Tyrosine phosphorylation controls PCNA function through protein stability
Shao-Chun Wang1,8, Yusuke Nakajima1,2,8, Yung-Luen Yu1,8, Weiya Xia1, Chun-Te Chen1,3, Cheng-Chieh Yang1,3, Eric W. McIntush4, Long-Yuan Li1,5, David H. Hawke6, Ryuji Kobayashi6 & Mien-Chie Hung1,3,7
1. Department of Molecular and Cellular Oncology, The University of Texas M.D. Anderson Cancer Center, Houston, TX 77030, USA.
2. Current address: Oral and Maxillofacial Surgery, Department of Oral Restitution, Division of Oral Health Sciences, Graduate School, Tokyo Medical and Dental University, 1-5-45 Yushima, Bunkyo- Ku, Tokyo 113-8549, Japan.
3. Graduate School of Biomedical Sciences, The University of Texas M.D. Anderson Cancer Center, Houston, TX 77030, USA.
4. Bethyl Laboratories, Inc., 25043 W. FM1097, Montgomery, TX 77356, USA.
5. Current address: Center for Molecular Medicine, China Medical University Hospital, No. 2 Yuh-Der Road, Taichung 404, Taiwan.
6. Department of Molecular Pathology, The University of Texas M.D. Anderson Cancer Center, Houston, TX 77030, USA.
7. Center for Molecular Medicine, China Medical University Hospital, Taichung 404, Taiwan.
8. These authors contributed equally to the work.
Correspondence to: Mien-Chie Hung1,3,7 e-mail: [email protected]
Abstract
The proliferating cell nuclear antigen (PCNA) is an essential protein for DNA replication and damage repair. How its function is controlled remains an important question. Here, we show that the chromatin-bound PCNA protein is phosphorylated on Tyr 211, which is required for maintaining its function on chromatin and is dependent on the tyrosine kinase activity of EGF receptor (EGFR) in the nucleus. Phosphorylation on Tyr 211 by EGFR stabilizes chromatin- bound PCNA protein and associated functions. Consistently, increased PCNA Tyr 211 phosphorylation coincides with pronounced cell proliferation, and is better correlated with poor survival of breast cancer patients, as well as nuclear EGFR in tumours, than is the total PCNA level. These results identify a novel nuclear mechanism linking tyrosine kinase receptor function with the regulation of the PCNA sliding clamp.
The proliferating cell nuclear antigen (PCNA) protein assembles in a homo-trimeric ring structure, encircling the DNA double helix, and functions as a mobile sliding clamp to recruit other proteins (such as DNA polymerases and ligases) to chromatin1. Functionally, nuclear PCNA protein is present in two populations: the detergent-resistant chromatin-bound form, which is involved in DNA replication and repair (Triton-resistant); and the chromatin- unbound form, which is not involved in DNA synthesis and can be readily released by detergent extraction (Triton-extractable)2. The chromatin-bound PCNA coordinates DNA replication and high polymerase processivity, and is also involved in multiple mechanisms of DNA-damage repair, including mismatch repair (MMR)3. These repair mechanisms also require DNA synthesis, and PCNA is the key molecule that controls the switch between processive and translesional DNA synthesis4, 5 . Consequently, preventing PCNA from loading onto the chromatin is a lethal deficiency for cells6. As well as DNA synthesis, recent findings highlight that PCNA modulates other critical cellular functions, including cell-cycle control, chromatin remodelling and gene expression, through a broad range of binding partners3, 7 . Despite the vital and versatile biological functions of PCNA, the mechanisms that regulate its activity in the nucleus remain to be explored.
EGF receptor (EGFR) is a tyrosine kinase receptor that has crucial roles in cell proliferation and tissue differentiation8. Although EGFR has conventionally been considered to be a membrane protein, it has also been detected in the nucleus and associated with chromatin9, 10,
11 . However, whether the tyrosine kinase activity of EGFR functions in the nucleus has not yet been determined, and if it does function in the nucleus, the targets of the kinase also remain to be determined. In the hepatocytes of regenerating liver, increased nuclear expression of EGFR is closely associated with active cell proliferation, as well as elevated expression of PCNA protein12, 13 . In cancer, nuclear EGFR expression is frequently associated with multiple tumour types14, 15, 16, 17 , including breast cancer9, 17 . In this regard, nuclear EGFR has been shown to be a poor prognosis tumour marker for cancer patients14, 17 . Interestingly, it is also well documented that EGFR expression in tumour tissues is correlated with the expression of PCNA by immunohistochemical staining, especially in the more advanced stages of disease18, 19 . Indeed, co-expression of EGFR and PCNA in breast tumours is associated with decreased responsiveness to primary chemotherapy20. Nevertheless, the underlying causes for the correlation between EGFR and PCNA have been elusive.
In the current study, we demonstrate that PCNA is subject to tyrosine phosphorylation at a specific site and that the phosphorylation enhances PCNA stability on the chromatin. Our results link the tyrosine kinase activity of nuclear EGFR to cell proliferation and DNA repair by regulating PCNA function.
Top of page
Results
Tyr 211 phosphorylation of PCNA
To gain further insights into the post-translational regulation of PCNA function, endogenous PCNA protein of the human cancer cell line A431 was purified by immunoprecipitation with a monoclonal PCNA antibody, followed by SDS gel electrophoresis. Electrospray ionization tandem mass (MS-MS) spectrometry analysis reproducibly identified a specific peptide containing phosphorylated Tyr 211 PCNA, indicating that Tyr 211 is phosphorylated under physiological conditions (see Supplementary Information, Fig. S1a). This tyrosine residue is conserved across different species (see Supplementary Information, Fig. S1a, inset). Indeed, phosphorylation at Tyr 211 was also detected in mouse cells (see Supplementary Information, Fig. S1b), indicating that Tyr 211 phosphorylation of PCNA is an evolutionarily conserved event. Consistently, tyrosine-phosphorylated PCNA was detected by western blotting using an anti-phosphotyrosine antibody (4G10) after immunoprecipitation by an anti-PCNA antibody (Fig. 1, DMSO lane).
Figure 1: Tyrosine phosphorylation of PCNA.
MDA-MB-468 cells were treated with DMSO, AG1478, U0126, LY294002, or a combination of these inhibitors (mix). Upper panel, endogenous PCNA protein was immunoprecipitated and the associated epidermal growth factor receptor (EGFR), and tyrosine phosphorylation of EGFR and PCNA (marked with an arrowhead) were examined with phosphotyrosine antibody 4G10. The amount of the immunoprecipitated PCNA was normalized to the amount of the corresponding immunoglobulin G heavy chain, and the relative level of each treatment was then determined using the PCNA level in the DMSO-treated sample (set as 1). IP, immunoprecipitation. Lower panel, western blots for treatment effectiveness.
Tyr 211 phosphorylation and protein stability of PCNA
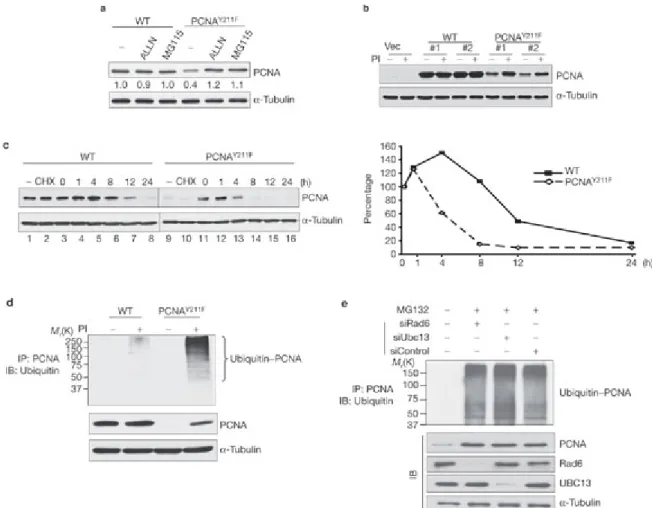
To explore the biological effects of Tyr 211 phosphorylation, a tyrosine-to-phenylalanine mutant of PCNA (PCNAY211F) was generated and both the wild-type and PCNAY211F were expressed in 293T cells by transient transfection. Western blot analysis showed that PCNAY211F was expressed at a moderately lower level compared with the wild type, whereas treatment with the proteasome inhibitor MG115 or ALLN recovered the protein level (Fig. 2a), indicating that Tyr 211 phosphorylation of PCNA modulates its stability. To further explore this possibility, stable transfectants expressing Flag-tagged wild-type or PCNAY211F were generated in the breast cancer cell line MDA-MB-468. Consistent with the transient transfection system, the protein level of mutant PCNA was about five times less than that of the wild type (Fig. 2b) and could be rescued by treatment with proteasome inhibitors. This observation was made both in independent stable clones (Fig. 2b) and in pooled stable transfectants (data not shown), excluding the possibility of clonal heterogeneity. Indeed, the estimated half-life of the wild-type PCNA protein (12 h) is significantly longer than that of PCNAY211F (5 h; Fig. 2c). These results indicate that cellular PCNA is subject to proteasome-dependent degradation and that phosphorylation on Tyr 211 prevents PCNA degradation. In line with these results, proteasome inhibitors caused more pronounced accumulation of poly-ubiquitinated PCNAY211F than the wild-type PCNA (Fig. 2d). PCNA is known to be ubiquitinated by the RAD6–RAD18 and UBC13–MMS2–RAD5 complexes under DNA-damage stress4, 5, 21, 22, 23 . To test whether the same mechanism also mediates the poly-ubiquitination that is associated with PCNAY211F, expression of the key complex components, RAD6 and UBC13, were knocked down by specific small interference RNA (siRNA). The results showed that downregulation of these genes did not affect the level of poly-ubiquitinated PCNAY211F (Fig. 2e), indicating that the protein stability of PCNA is controlled by a signalling pathway that is independent of the ubiquitination system induced by the RAD6–RAD18 and UBC13–MMS2–RAD5 complexes under DNA-damage stress.
Figure 2: Y 211 phosphorylation is required for maintaining PCNA protein stability.
(a) Ectopic expression of wild-type (WT) or Y211F Flag–PCNA in 293T cells with or without the proteasome inhibitors (PIs) ALLN and MG115 (5 M; 24 h). –, no treatment. Three micrograms of protein were loaded in each lane. (b) The expression levels of Flag–PCNA- WT and the Flag–PCNAY211F mutant in stable clones of MDA-MB-468 cells were examined. Vec, the pcDNA3 vector control line; PI, a mix of MG115, MG132 and ALLN (5 M each). (c) Left panel, the stable clones of PCNA-WT and PCNAY211F were either mock- treated (no treatment; lanes 1, 9), or treated by cycloheximide (50 M) (CHX; lanes 2, 10) or cycloheximide in the presence of the proteasome inhibitors as described in b (lanes 3–8 and 11–16). The level of Flag–PCNA protein at each time point was plotted (right panel). The levels of PCNA expression at time point 0 (lanes 3 and 11) were set at 100%. (d) Stable clones of Flag-tagged wild-type or PCNAY211F were treated with or without the PIs, as described in b. PCNA was then immunoprecipitated by an anti-Flag antibody and blotted with an anti-ubiquitin antibody. K, relative molecular mass in thousands; IB, immunoblotting; IP, immunoprecipitation. (e) A stable clone of Flag-tagged PCNAY211F was treated as indicated, and the poly-ubiquitinated PCNA was examined as in d. The levels of the indicated
proteins are also shown.
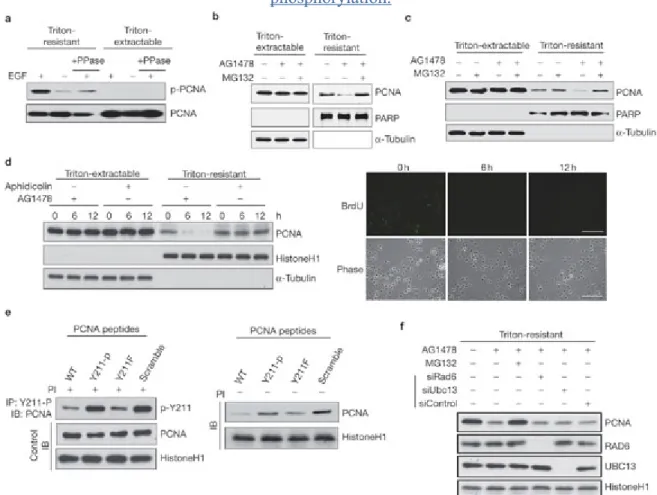
Interestingly, tyrosine phosphorylation of PCNA can be completely blocked by the EGFR- specific inhibitor AG1478 but not by the inhibitors of other kinases, including MAP kinase (UO126) and PI-3-kinase (PI(3)K; LY294002) (Fig. 1 and see below for further discussion), indicating that EGFR may have a major role in PCNA phosphorylation. Indeed, EGF preferentially stimulated tyrosine phosphorylation of Triton-resistant, chromatin-bound PCNA, but not the Triton-extractable form of the protein. The phosphorylation was further confirmed by treatment with phosphatase (Fig. 4a). These results demonstrate specific regulation of chromatin-bound PCNA by EGFR. The specificity of this regulation was also reflected at the level of protein stability, and the chromatin-bound and -unbound forms of PCNA protein responded differently to EGFR inhibition (Fig. 4b, c, and see Supplementary Information, Fig. S1c, d). In MDA-MB-468 cells, the protein level of chromatin-bound PCNA was decreased following inhibition of EGFR kinase activity for 16 h, whereas the level of the Triton-extractable, chromatin-unbound PCNA remained unchanged (Fig. 4b).
Treatment with proteasome inhibitors rescued chromatin-bound PCNA levels (Fig. 4b, c, f). A similar observation was made in another breast cancer cell line, MDA-MB-231, which expresses moderate levels of EGFR (see Supplementary Information, Fig. S1d), and in EGFR-expressing fibroblast cells (Fig. 4c)24. Thus, EGFR tyrosine kinase activity regulates the protein stability of chromatin-bound, rather than -unbound, PCNA. On the other hand, treatment with the DNA polymerase inhibitor aphidicolin suppressed DNA synthesis, but did not change PCNA protein levels (Fig. 4d), indicating that destabilization of PCNA by EGFR inhibition is not due to the suppression of DNA replication.
Figure 3: Nuclear EGFR associates and phosphorylates PCNA at tyrosine 211.
(a) Cell lysates were subject to immunoprecipitation (IP) and immunoblotting (IB) with the indicated antibodies. (b) Nuclear lysate of MDA-MB-468 cells was subject to immunoprecipitation and immunoblotting with the indicated antibodies. (c) A431 cells were serum-starved for 24 h, followed by stimulation by epidermal growth factor (EGF) for different times as indicated, and then lysed by NETN buffer. Endogenous PCNA was then immunoprecipitated by an anti-PCNA antibody, and the extent of tyrosine phosphorylation of PCNA (p-PCNA) was examined by the phosphotyrosine antibody PY100. The associated total EGFR and the phosphorylated EGFR (p-EGFR, as detected by the antibody 4G10) on the same western blot are also shown. (d) Confocal immunofluorescence co-localization of PCNA (red) and EGFR (green) in MDA-MB-468 cells. Cells were starved in the presence of 0.1% foetal bovine serum for 36 h, then treated with or without EGF (20 ng ml-1) for 15 min.
The co-localization of PCNA and EGFR is shown in the merged image (yellow). Scale bar, 5 m. (e) Top, autoradiograph of GST–PCNA and GST–PCNAY211F phosphorylation by the recombinant EGFR. Middle, autophosphorylation of EGFR. Bottom, Coommassie blue staining of the same gel. K, relative molecular mass in thousands. (f) The synthetic peptides as used in Figure 4e were subjected to a phosphorylation reaction by the recombinant EGFR kinase, as detected by the antibody 4G10. Ponceau staining of the peptides is shown for the loading control. Scramble, a sequence scrambled peptide. (g) MDA-MB-468 cells were serum-starved for 36 h, then treated with or without EGF for 15 min. Cells were then extracted with Triton-X-100 and the endogenous level of phospho-PCNA was determined by immunoprecipitation using the anti-phospho-Y211 antibody, followed by western blotting analysis using the PCNA antibody. HC, heavy chain. (h) As in g, except that cells were grown in normal medium and treated with or without AG1478, as described in Figure 4b.
Figure 4: Protein stability of the chromatin-associated PCNA is controlled by Y 211 phosphorylation.
(a) MDA-MB-468 cells were serum-starved for 24 h, then treated with EGF for 15 min.
Endogenous PCNA in the Triton-extractable and Triton-resistant fractions was immunoprecipitated, then incubated with or without lambda phosphatase (PPase), then blotted with the antibody 4G10. (b) MDA-MB-468 cells were treated with or without AG1478 (10
M for 16 h in the presence of serum) and MG132 (5 M), then fractionated with a Triton-X- 100-containing buffer. (c) The same experiment was performed for the NIH3T3-derived EGFR stable transfectant cell line, HER-5. (d) Left panel, MDA-MB-468 cells were treated as indicated, and the levels of PCNA at each time points were examined. Right panel, the effect of aphidicolin treatment on DNA replication was confirmed by BrdU incorporation (anti- BrdU antibody, green). Scale bar, 200 m. (e) Left panel, MDA-MB-468 cells were incubated with the indicated peptide in culture medium for 12 h, followed by Triton extraction.
Phospho-Tyr 211 PCNA in the chromatin-bound pool was immunoprecipitated and examined by the antibodies indicated. PI, proteasome inhibitor. Right panel, the chromatin-bound PCNA protein levels in the presence of indicated peptides were determined by immunoblotting. Scramble, the peptide with the same composite amino acids of the wild-type peptide but in a scrambled sequence. (f) MDA-MB-468 cells were treated as indicated and the levels of indicated proteins were determined by immunoblotting.
Full size image (42 KB)
To directly test the role of Y 211 phosphorylation on PCNA protein stability, peptides with either tyrosine, phenylalanine (Y211F) or phosphorylated tyrosine (Y211-p) in place of the residue were conjugated with a HIV-1 Tat-derived protein and transfected into cells. The Tat sequence is known to facilitate nuclear targeting of peptides25. We reasoned that the wild- type PCNA and PCNAY211F would be able to compete against PCNA protein for the kinase responsible for Y211 phosphorylation in the nucleus and, therefore, would inhibit the level of phospho-Y211 PCNA. On the other hand, the Y211-p peptide would not be as capable in competing for the kinase because it mimicks a phosphorylated product for the kinase. To examine the phosphorylation status of Y 211, an antibody recognizing the phosphorylated Y 211 was generated (see Supplementary Information, Fig. 1e). This antibody mainly recognizes the natural form of its cognate epitope, as it is able to immunoprecipitate the phosphorylated PCNA but is incapable of detecting the denatured protein by western blotting (data not shown). As expected, the wild-type and the Y211F peptides, but not the Y211-p peptide nor the peptide with a scrambled sequence, suppressed PCNA Y 211 phosphorylation (Fig. 4e, left panel) and downregulated the protein level of chromatin-bound PCNA (Fig. 4e, right panel), similar to the effect of AG1478 (Fig. 4b, c, d). As in the case of poly- ubiquitination shown in Figure 2e, downregulation of the chromatin-bound PCNA by AG1478 was not affected by knocking down the expression of RAD6 and UBC13, indicating a RAD6- and UBC13-independent mechanism (Fig. 4f).
Association and phosphorylation of PCNA by nuclear EGFR
Inhibition of EGFR abolished PCNA tyrosine phosphorylation, but not MAP kinase and PI(3)K (Fig. 1). As EGFR is capable of translocating to the nucleus9, we tested whether
EGFR phosphorylated PCNA in the nucleus. Indeed, interaction between endogenous EGFR and PCNA can be detected in different cell lines (Fig. 3a). The interaction between these two proteins was also demonstrated by reciprocal immunoprecipitation from lysates of 293T cells that had been cotransfected with PCNA and EGFR (see Supplementary Information, Fig. S2a, b) in a dose-dependent manner (see Supplementary Information, Fig. S2c). The interaction between EGFR and PCNA occurred in nuclear extracts (Fig. 3b). In different cell lines, EGF treatment quickly stimulated the interaction and increased tyrosine phosphorylation of PCNA (Fig. 3c and see Supplementary Information, Fig. S2d). Consistently, EGF treatment reproducibly stimulated nuclear colocalization of EGFR and PCNA, as observed by confocal immunofluorescence microscopy (Fig. 3d, and see Supplementary Information, Fig. S3), in which a single photoplane well within the nucleus was chosen to avoid the potential interference by cytoplasmic signals. Under these conditions, we did not observe nuclear localization of the membrane protein CD44, supporting the specificity of the EGFR nuclear staining (Fig. 3d). To further demonstrate the nuclear location of the EGFR–PCNA- containing complexes, sequential photosections of a nucleus were examined (see Supplementary Information, Fig. S4c).
AG1478 treatment suppressed PCNA tyrosine phosphorylation, as well as its association with EGFR (Fig. 1), indicating that PCNA may be a substrate of EGFR. Indeed, purified recombinant EGFR kinase domain phosphorylated the glutathione S-transferase (GST) fusion protein of the wild-type PCNA but not of PCNAY211F, indicating that EGFR phosphorylated Tyr 211 of PCNA (Fig. 3e). The EGFR kinase domain phosphorylated GST–PCNA in a dose- dependent manner, with the phosphate-to-substrate ratio plateau approaching one phosphate group per GST–PCNA molecule (see Supplementary Information, Fig. S2e).
Immunoprecipitated endogenous EGFR also exhibited the same kinase activity towards PCNA (data not shown). In addition, the wild-type peptide capable of blocking Tyr 211 phosphorylation of PCNA (Fig. 4e) was also a specific substrate of EGFR (Fig. 3f), further supporting the kinase-substrate relationship between PCNA and EGFR. Together, these results demonstrate that EGFR phosphorylates PCNA on Tyr 211. To further demonstrate this specific phosphorylation in vivo, cells were treated with or without EGF, and the chromatin- bound as well as the -unbound form of cellular PCNA was examined by the anti-phospho-Tyr 211 antibody. This experiment clearly demonstrates that EGF treatment specifically stimulated Tyr 211 phosphorylation of chromatin-bound form of PCNA (Fig. 3g).
Conversely, blocking EGFR activity diminished Tyr 211 phosphorylation of chromatin-bound PCNA (Fig. 3h). These results indicate that EGFR specifically targets the functional form of PCNA in the nucleus, stabilizing the PCNA protein that is associated with chromatin during cell proliferation (see also Fig. 5).
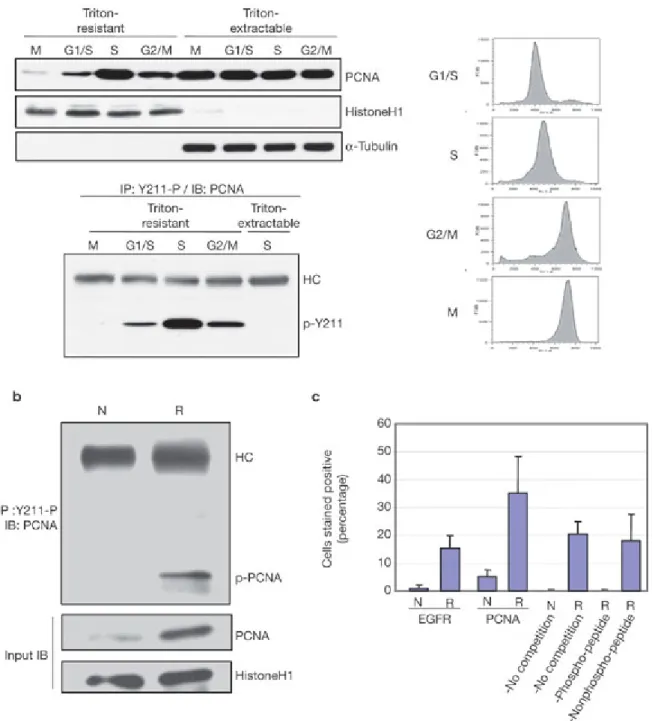
Figure 5: Tyr 211 phosphorylation of PCNA is associated with cellular and physiological proliferation.
(a) HeLa cells synchronized at M, G1-S, S and G2-M phases were extracted by Triton-X-100.
Endogenous PCNA levels (left, upper panel) and the corresponding phospho-Tyr 211 PCNA levels (left, lower panel) are shown. Right, the FACS plots for each cell-cycle stage. (b)
Nuclear phospho-Tyr 211 PCNA in regenerating liver. N, normal liver; R, regenerating liver 24 h after partial hepatectomy. The experiment was performed using male Fisher 344 rats using the procedure described previously13. (c) The percentage of liver cells with nuclear staining of EGFR and phospho-Tyr 211 PCNA in regenerating liver. Sequential tissue sections of the normal liver (N) and the regenerating liver 14 h after partial hepatectomy (R) were stained for EGFR, PCNA and phospho-Tyr 211 PCNA. Three different fields are scored.
Full size image (46 KB)
Tyr 211 phosphorylation and PCNA nuclear functions
The specific presence of phospho-Y211 in the chromatin-bound fraction indicates that Tyr 211 phosphorylation is associated with the genomic function of PCNA. To this end, the DNA- replication activity of the wild-type and PCNAY211F in the MDA-MB-468 stable transfectants (Fig. 2) was examined by measuring their DNA synthesis ability — for which the amount of PCNA-associated neo-synthesized genomic DNA was evaluated by chromatin immunoprecipitation (ChIP)26. Compared with wild-type PCNA, the chromatin immunocomplex that was derived from PCNAY211F contained significantly less neo- synthesized DNA (Fig. 6a, compare the wild-type and Y11F groups). The loss of PCNAY211F function can be attributed to its lower protein expression, as treatment by proteasome inhibitors recovered its protein level (Fig. 6b), and accordingly rescued the DNA synthesis activity (Fig. 6a, compare the WT+PI and the 211F+PI groups). In addition to DNA synthesis, the wild-type and PCNAY211F were also compared for MMR activity27. These two PCNA species were immunoprecipitated from cellular extracts by an anti-Flag antibody in the presence or absence of proteasome inhibitors. The purified PCNA was added into a DNA-repair mix and the associated MMR activity was measured, as described previously27.
The results showed that, although the mutant PCNA exhibited a much lower repair activity than the wild type (Fig. 6c, compare lanes 4 and 6), proteasome inhibition significantly rescued the repair function of mutant PCNA (Fig. 6c, compare lanes 8 and 10). These results also indicate that PCNAY211F still maintains intrinsic DNA synthesis and DNA MMR activities and it is the reduced protein expression of PCNAY211F that results in the decrease of these functions (Fig. 6a, c). Consistently, recombinant GST-wild-type and -PCNAY211F fusion proteins had the same binding ability to PCNA-associated partners critical for its function, such as FEN-1 and p21WAF1/CIP (Fig. 6d)1, 3, 7 . These GST fusion proteins exhibited identical DNA MMR activity when the same amounts of fusion protein were used in the assay (Fig. 6e). Thus, Tyr 211 phosphorylation is important for the physiological function of PCNA by regulating its protein stability.
Figure 6: Tyr 211 phosphorylation indirectly regulates the DNA repair and DNA replication functions of PCNA without changing its intrinsic activity.
(a) Stable clones of Flag-tagged PCNA or PCNAY211F PCNA were metabolically labeled by [3H]-thymidine and the associated neo-synthesized DNA was assessed by chromatin immunoprecipitation using an anti-Flag antibody or IgG. Asterisks, P < 0.05. (b) Immunoblotting analysis of wild-type and PCNAY211F expression. (c) Mismatch-repair assay using the stable clones. PBP, PCNA-binding p21 peptide. Statistical significance was determined by the Student's t-test from three independent experiments (P < 0.05). (d) Glutathione S-transferase (GST) pull-down assay using GST–PCNA-WT and GST–
PCNAY211F. (e) Mismatch-repair assay using GST–PCNA-WT and GST–PCNAY211F. A, mutant PBP without binding activity to PCNA; J, non-specific peptide; PBP, PCNA-binding p21 peptide; W, wild-type PBP. Inset, Coommassie Blue staining for GST, GST–WT and GST–Y211F fusion proteins. Statistical significance was determined by the Student's t-test from three independent experiments (P < 0.05).
As PCNA is involved in DNA synthesis and is known to be active in S phase, we examined the levels of phosho-Tyr 211 during cell-cycle progression. Although no change was found for the chromatin-unbound pool, the level of chromatin-bound PCNA is clearly regulated through the cell cycle, with a peak at S phase (Fig. 5a, upper panel), as was Tyr 211 phosphorylation (Fig. 5a, lower panel). Strikingly, the chromatin-unbound form of PCNA
contained no detectable Tyr 211 phosphorylation even at S phase, supporting the physiological function of Tyr 211 phosphorylation in DNA replication. The coordination of Tyr 211 phosphorylation with DNA synthesis implicates its critical role in tissue homeostasis, for which liver regeneration is a well-established physiological model. Liver tissue at resting state expresses very low levels of PCNA. After partial hepatectomy, robust DNA replication ensues within 24 h; increased PCNA protein expression occurs, which is a documented proliferation indicator during liver regeneration (Fig. 5b)12. The increase in PCNA level was strikingly associated with prominent Tyr 211 phosphorylation, indicating that increased PCNA protein stability by Tyr 211 phosphorylation is triggered in regenerating liver (Fig.
5b). It has been reported that EGFR is transported to the nuclei of regenerating liver cells13.
Indeed, partial hepatectomy triggered extensive nuclear localization of EGFR, as well as phospho-Tyr 211 PCNA (Fig. 5c, and see Supplementary Information, Fig. S4a, b).
Competition with the phospho-Tyr 211 peptide, but not the corresponding non-phospho peptide, abolished the nuclear immunoreactivity with the anti-phospho-Tyr 211 antibody, confirming the identity of phospho-Tyr 211 PCNA in the nucleus (Fig. 5c, and see Supplementary Information, Fig. S4b). The nuclear EGFR immunostaining was also verified by competition with the corresponding antigen peptide that was derived from EGFR, as reported previously (data not shown)17.
In addition to the hyper-proliferative liver tissues, expression of EGFR and phospho-Tyr 211 PCNA was also examined in consecutive-tissue sections that were derived from a cohort of human breast cancer tumour samples (see Supplementary Information, Fig. S4c, d, e).
Phospho-Tyr 211 PCNA was better correlated with the nuclear (P < 0.0001) than the membrane-cytoplasmic form of EGFR (P < 0.029). Total PCNA, on the other hand, revealed no significant correlation with cytoplasmic-membrane EGFR, and the correlation with nuclear EGFR was marginal (P > 0.067) (see Supplementary Information, Fig. S4f). It is noteworthy that, in the same cohort of tumour samples, the nuclear but not the total EGFR has been shown to be correlated with poor patient survival17. The distinct pathological characteristic of phospho-Tyr 211 PCNA versus total PCNA became more evident in that phospho-Tyr 211 PCNA expression was significantly correlated with poor overall survival (P < 0.0001), whereas total PCNA had no correlation with overall survival (P = 0.9564). On the other hand, phospho-PCNA showed no correlation with the expression of oestrogen receptor or progesterone receptor, or age (data not shown). Hence, phospho-Tyr 211 PCNA has prognostic value for cancer patients. Despite its wide application as a cancer marker, the correlation between PCNA expression and patient survival has been controversial. Studies either favouring its poor prognostic value for cancer patient survival28, 29 or challenging this correlation30, 31, 32, 33 have been reported. By showing that it is the level of the phospho- Tyr 211 PCNA rather than the total PCNA that bears a strong prognostic value in primary tumour tissues, our findings not only offer a resolution for the discrepancy but also provides a
significant biological relevance for phosphorylation of PCNA at Tyr 211.
Top of page
Discussion
It is well known that EGFR regulates multiple important physiological functions in cells8, 34, 35. EGFR9, and its family members ErbB-2 (Refs 36, 37), ErbB-3 (ref. 38) and ErbB-4 (ref.
39), have recently been demonstrated to translocate to the nucleus40, 41, 42, 43 , and a mechanism involving endocytosis and the nuclear-pore complex has been proposed44, 45 . The nuclear functions of these membrane receptors or their derivatives have been attributed to their ability to target gene promoters as transcriptional regulators9, 10, 11, 36, 37, 42, 43, 46 . It remains elusive whether these receptors also function as tyrosine kinases in the nucleus and, if they do, what their target substrates are in the nucleus. In the current study, we demonstrate that nuclear EGFR phosphorylates and, consequently, regulates the protein stability of chromatin-bound PCNA, which is the centerpiece of the nuclear machinery for genome replication. Phosphorylation at Tyr 211 stabilizes the functional form of PCNA during DNA synthesis, and is found at high levels in regenerating liver and in tumour tissues with high nuclear expression of EGFR, a molecular marker that is associated with poor prognosis14, 17 . PCNA has been widely used as a tumour marker, and this study shows that it is the distinct protein modification rather than the overall expression level of PCNA that determines the correlation with patient survival.
Our results show that Tyr 211 phosphorylation of PCNA occurs mainly in the chromatin- bound form, and we estimate that about 2% of PCNA is associated with nuclear EGFR, and a similar level of nuclear EGFR iss associated with PCNA (see Supplementary Information, Fig. S2f, g, h). It should be pointed out that the interaction between EGFR and PCNA may be subject to dynamic regulation, such as sub-nuclear localization and by participation of other nuclear factors in the complex; also, phosphorylation may not occur in a stoichiometric manner. Indeed, we estimated that about 6% of the chromatin-bound form of PCNA was phosphorylated on Tyr 211 by immunoprecipitation using the anti-phospho-Tyr 211 antibody (see Supplementary Information, Fig. S2g). Chromatin-associated PCNA is known to be subject to different modifications, including ubiquitination, sumoylation, and, as demonstrated here, phosphorylation. Ubiquitinated PCNA switches its associated DNA polymerase to engage in translesion synthesis of DNA. The complex nature of this regulation was demonstrated by showing that PCNA ubiquitination is reversible and that the ubiquitin- specific protease 1 negatively and specifically regulates PCNA ubiquitination47. Because Tyr 211 phosphorylation does not affect PCNA in the chromatin-unbound compartment and because phosphorylation is cell-cycle dependent, it is possible that PCNA Tyr 211 phosphorylation is reversible. Given the fact that the chromatin-bound PCNA is particularly vulnerable to proteasome-mediated degradation, it is likely that the degradation mechanism is
confined to the chromatin-associated compartment. After phosphorylation, stabilized PCNA could recruit different partners followed by de-phosphorylation. Alternatively, other more complicated mechanisms might exist.
The physiological importance of Tyr 211 phosphorylation in maintaining PCNA protein stability can be further supported by directly targeting Tyr 211 phosphorylation using synthetic peptides, which resulted in the destabilization of chromatin-bound PCNA.
Interestingly, although both the wild-type and PCNAY211F peptides successfully blocked Tyr 211 phosphorylation, the Tyr 211-p peptide did not. This is in line with our hypothesis that the Tyr 211-p peptide mimicks a phosphorylated product and therefore harbours a lower affinity to the cognate kinase. Consistently, the Tyr 211-p peptide exhibited poor binding to nuclear EGFR in vitro when compared with the wild-type and PCNAY211F peptides (see Supplementary Information, Fig. S2i). As a consequence, only weak co-localization of phospho-Tyr 211 PCNA and the nuclear EGFR was detected using the anti-phospho-Tyr 211 PCNA antibody (see Supplementary Information, Fig. S3b).
Our study shows that modification of PCNA by phosphorylation modulates its modification by poly-ubiquitination. It will be interesting to know whether the different post-translational modifications of PCNA constitute a functional code and modulate its activity temporally and spatially, for which the tyrosine kinase receptors in the nucleus may have an important role.
Top of page
Methods
Antibodies, chemicals and peptides.
The following antibodies and chemicals were purchased commercially: anti-EGFR (Santa Cruz, Santa Cruz, CA; NeoMarker, Freemont, CA); anti-PCNA, -histone H1 and -FEN1 (Bethyl, Montgomery, TX; Santa Cruz); anti-phosphotyrosine PY-100 (Cell Signalling, Beverly, MA); anti-phosphotyrosine 4G10 (Upstate, Lake Placid, NY); anti-PARP and -p21 (BD Biosciences, San Jose, CA); anti- -tubulin and -Flag (Sigma, St Louis, MO); anti- ubiquitin (Upstate); anti-CD44 (Santa Cruz); anti-BrdU (Abcam, Cambridge, MA); BrdU (Invitrogen, Carlsbad, CA); MG115, MG132, ALLN and AG1478 (CalBiochem, San DiegoCA); and cycloheximide and human recombinant EGF (Sigma). The lambda phosphatase was purchased from New England BioLabs (MA). The anti-phospho-Try 211 PCNA antibody was raised against a phosphorylated synthetic peptide and purified with the phosphopeptide column at Bethyl Laboratories, Inc. The synthetic peptides used to target Tyr 211 phosphorylation were derived from the antigen peptides for antibody induction by linking the amino-terminal end to the TAT-1 nuclear-localization sequence (GRKKRRQRRR)25, which were prepared and provided by Bethyl Laboratories, Inc. The siRNAs were purchased from Santa Cruz (control siRNA, sc-37007; Rad6 siRNA, sc-41677; Ubc13 siRNA, sc-
43551).
Cell extractions.
The cytoplasmic and nuclear fractions were prepared as described previously48. Briefly, cultured cells or liver tissues were lysed in buffer A (50 mM NaCl, 10 mM HEPES at pH 8, 500 mM sucrose, 1 mM EDTA, 0.5 mM Spermidine, 0.15 mM Spermine, 0.2% Triton-X- 100) on ice and dounced by a homogenizer. The nuclear pellet was washed and isolated. The nuclei were lysed in RIPA buffer (150 mM NaCl, 1% NP-40, 0.5% deoxycholate, 0.1% SDS, 50 mM Tris at pH 7.5, 25 mM NaF, 2 mM Na3VO4, 5 mM PMSF, 2 g ml-1 aprotinin) and by sonication (Sonics Vibra Cell, amplitude 20; Sonics & Materials, Newtown, CT). The fractionation efficiency was assessed by antibodies against -tubulin and PARP. Triton extraction for lysate was performed by incubating the cell monolayer in a 100-mm culture dish with 500 l of buffer A (100 mM NaCl, 300 mM sucrose, 3 mM MgCl2, 10 mM PIPES at pH 6.8, 1 mM EGTA, 0.2% Triton-X-100, 25 mM NaF, 2 mM Na3VO4, 5 mM PMSF, 2 g ml-1 aprotinin) for 5 min on ice. The cells were then collected by scraping and washed twice with the same buffer. The pellet was lysed in RIPA buffer. Protein expression in the lysates was analyzed by immunoblotting analysis. For total cell lysates, cells were lysed by NETN buffer (150 mM NaCl, 1 mM EDTA at pH 8.0, 20 mM Tris at pH 8.0, 0.5% NP-40, 25 mM NaF, 2 mM Na3VO4, 5 mM PMSF, 2 g ml-1 aprotinin).
Kinase assay.
For the assay using purified EGFR kinase, a baculovirus expressing GST–EGFR kinase domain fusion protein was purchased from Cell Signaling. To reduce background signal, a PCNA variant with all the tyrosine residues of the protein converted to phenylalanine was used for construction. GST–PCNA fusion proteins with either tyrosine or phenylalanine at residue 211 were then incubated with the purified EGFR kinase for kinase reaction. Ten units of the kinase were incubated at 30 °C for 30 min in 50 l of kinase buffer (60 mM HEPES at pH 7.5, 5 mM MgCl2, 5 mM MnCl2, 3 M Na3VO4 and 1.25 mM DTT) in the presence of 1 Ci -32P ATP and 5 g of the substrates. The reactions were then terminated and separated in 10% SDS–PAGE, followed by Coomassie blue staining and autoradiograph analysis to determine the relative phosphorylation level.
Confocal microscopy.
Cultured cells were fixed with methanol for 15 min at -20 °C, permeabilized with 0.1%
Tween-20 for 15 min and then immunostained with primary antibodies (1:200 dilution in phosphate-buffered saline (PBS) with 0.2% bovine serum albumin (BSA)) overnight at 4 °C.
For EGFR and PCNA, the primary antibodies used were from Santa Cruz. After three washes with PBS, the FITC- or Texas Red-conjugated secondary antibody was then applied for 45
min at room temperature. The nucleus was stained with TOPRO3. The images were captured with an OLYMPUS IX81 laser microscope. For Triton extraction, the cells were washed once with PBS and extracted with 0.2% Triton-X-100 in PBS for 5 min. After washing with PBS, the extracted cells were fixed as described above.
Mass spectrometry.
PCNA was isolated by immunopecipitation with a monoclonal antibody (Santa Cruz) and finally isolated by silver-stained SDS–PAGE. The protein band was excised, subjected to in- gel digestion with trypsin, desalted on a small reversed-phase column (POROS 20R2) and eluted directly into a metallized nanospray needle (Proxeon Biosystems, Odense, Denmark) in 5% formic acid and 60% acetonitrile. The needle was then positioned a few millimetres from the orifice of a QqTOF mass spectrometer (Qstar Pulsar-i) and a 1 kV spray potential was applied to acquire MS and MS-MS spectra of the desired ion (506.7 m/z).
PCNA Y 211 peptide EGFR kinase assay.
The 30 l reaction mix, which includes 60 mM HEPES at pH 7.5, 5 mM MgCl2, 5 mM MnCl2, 3 M Na3VO4, 1.25 mM DTT, 20 M ATP, 1 mM peptide and 10 U EGFR kinase (Cell Signaling), was incubated at 25 °C for 30 min. The volume of the reaction mixture was then brought up to 100 l by PBS, then passed through a spin column of 3 K molecular weight cut-off (Millipore, Billerica, MA). The filtrate was slot-blotted onto a nitrocellulose membrane system. The membrane was blocked with 2% BSA in T-PBS for 1 h at room temperature, followed by incubation with the anti-phosphotyrosine antibody (4G10; Upstate) at 4 °C for 12 h. The membrane was then washed three times with T-PBS and incubated with secondary antibody for 1 h at room temperature to be developed for chemiluminescent detection.
Mismatch-repair assay.
The assay was performed by following the procedure described previously27. To assay for the Flag-tagged wild-type and PCNAY211F, cells of the stable transfectants were washed and suspended in the hypotonic buffer (20 mM HEPES at pH 7.9, 5 mM KCl, 1.5 mM MgCl2, 1 mM DTT, 0.1 mM PMSF) and sonicated. Total cell lysate (1 mg) was then immunoprecipitated using an anti-Flag antibody (M2; Sigma). The protein G-agarose beads were then washed three times with the hypotonic buffer and incubated with the repair reaction mix (25 l) (30 mM HEPES at pH 7.8; 7 mM MgCl2; 4 mM ATP; 200 M of each of CTP, GTP and UTP; 100 M of each of dATP, dGTP, dTTP and dCTP; 40 mM creatine phosphate;
100 mg ml-1 creatine phosphokinase; 15 mM sodium phosphate at pH 7.5; 5 ng of the indicated heteroduplex DNA; and 50 g protein of the cytoplasmic extract prepared from HeLa cells). The endogenous PCNA function in the HeLa lysate was pre-depleted by the
inhibitory p21 peptide, as described previously27. Repair efficiency was calculated as 100 (1 minus mixed bursts derived from extract-treated sample/mixed bursts derived from untreated sample)27. Statistical significance was determined by the Student's t-test. P < 0.05 was considered to be significant.
DNA synthesis assay.
The PCNA-mediated [3H]-thymidine incorporation and chromatin immunoprecipitation were carried out as described previously26, except that anti-Flag antibody (M2; Sigma) was used for immunoprecipitation. Statistical significance was determined by the Student's t-test. P <
0.05 was considered to be significant.
Immunohistochemical staining.
IHC staining was performed as described previously. Sequential tissue sections were incubated with anti-phospho-Y211 PCNA, EGFR and PCNA. Sections were incubated with biotin-conjugated secondary antibody, followed by avidin biotin-peroxidase complex and amino-ethyl carbazole chromogen. The SPSS Kaplan–Meier survival curves were generated using scores of high versus low and negative PCNA and phospho-Y211 PCNA as strata.
Two-sided log-rank analysis was used to assess statistical significance, and the association between discrete variables was tested using the 2 test.
Note: Supplementary Information is available on the Nature Cell Biology website.
Acknowledgements
The anti-phospho-Tyr 211 antibody was generated by Bethyl Laboratories, Inc. as a collaboration. We also thank S. Dent, M. Van Dyke and D. Yu for helpful discussions and comments. This study was supported, in part, by the NIH RO1 109311 and NIH PO1 099031 grants, the National Breast Cancer Foundation, Inc. (to M.-C. H.), and the Cancer Center support grant CA16672.
References
Kelman, Z. & O'Donnell, M. Structural and functional similarities of prokaryotic and eukaryotic DNA polymerase sliding clamps. Nucleic Acids Res. 23, 3613–3620 (1995).
Bravo, R. & Macdonald-Bravo, H. Existence of two populations of cyclin/proliferating cell nuclear antigen during the cell cycle: association with DNA replication sites. J. Cell Biol. 105, 1549–1554 (1987).
Paunesku, T. et al. Proliferating cell nuclear antigen (PCNA): ringmaster of the genome. Int.
J. Radiat. Biol. 77, 1007–1021 (2001).
Haracska, L., Torres-Ramos, C.A., Johnson, R.E., Prakash, S. & Prakash, L. Opposing effects of ubiquitin conjugation and SUMO modification of PCNA on replicational bypass of DNA
lesions in Saccharomyces cerevisiae. Mol. Cell. Biol. 24, 4267–4274 (2004).
Kannouche, P.L. & Lehmann, A.R. Ubiquitination of PCNA and the polymerase switch in human cells. Cell Cycle 3, 1011–1013 (2004).
Jonsson, Z.O., Podust, V.N., Podust, L.M. & Hubscher, U. Tyrosine 114 is essential for the trimeric structure and the functional activities of human proliferating cell nuclear antigen.
EMBO J. 14, 5745–5751 (1995).
Maga, G. & Hubscher, U. Proliferating cell nuclear antigen (PCNA): a dancer with many partners. J. Cell Sci. 116, 3051–3060 (2003).
Yarden, Y. & Sliwkowski, M.X. Untangling the ErbB signalling network. Nature Rev. Mol.
Cell Biol. 2, 127–137 (2001).
Lin, S.Y. et al. Nuclear localization of EGF receptor and its potential new role as a transcription factor. Nature Cell Biol. 3, 802–808 (2001).
Lo, H.-W. et al. Nuclear interaction of EGFR and STAT3 in the activation of the iNOS/NO pathway. Cancer Cell 7, 575–589 (2005).
Hanada, N. et al. Co-regulation of b-Myb expression by E2F1 and EGF receptor. Mol.
Carcinog. 45, 10–17 (2006).
Assy, N. & Minuk, G.Y. Liver regeneration: methods for monitoring and their applications. J.
Hepatol. 26, 945–952 (1997).
Marti, U. & Hug, M. Acinar and cellular distribution and mRNA expression of the epidermal growth factor receptor are changed during liver regeneration. J. Hepatol. 23, 318–327 (1995).
Psyrri, A. et al. Quantitative determination of nuclear and cytoplasmic epidermal growth factor receptor expression in oropharyngeal squamous cell cancer by using automated quantitative analysis. Clin. Cancer Res. 11, 5856–5862 (2005).
Marti, U. et al. Nuclear localization of epidermal growth factor and epidermal growth factor receptors in human thyroid tissues. Thyroid 11, 137–145 (2001).
Tao, Y. et al. Nuclear accumulation of epidermal growth factor receptor and acceleration of G1/S stage by Epstein–Barr-encoded oncoprotein latent membrane protein 1. Exp. Cell Res.
303, 240–251 (2005).
Lo, H.-W. et al. Novel prognostic value of nuclear epidermal growth factor receptor in breast cancer. Cancer Res. 65, 338–348 (2005).
Jeziorski, A., Blonski, J.Z. & Niewiadomska, H. The expression of products of oncogens c- erbB2 and EGFR and proliferating antigens Ki67 and PCNA in primary invasive ductal cancer of female breast. J. Exp. Clin. Cancer Res. 19, 61–67 (2000).
Shrestha, P. et al. Proliferating cell nuclear antigen in breast lesions: correlation of c-erbB-2 oncoprotein and EGF receptor and its clinicopathological significance in breast cancer.
Virchows Arch. A Pathol. Anat. Histopathol. 421, 193–202 (1992).
Frassoldati, A. et al. Changes of biological features in breast cancer cells determined by primary chemotherapy. Breast Cancer Res. Treat. 44, 185–192 (1997).
Hoege, C., Pfander, B., Moldovan, G.-L., Pyrowolakis, G. & Jentsch, S. RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO. Nature 419, 135–141 (2002).
Stelter, P. & Ulrich, H.D. Control of spontaneous and damage-induced mutagenesis by SUMO and ubiquitin conjugation. Nature 425, 188–191 (2003).
Ulrich, H.D. How to activate a damage-tolerant polymerase: consequences of PCNA modifications by ubiquitin and SUMO. Cell Cycle 3, 15–18 (2004).
Kiyokawa, N., Lee, E.K., Karunagaran, D., Lin, S.-Y. & Hung, M.-C. Mitosis-specific negative regulation of epidermal growth factor receptor, triggered by a decrease in ligand binding and dimerization, can be overcome by overexpression of receptor. J. Biol. Chem. 272, 18656–18665 (1997).
Ammosova, T. et al. Nuclear targeting of protein phosphatase-1 by HIV-1 Tat protein. J. Biol.
Chem. 280, 36364–36371 (2005).
Hasan, S., Hassa, P.O., Imhof, R. & Hottiger, M.O. Transcription coactivator p300 binds PCNA and may have a role in DNA repair synthesis. Nature 410, 387–391 (2001).
Umar, A. et al. Requirement for PCNA in DNA mismatch repair at a step preceding DNA resynthesis. Cell 87, 65–73 (1996).
Kato, T. et al. New prognostic factors associated with long-term survival in node-negative breast cancer patients. Breast Cancer 6, 370–377 (1999).
Kato, T., Kameoka, S., Kimura, T., Nishikawa, T. & Kobayashi, M. The combination of angiogenesis and blood vessel invasion as a prognostic indicator in primary breast cancer. Br.
J. Cancer 88, 1900–1908 (2003).
Lee, J.S. et al. Correlation between angiogenesis, apoptosis and cell proliferation in invasive ductal carcinoma of the breast and their relation to tumor behavior. Anal. Quant. Cytol.
Histol. 23, 161–168 (2001).
Bukholm, I.R.K., Bukholm, G., Holm, R. & Nesland, J.M. Association between histology grade, expression of HsMCM2, and cyclin A in human invasive breast carcinomas. J. Clin.
Pathol. 56, 368–373 (2003).
Grossi, F. et al. Prognostic significance of K-ras, p53, bcl-2, PCNA, CD34 in radically resected non-small cell lung cancers. Eur. J. Cancer 39, 1242–1250 (2003).
Heimann, R., Ferguson, D., Recant, W.M. & Hellman, S. Breast cancer metastatic phenotype as predicted by histologic tumor markers. Cancer J. Sci. Am. 3, 224–249 (1997).
Arteaga, C.L. Overview of epidermal growth factor receptor biology and its role as a therapeutic target in human neoplasia. Semin. Oncol. 29, 3–9 (2002).
Baselga, J. Why the epidermal growth factor receptor? The rationale for cancer therapy.
Oncologist 7, 2–8 (2002).
Wang, S.-C. et al. Binding at and transactivation of the COX-2 promoter by nuclear tyrosine kinase receptor ErbB-2. Cancer Cell 6, 251–261 (2004).
Xie, Y.M. & Hung, M.C. Nuclear localization of p185neu tyrosine kinase and its association with transcriptional transactivation. Biochem. Biophys. Res. Commun. 203, 1589–1598 (1994).
Offterdinger, M., Schofer, C., Weipoltshammer, K. & Grunt, T.W. c-erbB-3: a nuclear protein in mammary epithelial cells. J. Cell Biol. 157, 929–940 (2002).
Ni, C.-Y., Murphy, M.P., Golde, T.E. & Carpenter, G. -Secretase cleavage and nuclear localization of ErbB-4 receptor tyrosine kinase. Science 294, 2179–2181 (2001).
Lo, H.W. & Hung, M.C. Nuclear EGFR signalling network in cancers: linking EGFR pathway to cell cycle progression, nitric oxide pathway and patient survival. Br. J. Cancer 94, 184–188 (2006).
Massie, C. & Mills, I.G. The developing role of receptors and adaptors. Nature Rev. Cancer 6, 403–409 (2006).
Carpenter, G. Nuclear localization and possible functions of receptor tyrosine kinases. Curr.
Opin. Cell Biol. 15, 143–148 (2003).
Wells, A. & Marti, U. Signalling shortcut: cell-surface receptors in the nucleus? Nature Rev.
Mol. Cell Biol. 3, 697–702 (2002).
Lo, H.W. et al. Nuclear-cytoplasmic transport of EGFR involves receptor endocytosis, importin 1 and CRM1. J. Cell Biochem. 98, 1570–1583 (2006).
Giri, D.K. et al. Endosomal transport of ErbB-2: a mechanism for nuclear entry of cell surface receptor. Mol. Cell. Biol. 25, 11005–11018 (2005).
Williams, C.C. et al. The ERBB4/HER4 receptor tyrosine kinase regulates gene expression by functioning as a STAT5A nuclear chaperone. J. Cell Biol. 167, 469–478 (2004).
Huang, T.T. et al. Regulation of monoubiquitinated PCNA by DUB autocleavage. Nature Cell Biol. 8, 341–347 (2006).
Asada, M. et al. Apoptosis inhibitory activity of cytoplasmic p21(Cip1/WAF1) in monocytic differentiation. EMBO J. 18, 1223–1234 (1999).