行政院國家科學委員會專題研究計畫 期中進度報告
人類 DNA 修復基因 hHR23A 與 hHR23B 之功能研究(1/3)
計畫類別: 個別型計畫 計畫編號: NSC92-2314-B-006-148- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立成功大學醫事技術學系 計畫主持人: 黃溫雅 計畫參與人員: 沈芳晴 蔡漢霓 謝惠娟 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 5 月 31 日
行政院國家科學委員會補助專題研究計畫
ˇ 期 中
進度報告
人類 DNA 修復基因 hHR23A 與 hHR23B 之功能研究(1/3)
計畫類別:ˇ 個別型計畫 □ 整合型計畫
計畫編號:NSC
92-2314-B-006-148
-
執行期間:92 年 08 月 01 日至 93 年 07 月 31 日
計畫主持人:黃溫雅
共同主持人:
計畫參與人員: 沈芳晴,謝惠娟,蔡漢霓
成果報告類型(依經費核定清單規定繳交):ˇ精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列管計
畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:成大醫技系
中 華 民 國 93 年 05 月 31 日
The involvement of the human homologs of the S. cerevisiae Rad23 A and B proteins
in nucleotide excision repair by RNA interference
Hui-Chuan Hsieh4, Hsien-Chang Chang4, Yu-Hsin Huang3, Fan-Ching Shen2, Yu-Ting Lai3, Tammy Wang3, Yi-Hsuan Hsieh1, Han-Ni Tsai2, Jui-He Tsai2 and Wenya Huang1,2,3
1
: Institute of Basic Medical Sciences; 2: Department of Medical Science and Biotechnology, 3: Institute of
Molecular Medicine,College of Medicine; 4: Institute of Biomedical Engineering, College of Engineering,
National Cheng-Kung University, Tainan 70101.
This work was supported by the National Scientific Council, Taiwan.
*
: To whom correspondence and requests for reprints should be addressed, at Department of Medical Technology, College of Medicine, National Cheng Kung University, Tainan 70101, Taiwan. Phone: (886) 6-235-3535 ext. 5766; Fax: (886) 6-236-3956; E-mail: [email protected]
Running title: hHR23 proteins in nucleotide excision repair by RNA intereference
Keywords: hHR23; Rad23; nucleotide excision repair; DNA repair; RNA interference
The abbreviations used are: NER, nucleotide excision repair; hHR23, human homolog of Rad23; RNAi, RNA interference; siRNA, small interfering RNA; KD, knock-down.
摘要
hHR23A 與 hHR23B 為酵母菌 Rad23 基因的同源基因。在本研究中我們已 RNA 甘擾技術來建構干 擾基因以了解這兩個基因的功能。我們發現,除了 hHR23B 以外,hHR23A 對於核苷酸切除修復機 制的進行也是非常重要的。總個而言,我們在此研究中將 hHR23A 與 hHR23B 的功能做了清楚的 陳述。
ABSTRACT
HHR23A and hHR23B are the human homologs of the S. cerevisiae Rad23. The hHR23B is associated with the NER factor xeroderma pigmentosum C protein ans is required for global genome repair. The function of hHR23A is not clear yet. In this study, we study the functions of hHR23A and hHR23B proteins by RNA interference techniques. The hHR23A and B siRNA constructs were shown able to diminish RNA levels of hHR23A or B proteins to around 50%, while the siRNA against one gene did not interfere the other hHR23 gene expression. Based on the analysis of Southwestern immunoblot and host cell reactivation assays, the hHR23A and B siRNA constructs caused decrease in DNA repair activity, indicating that the hHR23A and B are involved in nucleotide excision repair.
INTRODUCTION
Nucleotide excision repair is the major pathway for repairing DNA lesions that cause structural distortion. The DNA damage caused by ultraviolet irradiation and chemical cross-linking agent cisplatin are important examples as subjects for NER. The disease caused by NER deficiency is designated xeroderma pigmentosum (XP), which is characterized as skin pigmentosum, and ultimately metastatic skin cancer. In most of the cases, the individuals severely affected by XP have life span shorter than 20 years.
The XP genes are designated from XPA to XPG, each of which is involved in one (or more) step in NER. Among them, XPC is the major factor for damage recognition in the subpathway of global genome repair (GGR), which is involved in repairing DNA lesions on non-transcribed genes or on the non-transcribed strands of the active genes. hHR23A and B are human sequence homologs of the S. cerevisiae Rad23. The hHR23B is associated with XPC physically and functionally. Though highly homologous with hHR23B, the hHR23A has not been identified for its role in NER in vivo. Previous studies by using knock-out animal models have reported that the hHR23A is functionally redundant with hHR23B, and defects in hHR23A does not cause NER deficiency. In this study, we studied its role in cells by RNA interference techniques. We found that besides hHR23B, hHR23A is also required for NER.
MATERIALS AND METHODS
Plasmids, cell lines and antibodies - The RNA interference constructs for hHR23A and B were cloned into pSUPER (a generous gift from A, Netherlands). The DNA sequences used to generate hHR23A interfering RNA constructs are listed in Table 1. The DNA fragment of the interfering sequences were annealed into double-stranded DNA then cloned into pSUPER at the HindIII and BglII restriction enzyme sites. These constructs for RNA interference are sequenced for verification.
The human cervical cancer cell HtTA1 was used for the study. The cells were maintained in regular Dulbecco's modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 1× non-essential amino acid, and 1× of antibiotic/antimycotic mixture (Gibco BRL, Grand Island, NY). The cells were grown at 37°C with 5% CO2. Most of the reagents used for the reverse
transcription and PCR amplifications were purchased from Promega (Madison, WI). The specific antibody against hHR23A was derived from rabbit against the peptide, LADISDVEGEVG. The antibody against the hHR23B was purchased from Abgent, inc. Common chemicals used were purchased from Sigma Chemical Co. (St. Louis, MO).
Plasmid transfection – Lipofectamine reagent was used for plasmid transfection experiments. Briefly, 1
µg of the plasmid to be transfected was diluted to 50µl with OPTI-MEM®1 reduced serum medium, which contains 2% FBS. 2 µl of LipofectamineTM 2000 reagent was also diluted to 50 µl with OPTI-MEM®1, then allowed to sit for 5 minutes at room temperature. Solutions of the plasmid and LipofectamineTM 2000 were then mixed and allowed to sit for 20 minutes. The mixture solution was added to the culture cells in a 24-well plate, which had been transferred to the OPTI-MEM®1 medium. After transfection, the cells were incubated at 37°C for 20 hours in the presence of 5% CO2. The culture
medium was then switched to regular MEM medium for further incubation. To select for XPCWT/pcDNA3.1B-transfected cells, which are neomycin-resistant, the antibiotic G418 (1mg/ml) was supplemented to the growth medium.
MTT assay – A 3-[4, 5-dimethylthiazol-2-yl]-2, 5-diphenyl tetrazolium bromide (MTT) cell proliferation assay was used to detect cells’ sensitivity to UVC irradiation [12]. Briefly, cells were grown to 1 × 105 / well in a 24-well culture plate. The culture medium was removed, and cells were washed once with phosphate-buffered-saline (pH 7.4) and treated with 3 or 5 Joules/m2 of UVC irradiation. After 48 hours of incubation at 37ºC, half of the medium was removed. 0.5 mg/ml of MTT labeling reagent (Sigma) was added to each well with the UVC-treated or mock-treated cells. After 4 hours of incubation at 37˚C, 10% SDS/0.01 M HCl solubilization solution was added to wells, then cells were incubated at 37˚C for overnight. On the next day, the O.D.590nm was measured and converted to cell survival percentage as
compared to control cells.
Southwestern immunoblot – The HtTA1 cells transfected with RNA interference for hHR23A or hHR23B were irradiated with 254 nm ultraviolet lamp. The cells were incubated for various periods of time for DNA repair. After incubations, the genomic DNA in cells were extracted by standard phenol/chloroform methods, and diluted to 1 µg/µL in 0.3 N NaOH to denature the DNA strands. A total of 15 µg of genomic DNA from each set of cells was spotted on Nylon+ membrane then probed with antibody specifically against 6,4 photoproduct (a generous gift from Dr. , Japan), based on standard western blot analysis. After incubation with the primary antibody for one hour, the membrane was washed 3 times in TBST buffer, supplemented with 5% skim milk. The anti-mouse secondary antibody in the same buffer was then incubated with the membrane. The signals of 6, 4-photoproducts were visualized by ECL.
Host-cell reactivation assay - Host-cell reactivation activities on cisplatin-damaged plasmids were used to detect DNA repair activity in cells. To construct the plasmid substrate, pCMVluc DNA was treated in the dark with 5% cisplatin at 37°C for 2 hours. The treated plasmid DNA was purified by ethanol precipitation. To test the DNA repair activity in HtTA1 cells expressing various types of RNA intereference constructs, cisplatin-damaged pCMVluc as well as the hHR23A or hHR23B RNAi constructs were co-transfected into the HtTA1 cells. After transfection for 72 hours, cell lysates were collected and the luciferase activities were detected using a luciferase assay kit (ABI; Applied Biosystems).
RESULTS and DISCUSSION
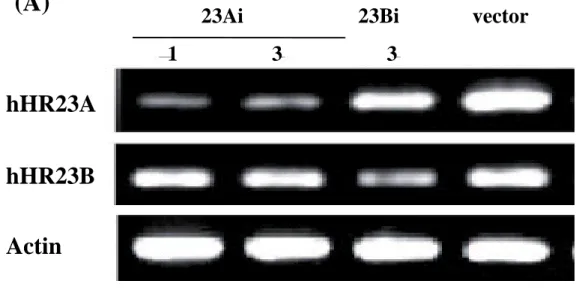
Inhibition of hHR23A and hHR23B expression by RNA interference - To specifically unhibit expression of hHR23A or hHR23B, the RNA interference constructs were generated. The sequences used to construct the RNAi plasmid for each of the hHR23A and B were derived form the regions that contain unique sequences. The data depicted in Figure 1 indicate that the RNAi construct against hHR23A was able to knock down the level of endogenous hHR23A by around 60%, as shown by RT-PCR and western blot. The RNAi construct against hHR23B knock down the level of hHR23B by 50%. And there was no cross-inhibition between two RNAi constructs. This indicates that the specific RNA interference against hHR23A or B could be used as a model for functional study of hHR23A and B in vivo.
Inhibition of nucleotide excision repair by RNA interference against the hHR23A and hHR23B genes - The DNA repair activity in HtTA1 cells, in which the hHR23A or hHR23B gene expressions were knocked down by RNA interference, was examined. The data presented in Figure 2 indicate after UV irradiation on the HtTA1 cells transfected with hHR23A or hHR23B RNAi KD constructs, the survival rates of cells were reduced. Survival rates of the hHR23Akd cells were reduced by 20%, and those of the
hHR23Bkd cells were reduced by 50%. The UV survival rates in the hHR23A and B double KD were lower than either of the single KD cells. This finding suggests that the endogenous hHR23A and B factors are required for NER.
To investigate the functional defects caused by the hHR23Akd and hHR23Bkd constructs, the host cell reactivation on the cisplatin-damaged pCMVLuc reporter plasmid was detected. Data depicted in Figure 3A indicate that the cells transfected with hHR23Akd and hHR23Bkd exhibited decreased DNA repair activity for recovery of cisplatin-DNA adducts. The hHR23Akd construct diminished the DNA repair activity by 30%, while the hHR23Bkd construct diminished the DNA repair by 50%. The double KD constructs diminished the DNA repair activity to higher extend. Therefore, though the RNAi knock-down constructs used in this study could not completely diminish the expression of endogenous hHR23A or B, the DNA repair activities have been seen significantly affected. To confirm the findings in HCR, analysis of Southwestern immunoblot was also employed in the cells carrying hHR23Akd or hHR23Bkd constructs. The data presented in Figure 3B clearly demonstrate that after transfected with hHR23Akd or hHR23Bkd constructs, the HtTA1 cells exhibited lower activity for repairing 6, 4-photoproducts, indicating that the hHR23A as well as hHR23B are involved in NER.
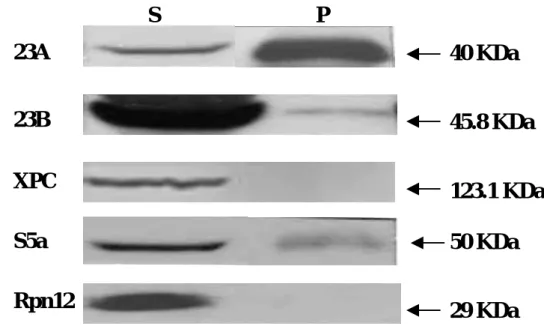
Protein complexes of hHR23A and B – hHR23A and hHR23B have been shown to interact with each other in vitro. Here we test for the protein complexes of hHR23A/B in vivo. By analysis of immunoprecipitation using the antibody against hHR23A, the hHR23B was shown to immuno-precipitate with the hHR23A. The proteasome factor S5A and ubiquitin, previously reported to associate with hHR23A and B, also appear in the same complex (Figure 4a). The Since the functional domains in hHR23A and B are quite homologous, it was speculated that the hHR23A/B proteins form homo- or hetero-oligomers in vivo. The ubiquitin-like (UBL), ubiquitin-associated (UBA) 1 and 2 functional domains were cloned into yeast two-hybrid vectors pGBKT7 (Gal4-BD) or pACT2 (Gal4-AD) and tested for their potential mutual interactions. The data summarized in Figure 4b indicate that the hHR23A and hHR23B directly interact with each other. This finding is consistent with our other study in analysis of immuno-precipitation. The hHR23B deleted of the UBL domain was also able to directly interact with the full-length hHR23A, indicating that the hHR23B associates with the hHR23A at the C-terminus.
The hHR23B has been shown to associated with xeroderma pigmentosum C protein, which is essential for global genome repair of NER. The function of hHR23A in vivo has not been reported. In this study we demonstrate the role of hHR23A in NER, suggesting that not only hHR23B, the hHR23A is also an NER factor.
REFERENCES
[1] E. C. Friedberg, G.. C. Walker, W. Siede, DNA repair and Mutagenesis, ASM, Washington DC, 1995. [2] P. C. Hanawalt, Transcription-dependent and transcription-coupled DNA repair responses, in: V. A. Bohr et al. (Eds.), DNA Repair Mechanisms, Munksgaard, Copenhagen, 1992, pp. 231-242.
[3] K. S. Sweder, P. C. Hanawalt, Preferential repair of cyclobutane pyrimidine dimers in the transcribed strand of a gene in yeast chromosomes and plasmids is dependent on transcription, Proc. Natl. Acad. Sci. USA. 89 (1992) 10696-10700
Fig. 1. Inhibition of expression levels of hHR23A and hHR23B by RNA interference.
(A)
(B)
hHR23A
hHR23B
Actin
23Ai 23Bi vector
1 3 3
hHR23A
hHR23B
Actin
1 3 3
Fig. 2. Cell survival of hHR23Akd and hHR23Bkd cells to UV irradiation.
(A)
0 20 40 60 80 100 120 0 5 10 15 20 25 30UV dose (J/m
2)
Su
rv
iv
al
p
e
rcen
ta
ge
(%)
0
µ
g 2
µ
g 4
µ
g 2
µ
g 4
µ
g
C Ai Ai Bi Bi
Ai+Bi V V
4
µ
g 2
µ
g 4
µ
g
(B)
Fig. 3 0 0.2 0.4 0.6 0.8 1 1.2(A)
Fig 4
(A)
(B)
pGBKT7 pACT2 Interaction
hHR23A hHR23B
+
hHR23B hHR23B
++
hHR23A
hHR23A
ΔUBL++
hHR23A UB
+++
hHR23B UB
+++
UB
hHR23A
ΔUBL++
S P
40 KDa
45.8 KDa
123.1 KDa
50 KDa
29 KDa
23A
23B
XPC
S5a
Rpn12
Table 1
Oligomer Oligomer sequence inhibited domain 23Ai-1(774-792)-forward 5’-GATCCCCGCTCCAGCAGCTGG GCCAGTTCAAGAGACTGGCCCA GCTGCTGGAGCTTTTTGGAAA-3’ XPC binding domain 23Ai-1(774-792)-reverse 5’-AGCTTTTCCAAAAAGCTCCAG CAGCTGGGCCAGTCTCTTGAACT GGCCCAGCTGCTGGAGCGGG-3’ 23Ai-3(534-552)-forward 5’-GATCCCCGCGGGTCGTGGCCG CCCTGTTCAAGAGACAGGGCGG CCACGACCCGCTTTTTGGAAA-3 UBA1 domain 23Ai-3(534-552)-reverse 5’-AGCTTTTCCAAAAAGCGGGTC GTGGCCGCCCTGTCTCTTGAACA GGGCGGCCACGACCCGCGGG-3 23Bi-3(615-633)-forward 5’-GATCCCCGCAAGTAATTGCAG CCCTGTTCAAGAGACAGGGCTG CAATTACTTGCTTTTTGGAAA-3’ UBA1 domain 23Bi-3(615-633)-reverse 5’-AGCTTTTCCAAAAAGCAAGTA ATTGCAGCCCTGTCTCTT GAAC AGGGCTGCAATTACTTGCGGG-3’