GAPDH (1:5000, Santa Cruz Biotechnology) or hnRNP (1:10000, Abcam, Cambridge, MA, USA) were designed as the internal standard.
2.4. Immunofluorescent staining of ICAM-1 and NF-κB p65
To examine ICAM-1 or NF-κB p65 expression in situ, confluent A549 cells (controls or cells treated for 24 h with eupafolin) on cover slips were incubated in the absence or presence of 3 ng/mL TNF-α for 4h at 37 °C and fixed in 4%
doi:10.6342/NTU201701930
8
paraformaldehyde in PBS, pH 7.4, for 15 min at RT. The cells were then reacted at 4 °C with rabbit anti-human ICAM-1 (1:100 dilution, Abcam, Cambridge, MA, USA) or anti-human NF-κB p65 antibodies (1:100 dilution, Epitomics, Burlingame, CA, USA) for overnight. After washes, the coverslips were incubated with DyLight 488 goat anti-rabbit polyclonal antibody (1:200 dilution, Abcam) for 1 h at 37 °C and observed by a fluorescence microscope.
2.5. siRNA knockdown of ERK, JNK, and p65
To perform the knockdown ERK, JNK, and p65 by siRNA, A549 cells were transfected with siRNA specifically targeting ERK, JNK, p65, or scrambled gene (ON-TARGET plus SMARTpool RNA duplexes; Thermo Scientific, Waltham, MA, USA). For transfection, siRNA (1 nM) was transfected into A549 cells (106)for 48 h according to the manufacturer's instruction. The siRNA results were evaluated by Western blotting.
2.6. Preparation of nuclear extracts and electrophoretic mobility-shift assay (EMSA)
The nuclear protein extracts and the EMSA conditions were prepared as described previously (Liang et al., 2013). Nuclear proteins were extracted using NE-PER reagent (Pierce, Rockford, IL, USA) according to the manufacturer's protocol. The NF-κB binding activity of equal amounts (10 μg) of nuclear protein was performed using the LightShift chemiluminescence EMSA kit (Pierce). The synthetic double-stranded oligonucleotides used as the probes in the gel-shift assay were
5′-AGTTGAGGGGACTTTCCCAGGC-3′ and 3′-TCAACTCCCCTGAAAGGGTCCG-5′.
doi:10.6342/NTU201701930
9
2.7. Epithelial cell–leukocyte adhesion assay
A549 cells, grown in 24-well dish, were pretreated with or without eupafolin for 24h and treated with TNF- for 4 h at 37℃, then washed three times with PBS. U937 cells, were labeled with 10 mM of BCECF-AM
(2',7'-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein acetoxymethyl, Boehringer Mannheim, Mannheim, Germany) for 1 h at 37 °C. Labeled U937 cells (106) were added to A549 cells (106) and incubation continued for 1 h. Nonadherent cells were removed by gentle washes with PBS. The number of U937 cells adhering to A549 cells was counted in six randomly selected images captured by fluorescent microscope (Zeiss) for each experiment.
2.8. Animal care and experimental procedures
All procedures involving experimental animals were performed in accordance with the guidelines for animal care of the National Taiwan University (No. 20130175) and complied with the Guide for the Care and Use of Laboratory Animals, NIH publication No. 86–23, revised 1985. Male 8-week-old C57BL6 mice, weighing between 25 and 35 g, were purchased from the National Taiwan University (Taipei, Taiwan). The mice were injected intraperitoneally (ip) with or without eupafolin (10 mg/2mL DMSO/Kg body weight/day) for 3 days and then were left untreated or were injected intratracheally with TNF-α (8 μg/Kg) for the next 1 day. Some mice were injected ip with an
equivalent volume of the DMSO vehicle (2mL/Kg body weight) as the control. They were then anesthetized by ip injection of 30-40 mg/kg pentobarbital and sacrificed. A part of lung tissues was immersion-fixed with 4% buffered paraformaldehyde and paraffin-embedded for immunohistochemistry; the remaining larger portion was
doi:10.6342/NTU201701930
10
immediately frozen in liquid nitrogen for protein isolation to detect the ICAM-1 level by Western blotting.
2.9. Immunohistochemistry
The sample size of lung is 5 x 5 x 5 mm. Sections (5 µm) were cut from the paraffin blocks. To determine the ICAM-1 expression in lung tissues, the sections were examined by immunostaining with ICAM-1 antibodies (1:100 dilution, Abcam). The sections were then incubated with biotin-conjugated goat anti-rabbit IgG (1:200 dilution, Vector lab, Cambridgeshire, UK) for 1h at room temperature (RT). Finally, the sections were stained with 3,3-diaminobenzidine tetrahydrochloride (DAB), counterstained with hematoxylin. To examine whether ICAM-1 was associated with type II alveolar
epithelial cells, the section was examined by double immunofluorescent staining for, respectively, ICAM-1 (1:100, BioLegend, CA, USA) and SP-D (marker for type II alveolar epithelial cells, 1:100, Bioss, Beijing, China) for 1h at RT. After washed with PBS, the section was then incubated with Alexa Fluro 488 conjugated goat anti-rat IgG (1:500, BioLegend, green) for ICAM-1 and Dylight 594 conjugated donkey anti-rabbit IgG (1:500, BioLegend, red) for SP-D. Finally, the slides were counterstained with DAPI and examined by fluorescent microscope.
2.10. Statistical analysis
The data are expressed as a fold value compared to the control value and are the means ± SEM for five separate experiments unless other specified. All statistical analyses were performed with one-way ANOVA, and then followed with Duncan's Multiple range test. Analyses were done using SigmaPlot software (Systat Software, Inc., Chicago, IL, USA). *P < 0.05 compared to the untreated cells. †P < 0.05 compared
doi:10.6342/NTU201701930
11
to the TNF-α-treated cells.
doi:10.6342/NTU201701930
12
第三章 結果
3.1. Eupafolin reduces TNF-α-induced upregulation of ICAM-1 in A549 cells
A549 cells were incubated with various concentrations of TNF-α at different time intervals. ICAM-1 expression was significantly upregulated with 1 ng/mL, 3 ng/mL or 10 ng/mL of TNF-α treatment for 4 h, 8 h, 16 h, and 24 h or with 0.3 ng/mL of TNF-α for 24 h (Fig. 2). We next analyzed the effect of eupafolin on ICAM-1 expression under inflammation. A549 cells were pretreated for 24 h with 1, 3, 10, 30, or 50 μM eupafolin before incubation with 3 ng/mL TNF-α for 4 h, TNF-α-induced ICAM-1 expression was reduced (2.3±0.3, 2.1±0.2, 1.6±0.2, 0.9±0.2, 0.6±0.1 fold of control levels, respectively).
The reductions caused by the three highest concentrations were significant (P<0.05, Fig.
3A). The effect of eupafolin on ICAM-1 expression was also confirmed by immunofluorescent staining (Fig. 3B). Cells treated for 4 h with 3 ng/mL TNF-α showed strong ICAM-1 expression (T) and this effect was inhibited by pretreatment with eupafolin (50 μM, ET). According to these results, 3 ng/mL TNF-α and 50 μM eupafolin were used in all subsequent experiments to evaluate the anti-inflammatory effects and molecular mechanisms of eupafolin treatment.
3.2. The inhibition of ERK1/2 and JNK phosphorylation mediates eupafolin-increased reduction in TNF-α-induced ICAM-1 expression
Previous studies have reported that TNF-α-induced inflammation includes the production of inflammatory cytokines via the MAPK pathways (Lee et al., 2011; Lee et al., 2013). We next investigated whether TNF-α-induced ICAM-1 expression was mediated through MAPKs phosphorylation. The phosphorylation of ERK1/2, p38, and JNK in A549 cells showed a significant increase at 15-30 min of TNF-α treatment and
doi:10.6342/NTU201701930
13
followed by a decline within 60 min (P<0.05, Fig. 4A). To determine the targets that were affected by eupafolin, the cells were preincubated with eupafolin for 24 h and then incubated with TNF-α for 5, 15, 30, 45, or 60 min. Eupafolin significantly inhibited TNF-α-induced ERK1/2, p38, and JNK phosphorylation (P<0.05). In addition, the PD98059 (50M, ERK1/2 inhibitor) or SP600125 (10M, 30M, 50M, JNK inhibitor) inhibited the TNF-α-induced ICAM-1 expression (PD: 40±13% of inhibition; SP:
26±10 % for 10M, 39±13% for 30M, and 43±15% for 50M, respectively, P<0.05).
In contrast, SB203580 (p38 inhibitor) had no effects (Fig. 4B). To further confirm the involvement of ERK1/2 and JNK in the TNF-α-induced ICAM-1 expression, we used siRNA transfection to knockdown the ERK1/2 or JNK expression in A549 cells. As shown in Fig. 5A, the expression levels of ERK1/2 or JNK were significantly
downregulated by siRNA transfection (P<0.05). Moreover, cells transfected with 1 nM ERK1/2- or with JNK-specific siRNA inhibited TNF-α-induced ICAM-1 expression (1.6±0.1 and 1.7±0.1 fold of control levels, respectively) (P<0.05, Fig. 5B). These results suggest that eupafolin inhibits TNF-α-induced ICAM-1 expression partly by inhibiting TNF-α-induced ERK1/2 and JNK phosphorylation.
3.3. The inhibition of AKT phosphorylation mediates eupafolin-increased reduction in TNF-α-induced ICAM-1 expression
The phosphatidylinositol 3-kinase (PI3K)/AKT signaling pathway is reported to be involved in adhesion molecule expression in TNF--treated various cells (Choi et al., 2012; Jang et al., 2012; Oh and Kwon, 2009). To investigate whether eupafolin affects TNF--induced PI3K/AKT activation, we examined the effect of eupafolin on the TNF-α-induced PI3K/AKT in A549 cells using Western blot analysis. The expression levels of phosphorylated PI3K and AKT were gradually increased after TNF-α
doi:10.6342/NTU201701930
14
stimulation, and pretreatment with eupafolin significantly attenuated the
phosphorylation of PI3K and AKT (P<0.05, Figs. 6A and 6B). Interestingly, eupafolin pretreatment completely blocked the increase of TNF-α-induced AKT phosphorylation.
Moreover, we determined whether the activation of PI3K/AKT was involved in TNF-α induced ICAM-1 expression. As shown in Fig. 6D, pretreatment with MK2206 (AKT inhibitor) caused a significant attenuation of ICAM-1 expression in TNF-α stimulated A549 cells (34±13% of inhibition, P<0.05). In contrast, pretreatment with LY294002 (PI3K inhibitor) did not reduce the ICAM-1 expression (P<0.05, Fig. 6C). These results suggest that eupafolin inhibits TNF-α-induced ICAM-1 expression partly by inhibiting TNF-α-induced AKT phosphorylation.
3.4. The inhibition of NF-κB activation and NF-κB p65 translocation mediates eupafolin-reduced ICAM-1 expression in TNF-α-treated A549 cells
We investigated whether eupafolin reduced TNF-α-induced ICAM-1 expression via NF-κB signaling because the promoter of ICAM-1 gene contains consensus binding sites for the transcription factor (Rahman et al., 1999). At first, we examined the levels of phosphorylated NF-κB p65 in TNF--treated A549 cells by Western blotting and immunofluorescence staining. The phospho-p65 level was higher in TNF--treated A549 cells than in control cells and that eupafolin pretreatment significantly reduced the effect (P<0.05, Fig. 7A). The similar result was obtained for IκB phosphorylation (P<0.05, Fig. 7B), which is responsible for NF-κB activation (Choi et al., 2012). The results of immunofluorescent staining were consistent with the Western blot finding of NF-κB p65. Control A549 cells (C) or cells incubated only with eupafolin (E) showed no nuclear NF-κB p65 staining, but strong staining in the cytoplasm. In contrast, A549 cells stimulated with TNF- for 1 h showed strong NF-κB p65 staining in the nucleus
doi:10.6342/NTU201701930
15
(T), and this effect was significantly decreased by pretreatment for 24 h with eupafolin (Fig. 8). Furthermore, TNF--induced ICAM-1 expression by A549 cells was blocked by preincubation of the cells for 1 h with 5-50 M Bay11-7082, a NF-κB inhibitor (Fig.
9A). To further confirm the involvement of NF-B in the TNF-α-induced ICAM-1 expression, we used p65-specific siRNA transfection to knockdown the p65 expression in A549 cells. The expression level of NF-B was markedly downregulated by siRNA transfection (P<0.05, Fig. 9B). Moreover, the level of TNF-α-induced ICAM-1
expression was also attenuated in the NF-B p65-depleted A549 cells (76±3% of
inhibition, P<0.05, Fig. 9C). Furthermore, we use Western blot analysis to determine the expression levels of NF-B p65 in the nuclear portion of A549 cells. The expression level of p65 in the nuclear portion of TNF-α-treated A549 cells was reduced by eupafolin (P<0.05, Fig. 10A). Gel-shift assays were performed to determine the effect of eupafolin on NF-κB activation in TNF--treated A549 cells. As shown in Fig. 10B, low basal levels of NF-κB binding activity were detected in both untreated cells and cells treated only with eupafolin, but binding was increased by treatment with TNF-
for 1 h and further decreased by pretreatment with eupafolin. These results suggest that eupafolin-reduced ICAM-1 expression in TNF--treated A549 cells was mediated by the inhibition of NF-κB activation.
3.5. TNF-α-induced ICAM-1 expression was mediated by AKT/ERK1/2/JNK/NF-B signaling pathway
There are multiple cross-talk points between PI3K and MAPKs pathways, whose co-ordinated action determines the cell fate (Aksamitiene et al., 2012; Bölck et al., 2014). In addition, AKT activation has been shown to activate MAPK pathway (Binion et al., 2009). To further elucidate the detailed pathway, we examine the crosstalk among
doi:10.6342/NTU201701930
16
AKT, MAPK, IB, NF-B p65 in TNF--treated A549 cells. A549 cells pretreated with PD98059, SP600125 or MK2206 for 24h, and then stimulated with TNF- for 15min.
As shown in Figs. 11A-11D, PD98059 had no effect on the phosphorylation of p65 and AKT. In contrast, SP600125 reduced the phosphorylation of p65 (21±8% and 29±10%
of inhibition for 10 M and 50 M, respectively), but not AKT. In addition,
pretreatment with MK2206 significantly attenuated the phosphorylation of ERK1/2, JNK and p65 (70±3%, 48±1%, and 60±6% of inhibition, respectively, P<0.05, Figs.
11E-11G). Moreover, to confirm whether MAPK/AKT was linked to IB
phosphorylation, we performed that A549 cells pretreated with PD98059, SP600125 or MK2206 for 24h, and then stimulated with TNF- for 5min. PD98059 had no effect on the expression of IB phosphorylation. But both SP600125 and MK2206 reduced the IB phosphorylation (59±17% and 69±13% of inhibition, respectively, P<0.05, Figs.
12A-12C). These data suggest that TNF-α-induced ICAM-1 expression in A549 cells was mediated by AKT/IB/ERK1/2, JNK/NF-B signaling pathway.
3.6. Eupafolin suppressed the adhesion of monocytes to TNF-α-stimulated A549 cells
To evaluate the influence of eupafolin on the epithelial cell-leukocyte interaction, we investigated the adhesion of U937 cells to TNF-α-stimulated A549 cells (Fig. 13A).
A549 cells pretreated with 3ng/mL TNF-α for 4 h (T) for 4 h substantially increased monocyte adhesion than control cells (C). Pretreatment of A549 cells with eupafolin for 24 h (E/T) reduced the number of U937 cells adherent to TNF-α-treated A549 cells by 52±6% compared to TNF-α alone (P<0.05, Fig. 13B). As expected, A 549 cells pretreated with 1 or 2 μg/mL anti-ICAM-1 antibody decreased the adhesion of U937 cells to TNF-α-treated A549 cells. This result showed that ICAM-1 plays the important
doi:10.6342/NTU201701930
17
role in the adhesion of U937 cells to TNF-α-treated A549 cells. The adherence of U937 cells to TNF-α-treated A549 cells was also markedly inhibited by the introduction of PD98059, SB203580, or SP600125 (73±5%, 57±8%, and 71±4% of inhibition, respectively, P<0.05). Similarly, the adherence of U937 cells to TNF-α-treated A549 cells was also inhibited by Bay11-7082, MK2206, or LY294002 (69±7%, 67±6 %, and 45±12% of inhibition, respectively, P<0.05).
3.7. Eupafolin reduces ICAM-1 expression in lung tissues in TNF-α-treated mice
To detect the effect of eupafolin on ICAM-1 expression under inflammation in vivo, lung tissues of TNF--treated mice were examined by Western blotting and immunohistochemical staining. TNF- significantly induced the ICAM-1 expression in lung tissues and pretreatment with eupafolin could downregulate the ICAM-1 level by Western blotting (53±15%, P<0.05, Fig. 14A). Fig. 14B shows that no ICAM-1 staining was seen on the lung tissues in the control (C) and eupafolin-treated (E) groups,
whereas strong ICAM-1 staining was seen on the epithelial cells in the TNF-α-treated group (T) by the immunohistochemical staining. The stronger ICAM-1 expression was closely associated with type-II alveolar cells, which were identified by SP-D (Fig. 14C).
In contrast, preadministration of eupafolin showed weak ICAM-1 staining in TNF-α-treated mice (ET).
doi:10.6342/NTU201701930
18
第四章 討論
In the present study, our results showed that eupafolin significantly reduced ICAM-1 expression and monocytic cell line U937 adhesion in TNF-α-stimulated A549 cells in vitro. The influence was partly mediated through inhibition of
AKT/ERK1/2/JNK phosphorylation and NF-κB activation. Furthermore, eupafolin attenuates ICAM-1 expression in lung tissues of TNF-α-treated mice in vivo.
P. nodiflora, an important ingredient of herbal tea, has long been used in traditional medicine to treat inflammatory diseases (Lai et al., 2011). The bioactive compounds isolated from P. nodiflora include flavonoids (Tom´as-Barber´an et al., 1987), essential oils, resin (Elakovich and Stevens., 1985), quinol (Siddiqui et al., 2009), cyclohexenone (Ravikanth et al., 2000), and steroids (Wang and Huang., 2005), which are responsible for its antiseptic, antitussive, antipyretic, antiurolithiatic, antidiabetic, antinociceptive, and anti-inflammatory effects (Balakrishnan et al., 2010; Forestieri et al., 1996). Eupafolin, a flavonoid isolated from P. nodiflora, was chosen for using in the present research, which possesses the anti-inflammatory action (Lai et al., 2011; Maas et al., 2011). Eupafolin promoted iron release from ferritin and donated electrons to the stable free radical DPPH (Dabaghi-Barbosa et al., 2005). Eupafolin protected cultured neurons against glutamate-induced oxidative stress (Kim et al., 2002) and inhibited xanthine oxidase activity (Sanz et al., 1994). The recent study has showed that eupafolin inhibited pro-inflammatory iNOS and COX-2 protein expressions in LPS-stimulated RAW264.7 macrophages (Lai et al., 2011). In addition, eupafolin exhibited anti-tumor effects on MK-1 (human gastric adenocarcinoma), B16-F10 (murine melanoma), HeLa (human cervical adenocarcinoma) cells, and prostate cancer cells (Abe et al., 2002;
Chung et al., 2010; Ko et al., 2014; Liu et al., 2014). Eupafolin lessened virus-induced
doi:10.6342/NTU201701930
19
upregulation of IL-6 and RANTES in RD cells, derived from a human
rhabdomyosarcoma (Wang et al., 2013). Our previous study demonstrated that
eupafolin downregulated melanogenesis (Ko et al., 2014). The present study is the first to report that eupafolin strongly reduces the ICAM-1 expression in TNF-α-treated A549 cells and pulmonary tissues in TNF--treated mice. In addition, eupafolin treatment markedly inhibited leukocyte adhesion to these cells by the inhibiting ICAM-1 expression. The induced expression of adhesion molecules, especially ICAM-1, has been reported to be associated with airway inflammation and the migration and
recruitment of lymphocytes (Lee and Yang, 2013; Qureshi et al., 2003), indicating that an additional mechanism by which eupafolin treatment may be important in preventing the progression of airway inflammation.
MAPK pathways, such as phosphorylation of ERK, JNK, and p38 play the important role in the expressions of proinflammatory mediators, which lead to the initiation and progression of lung inflammation (Lee et al., 2013). The present study demonstrated that TNF-α caused strong activation of three MAPK subtypes in human alveolar epithelial A549 cells, as reported in previous studies (Lee et al., 2013; Jang et al., 2012; Oh and Kwon, 2009). However, the involvement of their activation in the protective mechanism of eupafolin has not been detected. Our results showed that eupafolin decreased TNF-α-induced ERK1/2, JNK and p38 phosphorylation. The increase in ICAM-1 expression induced by TNF-α was markedly suppressed in the presence of an ERK1/2 inhibitor or a JNK inhibitor, but not a p38 inhibitor. ICAM-1 expression was also inhibited by ERK1/2 or JNK-specific siRNA. Based on the results, we suggest one of the signals by which eupafolin attenuates TNF-α-induced ICAM-1 expression involves a reduction in ERK1/2 and JNK activation. Consistent with our results, eupafolin specifically reduced virus-induced upregulation of IL-6 and RANTES
doi:10.6342/NTU201701930
20
by inhibiting the ERK1/2 signaling pathway (Wang et al., 2013). Another study showed that nodiflora extract significantly inhibited the phosphorylation of ERK1/2 and JNK in LPS-treated RAW 264.7 macrophages (Balakrishnan et al., 2010). In contrast, eupafolin significantly induced the phosphorylation of ERK1/2 and p38 MAPK correlate well with the suppression of melanogenesis in B16F10 mouse melanoma cells (Ko et al., 2014). The differences between the above results in terms of the pathways involved may be related to differences in cell type, inducers, and cytokines.
The transcription factor NF-κB was served as the major activator in the regulation of inflammatory responses (Karin et al., 2002). MAPKs have been shown to
phosphorylate NF-κB transcriptional activity. Our results demonstrated that the
activation of NF-κB is necessary for TNF-α-induced ICAM-1 expression in A549 cells and that is in accordance with previous reports (Banerjee et al., 2002; Oh and Kwon, 2009; Wu et al., 2014). Our study further demonstrated that the eupafolin-decreased ICAM-1 expression in TNF--treated A549 cells was mediated through inactivation of NF-κB binding activity. The result is similar with a previous report that nodiflora extract inhibited LPS-induced TNF-, IL-1, and IL-6 production might be related the
reduction of NF-κB activation in RAW 264.7 macrophages (Balakrishnan et al., 2010).
NF-κB is activated by signals possibly involving of the IκB phosphorylation and its dissociation from the inactive cytoplasmic complex, followed by translocation of the active p50/p65 dimer to the nucleus and induced gene expression (Choi et al., 2012). We demonstrated that the eupafolin-induced decrease in ICAM-1 expression was mediated through inhibition of IκB phosphorylation and p65 translocation.
PI3K and AKT pathway have been implicated that they played the crucial role in activation of inflammatory mediators, inflammatory cell recruitment and immune cell function (Koyasu et al., 2003). This notion is confirmed by our observation that TNF-
doi:10.6342/NTU201701930
21
activated the phosphorylation of PI3K and AKT. In the present study, the increase in ICAM-1 expression induced by TNF-α was markedly suppressed in the presence of an AKT inhibitor, but not a PI3K inhibitor. In addition, eupafolin decreased
TNF-α-induced PI3K and AKT phosphorylation. Eupafolin seems to have a more pronounced effect on AKT phosphorylation and subsequently reduced ICAM-1 expression. Thus, one of the mechanisms by which eupafolin reduces TNF-α-induced ICAM-1 expression involves a reduction in AKT activation. Moreover, it has been reported that there are multiple cross-talk points between PI3K and MAPKs pathways, whose co-ordinated action determines the cell fate (Aksamitiene et al., 2012; Bölck et al., 2014). In our study, the phosphylation of AKT was not affected by the inhibition of ERK1/2 and JNK, but the phosphylation of ERK1/2 and JNK was affected by the inhibition of AKT. These findings indicate that AKT is the upstream regulator of IB/ERK/JNK activation. Together these results suggest that eupafolin treatment inactivates TNF-α-induced AKT phosphorylation, which in turn reduces the phosphorylation of IB/ERK1/2/JNK MAPK cascades and NF-B pathways, and subsequently suppressed ICAM-1 expression, resulting in the decreased binding of leukocytes. Because the inflammation is involved in many kinds of chronic and acute lung tissues and it is characterized by the production of proinflammtory cytokines, the enhanced monocyte adhesion, and the accompanying inflmmatory signal (Lee and Yang, 2013), eupafolin may provide a new therapeutic approach for the prevention of
inflammation and lung diseases.
In summary, this study provides the first evidence that eupafolin reduces ICAM-1 expression under inflammatory conditions both in vitro and in vivo and also decreases leukocyte adhesion to alveolar epithelial cells. Our results show that the eupafolin inhibited ICAM-1 expression in A549 cells through blockade of AKT, ERK1/2, JNK,
doi:10.6342/NTU201701930
22
and NF-κB phosphorylation (Fig. 15). Eupafolin, an active component of P. nodiflora, exerts the anti-inflammatory effect on pulmonary epithelial cells in the present study.
Based on these findings, eupafolin should be considered as a novel therapeutic agent targeting epithelial activation in pulmonary inflammation.
doi:10.6342/NTU201701930
23
參考資料
Abe, F., Nagao, T., Okabe, H., 2002. Antiproliferative constituents in plants 9. Aerial parts of Lippia dulcis and Lippia canescens. Biological and Pharmaceutical Bulletin 25, 920-922.
Aksamitiene, E., Kiyatkin, A., Kholodenko, B. N., 2012. Cross-talk between mitogenic Ras/MAPK and survival PI3K/Akt pathways: a fine balance. Biochemical Society Transactions 40, 139-146.
Balakrishnan, G., Janakarajan, L., Balakrishnan, A., 2010. Lakshmi, B. S., Molecular basis of the anti-inflammatory property exhibited by cyclo-pentano phenanthrenol isolated from Lippia nodiflora. Immunological Investigations 39, 713-739.
Banerjee, T., Valacchi, G., Ziboh, V. A., van der Vliet, A., 2002. Inhibition of TNFα-induced cyclooxygenase-2 expression by amentoflavone through suppression of NF-κB activation in A549 cells. Molecular and Cellular Biochemistry 238, 105-110.
Barnes, P.J., 2011. Similarities and differences in inflammatory mechanisms of asthma and COPD. Breathe 7, 229-238.
Binion, D. G., Heidemann, J., Li, M. S., Nelson, V. M., Otterson, M. F., Rafiee, P., 2009. Vascular cell adhesion molecule-1 expression in human intestinal microvascular endothelial cells is regulated by PI 3-kinase/Akt/MAPK/NF-κB:
inhibitory role of curcumin. American Journal of physiology-Gastrointestinal and Liver Physiology 297, G259-G268.
Bölck, B., Ibrahim, M., Steinritz, D., Morguet, C. Dühr, S., Suhr, F., Lu-Hesselmann, J., Bloch, W., 2014. Detection of key enzymes, free radical reaction products and activated signaling molecules as biomarkers of cell damage induced by benzo [a]
doi:10.6342/NTU201701930
24
pyrene in human keratinocytes. Toxicology in Vitro 28, 875-884.
Chen, C. C., Chou, C. Y., Sun, Y. T., Huang, W. C., 2001. Tumor necrosis factor α-induced activation of downstream NF-κB site of the promoter mediates epithelial ICAM-1 expression and monocyte adhesion: Involvement of PKCα, tyrosine kinase, and IKK2, but not MAPKs, pathway. Cellular Signalling 13, 543-553.
Choi, K. W., Um, S. H., Kwak, J. H., Park, H. J. Kim, K. H., Moon, E. Y., Kwon, S. T., Pyo, S., 2012. Suppression of adhesion molecule expression by phenanthrene-containing extract of bulbils of Chinese Yam in vascular smooth muscle cells through inhibition of MAPK, Akt and NF-κB. Food and Chemical Toxicology 50, 2792-2804.
Chung, K. S., Choi, J. H., Back, N. I., Choi, M. S. Kang, E. K., Chung, H. G., Jeong, T.
S., Lee, K. T., 2010. Eupafolin, a flavonoid isolated from Artemisia princeps, induced apoptosis in human cervical adenocarcinoma HeLa cells. Molecular Nutrition & Food Research 54, 1318-1328.
Dabaghi-Barbosa, P., Rocha, A. M., Da Cruz Lima, A.F., De Oliveira, B. H., Martinelli De Oliveira, M. B., Skare Carnieri, E. G., Cadena, S. M., Merlin Rocha, M. E., 2005. Hispidulin: Antioxidant properties and effect on mitochondrial energy metabolism. Free Radical Research 39, 1305-1315.
Demir, T., Yalcinoz, C., Keskinel, I., Demiröz, F., Yildirim, N., 2002. sICAM-1 as a serum marker in the diagnosis and follow-up of treatment of pulmonary tuberculosis. The International Journal of Tuberculosis and Lung Disease 6, 155-159.
Elakovich, S. D., Stevens, K. L., 1985. Volatile constituents of Lippia nodiflora. Journal of Natural Products 48, 504-506.
Forestieri, A. M., Monforte, M. T., Ragusa, S., Trovato, A., Iauk, L., 1996.
doi:10.6342/NTU201701930
25
Antiinflammatory, analgesic and antipyretic activity in rodents of plant extracts used in African medicine. Phytotherapy Research 10, 100–106.
Jang, J. H., Yang, E. S., Min, K. J., Kwon, T. K., 2012. Inhibitory effect of butein on tumor necrosis factor-α-induced expression of cell adhesion molecules in human lung epithelial cells via inhibition of reactive oxygen species generation, NF-κB activation and Akt phosphorylation. International Journal of molecular and cellular medicine 30, 1357-1364.
Karin, M., Cao, Y., Greten, F. R., Li, Z. W., 2002. NF-κB in cancer: from innocent bystander to major culprit. Nature Reviews Cancer 2, 301-310.
Kim, H., Hwang, J. S., Woo, C. H., Kim, E. Y., Kim, T. H., Cho, K. J., Seo, J. M., Lee, S. S., Kim, J. H., 2008. TNF-α-induced up-regulation of intercellular adhesion molecule-1 is regulated by a Rac-ROS-dependent cascade in human airway epithelial cells. Experimental & Molecular Medicine 40, 167-175.
Kim, S. R., Park, M. J., Lee, M. K., Sung, S. H., Park, E. J., Kim, J., Kim, S. Y., Oh, T.
H., Markelonis, G. J., Kim, Y. C., 2002. Flavonoids of Inula Britannica protect cultured cortical cells from necrotic cell death induced by glutamate. Free Radical Biology and Medicine 32, 596-604.
Ko, H. H., Chiang, Y. C., Tsai, M. H., Liang, C. J., Hsu, L. F., Li, S. Y., Wang, M. C., Yen, F. L., Lee, C. W., 2014. Eupafolin, a skin whitening flavonoid isolated from Phyla nodiflora, downregulated melanogenesis: Role of MAPK and Akt pathways.
Journal of Ethnopharmacology 151, 386–393.
Koyasu, S., The role of PI3K in immune cells, 2003. Nature Immunology 4, 313-319.
Lai, Z. R., Ho, Y. L., Huang, S. C., Huang, T. H., Lai, S. C., Tsai, J. C., Wang, C. Y., Huang, G. J., Chang, Y. S., 2011. Antioxidant, anti-inflammatory and antiproliferative activities of Kalanchoe gracilis (L.) DC stem. The American
doi:10.6342/NTU201701930
26
Journal of Chinese Medicine 39, 1275-1290.
Lee, C. W., Lin, C. C., Lee, I. T., Lee, H. C., Yang, C. M., 2011. Activation and induction of cytosolic phospholipase A2 by TNF ‐ α mediated through Nox2, MAPKs, NF‐κB, and p300 in human tracheal smooth muscle cells. Journal of Cellular Physiology 226, 2103-2114.
Lee, I. T., Lin, C. C., Lee, C. Y., Hsieh, P. W., Yang, C. M., 2013. Protective effects of (−)-epigallocatechin-3-gallate against TNF-α-induced lung inflammation via ROS-dependent ICAM-1 inhibition. The Journal of Nutritional Biochemistry 24, 124-136.
Lee, I. T., Yang, C.M., 2013. Inflammatory signalings involved in airway and pulmonary diseases. Mediators of Inflammation ID 791231.
Liang, C. J., Yen, Y. H., Hung, L. Y., Wang, S. H., Pu, C. M., Chien, H. F., Tsai, J. S., Lee, C. W., Yen, F. L., Chen, Y. L., 2013. Thalidomide inhibits fibronectin production in TGF-1-treated normal and keloid fibroblasts via inhibition of the p38/Smad3 pathway. Biochemical Pharmacology 85, 1594-1602.
Liu, K., Park, C., Chen, H., Hwang, J., Thimmegowda, N. R., Bae, E. Y., Lee, K. W., Kim, H. G., Liu, H., Soung, N. K., Peng, C., Jang, J. H., Kim, K. E., Ahn, J. S., Bode, A. M., Dong, Z., Kim, B. Y., Dong, Z., 2014. Eupafolin suppresses prostate cancer by targeting phosphatidylinositol 3 ‐ kinase ‐ mediated Akt signaling.
Molecular Carcinogensis DOI 10.1002/mc.22139.
Maas, M., Deters, A. M., Hensel, A., 2011. Anti-inflammatory activity of Eupatorium perfoliatum L. extracts, eupafolin, and dimeric guaianolide via iNOS inhibitory
activity and modulation of inflammation-related cytokines and chemokines. Journal of Ethnopharmacology 137, 371-381.
Oh, J. H., Kwon, T. K., 2009. Withaferin A inhibits tumor necrosis factor-α-induced
doi:10.6342/NTU201701930
27
expression of cell adhesion molecules by inactivation of Akt and NF-κB in human pulmonary epithelial cells. International Immunopharmacology 9, 614-619.
Qureshi, M. H., Cook-Mills, J., Doherty, D. E., Garvy, B. A., 2003. TNF-α-dependent ICAM-1-and VCAM-1-mediated inflammatory responses are delayed in neonatal mice infected with Pneumocystis carinii. Journal of Immunology 171, 4700-4707.
Rahman, A., Anwar, K. N., True, A. L., Malik, A. B., 1999. Thrombin-induced p65 homodimer binding to downstream NF-κB site of the promoter mediates endothelial ICAM-1 expression and neutrophil adhesion. Journal of Immunology 162, 5466-5476.
Ravikanth, V., Ramesh, P., Diwan, P. V., Venkateswarlu, Y., 2000. Halleridone and Hallerone from Phyla nodiflora as taxonomic markers. Biochemical Systematics and Ecology 28, 905-906.
Rosseau, S., Selhorst, J., Wiechmann, K., Leissner, K., Maus U., Mayer K., Grimminger F., Seeger W., Lohmeyer J., 2000. Monocyte migration through the alveolar epithelial barrier: adhesion molecule mechanisms and impact of chemokines.
Journal of Immunology 164, 427-435.
Sanz, M. J., Ferrandiz, M. L., Cejudo, M., Terencio, M. C., Gil, B., Bustos, G., Ubeda, A., Gunasegaran, R., Alcaraz, M. J., 1994. Influence of a series of natural flavonoids on free radical generating systems and oxidative stress. Xenobiotica 24, 689-699.
Siddiqui, B. S., Ahmed, F., Ali, S. K., Perwaiz, S., Begum, S., 2009. Steroidal constituents from the aerial parts of Lippia nodiflora Linn. Natural Product Research 23, 436-441.
Tomás-Barberán, F. A., Harborne, J. B., Self, R., 1987. Twelve 6-oxygenated flavone sulphates from Lippia nodiflora and L. canescens. Phytochemistry 26, 2281-2284.
doi:10.6342/NTU201701930
28
Wang, C. Y., Chiang, T. H., Chen, C. L., Tseng, P. C., Chien, S. Y., Chuang, Y. J., Yang, T. T., Hsieh, C. Y., Choi, P. C., Lin, C. F., 2014. Autophagy facilitates cytokine-induced ICAM-1 expression. Innate Immunity 20, 200-213.
Wang, C.Y., Huang, S. C., Lai, Z. R., Ho, Y. L. Jou, Y. J., Kung S. H., Zhang, Y., Chang, Y. S., Lin, C. W., 2013. Eupafolin and ethyl acetate fraction of Kalanchoe gracilis stem extract show potent antiviral activities against enterovirus 71 and coxsackievirus A16. Evidence-based Complementary and Alternative Medicine, ID 591354.
Wang, Y. C., Huang, T. L., 2005. Screening of anti‐Helicobacter pylori herbs deriving from Taiwanese folk medicinal plants. FEMS Immunology and Medical Microbiology 43, 295-300.
Wu, C., Wang, H., Xu, J., Huang, J., Chen, X., Liu, G., 2014. Magnolol inhibits tumor necrosis factor-α-induced ICAM-1 expression via suppressing NF-κB And MAPK Signaling pathways in human lung epithelial cells. Inflammation 37, 1957-1967.
Yang, Y. P., Lu, S. Y., Chen, T. T., 1998. Verbenaceae. Flora of Taiwan 127, 421.
doi:10.6342/NTU201701930
29
附圖
Figure 1
Figure 1: Chemical structure of eupafolin.