Swelling activated chloride currents in the electrical activity of pulmonary vein cardiomyocytes
S. H. Lee¶, Y. C. Chen§, S. Y. Chen†, C. I. Lin§, Y. J. Chen* and S. A. Chen‡
*Taipei Medical University; †Taipei Medical University-Wan Fang Hospital; ‡National Yang-Ming University, School of Medicine; §National Defence Medical Center; ¶Shin Kong Wu Ho-Su Memorial Hospital, Fu Jen Catholic University, Taiwan
ABSTRACT
Background Pulmonary veins (PVs) contain cardiomyocytes with a high arrhythmogenicity for inducing atrial fibrillation. The swelling-activated outwardly rectifying Cl– currents (ICl,swell) are important in the electrical activity of cardiomyocytes. This study was to investigate whether ICl,swell play a role in the PV electrophysiological characteristics.
Materials and methods A whole-cell patch clamp was used to investigate the action potentials and ICl,swell in isolated rabbit single PV and atrial cardiomyocytes during immersion in isotonic (290–300 mosm L–1) and hypotonic (220–230 mosm L–1) solutions. The cell length and cell width were measured using confocal microscopy.
Results Hypotonic solution induced larger ICl,swell in the PV cardiomyocytes with pacemaker activity than those in the PV cardiomyocytes without pacemaker activity or atrial cardiomyocytes. Hypotonic solution shortened the action potential duration and increased the cell width to a greater extent in the PV cardiomyocytes than in the atrial cardiomyocytes. Moreover, hypotonic solution decreased the PV firing with a decrease in the transient inward currents and delayed after depolarizations.
Conclusions These findings suggest that the ICl,swell plays an important role in the electrical activity of the PV cardiomyocytes.
Keywords Atrial fibrillation, hyposmolarity, pulmonary vein, swelling activated chloride currents.
Eur J Clin Invest 2008; 38 (1): 17–23
Introduction
Atrial fibrillation is the most common cardiac arrhythmia seen in clinical practice and induces cardiac dysfunction and strokes [1,2]. Myocardial ischaemia has been shown to be an important factor in the occurrence of atrial fibrillation [3,4].
However, the underlying mechanisms of the pathophysiological mechanisms still remain unclear. The metabolites which accumulate in cardiomyocytes during ischaemia would increase the intracellular osmolarity and cause the development of an osmotic gradient between the intracellular and extracellular milieu and produce cell swelling [5]. The swelling-activated Cl– currents (ICl,swell) have been found in mammalian cardiomyocytes and modulate the cardiac electrical activity [6–10]. Previous studies have shown that ICl,swell will shorten the action potential (AP) duration and facilitate the genesis of re-entrant circuits [10].
Therefore, ICl,swell may play an important role in the pathophysiology of atrial fibrillation during myocardial ischaemia.
Pulmonary veins (PVs) are known to be important sources of ectopic beats which initiate paroxysmal atrial fibrillation or are the foci of ectopic atrial tachycardia and focal atrial fibrillation [11–13].
PVs also play a vital role in the maintenance of atrial fibrillation [14,15]. Previous studies have shown that PVs contain a mixture of working myocardium and pacemaker cells with distinctive electrical activity and high arrhythmogenesis [16–20]. PV cardiomyocytes receive their blood supply from the left circumflex coronary artery. Therefore, circumflex coronary artery disease may induce ischaemia in PV cardiomyocytes and produce cell swelling. Nevertheless, knowledge about ICl,swell in the PV cardiomyocytes has been limited. Since different cardiomyocytes have dissimilar ICl,swell [21], ICl,swell may play a crucial role in the PV electrical activity and produce ischaemia-induced atrial fibrillation. The purpose of the present study was to compare the ICl,swell among the PV and atrial cardiomyocytes and to investigate the role of the ICl,swell in PV arrhythmogenesis.
Materials and methods
Isolation of PV cardiomyocytes
PV and atrial cardiomyocytes were enzymatically dissociated using the same procedure as described previously [22,23]. In brief, the heart with the lungs was removed from anesthetized rabbits after a mid-line thoracotomy. The PV and atrial cardiomyocytes were perfused in a retrograde manner via a polyethylene tubing cannulated through the aorta and left ventricle into the left atrium. The free end of the polyethylene tube was connected to a Langendorff perfusion column for perfusion with oxygenated normal Tyrode’s solution at 37 °C [containing:
NaCl 137, KCl 5·4, CaCl2 1·8, MgCl2 0·5, N-(2-Hydroxyethyl) piperazine-N′-(2-ethanesulfonic acid) (HEPES) 10 and glucose 11 mm; the pH was adjusted to 7·4 by titrating with 1 N NaOH]. The perfusate was replaced with oxygenated Ca2+-free Tyrode’s solution containing 300 units mL–1 of collagenase (Type I, Sigma, ST. Louis, Missouri, USA) and 0·25 units mL–1 of protease (Type XIV, Sigma) for 8–12 min. The atrial and proximal PVs (8–12 mm) were isolated and were gently shaken in Ca2+-free oxygenated Tyrode’s solution until single
cardiomyocytes were obtained. The solution was then gradually changed to normal oxygenated Tyrode’s solution. The cells were allowed to stabilize in the bath for at least 30 min before the experiments.
Measurement of cell width and length in hypotonic and isotonic solutions
The PV and atrial cardiomyocytes were placed in an experimental chamber of a Zeiss LSM 510 laser scanning microscope (Axiovert 100 A, Carl Zeiss Co., Jena, Germany). The cells were allowed to settle on the bottom of the experimental chamber for 10 min. The cell width and length were measured in a frame scan mode composed of 512 × 512 pixels while immersed in the isotonic and hypotonic solutions at 35 ± 1 °C.
Electrophysiological study
A whole-cell patch-clamp was performed using an Axopatch 1D amplifier (Axon Instruments, Foster City, CA, USA) at 35 ± 1 °C, and borosilicate glass electrodes (outer diameter, 1·8 mm) were used, with tip resistances of 3–5 MΩ. Before the formation of the membrane-pipette seal, the tip potentials were zeroed in Tyrode’s solution (containing: NaCl 137, KCl 5·4, CaCl2 1·8, MgCl2 0·5, HEPES 10 and glucose 11 mm; the pH was adjusted to 7·4 by titration with 1 N NaOH). The AP and transient inward currents of the cardiomyocytes were measured during superfusion with the isotonic solution (290–300 mosm L–1) and hypotonic solution (220–230 mosm L–1). The isotonic solution contained (in mm): NaCl 100, KCl 5, MgCl2 1, CaCl2 1·5, glucose 10, HEPES 10 and mannitol 70, titrated to a pH of 7·4 with NaOH. The components of the hypotonic solution (220–230 mosm L–1) were the same as those for
the isotonic medium except that mannitol was omitted. The APs and ionic currents were recorded in the current-clamp mode and in the voltage-clamp mode, respectively [20]. A small
hyperpolarizing step from a holding potential of –50 mV to a testing potential of –55 mV for 80 ms was delivered at the beginning of each experiment. The area under the capacitative currents was divided by the applied voltage step to obtain the total cell capacitance. Voltage command pulses were generated by a 12-bit digital-to-analog converter controlled by pCLAMP software (Axon Instruments). Recordings were low pass-filtered at half the sampling frequency. Data were sampled at rates varying from 2 to 25 kHz.
The AP duration at 90% (APD90) of the full repolarization was measured during APs elicited by 2 Hz electrical stimulations. Only APs with stable morphology during paired stimulation were selected for measurement. The delayed after depolarization was defined as the presence of a spontaneous depolarization of the impulse after full repolarization had occurred. The micropipettes were filled with a solution containing (in mm) KCl 20, K aspartate 110, MgCl2 1, Mg2ATP 5, HEPES 10, EGTA 0·5, LiGTP 0·1, and Na2 phosphocreatine 5, titrated to a pH of 7·2 with KOH for the transient inward currents and AP experiments. To measure the activity of the volume-regulated Cl– channels, KCl was replaced by CsCl in the media and the pipette solutions contained (in mm):
CsCl 40, Cs-aspartate 100, MgCl2 1, CaCl2 1·93, EGTA 5, ATP 2, GTP 0·5, and HEPES 5. The pipette solutions were adjusted to a pH of 7·2 with CsOH. The recordings were low pass-filtered at half the sampling frequency. Data were sampled at rates varying from 2 to 25 kHz.
The current-voltage relationship of the ICl,swell was measured as the difference between the elicited chloride currents while immersed in the isotonic and hypotonic solutions, which were induced from clamped potentials from a hold potential of –40 mV to test potentials from –100 to +100 mV with an increment of 20 mV for a duration of 0·3 s.
The transient inward current was induced by clamped potentials from –40 to +40 mV for a duration of 3 s and then repolarized to –40 mV. The amplitude of the transient inward current was measured as the difference between the peak of the transient current and the mean of the current just before and after the transient current [20].
Statistical analysis
Continuous variables are expressed as the mean ± S.E.M.
The differences among the atrial cardiomyocytes and PV cardiomyocytes with and without pacemaker activity were analyzed by a one-way anova. Multiple comparisons were analyzed with the Bonferroni t-test. The differences before and after the superfusion of the hypotonic solution were analyzed by a paired T-test. A P-value lower than 0·05 was considered to be statistically significant.
Results
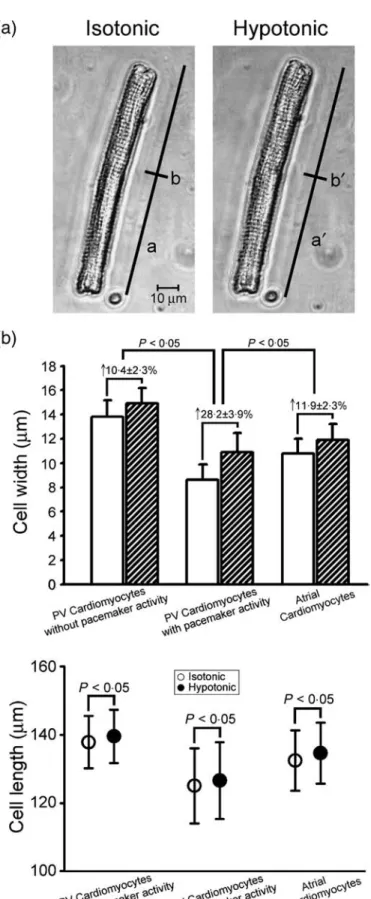
Effect of the hypotonic solution on the cellular size Compared to those observed in the isotonic solution,
immersion in the hypotonic solution increased the cell width and cell length in the PV and atrial cardiomyocytes (Fig. 1a). Moreover, hypotonic solution increased the cell width in the PV cardiomyocytes with pacemaker activity to a greater extent than those in the atrial cardiomyocytes or PV cardiomyocytes without pacemaker activity (Fig. 1b). However, hypotonic solution increased the cell length to a similar extent among the atrial cardiomyocytes and PV cardiomyocytes with or without pacemaker activity (Fig. 1c).
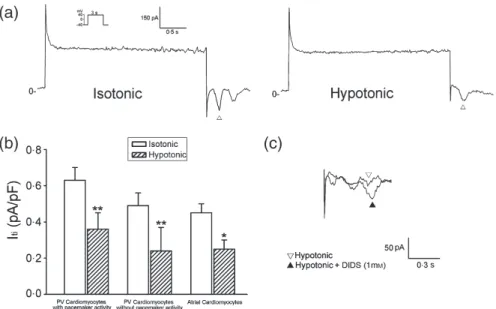
ICl,swell of the PV and atrial cardiomyocytes Figure 2a shows the recordings of the ICl,swell in the atrial cardiomyocytes and PV cardiomyocytes with and without pacemaker activity. The current densities of the ICl,swell differed among the atrial cardiomyocytes and PV cardiomyocytes with or without pacemaker activity. Figure 2b shows the I-V relationship of the ICl,swell in the PV and atrial cardiomyocytes. The ICl,swell in the PV cardiomyocytes were larger than those in the atrial cardiomyocytes. In addition, the PV cardiomyocytes with pacemaker activity had larger ICl,swell than those without pacemaker activity. The administration of 1 mm of ICl,swell blocker, DIDS (4,4′-diisothiocyanostilbene-2,2′-disulfonic acid) [24], reduced the swelling-activated outward currents. Those findings confirmed that these currents belonged to the ICl,swell (Fig. 2c).
Effects of hypotonic solution on the PV electrical activity
As in the example shown in Fig. 3a, the exposure of the cells to the hypotonic solution decreased the PV firing rates from 2·2 ± 0·2 to 1·1 ± 0·3 Hz (n = 9, P < 0·05) in the PV cardiomyocytes with pacemaker activity. In the PV cardiomyocytes without pacemaker activity, hypotonic solution shortened the APD90 and depolarized the resting membrane potential (Fig. 3b). In the atrial
cardiomyocytes, the hypotonic solution also decreased the APD90.
Figure 1 The change in the cell size occurring between isotonic and hypotonic solutions. Panel a shows the cell images from a pulmonary vein (PV) cardiomyocyte in an isotonic and hypotonic solution superfusion. The a, b (88·8, 13·7μm) and a′, b′ (90·6, 14·8μm) show the cell length and width of a PV cardiomyocyte in the isotonic and hypotonic solutions. Panel b shows the average cell length and width in atrial (n = 14) and PV cardiomyocytes with (n = 9) and without (n = 12) pacemaker activity.
Those effects could be reversed by the administration of DIDS (1 mm). However, compared with those in the atrial
cardiomyocytes, hypotonic solution induced a significantly greater APD90 shortening in the PV cardiomyocytes with a constant speed to reach a steady state (Fig. 3b,c). In contrast, hypotonic solution shortened the APD90 with a relatively slow response in atrial cardiomyocytes (Fig. 3c). Moreover, in the PV cardiomyocytes with delayed after depolarizations, hypotonic solution significantly decreased the amplitude of the DADs from 10·0 ± 1·7 to 1·6 ± 0·7 mV (n = 8, P < 0·01, Fig. 3d).
Effects of the hypotonic solution on the transient inward currents
In order to evaluate the mechanism of hypotonic solution-induced decrease of delayed after depolarization, we compared the transient inward currents of cardiomyocytes during immersion in isotonic and hypotonic solutions. Hypotonic solution decreased the transient inward currents both in the atrial cardiomyocytes and PV cardiomyocytes (Fig. 4). Those effects could be reversed by the administration of DIDS (1 mm, Fig. 4c). Therefore, the activation of ICl,swell may reduce the transient inward currents, decreasing hypotonic solution-induced delayed after
depolarizations.
Discussion
In this study, for the first time, we identified the existence of the ICl,swell in the PV cardiomyocytes and found different current densities of the ICl,swell between the PV and atrial cardiomyocytes.
Those results confirmed that the PV cardiomyocytes contained distinct electrophysiological characteristics [25]. Similarly, ICl,swell in atrial and sinoatrial node cells also differed [26]. The results of this study showed that the changes in the cell width during immersion in hypotonic solution were greater in the PV cardiomyocytes than in the atrial cardiomyocytes. These findings were hypothesized to arise from different cytoskeletons between the atrial cardiomyocytes and PV cardiomyocytes [27], since the PV cardiomyocytes have to adapt themselves to the vascular structure and they should have a more flexure structure with a susceptibility to volume changes. In addition, the significant increase in the cell width in the PV cardiomyocytes may cause larger ICl,swell. The PV and atrial cardiomyocytes are known to have different embryological origins and may have different cell structures [18]. Taken together, those findings suggested that non-uniform ICl,swell existed in the cardiomyocytes.
In this study, we found that osmotic swelling induced a greater APD90 shortening in the PV cardiomyocytes as compared to those Figure 2 The example of ICl,swell tracings (a) and the I-V relationship (b) during osmotic swelling in the atrial cardiomyocytes (n = 7) and PV cardiomyocytes with (n = 7) or without (n = 7) pacemaker activity. *P < 0·05 vs. atrial cardiomyocytes, #P < 0·05 vs. PV cardiomyocytes without pacemaker activity. The insets in the current traces show the various clamp protocols. (c) shows that ICl,swell blocker (DIDS, 1 mM) reduced hypotonic solution-induced ICl,swell in a PV cardiomyocyte.
in the atrial cardiomyocytes. Activation of the ICl,swell is known to shorten the AP duration in cardiomyocytes [10]. The larger ICl,swell in the PV cardiomyocytes will produce greater APD90 shortening with a rapid time course during immersion in hypotonic solution.
As micro re-entry plays a critical role in the PV arrhythmogenesis, the significant hypotonic solution-induced APD90 shortening will facilitate the genesis of micro re-entry circuits in the PVs and enhance the PV arrhythmogenesis to induce atrial fibrillation during myocardial ischaemia. Moreover, osmotic swelling may cause a decrease in conduction velocity due to a slight
depolarization of diastolic potential with inactivation of the Na+ current [28]. This effect may also enhance the occurrence of micro
re-entry. Since human and rabbit PVs have similar
electropharmacological characteristics [25], our findings should be comparable with the clinical situations.
ICl,swell has been found in pacemaker cells; however, ICl,swell and cell swelling in the pacemaker activity of cardiomyocytes has been obscure. The results of this study first showed that cell swelling may reduce the PV firing rates. Transient inward currents are known to induce delayed after depolarizations and play an important role in the PV firing rates. In this study, activation of ICl,swell would decrease the transient inward currents and reduce the PV firing rates or delayed after depolarizations. Those effects may be caused by ICl,swell-induced APD90 shortening, which would Figure 3 Effects of hypotonic solution on the PV electrical activity and action potential (AP) morphology. (a) shows an example of hypotonic solution reducing the PV firing rates from 1·8 Hz to 0·8 Hz and also inhibiting the delayed after depolarizations (↓).
Right panel shows the time course of the osmotic swelling on the PV firing rates. (b) shows the superimposed tracings and the average resting membrane potential (RMP) and AP duration at 90% of the full repolarization (APD90) in the PV (n = 8) and atrial cardiomyocytes (n = 8) during isotonic and hypotonic solutions. Hypotonic solution-induced a larger decrease in the APD90 in the PV cardiomyocytes than in the atrial cardiomyocytes. (c) shows the time course of hypotonic solution on the APD90 in a PV cardiomyocyte and atrial cardiomyocyte, respectively. (d) shows an example of the hypotonic solution reducing the amplitude of the delayed after depolarization (↓) in a PV cardiomyocyte. The APs were elicited by electrical stimuli delivered at 2Hz.
**P < 0·01, and ***P < 0·005 vs. the isotonic solution.
reduce the calcium entrance and decrease the intracellular calcium to decrease Na+/Ca2+ activity and transient inward currents. An opposite effect of the arrhythmogenesis of the APD90 shortening, reducing the PV firing rates and delayed after depolarizations by ICl,swell, may decease the PV arrhythmogenic activity.
Taken together, ICl,swell have complex effects on the PV arrhythmogenesis.
Heart failure is an important risk factor of atrial fibrillation and produces haemodynamic perturbations that cause cell swelling [1].
However, this study showed that cell swelling will reduce the PV firings rates and delayed after depolarizations. Those effects differed from the known effects of stretch on the PV
cardiomyocytes [29], whereas stretch will increase PV firing rates and triggered activity. It also has been shown that stretch and swelling have different electrophysiological effects. Stretch may generate early and delayed after depolarizations [30]. In contrast, similar to that in this study, cell swelling reduces the triggered activity [28].
Conclusions
This study found different hypotonic effects on the PV and atrial cardiomyocytes. The significant ICl,swell in the PV cardiomyocytes suggests that ICl,swell play a crucial role in the PV electrical activity and arrhythmogenesis.
Address
*Taipei Medical University, Graduate Institute of Clinical Medicine (Y. J. Chen); †Division of Cardiovascular Medicine, Taipei Medical University-Wan Fang Hospital (S. Y. Chen);
‡National Yang-Ming University, School of Medicine; Division of Cardiology and Cardiovascular Research Center, Veterans General Hospital-Taipei, Taipei (S. A. Chen); §Department of Biomedical Engineering and Institute of Physiology, National Defense Medical Center (Y. C. Chen, C. I. Lin); ¶Shin Kong Wu Ho-Su Memorial Hospital and Department of Medicine, Fu Jen Catholic University (S. H. Lee).
Correspondence to: Yi-Jen Chen, MD, PhD, Graduate Institute of Clinical Medicine, Taipei Medical University 250 Wu-Xin Street, Taipei, Taiwan. Tel.: 886-2-28757156;
fax: 886-2-29339378; 886-2-28735656;
e-mail: [email protected]
Received 2 April 2007; accepted 20 September 2007
Acknowledgements
The present work was supported by the Topnotch Stroke Research Centre Grant, Ministry of Education and grants NSC 95-2314-B-010-017, NSC 95-2314-B-010-025, NSC 95-2314-B-016-015, NSC 95-2314-B-038-026, V95C1-059, V95A-008, SKH-8302-96-DR-01 and SKH-TMU-94-01 from Shih Kong Wu Ho-Su Memorial Hospital.
Figure 4 Effects of hypotonic solution on the transient inward currents. (a) shows that hypotonic solution reduces the transient inward currents in a PV cardiomyocyte with pacemaker activity. (b) shows the average transient inward currents of the atrial cardiomyocytes (n = 7) and PV cardiomyocytes with (n = 10) or without (n = 7) pacemaker activity during the isotonic and hypotonic solutions. *P < 0·05 and **P < 0·01 vs. the isotonic solution superfusion. (c) shows the administration of DIDS (1 mM) reversing the hypotonic solution-induced decrease in the transient inward currents. The inset in the current traces shows the clamp protocol.
References
1 Kannel WB, Abbott RD, Savage DD, McNamare PM. Epidemiologic features of chronic atrial fibrillation. N Engl J Med 1982;306:1018–22.
2 Wolf PA, Abbott RD, Kannel WB. Atrial fibrillation as an independent risk factor for stroke. The Framingham Study. Stroke 1991;22:983–8.
3 Allessie MA, Boyden PA, Camm AJ, Kleber AG, Lab MJ, Legato MJ et al. Pathophysiology and prevention of atrial fibrillation. Circulation 2001;103:769–77.
4 Sinno H, Derakhchan K, Libersan D, Merhi Y, Leung TK, Nattel S.
Atrial ischemia promotes atrial fibrillation in dogs. Circulation 2003;107:1930–6.
5 Jennings RB, Ganote CE. Structural changes in myocardium during acute ischemia. Circ Res 1974;34:III156–72.
6 Tseng GN. Cell swelling increases membrane conductance of canine cardiac cells: evidence for a volume-sensitive Cl channel. Am J Physiol 1992;262:C1056–67.
7 Sorota S. Swelling-induced chloride-sensitive current in dog atrial cells revealed by whole cell patch clamp method. Circ Res 1992;70:679–87.
8 Hagiwara N, Masuda H, Shoda M, Irisawa H. Stretch-activated anion currents of rabbit cardiac myocytes. J Physiol (Lond) 1992;
456:285–302.
9 Du XY, Sorota S. Cardiac swelling-induced chloride current depolarizes canine atrial myocytes. Am J Physiol 1997;272:H1904–16.
10 Vandenberg JI, Bett GC, Powell T. Contribution of a
swelling-activated chloride current to changes in the cardiac action potential. Am J Physiol 1997;273:C541–7.
11 Walsh EP, Saul JP, Hulse E, Hulse JE, Rhodes LA, Hordof AJ et al.
Transcatheter ablation of ectopic atrial tachycardia in young patients using radiofrequency current. Circulation 1992;86:1138–46.
12 Haissaguerre M, Jais P, Shah DC, Takahashi A, Hocini M, Quiniou G et al. Spontaneous initiation of atrial fibrillation by ectopic beats originating in the pulmonary veins. N Engl J Med 1998;
339:659–66.
13 Chen SA, Hsieh MH, Tai CT, Tsai CF, Prakash VS, Yu WC et al.
Initiation of atrial fibrillation by ectopic beats originating from the pulmonary veins: Electrophysiological characteristics,
pharmacological responses, and effects of radio frequency ablation.
Circulation 1999;100:1879–86.
14 Pappone C, Rosanio S, Oreto G, Tocchi M, Gugliotta F, Vicedomini G et al. Circumferential radio frequency ablation of pulmonary vein ostia: a new anatomic approach for curing atrial fibrillation.
Circulation 2000;102:2619–28.
15 Sueda T, Imai K, Ishii O, Orihashi K, Watari M, Okada K. Efficacy of pulmonary vein isolation for the elimination of chronic atrial fibrillation in cardiac valvular surgery. Ann Thorac Surg 2001;
71:1189–93.
16 Nathan H, Eliakim M. The junction between the left atrium and the pulmonary veins: an anatomic study of human hearts. Circulation 1966;34:412–22.
17 Saito T, Waki K, Becker AE. Left atrial myocardial extension onto pulmonary veins in humans: anatomic observations relevant for atrial arrhythmias. J Cardiovasc Electrophysiol 2000;11:888–94.
18 Blom NA, Gittenberger-de Groot AC, DeRuiter MC, Poelmann RE, Mentink MM, Ottenkamp J. Development of the cardiac conduction tissue in human embryos using HNK-1 antigen expression: possible relevance for understanding of abnormal atrial automaticity.
Circulation 1999;99:800–6.
19 Cheung DW. Electrical activity of the pulmonary vein and its interaction with the right atrium in the guinea-pig. J Physiol (Lond) 1981;314:445–6.
20 Chen YJ, Chen SA, Chen YC, Yeh HI, Chan P, Chang MS et al. Effects of rapid atrial pacing on the arrhythmogenic activity of single cardiomyocytes from pulmonary veins: implication in initiation of atrial fibrillation. Circulation 2001;104:2849–54.
21 Vandenberg JI, Yoshida A, Kirk K, Powell T. Swelling-activated and isoprenaline-activated chloride currents in guinea pig cardiac myocytes have distinct electrophysiology and pharmacology.
J General Physiol 1994;104:997–1017.
22 Chen YC, Chen SA, Chen YJ, Chang MS, Chan P, Lin CI. Effects of thyroid hormone on the arrhythmogenic activity of pulmonary vein cardiomyocytes. J Am Coll Cardiol 2002;39:366–72.
23 Wongcharoen W, Chen YC, Chen YJ, Chang CM, Yeh HI, Lin CI et al. Effects of a Na+/Ca2+ exchanger inhibitor on pulmonary vein electrical activity and ouabain-induced arrhythmogenicity.
Cardiovasc Res 2006;70:497–508.
24 Sorota S. Insights into the structure, distribution and function of the cardiac chloride channels. Cardiovasc Res 1999;42:361–76.
25 Chen YJ, Chen SA. Electrophysiology of pulmonary veins.
J Cardiovasc Electrophysiol 2006;17:220–4.
26 Hagiwara N, Masuda H, Shoda M, Irisawa H. Stretch-activated anion currents of rabbit cardiac myocytes. J Physiol 1992;456:285–302.
27 Henson JH. Relationships between the actin cytoskeleton and cell volume regulation. Microsc Res Tech 1999;47:155–62.
28 Hiraoka M, Kawano S, Hirano Y, Furukawa T. Role of cardiac chloride currents in changes in action potential characteristics and arrhythmias. Cardiovasc Res 1998;40:23–33.
29 Chang SL, Chen YC, Chen YJ, Wangcharoen W, Lee SH, Lin CI et al.
Mechanoelectrical feedback regulates the arrhythmogenic activity of pulmonary veins. Heart 2007;93:82–8.
30 Tavi P, Laine M, Weckstrom M. Effect of gadolinium on
stretch-induced changes in contraction and intracellularly recorded action- and afterpotentials of rat isolated atrium. Br J Pharmacol 1996;118:407–13.