國立宜蘭大學園藝學系 Department of Horticulture
National Ilan University
碩士論文 Master Thesis
孤挺花花芽發育、種球貯藏及切花適期之研究 Study on flower buds development, bulbs storage and cut
flower stages in Hippeastrum hybridum Hort.
指導教授:朱 玉 博士 Yu Chu Ph. D.
研究生:楊信彥
Hsin Yen Yang
中華民國九十六年七月
July, 2007
內容目次(Contents)
摘要(Abstract)--- 1
Summary --- 2
前言(Introduction)--- 4
前人研究(Literature review)--- 6
一、 植物學分類及生育特性---6
二、 育種歷史及目標---7
三、 球根花卉之休眠及開花生理---8
四、 生長與開花---11
(一)、內在因子---11
1、球根大小---11
2、品種特性---12
(二)、外在因子---13
1、光線---13
(1)、光強度---13
(2)、光週期---14
2、溫度---14
3、水分---15
五、 球根貯藏---16
(一)、貯藏溫度---16
(二)、貯藏時間---17
六、 影響切花瓶插壽命之因子---19
(一)、品種---19
(二)、水分---19
(三)、乙烯---20
(四)、糖---20
(五)、保鮮劑---20
(六)、切花適期---21
(七)、其他---21
材料與方法(Materials and methods)---22
一、 植物材料---22
二、 其他材料---24
三、 實驗環境---24
四、 試驗方法---24
(一)、生育特性調查---24
1、全年生育調查---24
2、不同栽植期對鱗莖內部花芽發育之影響---25
(二)、去除花芽與貯藏溫度及時間對開花之影響---25
(三)、貯藏時間對開花之影響---26
(四)、不同切花適期對切花瓶插之影響---26
五、 試驗設計與統計分析---27
六、 實驗調查項目說明---27
結果(Results) ---29
一、 生育特性調查---29
(一)、全年生育調查---29
1、鱗莖內部花芽之觀察---29
2、孤挺花於不同時期之生長變化---34
3、分生子球之觀察---39
(二)、不同栽植期對鱗莖內部花芽發育之影響---43
二、 去除花芽與貯藏溫度及時間對開花之影響---43
三、 貯藏時間對開花之影響---43
四、 不同切花適期對切花瓶插之影響---54
討論(Discussion)---61
一、 生育特性調查---61
二、 去除花芽與貯藏溫度及時間對開花之影響---63
三、 貯藏時間對開花之影響---64
四、 不同切花適期對切花瓶插之影響---65
參考文獻(Reference)---66
圖目次
圖 1. 本研究所使用之孤挺花品種。---23
圖 2. 孤挺花切花之不同採收標準。---28
圖 3. 孤挺花種球於不同時期解剖情形。---31
圖 4. 孤挺花於不同時期之花芽數變化。---32
圖 5. 孤挺花於 10 月份及 2 月份之花芽長度。---33
圖 6. 孤挺花周徑於不同時期之變化。---35
圖 7. 孤挺花全株鮮重於不同時期之變化。---36
圖 8. 孤挺花地下部重量於不同時期之變化。---37
圖 9. 孤挺花葉長於不同時期之變化。---38
圖 10. 孤挺花葉數於不同時期之變化。---40
圖 11. 孤挺花根長於不同時期之變化。---41
圖 12. 子球於不同時期在鱗莖內發育之狀況。---42
圖 13. 孤挺花鱗莖經不同栽植期之解剖情形。---44
圖 14. 花芽切除與低溫貯放 15、30 及 45 天鱗莖解剖情形。---45
圖 15. 花芽不切除與低溫貯放 15、30 及 45 天鱗莖解剖情形。---46
圖 16. 花芽切除與常溫貯放 15、30 及 45 天鱗莖解剖情形。---47
圖 17. 花芽不切除與常溫貯放 15、30 及 45 天鱗莖解剖情形。---48
圖 18. 孤挺花於 7 ℃貯藏下周徑與鮮重損失百分率之變化。---49
圖 19. 孤挺花於 7℃貯藏下種球腐爛情形。---51
圖 20. 孤挺花‘Apple Blossom’切花於不同切花適期之鮮重變化。---55
圖 21. 孤挺花‘Lady Jane’切花於不同切花適期之鮮重變化。---56
圖 22. 不同切花適期對孤挺花 ‘Apple Blossom’ 花瓣著色之狀況。---59
圖 23. 不同切花適期對孤挺花 ‘Lady Jane’ 花瓣著色之狀況。---60
表目次
表 1. 不同貯藏時間對孤挺花 ‘Blossom Peacock’ 開花之影響。---52 表 2. 不同貯藏時間對孤挺花‘Blossom Peacock’第一花莖小花數及開放天數
之影響。---53 表 3. 不同切花適期對孤挺花切花瓶插壽命之影響。---57 表 4. 不同切花適期對孤挺花切花小花開放率之影響。---58
摘 要(Abstract)
本研究了解孤挺花之花芽於全年發育狀況,並觀察貯藏、栽植時間等處理對 孤挺花開花之影響,最後探討孤挺花之切花適期。試驗中,明顯可看出花芽於7 月份開始變大外推,而於10 月份花芽已超過 3 cm 且數量已增加到 3 個,隔年 2 月花芽已冒出鱗莖頸部。另外,在開花後所分化的第一個花芽,有很高的機率會 在鱗莖內乾枯死亡。種球利用去除花芽及貯放溫度與時間開花之影響,結果顯示 花芽去除及貯放溫度對開花影響不明顯,花芽發育也差異不大。周徑、全株重、
地下部鮮重及葉長都在花期過後開始有增加的趨勢,根長則在花期過後迅速增 長,7 月份後即無明顯變化。觀察子球自然分生方面,子球皆與花芽固定隔兩層 鱗片,且與花芽著生位置相同都在開花軸上。孤挺花利用7 ℃低溫貯藏,結果顯 示雖然低溫可抑制花芽的伸長,但經長期貯藏後開花率低,花莖及小花開放天數 短,但有促進提早開花之作用。另外貯藏至24 週時,種球開始有腐爛的現象發 生。
孤挺花於花莖抽出時,切取苞葉期、著色期及彎曲期三階段進行瓶插試驗。
結果顯示以苞葉期採收之切花,雖瓶插壽命較長,但小花開放率低,花瓣著色不 佳,影響觀賞價值。於彎曲期採收者,雖花朵著色佳,小花開放率高,但瓶插壽 命短。而以著色期採收最為適當,有較長的瓶插壽命且花朵著色亦較佳。
Summary
The purpose of this research lies in understanding development states of flower buds of Hippeastrum hybridum Hort. in whole growing year, and observation effect of storage, plating times on flowering. Finally, we test the cut flower stages. In
experiment, the flower buds began greatening obviously in July. The flower bud has exceeded 3 cm already, and the number has increase to 3 already during October. And the flower buds take out from the bulb neck during February following year. Another, the first buds has very high rate of death and blasting in the bulb after flowering.
Observe the flowering of bulbs use to remove the flower buds and storage temperature.
The result shows it is not evidence flowering by removed the flower bud and storage temperature, and the difference not glaringly at development of flower buds. The bulb sizes, fresh weight of all plant and underground part, and leaf length have trend of increase after flowering. The root length rapidly increasing after flowering, but nearly it isn’t change obviously after July. At observe multiplication of bulblets naturally, all bulblets and flower buds off two layers of scales steady, and with the starting site of flower bud alike, all in the floral axis. Hippeastrum hybridum Hort. storage in 7 ℃ low temperatures, the result shows though low temperature can suppress the extending of flower buds, but after long time storage, the flowering rate is low, the scapes length and longevity of florets are short, but have function of promote the flowering early. In addition, it have phenomena of rotten while storage until 24 weeks.
It is cut spathe stages, color stages and bend stages when scapes of Hippeastrum hybridum Hort. is sprout, begin to experiment of vase. Result show the cut flower of spathe stages harvest, although have a long vase life relatively, but the percentage of floret blooming is low, the petal color not well, and influences ornamental value.
Those who of bend stages harvest, although the flower color well, percentage of floret blooming highest, but the vase life are short. And the color stages is the most suitable, there have longer of vase life and flower colors also better.
前 言(Introduction)
孤挺花(Hippeastrum hybridum Hort.)為原生於熱帶美洲之球根花卉。台灣 的栽培始於1911 年由鈴木三郎氏從新加坡引入(王, 1995),園藝品種於 1960 年自 英國引入第一批,1967 及 1969 年張碁祥氏由日本、荷蘭又引入一批(董, 1985)。
一般在全台各地栽培的為劍瓣種,花中型、花色只有紅色系及白色系,觀賞價值 不高。該種類容易生長、易繁殖且開花容易而遍佈全台,但並未形成商業性消費 (黃, 1980; 劉, 1996)。而目前市面上的孤挺花,花朵大型且有單瓣、重瓣及半重 瓣,顏色豐富,有白色、黃色、紅色、粉色、桔色甚至淡綠色、藍色都有,絕大 部分是藉由進口商從荷蘭,南非或澳洲所搜羅和進口而來(林, 2000)。
孤挺花的育種及生產目前以荷蘭的Ludwing、Warmenhove 公司或南非的 HADECO 公司為主要的新品種培育機構,而又以 HADECO 公司為世界最大的孤 挺花養殖場,而且年年有新品種推出,增加孤挺花市場的多采多姿(林, 2000;
Okubo, 1993)。孤挺花在歐美是頗受歡迎的盆花、花壇以及切花之重要球根花卉,
而在台灣主要以庭園及盆花方式栽培利用(呂, 2000; 劉, 2005)。然隨著新品種陸 續由國外引進栽培,加上孤挺花環境適應力強,已受到各界愛花人士的喜愛,而 孤挺花藍色品種 ‘Worsleya Rayner’,為巴西國寶,台灣貿易商從紐西蘭引進時,
每粒售價甚至高達6 仟元(林, 2000)。然孤挺花目前在台灣除了業餘的零星栽培 外,尚有各農業學校、種苗場及其他農業研究單位的培育、研究及推廣,消費量 有逐年增加的趨勢,可見為一個具有高度發展潛力的熱帶球根花卉(劉, 1996; 張, 1998; 林, 2000)。
雖然孤挺花在台灣尚未形成產業,但相較於荷蘭,台灣地處亞熱帶對培育鱗 莖有更良好的環境條件,但種球卻由荷蘭外銷至台灣,此點值得探討與改進(劉, 1996)。而孤挺花在台灣可周年生長(朱, 2004),若是在溫室的環境中,給予不斷 的供水可保持植株的常綠。但一般孤挺花之開花期都集中於3-5 月,即於清明節
前後。而雖大多數的孤挺花,利用盆花或直接以鱗莖的方式出售,但是仍有相當 潛力當作切花,因其瓶插壽命平均有10 天或更久(王, 1990)。
本研究中,藉由定期之鱗莖剖面觀察花芽,了解孤挺花鱗莖內部花芽於一年 中不同時期之發育狀況,以及概略調查全年之生長變化,以供產期調節或其他研 究用。並利用不同的貯放溫度與時間及去除花芽,以觀察孤挺花於夏天開花之狀 況。另利用長期低溫貯藏來探討花期分散之可行性。再則探討將來孤挺花於生產 切花之產業應用時,提供適當的採收適期。
前人研究(Literature review)
一、 植物學分類及生育特性
孤挺花(Hippeastrum hybridum Hort.)在植物分類上屬於單子葉植物亞綱 (subclass Monocotyledonae) 石蒜科(Amaryllidaceae)孤挺花屬(Hippe- astrum)多 年生草本有皮鱗莖(tunicated bulbs)之球根花卉(劉等, 2004);孤挺花原產於熱帶 美洲,主要分布於中南美洲熱帶地區的巴西、祕魯、墨西哥至智利、阿根廷等地 (Rees, 1985)。而目前種球的主要生產地為荷蘭、以色列、南非及史瓦濟蘭(王, 1995;
朱, 2004; 劉, 2005)。
孤挺花一般常用的英文名稱為Amaryllis,但又因原生種的不同,又有 Barbados Lily (H. equestre; H. puniceum)、Butterfly Amaryllis (H. papilio)、Dutch Amaryllis、Equestrian Star-Flower (H. eqestre; H. puniceum)、Green Amaryllis (H.
calyptratum)、Knight’s Star Lily,Lily of the Palace (H. aulicum),Mexican Lily (H.
reginae),South African Amaryllis 等(Okubo, 1993);別名為大孤挺花、百枝蓮、石 蒜花、喇叭花和華冑蘭及鼓吹花(王, 1995; 劉, 2005);而 Hippeastrum 是由希臘語 騎士(hippeus)和星星(astron)兩個字組合而成(Huely et al., 1992)。
孤挺花球根為葉片基部肥大之鱗片(leaf scales)組成,最外層包覆之咖啡色皮 膜由養分耗盡之鱗片所形成,具有保護及防止水分散失之功用,屬於有皮鱗莖 類。葉為帶形或線形,花莖中空,花序為繖形花序(umbel),由 2~6 朵花所形成,
每一朵花是由六枚花被(perianth)所組成,花萼與花瓣不易區分(劉等, 2004)。雄蕊 6 枚,花柱較雄蕊長,柱頭三裂,三心皮,子房下位,果實為蒴果(劉, 2005)。花 朵大而豪華,但較缺乏香氣,花色從深紅、粉紅、白色、桔色、黃色、藍色都有 (王, 1990),甚至目前已有黑紅絨的花色以及印度也傳出已育出帶銀色的品種,且 也已有帶香味的品種問世。孤挺花每朵花是左右對稱,花藥附在不等長的花絲
上,花絲彎下伸展。種子黑色,圓而扁平,質薄,種子壽命短(黃, 1980)。種球繁 殖可用種子(實生法)、自然分球法以及種球切割法,而種球切割法又因不同切割 或培養方式基本上可分為基底切割法、瓣狀鱗片法、雙鱗片法及微體繁殖法(張, 1998; 劉, 2005),而目前在商業上則以雙鱗片繁殖法為主。
孤挺花因原產於熱帶之中南美洲,因此較不耐寒,在台灣像屏東可露地栽 培,但夏季陽光太強,溫度也太高。孤挺花幼苗和鱗莖生長,最低溫可忍受到8℃,
低於8℃則停止生長,不過最合適生長溫度是日溫 23℃,夜溫 18℃(王, 1995);
日溫28℃,夜溫 23℃則有利於葉片生長以及促進種球的發育,但會抑制花芽形 成(劉, 2005)。孤挺花栽培介質以砂質土壤,排水良好,最適 pH 值在 5.5-7 之間,
稍含有機質較佳(王, 1995; 江, 1998; 劉, 2005; 羅, 2005)。病蟲害方面有炭疽病(楊, 1996)、毒素病(黃, 1980)、赤斑病、蛞蝓、蝸牛(董, 1985)、石蒜夜蛾及根蜘蛛(黃, 1980)等。
二、 育種歷史及目標
孤挺花之育種歷史已超過 200 年,於早期育出之雜交種,主要都集中於 H.
aulicum、H. psittacinum、H. puniceum、H. reginae、H. reticulatum、H. striatum 和 H. vittatum。而最早育出的雜交種為 Johnsonii 氏在 1799 年由 H. vittatum 和 H.
reginae 於英國育成。而於 19 世紀初期,在南美洲又發現許多新種,並且引進歐 洲。然孤挺花育種最重要的兩群即是Reginae 和 Leopoldii 之雜交群。Reginae 群 是由荷蘭Jan de Graff 父子利用 H. vittatum、H. striatum,和 H. psittacinum 雜交選 育出適合於歐洲之優良品種。而由Richard Pearce 發現的 H. leopoldii 和 H.
pardinum,從南美洲安地斯山脈引進英國,具有花大、開展及對稱之生育特性。
之後Veitch 和 Sons 持續進行育種雜交,發展出 Leopoldii 雜交群。除這兩群外,
Vittatum 群是由 H. vittatum 與其他種(species)雜交育種而成。Reticulatum 雜交群 是在夏末至秋季開花,主要是巴西種 H. reticulatum,比其他雜交群矮小、常綠,
葉脈中肋常有黃色或白色條帶(劉, 2005)。而目前普遍栽培的品種大都由 H.
aulicum,H. reginae,H. reticulatum,H. solandriflorum,及 H. vittatum 等經過多 年雜交而成,所以用hybridum 的種名(黃, 1980; Okubo, 1993)。
而孤挺花的原生種約有60 至 70 種,現今之栽培種乃經由原種選種後在經多 年雜交育種選育而來。花色從白色、粉色、桔色、紅色、淡綠色等,花瓣有單瓣、
半重瓣、重瓣。目前雜交栽培種已從二倍體發展到三倍體和四倍體。因此花朵特 性也傾向於大花、花瓣寬大、花形圓、平面化等。而目前未來孤挺花主要育種目 標為(1)耐寒品種(2)具香味品種(3)新花色(尤其是金黃色、藍色到紫色系列)(4)抗病 品種等(劉等, 2004)。
三、 球根花卉之休眠及開花生理
學者們常利用不同的觀點及方式將球根花卉做歸納及分類,以利研究者或栽 培者了解與應用及研究。而球根花卉中為了解其開花生理,常依據其不同的開花 時期、休眠時間做為分類之準則。下面即提出幾種以休眠、花芽分化及形成的時 期或開花季節的不同而做的分類,供作參考。
球根花卉若以葉片於一年四季中的生長、老化情形及休眠時期,可將其概分 為三類:
(一) 常綠性球根花卉:一般原生於熱帶地區者多為此類,如文珠蘭、孤挺花等,
但如遇到不適宜的環境時亦會落葉,如乾旱或於溫帶地區種植時遇低溫才會造 成落葉。
(二) 落葉性而於夏季遇高溫時休眠:在溫帶地區,此類球根在春季葉片伸出土面 而後開花,夏季溫度高土壤乾燥時,葉片便枯萎老化,此時採收球根。如鬱金 香及風信子於夏天採收時及貯藏期間,鱗莖內的生長點繼續分化葉片,再分化 花芽,等秋季溫度較低時,再種回田裡,經冬季低溫之後於隔年春天再重新抽
梢開花。
(三) 落葉性而於冬季時休眠:此類於溫帶為夏季開花,且具有較高的生長速率,
但於冬季低溫葉片即停止生長或進入休眠,等滿足其低溫需求才抽梢並於隔年 夏天開花,如唐菖蒲與百合等(李和林, 2001)。
而若依其花芽創始及分化(flower initiation and differentiation)始於休眠時的前 後,可將球根花卉概分為三類:
(一) 花芽分化於種球進入休眠前即開始,如鬱金香等,其花芽分化與抽花養分主 要由母球供應。
(二) 於種球休眠後之莖梢抽長時花芽分化,如鐵炮百合、唐菖蒲等,而其花芽分 化除了受到溫度影響之外,光度若不足常會有花苞夭折或消蕾的現象。
(三) 在合適的氣候下,種球並無真正的休眠期,故此類型在種球夠大後,每形成 一定的葉片數便能花芽分化形成花芽,如孤挺花、亞馬遜百合等,其葉及花 梗的抽出受溫度及水分的影響較大(Rees, 1985)。
而若依照花芽分化及形成的時期,可將其分為七種類型:
(一)花芽在春或夏季初開花之後,或在當年種球採收時形成,如:西洋水仙、台 灣一葉蘭等。
(二)花芽是在葉片枯黃萎凋,種球採收及貯藏之後形成,如:鬱金香、風信子等。
(三)植株在定植後,經過冬季或初春的低溫才形成花芽,如:球根鳶尾、鐵炮百 合等。
(四)花芽於種球貯藏時開始分化,但於田間定植時才完成,如:球根海棠。
(五)花芽在春天定植抽梢後的涼溫(約 20-23℃)下形成,如:唐菖蒲。
(六)植株在開花前的一年多之前便開始形成花芽,如:納麗石蒜。
(七)植株每形成一定的葉片數之後便能形成花芽,如:孤挺花每形成 4 片葉子便
可進入花芽分化(李和林, 2001),風雨蘭則是每形成 3 片葉子就可分化 1 個花 序(Kerling, 1941; Genjiro and Hideo, 1997)。
然在球根花卉的研究中,許多石蒜科球根花卉被用來研究了解其成長週期性 及調查溫度於花芽創始和發育上的作用。獲得的結果總結如下:
(一)花芽在季節的確切時刻形成,依季節分為:
1.花芽在春天和初夏期間的鱗莖採收前形成,而再依其開花時節的不同而分 為:
(1)在夏末到秋天開花:紅花石蒜(Lycoris radiate),鹿蔥(Lycoris squamigera)、
黃花冬水仙(Sternbergia lutea)和秋雪片蓮(Leucojum autumnale)。在這些種 類,低溫為開花誘導所必須以及花芽創始和發育到開花的最適溫度為20 到30℃之間的範圍。
(2)在一年之後的春天開花:夏雪片蓮(Leucojum aestivum)。在這個種類,花 芽創始與花器進一步分化的最適溫度為20-25℃且開花需求為暴露在 10-15℃範圍間的低溫。
2.花芽在鱗莖採收後的夏天形成,而開花時期在冬天:中國水仙(Narcissus tazetta var. chinensis),25℃為花芽在心皮形成階段和開花之間發育的最適溫 度。
3.花芽在夏天的生長期間形成,而開花時期在夏末到秋天:晚香玉(Polianthes tuberosa)(後被歸類至龍舌蘭科),開花誘導必須暴露在高溫 30℃下 2 個星期。
4.花芽在秋天的生長期間形成,而在一年之後的夏天開花:蜘蛛百合
(Hymenocallis speciosa)。在這個種類,在花芽創始後先暴露在高溫 20℃之上 和長的日長下,而花芽形成和開花需求的溫度範圍為從15 到 20℃。
(二)花芽選擇性的在整個同化作用期間葉片形成時形成,而再依其開花時節的不 同而分為:
1.在 1 月到 5 月開花:垂筒花(Cyrtanthus mackenii),每年形成 2-3 個花序,而 開花需求為在心皮形成階段需暴露在10℃下 3 星期或 15℃下 4 星期。
2.在 3 月開花:紅花君子蘭(Clivia miniata),每年形成 1 個花序,而開花需求 為在心皮形成階段以後暴露在10℃的低溫下 8-9 個星期。
3.在初夏開花:香粉文珠蘭(Crinum x powellii)和櫻茅(Rhodohypoxis baurii),每 年分別形成5-6 及 24-26 花序。而香粉文珠蘭的開花必須在心皮形成階段暴 露在15/12℃下 30 天或在 6℃下 45 天。櫻茅的開花需求為暴露在 6℃下 12 個星期或10℃下 16 個星期。
4.在夏天開花:白花蔥蘭(Zephyranthes candida),每年形成 3-4 花序。開花則 在25-30℃的高溫下(今西, 1999)。
四、 生長與開花
影響植物體生長發育及開花的因子,相當複雜且多,基本上可分為植物體本 身與外在環境的影響因子,即內在因子和外在因子。
(一)、內在因子:
在球根花卉中,其內在因子有球根的大小或重量、品種、葉片或根長等因子。
1、球根大小
一般而言,種球大小是決定該種球根花卉是否具開花能力的指標之ㄧ(呂, 2000; 李和林, 2001)。然而種球需要多大才能開花,亦因種類或品種的不同而有 所不同,尤其鱗莖類種球大多需培養2-3 年或以上才能開花(賴, 1995)。
在台灣一葉蘭(Pleion formosana)中,球莖內澱粉含量隨採收成熟度愈高而增 加,此外,晚期採收,花朵較大,花梗長度及花朵壽命也較早期採收者為長,其
次,花芽大小及乾物重亦隨採收成熟度而增加。若以落葉為分界點,11 月中旬落 葉後採收之球莖於23℃貯藏一個月後即陸續萌芽,至達 50%萌芽天數較貯藏於 5
℃者為少。而落葉前採收者在23℃貯藏 8 週期間無萌芽現象,種植後之萌芽速率 較於5℃貯藏 8 週者慢,而 5℃冷藏雖可促進萌芽開花,然開花仍不整齊,與 23
℃貯藏者同(滕等人, 1985)。
於紫燈花 ‘Queen Fabiola’ 中,其開花率在母球重量約 0.6 公克以下,即迅速 下降,而在母球重約0.25 公克時,開花率僅達約 50%,且在母球重量≦ 0.1 公 克時,則不開花,但母球大小對花莖長度並無顯著性的影響,而在≦ 0.6 公克的 球莖,每一花序上的小花數明顯地降低,而隨著母球大小的增加,其子球的數量 及重量上也隨之顯著性的增加(Han et al., 1991)。石蒜科地哥飛花屬植物 Zephyra elegans D. Don 中,較大的球根具有較高的萌發率,而開花率及到達開花日數上 則沒有達顯著性差異,但是在開花品質上,除了花莖長度沒有差異外,其重量、
分支數及小花數則隨球根大小的增加而增加(Yañez et al., 2005)。
而一個孤挺花的成熟鱗莖周徑大約在20 公分以上,是由葉的基部肥大的葉 鱗片所組成,大約含有6 個枝葉組(Shoot units),每個枝葉組一般含有 4 片葉片 (3 片全周葉及 1 片半周葉)和一個花序,其中 2 個枝葉組沒有地上部,2 組有萌發 的葉片,而2 組未萌發。一般是由後 4 組萌發開花,花序生長之位置為葉片萌發 時的側面,並相對形成一軸線,在此稱為開花軸。其花芽分化在一年之中會發生 數次,但是有些花芽不會發育達到開花階段,而在鱗莖內枯死(Rees, 1985)。
2、品種特性
由於各品種常由遺傳育種學家利用不同種質來源進行雜交選育,故品種間的 生育特性不盡相同。於納麗石蒜七個試驗品種中,六個品種屬先開花再長葉型,
而另一品種則為常綠型(賴, 1995)。
美國於11 月從荷蘭進口孤挺花鱗莖運至北卡州,接著貯藏於 13℃下直到種 植於不同商業介質中,栽培於22-24 ℃之自然光照中,當第一花莖達到 30 cm 高 的銷售期時發現,不管在任何商業介質中,‘Apple Blossom’都較‘Red Lion’早開花 (De Hertogh and Tilley, 1991)。孤挺花鱗莖於花朵凋謝後經 5 個月的養球才可再開 第二次花,而品種之間的開花率為: ‘Picotee’ > ‘Red Lion’ > ‘Wonderland’ (劉, 1996)。
(二)外在因子:
植物體從種子發芽生長開始,無一階段不受到環境的控制與影響,而構成植 物生長環境的因子有光線、溫度、水分、土壤及大氣等因子,這些因子以單獨或 共同影響著植物的生長與開花。
1、光線
光線為植物生長中重要的環境因子之一,直接影響著植物的生長與開花,而 一般光線影響植物生長的情形,有光強度(light intensity)、光照時間(light duration) 及光質(light quality)。
(1)、光強度
光強度直接影響著植物體的光合作用速率,光強度愈強,其光合作用固 定CO2的速率愈快,但超過某一程度,即不再增加,稱之光飽和點(light saturation point)。
亞洲型百合‘Enchantment’中,以連續的補充光照(至少 20 瓦/平方公尺) 和1000 ppm CO2的組合來做處理,可使之減少落蕾,縮短生長至開花的時 間,改進植株及花朵的開花品質(Hendricks, 1996)。而適度遮蔭可促進節間伸 長,但過度遮蔭卻會造成碳水化合物下降而產生消苞及落蕾(許, 1995)。鐵炮
百合自花芽分化後遮蔭85% (1.4 mol‧m-2‧d-1 photons)會使得花苞產生消蕾 現象(Miller and Langhans, 1989a; 1989b)。
(2)、光週期
於紫燈花 ‘Queen Fabiola’ 中,長的光週期雖能夠加速開花但卻會降低 其開花品質以及開花率(Han et al., 1991)。鐵砲百合‘Ace’於低溫下(4.4℃) 貯 藏0~6 週之後,移入最低 21℃的溫室中,以不同日長條件栽培,結果顯示低 溫可由長日(8.5 小時之自然光加 4 小時之暗期中斷) 替代而提早使開花率達 到100%(李, 1992)。
在東方型百合 ‘Bonfire’、‘Casa Blanca’、‘Furore’、‘Joume’s End’、
‘Laura’、‘Pink Virtuoso’及‘Star Gazer’七個品種中,從一月底開始每日以日光 燈補充照明以延長光週期,大約六星期,明顯會較快開花(Beattie and White, 1993)。而孤挺花經由 4 年的實驗顯示,日長對生長及花芽發育並沒有影響,
但在短日下,其生育有較為衰弱的現象,可能是因光合成產物減少的緣故 (Hayashi, 1974)。
2、溫度
球根花卉的種類非常多,分布地亦廣,故對其生長開花所需求的溫度也有很 大的變化。溫度會直接影響球根花卉的生長與開花,也影響發育,花色等品質。
於爆竹百合(Lachenalia aliides) ‘Pearsonii’品種中,到花日數隨著促成栽培的 溫度提高而增加。而開花率隨著促成栽培溫度的提高而下降,即促成栽培的溫度 較高,花序的夭折率則會增加。鱗莖在前四週都是16 ℃的促成溫度處理中,相 較於前四週24 ℃的處理,明顯有較高的開花率且達開花日數也較短(Roh, 2005)。而於紫燈花 ‘Queen Fabiola’中,隨著夜溫的增加花莖長度有明顯的縮短
(Han et al., 1991)。東方百合在夏季高的溫室促成溫度下,會加速開花但畸形花朵 的數量亦同樣增加,而在15.5 ℃的促成溫度下,花莖的長度較長於 25 ℃的促成 溫度(Lee and Roh, 2001)。
石蒜科地哥飛花屬植物 Zephyra elegans D.Don 生長於日夜溫 15/10 ℃或 20/15 ℃下,萌芽及開花較於 25/20 ℃下為迅速,且在日夜溫 25/20 ℃下生長者開 花品質也較為低劣(Yañez et al., 2005)。菊花於高溫下,會增加每株植株的花朵數 量,但卻會縮減每朵花朵的大小(Carvalho et al., 2005)。而於孤挺花方面,Hayashi 和Suzuki (1970)利用日夜溫 28/23 ℃,23/18 ℃,18/13 ℃,13/8 ℃處理,結果顯 現種苗和鱗莖生長的最低溫約為8 ℃,而最佳的生長溫度為日夜溫 23/18 ℃,而 在28/23 ℃的處理中,能夠快速促進葉片生長,但鱗莖發育和花芽創始卻會受到 抑制。
3、水分
水分管理為植物栽培生長中一個相當重要的環境因子。而一般植物缺水時常 造成光合作用減少,碳水化合物比例及醣類代謝途徑的改變,組織內RNA 及蛋 白質含量減少,型態改變(如葉片脫落、葉面積減少等),產量、品質的減少及下 降(朱, 1990)。在薑荷花中,於種植後若缺乏灌溉會導致萌芽延遲,側芽數減少,
而如生長期缺水則生長勢容易衰弱,影響其花莖及種球的生長與發育,最後使品 質及產量降低與減少,尤其是對薑荷花切花之瓶插壽命的縮短最為顯著(張, 2007)。
於1984 年 1 月 8 日利用鱗莖周徑 19-32 公分的孤挺花 ‘Red Lion’ 種植於 12.5 公分的塑膠盆中,之後使植株生長在日溫18℃,夜溫 15.5℃下的溫室中,鱗莖於 種植後4-8 週開花,標記開花與未開花者,僅只使用有開花的鱗莖做隨後的灌溉 處理。而從5 月初到 9 月中旬使用遮陰減少光量子流(photosynthetic photo flux
density, PPF) 50%,再從 9 月初進行斷水處理,處理組別分別為 a)不斷水連續灌 溉(對照組);b)斷水 8 週;c)斷水 4 週;d)斷水 2 週。結果顯示,斷水處理 8 週的 第一花莖在40 天內有 61%開花,而在 60 天內即達 100%,第二花莖則達 28%,
而斷水處理4 週的第一花莖則在 80 天內有 28%開花,140 天內達 100%開花,而 斷水2 週者則在 160 天內完全開花,但連續灌水者卻僅只有 83%開花。斷水處理 有效地促進孤挺花 ‘Red Lion’ 提早開花,而在第一及第二花莖上的花朵數量上並 不會造成影響(Boyle and Stimart, 1987)。
除了內在及外在因子外,人為的栽培操作管理亦會影響球根植物的生長發育 及開花,如種植深度與距離。
球根的種植深度一般為球根本身高度的2~3 倍。但一般小型球根類才適用於 此通則,對較大的球根則不適用,深度需較為淺,使球根頂端或頸部與土面相平 或稍高,有些球根甚至需露出一半於土面之上(董, 1985)。於紫燈花 ‘Queen Fabiola’ 品種中,開花比率隨著種植深度的增加而減少(Han et al., 1991)。而孤挺 花的種植深度則以頸部於表土之上或鱗莖露出介質面1/2~1/3 為宜(黃, 1980; 董, 1985; 王, 1995)。孤挺花的種植密度如果太密會抑制其花芽創始(Hayashi, 1972),
栽培上以球距20 公分為適當(黃, 1980)。
五、 球根貯藏
球根貯藏是一般球根花卉用來調節分散花期、促進花芽分化及發育和打破休 眠等常用之手段,而貯藏又常以貯藏溫度及時間來作為調控因素。而貯藏能力與 種類、品種特性、貯藏溫度及採收技術有關。
(一)、貯藏溫度
在爆竹百合 ‘Pearsonii’ 中,當鱗莖貯藏在 10℃,12.5℃或 15℃45 天時,與
在20℃及 25℃時相較下,開花增加,但葉片長度和小花數則減少。而隨著鱗莖 貯藏溫度的提高,開花日數上也隨著增加。而鱗莖貯藏在10℃,12.5℃,15℃下 15 天,而在 17/15℃的促成栽培溫度下,顯示達開花日數小於 110 天,而且小花 數都在17 朵以上。然葉片長度亦隨著鱗莖貯藏溫度的提高而增長。而貯藏在較 高溫度(20℃,25℃)下的鱗莖,雖有比較高的開花率,但是相對地達開花日數也 隨著拉長(Roh, 2005)。而在另一品種 ‘Ronina’ 中,將鱗莖貯放於高溫,中溫及低 溫下,而高溫處理者,鱗莖發育較多的單位,但也發生花序的消蕾現象,然而在 低溫的處理中,雖然發育較為緩慢,但大體上其鱗莖生長品質較中溫及高溫為優 (Toit et al., 2002)。
台灣一葉蘭以5℃冷藏者之開花率為最高,達 83.5%,而於 10℃及 23℃貯藏 者只 70%左右(滕等人, 1985)。而納麗石蒜乾藏於 2℃,相對溼度於 80%下的環 境約2 個月,可以打破鱗莖休眠且開花率較高,但在 15℃下未貯藏的鱗莖也會打 破休眠而生長發育,然與貯藏在5℃或 9℃者相比較下,則需要較長的時間(賴, 1995 )。
在溫帶地區的孤挺花開花後,地上部枯萎或因寒冷氣候而將鳞莖挖起貯藏,
而貯藏溫度對孤挺花的開花亦會造成影響;在早期Luyten (1926)於 9 月份鳞莖採 收之後利用27、23 或 17℃貯藏 4.5 週,之後再將所有鱗莖於 23℃下貯放 4 週後 種植於20℃的土壤及 17-24℃的環境下。貯藏在 27℃下沒有開花的現象,而以 17℃
貯藏者具有較高的開花率,而貯藏於23℃者開花非常的不規律且即使第一花序已 經伸長及花朵開放,第二花序也是很少出現。低溫能夠預防於貯藏期間過度的萌 芽,而且能促進於較高溫生長的一致性(Okubo, 1993)。
(二)、貯藏時間
於爆竹百合 ‘Pearsonii’ 中,鱗莖貯藏在 10℃下 15,30,45 天,而在 17/15
℃的促成栽培溫度下,可看到貯藏15 天的鱗莖葉片較長,而隨著貯藏時間的增 加,植株葉片也大幅縮短,在其他貯藏溫度(12.5℃,15℃,20℃,25℃)下,亦 是如此。且貯藏在10℃下 15,30,45 天,可發現不管促成栽培的溫度在 17/15
℃或21/19℃下,達開花日數隨著貯藏時間的增加而縮短。但貯藏於 15℃,20℃,
25℃下 15,30,45 天,達開花日數並無隨著貯藏時間的增加而縮短,甚至有增 加的趨勢(Roh, 2005)。
台灣一葉蘭在 23℃室溫貯藏 4 週較 8 週知開花率略高,因球莖於室溫乾貯(dry storage)8 週,不少球莖已萌芽長根,已萌發之芽體得不到充分的水分供應而致消 蕾。將台灣一葉蘭球莖貯於1~2℃,20 週後因花芽伸長,故降低貯溫至 0~-2℃
間以抑制其伸長,到23 週,不論中或大球,開花率仍在 90%以上,花朵大小,
花梗長及花朵壽命未見減少,23 週後苞片出現失水焦褐現象,花徑變小,花梗變 短,每花壽命亦減少,然至31 週時仍有 90%之開花率與觀賞價值,31 週以後因 花瓣不易張開,花瓣顯著變小,花苞顯色後之消蕾率過高而使開花率驟降,至49 週時只有30~50%(滕等人, 1985)。
宮燈百合(Sandersonia aurantiaca)塊莖的萌芽率隨貯藏時間的增加而遞減(60 天94.3%,90 天 93.8%,120 天 88.5%,150 天 86.5%),而萌芽天數在貯藏 30 天時為58.8 天,而貯藏 150 天者則縮短為 17.5 天,可見萌芽天數隨貯藏時間的 增加而減少,而種植到採收的天數亦然(Clark, 1995)。而番紅花 Crocus sativus L.
的花朵數及花朵大小隨著冷藏的時間增加而漸減(Molina et al., 2005)。
然而在孤挺花的貯藏上,利用涼溫15℃及高溫 30℃不同天數做變溫處理,
利用15℃涼溫促使花芽萌發,而 30℃用來促進花莖伸長,總處理天數為 60 天,
有6 種不同的組合處理:(1)15℃10 天,30℃50 天、(2)15℃20 天,30℃40 天、(3)15
℃30 天,30℃30 天、(4)15℃40 天,30℃20 天、(5)30℃60 天、(6)自然環境。處 理組於74-91 天開花,然對照組則是在 160 天開花,而以乾燥方式貯藏,則發現
溫度處理者在127-132 天開花,但對照組則在 181 天開花且開花時缺少葉片,但 卻可節省空間並適合於生產切花(劉, 1996)。
另Bose 氏等人(1979)將孤挺花 ‘Fire Dance’ 品種之鳞莖放置於 30、17、10、
5℃的環境下 30-90 天,以 30℃貯藏者有促進植株高度的作用,株高為 47.6 公分,
而在 5℃下貯藏者之株高則為 39.8 公分,且於較高溫度貯藏下,葉片數、花莖數 及花朵數均較低溫貯藏為高;而隨著貯藏時間的增加則開花延遲且降低開花品質 (Bose et al., 1979)。
六、 影響切花瓶插壽命之因子
花朵的壽命依花卉種類的不同,有相當大的變化。而大部分的種類,花朵在 植株上的壽命都較長,一旦切離母株插在水中,都會縮短花朵的壽命,這種切離 植株後的花朵壽命,即稱為切花的瓶插壽命(vase life)。而影響瓶插壽命的內在及 外在因子相當多,其中以切花種類、品種、養水分、栽培環境、水中微生物、花 朵老化等問題最為重要(黃, 2002)。
(一)、品種
火鶴花於四個品種間的瓶插壽命比較上,以 ‘Pistache’ 的瓶插壽命為最長,
‘Sultana’ 次之, ‘Tropical’ 及 ‘Fantasia’ 最短(葉, 2004)。
(二)、水分
水溶液中有很多因子影響著切花的瓶插壽命及品質,如pH 值、水中含氧量、
蔗糖濃度等,如在玫瑰切花中,這些因子都對其造成吸水上的影響(Conrado et al., 1980);切花的吸水能力除了受到環境因子及其他因子影響外,切花本身因子也會 影響到切花的吸水能力,如器官、組織結構及型態上的差異。玫瑰 ‘Sonia’ 於採
收後暴露於空氣中3 小時,其花梗的吸水能力不會受到影響,但如超過 24 小時,
就會受到嚴重阻礙(van Doorn, 1994)。
(三)、乙烯
在許多研究報告中皆顯示乙烯不僅會加速花朵老化且也會縮短切花的瓶插 壽命,於福祿考切花中,將切花暴露於1 μL/L 濃度的乙烯下 12 小時,達 50 %落 花率,且開花數減少及較短的瓶插壽命(Poart et al., 1995)。而鳳尾耆、雞冠花、
糙葉向日葵、吊鐘柳和錦帶花切花暴露在0.2 或 1.0 μ/L 的乙烯下,亦明顯的縮短 瓶插壽命(Redman et al., 2002)。而鬱金香切花於採收後花朵的呼吸率及乙烯量皆 低,其花瓣老化主要是與脂氧化酶活性的快速上升有關(Jones and McConchie, 1995)。
(四)、糖
蔗糖的添加在嘉蘭上明顯增加了瓶插壽命,且能增進未成熟花芽的開放及著 色(Jones and Truett, 1992),亦能輕微增加孤挺花 ‘Zanzibar’ 的瓶插壽命,且能促 進所有小花的完全開放,也能改善孤挺花切花花梗基部裂開的反捲問題(Halevy and Kofranek, 1984)。而在雞冠花和糙葉向日葵上,蔗糖的添加反而會減少瓶插壽 命(Redman et al., 2002)。
(五)、保鮮劑
嘉蘭切花使用殺菌劑8-HQC (250 mg/L)、DICA (50 mg/L)及 Physan-20 (50 mg/L)明顯可以延長瓶插壽命(Jones and Truett, 1992)。玫瑰使用 200 mg/L 的 8-HQS 及30 g/L 加 200 mg/L 8-HQS 處理切花,結果顯示不管單獨使用 8-HQS 或是與蔗 糖組合使用都可延長瓶插壽命,尤其以蔗糖加8-HQS 處理最為有效,不僅可延長 瓶插壽命更可增加花徑大小及鮮重,且8-HQS 和蔗糖一起使用與單獨使用 8-HQS
及對照組比較,明顯可增加花瓣中葡萄糖、果糖及蔗糖的含量(Ichimura et al., 1999)。洋桔梗切花使用 0.2 mM 硫代硫酸銀(silver thiosulfate complex; STS)與 4 % 蔗糖合用,與單獨使用0.2 mM 硫代硫酸銀比較,更能延長瓶插壽命,且可促進 高位芽開放及增進花瓣的著色(Shimizu and Ichimura, 2005)。HICP
(2-hydroxy-3-ionene chloride polymer)在玫瑰中明顯的可以延長其瓶插壽命
(Ueyama and Ichimura, 1998)。使用 500 mg/L 的 8-HQC 會增加糙葉向日葵和錦帶 花的瓶插壽命但卻會減少鳳尾耆的瓶插壽命,而鳳尾耆和雞冠花使用STS 可增加 瓶插壽命(Redman et al., 2002)。
(六)、切花適期
紫羅蘭切花依照不同的販賣市場而有不同的採收標準,如是以國內市場為目 的,應在花穗上小花達1/2(花開 10 朵)開放時採收,但如以日本市場為銷售地時,
因需經過一段貯藏時期,故以花穗小花達1/3(花開 7 朵)開放時採收,如此才能有 最佳的瓶插壽命(張等, 2003)。
(七)、其他
非洲菊四個品種在切花採收前噴施及採收後浸泡或注射0,0.5,1.0,1.5 % 的氯化鈣,結果顯示 ‘Campitano’ 、 ‘Dino’ 及 ‘Testarossa’ 三品種於注射 1.0%
的氯化鈣後與對照組比較,明顯可以延長瓶插壽命及降低垂頸率。而 ‘Sangria’ 於 採前噴施1.0-1.5 %的氯化鈣會有較長的花朵壽命及較低的垂頸率。鈣處理與對照 組相比較約可增加3-4 天的瓶插壽命及延遲垂頸 3-5 天,然而高濃度(1.5 %)的氯 化鈣雖能增加花莖鈣的含量,但對瓶插壽命並沒有額外的貢獻(Gerasopoulos and Chebli, 1999)。在玫瑰的數個栽培品種中,隨著空氣濕度的增加,而瓶插壽命則 雖之減少(Mortensen and Fjeld, 1998; Mortensen and Gislerød, 1999)。
材料與方法(Materials and methods)
一、 植物材料
試驗一中第一項全年生育調查試驗所使用之種球為取自本系所系館二樓 陽台,其鱗莖周徑平均為25 公分以上,品種為 ‘Apple Blossom’ (圖 1A)。第 二項不同栽植期對鱗莖花芽之影響,則為95 年 10 月份購自芊卉種苗公司,
經貯藏後,依不同日期取出種植,在96 年 6 月份切球觀察其內部花芽狀態,
品種為 ‘Blossom Peacock’ (圖 1B)。而試驗二花芽去除與貯藏溫度及時間試驗 使用之種球為95 年 1 月份自芊卉種苗公司購得,種球周徑為 25 公分以上,
品種亦為‘Apple Blossom’。試驗三貯藏時間之試驗所使用種球為 95 年 10 月 份購自芊卉種苗公司,鱗莖周徑平均為29.7 公分,重量為 375.1 公克,品種 為 ‘Blossom Peacock’ 。而切花採收適期試驗所採用的品種為重瓣 ‘Lady Jane’ (圖 1C)以及單瓣 ‘Apple Blossom’ 係由所栽培之種球於 96 年春季抽出 的花梗。本研究所使用品種之特性如下:
1、 蘋果花 ‘Apple Blossom’
屬大花單瓣品種,花朵顏色淡粉有如蘋果花而得名,花莖長度約可達 60 cm,每枝花莖可開 4-6 朵花,此品種非常容易自然分球,可供作為盆花及切 花(圖 1A)。
2、 花孔雀 ‘Blossom Peacock’
由日本選育而得。花為重瓣,花瓣外缘為紅色,中肋為白色,每一花莖 有4-6 朵花,花莖長度約為 50~65 cm (圖 1B)。
圖1. 本研究所使用之孤挺花品種:蘋果花(A)、花孔雀(B)、珍女士(C)。
Fig. 1. Hippeastrum hybridum Hort. cvs. Apple Blossom (A)、Blossom Peacock (B)、Lady Jane (C) were used for experiments.
C A
B
3、珍女士 ‘Lady Jane’
屬大花重瓣品種,花朵深橘色,帶淺白色線條,每一花莖約可開
4-6 朵花,適合做為盆花栽培(圖 1C)。
二、 其他材料
試驗所使用之栽培介質為 1:1:1 (v/v)之二號蛭石(南海蛭石工業股份有 限公司)與真珠石(正東瀛股份公司)與泥炭土(荷蘭進口),以介質自動攪拌機 混合均勻備用,栽培容器為使用自華興園藝資材行所購得之八吋橙色圓形花 盆。
三、 試驗環境
全年生育調查及花芽去除與貯藏溫度試驗之栽培環境為宜蘭大學園藝學 系系館二樓陽台,而不同栽植期對鱗莖花芽之影響及貯藏時間之試驗的栽培 環境為本系E 棟簡易溫室;切花試驗於切花採收後,於本系館第二實驗室進 行瓶插試驗,實驗室內溫度控制在25℃左右,而試驗一至試驗三於試驗期間 控制澆水,維持介質之濕潤度,以利植株之生長及發育。
四、 試驗方法
(一)、生育特性調查
1、全年生育調查
本試驗目的在於觀察孤挺花全年鱗莖及葉,根及子球等生長發育之變化 以及鱗莖內部的花芽於全年分化發育之狀況與變化,以了解孤挺花之生育習
性並提供作為生產及花期調節之參考。
試驗地點為宜蘭大學園藝學系系館二樓陽台,於95 年 1 月冬季時從二 樓陽台採收周徑25 公分以上之孤挺花 ‘Apple Blossom’ 品種開花球,於採收 時去除葉片,鱗莖頸部留2-3 公分,清洗乾淨之後放置於平均約 23 ± 1℃的 室溫下3 日待傷口完全癒合後,約留 5-10 公分長的根並將乾燥或有損傷的 根去除後種植於八吋橙色圓形花盆,種植後充分澆水並且於試驗期間維持介 質濕潤度。
從95 年 3 月份下旬開始,每隔四週隨機選取 2 球於開花軸之軸面切開,
以觀察鱗莖的剖面及調查花芽狀況與測量其他生育性狀。
2、不同栽植期對鱗莖內部花芽發育之影響
本試驗目的在利用貯藏後,經不同種植日期而開花後之孤挺花種球對後 期鱗莖內部花芽發育之影響,以探討孤挺花經不同花芽發育狀態對產期調節 之可能性。
試驗地點為宜蘭大學園藝學系簡易溫室E 棟,種球於 95 年 10 月從芊卉 種苗公司購入,置於7℃冰箱中貯放,相對溼度控制於 80%以上,每隔四個 星期取出10 顆,其中 2 球切開觀察鱗莖內部花芽狀況,另外 8 球種植使其 開花,後於96 年 6 月份取球切開觀察其內部花芽發育狀態。
(二)、去除花芽與貯藏溫度及時間對開花之影響
本試驗目的在於利用花芽去除與否及不同冷藏溫度處理來觀察孤挺花 的開花情形以及內部花芽之發育狀況,並作為孤挺花產期調節的可行性。
於95 年 1 月冬季時將從芊卉種苗公司購得之開花球種植後,其中一部
分於花莖抽出時剪除抽出之花莖,而另一部分任其正常開花,而於培養四個 月之後掘起,剪除葉片並且留下5-10 公分的根,之後置於低溫(13℃)及常溫 (23℃)貯放,分別於貯放 15,30,45 天之後取出種植,種植後充分澆水並且 於試驗期間維持介質濕潤度。觀察夏季開花情形,以及於生長四個月後,同 實驗一於每處理隨機選取2 球於開花軸之軸面切開,以觀察鱗莖剖面之花芽 分化及發育狀態。
(三)、貯藏時間對開花之影響
本試驗目的在於利用長期之7℃低溫貯藏來觀察對孤挺花開花性狀之影 響以及調節與分散孤挺花之開花期,並探討作為孤挺花利用貯藏延後產期之 可行性。
試驗地點為宜蘭大學園藝學系簡易溫室E 棟,種球於 95 年 10 月從芊卉 種苗公司購入142 顆孤挺花 ‘Blossom Peacock’ ,於 95 年 10 月 24 日將購入 之開花球測量周徑及重量,留下12 顆種球,10 顆種植,2 顆切開觀察鱗莖 內部花芽狀態,其餘130 顆種球使用兩層報紙包裹並裝箱,置於 7℃冰箱中 貯放,相對溼度控制於80%以上,每隔四個星期取出 10 顆種球量測其周徑 及鮮重損失之百分比,其中2 球切開觀察鱗莖內部花芽狀況,另外 8 球種植 觀察其開花情形,調查第一及第二花莖長度、花莖數量、開花率、到花日數、
第一花莖小花數及第一花莖上第1 至第 4 朵小花之開放天數。種植後充分澆 水,並維持其栽培期間之介質濕潤度。
(四)、不同切花適期對切花瓶插之影響
本試驗目的在於利用不同發育程度的花莖進行瓶插試驗,觀察孤挺花之 切花瓶插情形,並作為孤挺花應用於切花之可行性及切花採收適期之指標。
於96 年春季時,以所栽植的孤挺花品種中,選擇 ‘Apple Blossom’ 及‘Lady Jane’ 為瓶插試驗之品種,而採收標準設定為 3 個階段(圖 2),苞葉期(Spathe stages):小花未突出苞葉、著色期(Color stages):小花突出苞葉並著色及彎 曲期(Bend stages):小花梗與花莖彎曲成約 90°,待花莖達設定之採收階段 時,將花莖從鱗莖頸部位置切除,秤重後插於錐形瓶中,每錐形瓶添加自來 水300 ml,於瓶插後一天換水一次,之後每兩天換水一次,每處理二重複,
每重複3 支,調查項目有瓶插壽命、小花開放率及鮮重變化率。
五、 試驗設計與統計分析
所有試驗皆採完全逢機設計(Complete Randomized Design, CRD)。而本 研究所調查的各資料,採用SPSS 統計分析軟體,以鄧肯氏多變域變方分析 (Duncan’s Multiple-Range Test),檢驗各處理間 5%的差異顯著性。
六、 試驗調查項目說明
1、 花莖長度:第一及第二花莖長度,當花莖上第一朵小花開放時測量,花 莖長度的測量是由鱗莖頸部至花朵下方的花頸部位之長度。
2、 到花日數:為種植後至第一花莖上第一朵小花開放之天數,花朵上以兩 片花瓣打開視為開放。
3、 小花數:為第一花莖上的小花開放數量。
4、 小花開放天數:為小花開放日至小花萎凋日之天數,小花花瓣顏色變深 且失去彈性並皺縮視為萎凋。
5、 瓶插壽命:為切花採收日至瓶插終止日,瓶插終止為 2/3 的小花凋謝或 切花花梗因腐爛折斷而無法繼續觀賞時。
6、 切花鮮重變化:為切花於瓶插期間之鮮重變化百分率,測定方法如下:
鮮重變化(%/日)=(當日花重-前次花重)/原始花重×100/日數。
圖2. 孤挺花切花之不同採收標準:苞葉期,小花未突出苞葉、著色期,小花突出苞葉
且著色、彎曲期,小花梗與花莖彎曲成90°。
Fig. 2. Different harvest standard of Hippeastrum hybridum Hort. for cut flowers:Spathe stages, the florets has not stand out the spathe, Color stages, the florets stand out the spathe and petal color, Bend stages, the floret pedicel and scapes bend to the 90°.
Spathe stages Color stages Bend stages
結 果(Results)
一、生育特性調查 (一)、全年生育調查
1、鱗莖內部花芽之觀察
而從鱗莖的剖面觀察中,06 年 3 月份在球體中可看到一個開花中的花梗 基部及一個花芽,而4 月份則可明顯看到兩個在開花的花梗基部,5 月 18 日 可看到開花後的花梗基部開始乾枯萎縮,而6 月 15 日的鱗莖剖面可以看到靠 近中心軸的位置有一個小的花芽,而於7 月 13 日時可明顯看出花芽開始從內 側的中心軸位置上往鱗莖外圍推出(圖 3)。而於 8 月 10 日的觀察中,鱗莖可看 到兩個花芽,而最大的花芽卻有乾扁的現象,且在接下來的月份除12 月 28 日沒有觀察到之外,幾乎每個月都可以觀察到在開完花之後所分化的第一個花 芽都於鱗莖內乾枯死亡(圖 3)。而於 06 年 10 月 5 日的觀察中,花芽位置已較 7 月份更為外推,而且數量增加達到 3 個(圖 4),最大花芽長度也超過 3 公分,
第二花芽也已超過2 公分(圖 5A),而於 06 年 11 月 2 日至隔年 1 月 25 日,鱗 莖內部並無明顯變化,但於07 年 2 月 22 日最後一次觀察中,花芽已快突出鱗 莖,且於花芽長度上,最大的花芽已超過7 公分,第二花芽的長度也超過 5 公分(圖 5B)。
在花芽數量上,於06 年 3 月 23 日的調查中,平均有 2 個花芽,而於 4 月份和5 月份的開花時節花芽數量為最低,平均為 0.5 個,隨後鱗莖繼續生長 分化花芽,於7 月份數量提升回到 2 個,而 10 月份之後,花芽數量也都超過 2 個(圖 4)。
接續下一頁(next page continuous)
06/03/23 06/04/20
06/05/18 06/06/15
06/07/13 06/08/10
06/09/07 06/10/05
圖3. 孤挺花種球於不同時期解剖情形。箭號所指為花芽或盲芽及花莖基部。
Fig. 3. Anatomy of Hippeastrum hybridum Hort. bulbs in different dates. Arrow showed flower buds or blind buds and base of flower stalk.
06/11/02 06/11/30
06/12/28 07/01/25
07/02/22
Month/Day/Year 3/1/06 4/1/06 5/1/06 6/1/06 7/1/06 8/1/06 9/1/06
10/1/06 11/1/06
12/1/06 1/1/07 2/1/07 3/1/07
No. of flower buds / bulb
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
圖4. 孤挺花於不同時期之花芽數變化。
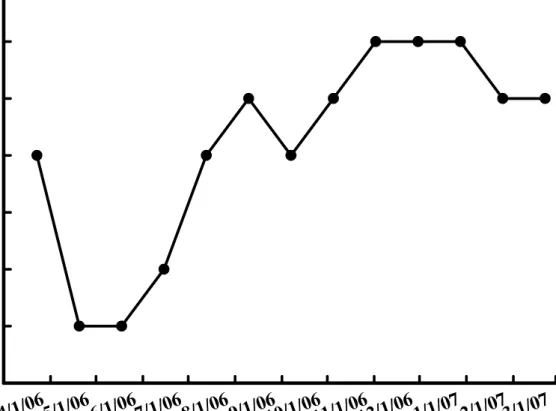
Fig. 4. Changes of number of flower buds on Hippeastrum hybridum Hort. in different dates.
圖5. 孤挺花於 10 月份(A)及 2 月份(B)之花芽長度。
Fig. 5. Length of flower buds of Hippeastrum hybridum in October (A) and February (B).
A
B
2、孤挺花於不同時期之生長變化
(1)、周徑
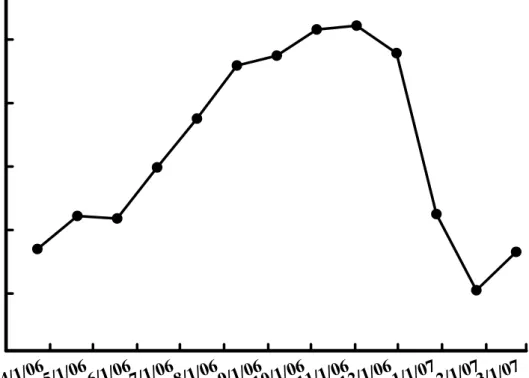
在周徑的變化上,明顯可看出孤挺花於3-4 月的開花期當中,周徑逐漸縮 減變小,而於5 月份的觀察中,周徑甚至降低至 22 公分以下,而於花期過後 的6 月夏季,周徑又逐漸增加變大,且於 11 月底的調查中,周徑已達到 30 公分(圖 6)。
(2)、全株重
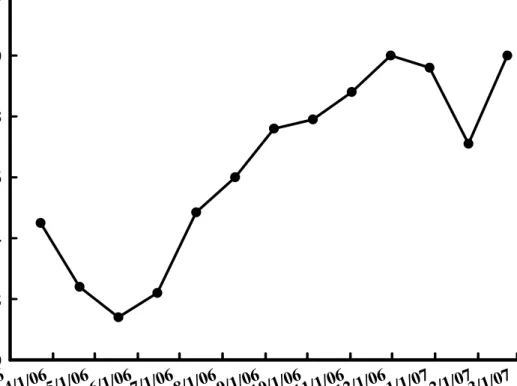
在全株重的調查方面,與周徑相似,亦是在 3-5 月的開花期逐漸降低,甚 至於5 月份,全株鮮重甚至降低至 200 公克以下,而於花期過後的 6 月至 7 月重量明顯迅速增加,並且於9 月份的調查,已接近 600 公克,而於 11 月底 調查可看出,已經將近700 公克,但在 12 月及 1 月份的調查中,重量卻呈現 較低的趨勢,而2 月份則卻又升高(圖 7)。
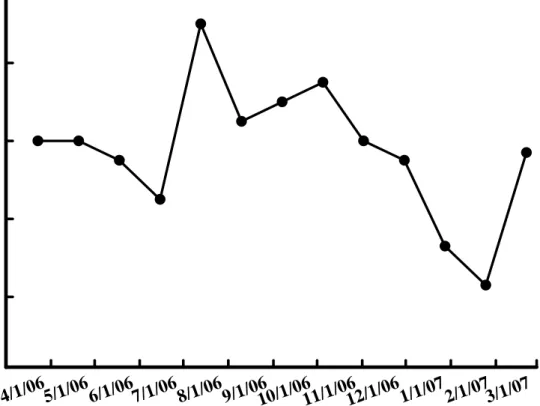
(3)、地下部鮮重
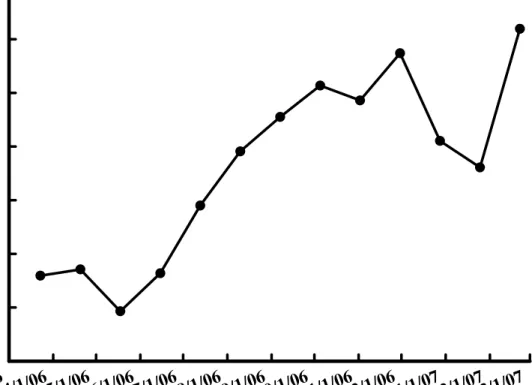
在地下部鮮重方面,在3 月份約為 250 公克,而與周徑和鮮重同樣於 5 月份的調查中達到最低,為近150 公克,而於隨後的生長開始逐漸增加,於 7 月份已超過300 公克,於 10 月份超過 400 公克,而於 11 月底甚至超過 450 公克,之後在12 月卻開始下降,於 1 月份下降至約 350 公克,2 月份才又開 始上升(圖 8)。
(4)、葉片長度
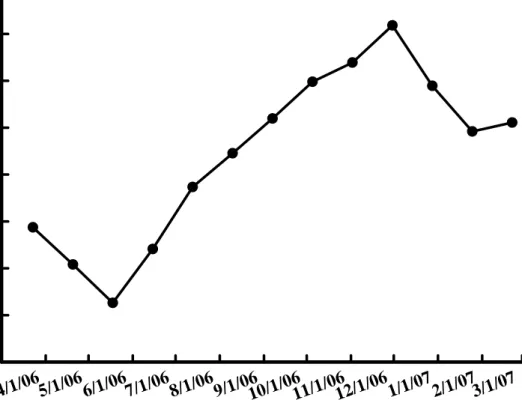
在葉片長度方面,於3 月份一開始的調查中為最低,不到 15 公分,而隨 後開始逐漸增長,於夏季的6 月份,葉片的長度上已超過 40 公分,5 月份至 8 月份為葉片快速增長期,而約於9 月份時生長逐漸趨於緩和,長度約在 80 公 分左右,於10 月初至 11 月底的調查中,葉片長度已經沒有太大的變化,也都
Month/Day/Year 3/1/06
4/1/06 5/1/06
6/1/06 7/1/06
8/1/06 9/1/06
10/1/06 11/1/06
12/1/06 1/1/07 2/1/07
3/1/07
Bulb size (cm)
20 22 24 26 28 30 32
圖6. 孤挺花周徑於不同時期之變化。
Fig. 6. Changes of bulb size of Hippeastrum hybridum Hort. in varies dates.
Month/Day/Year 3/1/06
4/1/06 5/1/06
6/1/06 7/1/06
8/1/06 9/1/06
10/1/06 11/1/06
12/1/06 1/1/07 2/1/07
3/1/07
Fresh weight (g)
100 200 300 400 500 600 700 800
圖7. 孤挺花全株鮮重於不同時期之變化。
Fig. 7. Changes of fresh weight of Hippeastrum hybridum Hort. in varies dates.
Month/Day/Year 3/1/06
4/1/06 5/1/06
6/1/06 7/1/06
8/1/06 9/1/06
10/1/06 11/1/06
12/1/06 1/1/07 2/1/07
3/1/07
Bulb and root weight (g)
100 150 200 250 300 350 400 450 500
圖8. 孤挺花地下部重量於不同時期之變化。
Fig. 8. Changes of bulb and root weight of Hippeastrum hybridum Hort. in varies dates.
Month/Day/Year 3/1/06
4/1/06 5/1/06
6/1/06 7/1/06
8/1/06 9/1/06
10/1/06 11/1/06
12/1/06 1/1/07 2/1/07
3/1/07
Leaf length (cm)
0 20 40 60 80 100
圖9. 孤挺花葉長於不同時期之變化。
Fig. 9. Changes of leaf length of Hippeastrum hybridum Hort. in varies dates.
是在80 公分上下,但於 12 月份的調查中,葉片迅速下降至近 20 公分,1 月 份甚至沒有葉片生長,而於2 月份則又開始上升(圖 9)。
(5)、葉片數
而在葉片的數量變化方面,在 3 月到 4 月期間,葉片數量約在 4 片左右,
而於6 月份到 7 月份夏季期間,葉片快速生長抽出,由 6 月份的不到 3 片迅速 抽出成7 片葉子,且於不同期間的調查中,7 月份所調查的葉片數量為最高,
而隨後在8 月份的調查則又開始下降至不到 5 片,而於 8 月份到 10 月份的夏 秋季轉換期之間,葉片數量亦都在5 片左右,且在 11 月份葉片數量就逐漸地 萎黃凋謝而一直減少,且於冬季的 1 月份,幾乎沒有葉片的生長,而於 2 月份 的調查,葉片數量才恢復到4 片左右(圖 10)。
(6)、根長
在根長方面,3 月至 5 月之間的開花期,根沒有明顯生長情形,長度皆約 在20 公分以下,而於花期過後,5 月至 7 月份根急速生長,6 月份已接近 30 公分,而在7 月份根的生長速度開始趨於平緩,7 月份之後根長都大約保持在 35 公分上下,至隔年 2 月份亦都保持在 35 公分左右,而無太大的差異(圖 11)。
3、分生子球之觀察
在不同時期的鱗莖解剖中發現,從7 月份開始之後的每個月份(圖 12),幾 乎都可在孤挺花鱗莖內較外層單位的鱗片中觀察到有子球分蘗增生之現象,即 開完花或盲芽的單位。而於鱗莖外的分生子球幾乎在全年每個月份都可以觀察 到。而且觀察中發現,鱗莖內部的子球著生位置皆於每個枝葉組的最內層全周 葉外側,且在觀察中,明顯可看出與花芽不同的是,花芽會與黃色的莖盤有一 段黃色結構的連結,但小鱗莖則沒有,且不管此枝葉組有3 或是 5 片鱗片,小 鱗莖都固定與花芽隔兩層鱗片,即一片全周葉以及一片半周葉,而其他的鱗
Month/Day/Year 3/1/06
4/1/06 5/1/06
6/1/06 7/1/06
8/1/06 9/1/06
10/1/06 11/1/06
12/1/06 1/1/07 2/1/07
3/1/07
No. of leafs
0 2 4 6 8
圖10. 孤挺花葉數於不同時期之變化。
Fig. 10. Changes of number of leaf of Hippeastrum hybridum Hort. in varies dates.
Month/Day/Yeat 3/1/06
4/1/06 5/1/06
6/1/06 7/1/06
8/1/06 9/1/06
10/1/06 11/1/06
12/1/06 1/1/07 2/1/07
3/1/07
Root length (cm)
15 20 25 30 35 40
圖11. 孤挺花根長於不同時期之變化。
Fig. 11. Changes of root length of Hippeastrum hybridum Hort. in varies dates.
圖12. 子球於不同時期在鱗莖內發育之狀況。箭號處為子球著生位置。
Fig. 12. State of bulblets development in the bulbs of varies dates. Arrow showed the staring site of the bulblets.
06/11/30 06/11/02
06/10/05 06/09/07
06/08/10 06/07/13
片葉中則沒有發現有子球自然分生,而以軸線來看,子球的著生位置與花芽 著生位置相同,也就是亦固定著生在開花軸線上。而在除了開花軸線之外的位 置,於鱗莖內部的觀察中,完全沒有分生子球,鱗莖外則是幾乎沒有分生子球 或是與開花軸線上所增生的子球相比較,都相對較小且少。
(二)、不同栽植期對鱗莖內部花芽發育之影響
孤挺花種球經由不同栽植期後,於07 年 6 月的種球解剖觀察顯現,於 06 年10 月 24 日栽植者,至隔年 6 月時,花芽已超過鱗莖頸部,而 11 月 21 日及 12 月 19 日栽植者,花芽已近鱗莖頸部,1 月及 2 月所栽植者,花芽則相對較 小(圖 13)。
二、去除花芽與貯藏溫度及時間對開花之影響
於試驗中,所有處理在夏季時節之開花率皆不到20%或是等於零(未顯示 數據)。而於 11 月中旬從每處理中選取兩球作切球觀察,觀察其中之花芽狀 態,發現鱗莖內之花芽大都處於剛分化完成左右之階段,且於鱗莖中也發現有 乾扁的花芽(圖 14B 和 C,圖 15A 和 B,圖 16B 和 C,圖 17A、B 和 C)。而貯 放前的花芽去除處理與對照組(任其自然開花)相比,對種球花芽分化及發育的 影響差異不甚明顯。本實驗中,低溫及常溫貯放對種球的開花及花芽也不明 顯,貯放天數亦然。夏季時開花率低,而且時至12 月初的觀察,仍無明顯的 開花現象。
三、貯藏時間對開花之影響
於貯藏期間,鱗莖的周徑及球重逐漸減少,周徑的變化率較少且平緩,而 重量的變化在貯藏前期明顯快速下降,於貯藏12 週時調查的結果顯示,鮮重 已減少近5%,貯藏至 28 週時,鮮重甚至下降到原始重量的 92%以下(圖 18)。
圖13. 孤挺花鱗莖經不同栽植期之解剖情形。
Fig. 13. Anatomy of Hippeastrum hybrudum Hort. bulbs by varies from planting times. Arrow shows the flower buds.
06/10/24
06/02/13
06/01/16 06/12/19
06/11/21
圖14. 花芽切除與低溫貯放 15(A)、30(B)及 45(C)天鱗莖解剖情形。
Fig. 14. Anatomy of bulbs removed flower buds and low temperature storage for 15(A), 30(B) and 45(C) days. Arrow shows the flower bud or blind buds.
A
C B
圖15. 花芽不切除與低溫貯放 15(A)、30(B)及 45(C)天鱗莖解剖情形。
Fig. 15. Anatomy of bulbs not removed flower buds and low temperature storage for 15(A), 30(B) and 45(C) days. Arrow shows the flower bud or blind buds.
A
B C
圖16. 花芽切除與常溫貯放 15(A)、30(B)及 45(C)天鱗莖解剖情形。
Fig. 16. Anatomy of bulbs removed flower buds and room temperature storage for 15(A), 30(B) and 45(C) days. Arrow shows the flower bud or blind buds.
B C
A
圖17. 花芽不切除與常溫貯放 15(A)、30(B)及 45(C)天鱗莖解剖情形。
Fig. 17. Anatomy of bulbs not removed flower buds and room temperature storage for 15(A), 30(B) and 45(C) days. Arrow shows the flower bud or blind buds.
A
B C
Weeks after storage
0 4 8 12 16 20 24 28 32
Percentage (%)
90 92 94 96 98 100 102
Bulb size Fresh weight
圖18. 孤挺花於 7 ℃貯藏下周徑與鮮重損失百分率之變化。
Fig. 18. Changes of bulbs size and fresh weight loss percentage of Hippeastrum hybridum Hort. under 7 ℃ storage.
而隨著貯藏時間的增加,有發現部分鱗莖於貯藏時有腐爛的現象發生,在貯藏 20 週以下,尚無發現此現象,但於貯藏 24、28 及 32 週時,即發現鱗莖中開 始有腐爛的現象發生,貯藏24 週時,鱗莖內只有花芽部位稍微有腐爛(圖 19A),而貯藏至 28 週時,此現象已明顯可在少數鱗莖內可觀察到(圖 19B),
而貯藏至32 週時,有腐爛現象的鱗莖內已經相當程度的嚴重腐爛現象,幾乎 整個種球都完全腐爛(圖 19C)。
試驗結果顯示,第一個花莖及第二個花莖的長度都隨著貯藏時間的增加,
而逐漸縮短,貯藏0 到 16 週的第一個花莖,長度在 34~46 公分之間,而沒有 顯著性的差異,而貯藏28 週的種球於種植後,並沒有產生第二個花莖,從貯 藏0 到 12 週抽出的第二個花莖,長度都較第一個花莖為長,在 51~61 公分之 間,而抽出的花莖數量由試驗開始的2 支到貯藏 28 週時只剩抽出 1 支,且達 顯著性差異。而到花日數方面,除貯藏0 到 16 週的處理,沒有顯著性差異之 外,之後的貯藏則隨著時間的增加而縮短。而於貯藏0 到 16 週的處理,到花 日數在44~49 天之間,而貯藏 20 週之後,到花日數明顯開始減少至約 30 天 以下。各處理的開花率方面,亦隨著貯藏時間的增加而開花率逐漸減少,於貯 藏0~12 週的處理上,開花率保持在 100%,而貯藏 16、20 和 24 週的開花率 分別為87.5%、75%及 75%,而貯藏到 28 時,開花率明顯不到一半,為 37.5%(表 1)。
而在第一個花莖上小花方面,小花的數量上沒有明顯的差異,只有稍微隨 著貯藏的時間增加而減少,貯藏0 到 24 週的小花數量為在 3.7~4.8 之間,而 貯藏至28 週時,小花的數量則平均降至只剩 3 朵。而於第一花梗上的小花開 放天數中,明顯可看出第一朵至第四朵的開放天數有下降的趨勢,在貯藏0~8 週的小花開放天數,第一到第四朵都有約超過10 天,而於之後則慢慢減少至 不到10 天,甚至於貯藏 28 週之後的開放天數,平均已下降至 5 天左右(表 2)。
圖19. 孤挺花於 7℃貯藏下種球腐爛情形:貯藏 24 週(A)、貯藏 28 週(B)、貯藏 32 週(C)。
Fig. 19. Condition of bulb rotten of Hippeastrum hybridum Hort. under 7 ℃ storage for 24 weeks (A), 28 weeks (B) and 32 weeks (C).
A
B C