行政院國家科學委員會專題研究計畫 成果報告
轉錄因子 alpha-Pal/NRF-1 在神經細胞分化及學習與記憶 之角色
計畫類別: 個別型計畫
計畫編號: NSC93-2320-B-006-043-
執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立成功大學生理學科(所)
計畫主持人: 黃阿敏
計畫參與人員: 張文騰, 邱榮敬
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 94 年 10 月 31 日
行政院國家科學委員會補助專題研究計畫 ■成 果 報 告
□期中進度報告
轉錄因子 α-Pal/NRF-1 在神經分化之角色
Role of transcription factor α-Pal/NRF-1 in neuronal differentiation
計畫類別:■ 個別型計畫 □ 整合型計畫 計畫編號:NSC 93-2320-B-006-043-
執行期間: 93 年 8 月 1 日 至 94 年 7 月 31 日
計畫主持人:黃阿敏 共同主持人:
計畫參與人員: 張文騰、邱榮敬
成果報告類型(依經費核定清單規定繳交):□精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:
中 華 民 國 94 年 10 月 28 日
附件一
中文摘要
本計畫原本申請為期兩年,審查通過為一年計畫,因此針對第一年的計畫內容執行。
延續前兩年的研究成果,本年度對於α-Pal/NRF-1 促進神經軸突生長的機制進一步探討。本 年度主要進展為:(一)發現 α-Pal/NRF-1 確實促進神經軸突生長,其所調控的下游基因 integrin-associated protein (IAP)及其蛋白產物參與其中。如果將 IAP 基因的反義 cDNA 送入 神經纖維瘤細胞株 IMR-32 中,發現 IAP 蛋白含量顯著下降,並且抑制 α-Pal/NRF-1 所促進 的神經軸突生長。此項結果與前一年的結果合併發表於 2005 年 Biochemical and Biophysical Research Communication。(二)確認 mitogen-activated protein kinase (MAPK)訊息路徑參 與α-Pal/NRF-1 所調控的神經軸突生長。以 U0126 抑制 MAPK 上游訊息分子時,顯著抑制 IMR-32 細胞神經軸突生長。進一步發現,U0126 能抑制大量表現 α-Pal/NRF-1 和血清剝奪 所促進的 IAP 啟動子活性、α-Pal/NRF-1 與 DNA 結合能力和 IAP mRNA 表現量。以上結果 確認 α-Pal/NRF-1 促進神經軸突生長的角色,MAPK 訊息路徑參與其中,此現象可能經由 活化α-Pal/NRF-1 與 IAP 基因啟動子 DNA 結合的能力,並且與 IAP 啟動子活性及 IAP 基因 表達增加有關。
關鍵字:神經軸突生長、訊息路徑、血清剝奪、轉錄因子、啟動子活性、基因表達
英文摘要
This project was originally submitted as a two-year project. The final fund is for one year.
Therefore, we performed the project of the first year. In this study, we confirmed the role of
-Pal/NRF-1 in neurite outgrowth and investigated whether the mitogen-activated protein kinase protein involved in the increase of neurite outgrowth by-Pal/NRF-1. Results showed that if the antisense IAP cDNA was introduced into the IMR neuroblastoma cells, the IAP protein level was reduced and the percentage the neurite-bearing cells was decreases. These results, combined with the results from previous study, were published in Biochemical and Biophysical Research Communication (2005). We also confirmed that the mitogen-activated protein kinase (MAPK) pathway is involved in -Pal/NRF-1-increased neurite outgrowth. The MAPK kinase (MEK) inhibitor, U0126, inhibited neurite outgrowth increased by ectopic expression of-Pal/NRF-1 as well as serum deprivation in IMR-32 cells. Further results demonstrated that U0126 inhibited the IAP promoter activity, the DNA binding activity of -Pal/NRF-1 and the IAP mRNA levels in cells overexpressed with -Pal/NRF-1 or cultured in serum free medium. These results suggest that the MAPK signaling cascade is one of the upstream signaling pathways that are involved in the activation of-Pal/NRF-1 during neurite outgrowth.
Key words: neurite outgrowth, signaling pathway, serum deprivation, transcription factor, promoter activity, gene expression
研究目的
1. To correlate the protein level of integrin-associated protein and neurite outgrowth
2. To examine if mitogen-activated protein kinase signaling pathway involved in the enhancement ofα-Pal/NRF-1 in neurite outgrowth
前言
We have found that the transcription factor-Pal/NRF-1 is a critical activator of the IAP gene in the human neuroblastoma cells IMR-32.-Pal/NRF-1 belongs to a new class of transcription factors that contain a unique putative basic leucine zipper (bZip) DNA-binding domain (Efiok et al., 1994). -Pal was first discovered as a key transcription factor in the eukaryotic initiation factor 2gene, which is a target of the post-translation mechanisms when eukaryotic cells respond to growth, metabolic, and other signals (Efiok et al., 1994; Jacob et al., 1989). NRF-1 was discovered independently as a nuclear transcription factor that is important for the regulation of mitochondrial genes (Evans and Scarpulla, 1990). Later studies found that the -Pal/NRF-1 recognition sequence can be identified in many other genes involved in energy transduction, translation/protein turnover, DNA synthesis/repair, and cellular proliferation (Efiok et al., 1994; Efiok and Safer, 2000). This suggests that -Pal/NRF-1 functions to modulate transcription of metabolic genes required for a variety of biological functions, including cellular growth.
-Pal/NRF-1 shows strong homology in the DNA-binding domain with two invertebrate genes, sea urchin P3A2 and Drosophila erect wing gene (ewg). The homology is restricted in the DNA-binding domain (Virbasius et al., 1993; Efiok, et al., 1994). These two genes have been implicated in embryonic or larval development (Calzone et al., 1991; DeSimone and White, 1993). On the other hand, -Pal/NRF-1 shows 91% identity to its homologue in zebrafish, not really finished (nrf) (Becker et al., 1998). Both ewg and nrf have been associated with the development of the central nervous system (DeSimone and White, 1993, Becker et al., 1998). These studies suggest that -Pal/NRF-1 may regulate the growth of neurons, including neurite outgrowth and synaptogenesis, during development. Therefore, we hypothesize that -Pal/NRF-1, as a critical transcription factor of IAP, will play a role in neurite differentiation, such as neurite outgrowth.
The next question is: what is the upstream signaling pathway of -Pal/NRF-1 in the regulation of neurite outgrowth? Mitogen-activated protein kinase (MAPK) pathway is well known to regulate neuronal differentiation, neurite outgrowth, and neuronal survival (Patapoutian and Reichardt, 2001; Miller and Kaplan, 2003). Therefore, we also test the possibility that the MAPK pathway is involved in the activation of -Pal/NRF-1 in the function of promoting neurite outgrowth.
研究方法
Cell Culture –Human neuroblastoma cell line IMR-32 was obtained and maintained as described previously (Chang and Huang, 2004). IMR-32 cells were grown in minimum essential medium Eagle with Earle’s salt base (Sigma-Aldrich, St Louis, MO) supplemented with or without 10% fetal bovine serum (FBS) (Hyclone, Logan, UT). Cells were cultured at 37°C in a humidified chamber containing 5% CO2. The MAPK kinase (MEK) inhibitors PD98059 (25 or 50M) and U0126 (10, 25 or 40 M) were used to inhibit the activity of MAPK.
Gel electrophoretic mobility shift assay (EMSA) –Preparation of nuclear proteins, labeling of oligonucleotides, and EMSA were performed as described previously (5). The oligonucleotides IAP -Pal/NRF-1 (f), 5’-GAG TGC GCG TGC GCG GCT CT, and IAP -Pal/NRF-1 (r), 3’-TCA CGC GCA CGC GCC GAG AG, were annealed and labeled with [-32P]dCTP using a
fill-in reaction. Three g of nuclear extract and 2 104 cpm of 32P-labeled double-stranded oligonucleotides were added to a reaction mixture containing 10 mM Tris-Cl (pH 7.5), 50 mM sodium chloride, 0.5 mM dithiothreitol, 0.5 mM EDTA, 1 mM magnesium chloride, 4% glycerol, and 0.05g poly(dI-dC)poly(dI-dC). The binding reaction was performed at 4°C for 30 min and analyzed on a 4% native polyacrylamide gel at 10 V/cm for 2.5 h. The gel was dried and analyzed using autoradiography. The intensity of shifted bands was quantified using 1-D advanced Universal Software (American Applied Biotechnology, Fullerton, CA).
RT-PCR –For the analysis of IAP transcripts after serum deprivation in IMR-32 cells, the IAP cDNAs were amplified using the primer pair Hiap14 (5’-TAA CCT CCT TCG TCA TTG CC) and Hiap15 (5’-CGT AAG GGT CTC ATA GGT G). PCR parameters were: 94°C for 3 min, 94°C for 20 s, 53°C for 25 s, 72°C for 25 s, for 24 cycles, and 72°C for 7 min. The PCR products were analyzed on a 2% agarose gel in 0.5TBE. The intensity of bands was quantified using 1-D advanced Universal Software (American Applied Biotechnology).
Transient transfection –The plasmids pCMS--Pal/NRF-1-FL and pCMS-GFP empty vector were transfected into IMR-32 cells using the calcium phosphate precipitation method (17). Cells (1.5 105) were cultured on 6-well plates for 24 h. The medium was exchanged with 2 ml of fresh medium, and cells were incubated for additional 1 h for the subsequent transfection.
Plasmid DNA (1 g) was mixed with 5 l of 2.5 M calcium chloride in a volume of 50 l containing 0.1TE buffer. The mixture was then mixed with one volume of 2HEPES buffer and incubated for 1 min at room temperature. The mixture was added to the cells for transfection.
After 12 h of transfection, the medium was exchanged with fresh medium and the cells were grown for neurite measurement on the following days.
Promoter assay –The IAP promoter activity was assayed as described previously (5).
IMR-32 cells (1.5105) were plated on each well of 6-well plates for 24 h and transfected with 50 ng of the IAP promoter construct pGL3-232 alone or co-transfected with 50 ng of pGL3-232 and 0.5 g of the control vector pcDNA3.1 or the pcDNA3.1-NRF-1 construct for different experiments using the calcium phosphate precipitation method (17). After a 12-h transfection, the medium was changed to fresh medium and cells were cultured for the subsequent analyses. The cell lysates were prepared by scraping the cells from plates with 1passive lysis buffer (Promega, Madison, WI). The IAP promoter activity was determined using the Luciferase Reporter Assay System (Promega) and the total proteins were quantified with BCA Protein Assay Reagent (Pierce Biotechnology Inc., Rockford, IL). Due to cell loss of cultures in serum-free medium, luciferase activity was normalized with the concentration of the total protein in each sample.
Measurement of neurite outgrowth –To determine the neurite process length, GFP-positive cells were observed under the fluorescent microscope (Leica). The length of the longest process for each of > 100 cells was determined for each sample using Metamorph software (Universal Imaging Corporation, Downingtown, PA). A process longer than 25 m is defined as a neurite, which is about 1.5 times cell-body diameter.
Statistical analysis –All results were analyzed and graphs were drawn using GraphPad Prism 3.0 software (Cadmus Professional Communications, Richmond, VA). Statistical analysis was performed using unpaired t tests for pairwise comparisons. Statistical significance was set at p <
0.05.
結果及討論
U0126 inhibited-Pal/NRF-1-induced neurite outgrowth
The involvement of the MAPK pathway in the function of -Pal/NRF-1 in neurite outgrowth was examined by the treatment of U0126 in IMR-32 cells transiently transfected with the cDNA construct containing the full length-Pal/NRF-1, pCMS--Pal/NRF-1-FL. As reported in the previous study, transfection of full length-Pal/NRF-1 cDNA significantly increased the percentage of neurite-bearing cells (Chang et al., 2005). U0126 inhibited the percentage of neurite-bearing cells in-Pal/NRF-1-transfected cells in a dose- and time-dependent manner (Fig.
1). We lowered down the concentrations of U0126 to 10 or 25M to avoid non-specific effects.
As a comparison, U0126 treatment for 1 day at these concentrations did not decrease the percentage of neurite-bearing cells in empty vector-transfected IMR-32 cells, although prolonged treatment of U0126 for 2 or 3 days slightly decrease neurite outgrowth in these cells (Fig. 1).
These results revealed that the MAPK pathway is involved in -Pal/NRF-1-induced neurite outgrowth.
U0126 decreased the IAP/CD47 promoter activity increased by overexpression of -Pal/NRF-1 and serum deprivation
The function of -Pal/NRF-1 in neurite outgrowth is mediated through its downstream gene IAP/CD47 (Chang et al., 2005). Therefore, we examined if U0126 inhibited the activation of promoter activity of IAP/CD47 by -Pal/NRF-1. As reported before (Chang and Huang, 2004), transfection of full length -Pal/NRF-1 cDNA significantly increased the IAP promoter activity.
However, the increases were significantly reduced by the treatment of U0126 at 10 and 25 M for 24 h (Fig. 2A). Serum deprivation increased P promoter activity both at 8 and 16 hr after treatment. This increase was significantly reduced by the treatment of 25M of U0126 (Fig. 2B) U0126 decreased the DNA binding activity of-Pal/NRF-1 increased by serum deprivation
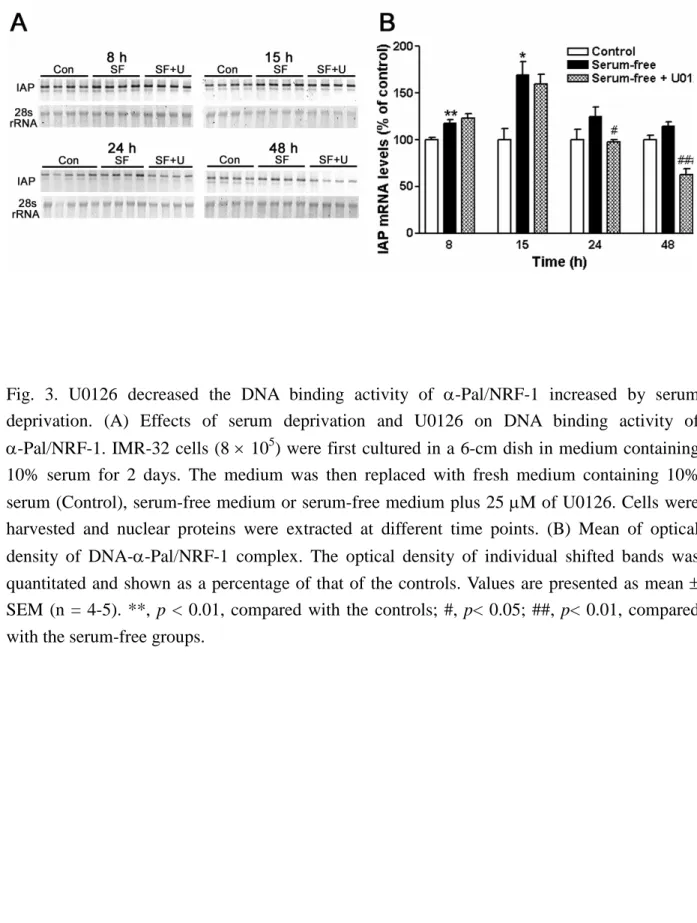
A gel electrophoretic mobility shift assay (EMSA) was used to determine the DNA binding activity of -Pal/NRF-1 after serum deprivation and U0126 treatment. The binding activity of-Pal/NRF-1 was significantly increased at 8 h and returned to the control level at 16 and 24 h after serum deprivation (Fig. 3A). U0126 was treated for 4, 8, 16 and 24 hr. When treated with 25 M of U0126 for 16 h, the DNA binding activity of -Pal/NRF-1 was significantly reduced. Interestingly, the DNA binding activity of -Pal/NRF-1 was significantly increased after 4-h treatment of U0126. (Fig. 3B)
U0126 decreased the IAP mRNA level increased by serum deprivation
Semi-quantitative RT-PCR was used to measure the IAP mRNA level after serum deprivation. The expression levels of IAP transcripts were significantly increased at 8 h and peaked at 15 h after serum starvation. Increases in the IAP mRNA levels were reduced by treatment with U0126 and the reduction reached the significant level at 24 h and 48 h (Fig. 5A and B).
參考文獻
Becker, T. S., Burgess, S. M., Amsterdam, A. H., Allende, M. L., and Hopkins, N. (1998) not really finished is crucial for development of the zebrafish outer retina and encodes a transcription factor highly homologous to human nuclear respiratory factor-1 and avian initiation binding repressor. Development 125:4369-4378.
Calzone, F. J., Höög, C., Teplow, D. B., Cutting, A. E., Zeller, R. W., Britten, R. J., and Davidson, E. H. (1991) Gene regulatory factors of the sea urchin embryo: I. Purification by affinity
chromatography and cloning of P3A2, a novel DNA-binding protein. Development 112, 335-350
DeSimone, S. M., and White, K. (1993) The Drosophila erect wing gene, which is important for both neuronal and muscle development, encodes a protein which is similar to the sea urchin P3A2 DNA binding protein. Mol. Cell. Biol. 13:3641–3649.
Efiok, B. J. S., Chiorini, J. A., and Safer, B. (1994) A key transcription factor for eukaryotic initiation factor-2is strongly homologous to developmental transcription factors and may link metabolic genes to cellular growth and development. J. Biol. Chem. 269:18921-18930.
Efiok, B. J. S., and Safer, B. (2000) Transcriptional regulation of E2F-1 and eIF-2 genes by -Pal:
a potential mechanism for coordinated regulation of protein synthesis, growth, and the cell cycle. Biochim. Biophy. Acta 1495:51-68.
Evans, M. J., and Scarpulla, R. C. (1990) NRF-1: a trans-activator of nuclear encoded respiratory genes in animal cells. Genes Dev. 4:1023-1034.
Jacob, W. F., Silverman, T. A., Cohen, R. B., and Safer, B. (1989) Identification and characterization of a novel transcription factor participating in the expression of eukaryotic initiation factor 2.J. Biol. Chem. 264, 20372-20384.
Miller F. D. and Kaplan, D. R. (2003) Signaling mechanisms underlying dendrite formation. Curr. Opin. Neurobiol. 13, 391-398.
Patapoutian, A. and Reichardt, L. F. (2001) Trk receptors; mediators of neurtrophin action.
Curr. Opin. Neurobiol. 11, 272-280.
Virbasius, C. A. Virbasius, J. V. and Scarpulla, R. C. (1993) NRF-1, an activator involved in nuclear-mitochondrial interactions, utilizes a new DNA-binding domain conserved in a family of developmental regulators. Genes Dev. 7, 2431-2445.
計畫成果自評:
此計畫原本申請為兩年期,審查通過為一年期,所以執行第一年的計畫。延續前兩年 的研究成果,本年度對於α-Pal/NRF-1 促進神經軸突生長的機制進一步探討。本年度主要進 展為:(一)發現α-Pal/NRF-1 確實促進神經軸突生長,其所調控的下游基因 integrin-associated protein (IAP)及其蛋白產物參與其中。如果將 IAP 基因的反義 cDNA 送入神經纖維瘤細胞株 IMR-32 中,發現 IAP 蛋白含量顯著下降,並且抑制 α-Pal/NRF-1 所促進的神經軸突生長。
此 結 果 與 前 一 年 的 結 果 合 併 發 表 在 2005 年 Biophysical and Biochemical Research Communication (334:199-209, 2005)。
(二)確認 mitogen-activated protein kinase (MAPK)訊息路徑參與 α-Pal/NRF-1 神 經軸突生長。以 U0126 抑制 MAPK 上游訊息分子時,顯著抑制 IMR-32 細胞神經軸突生長。
進一步發現,U0126 抑制 α-Pal/NRF-1 以及剝奪血清所促進的 IAP 啟動子活性、α-Pal/NRF-1 與 DNA 結合能力和 IAP mRNA 表現量。此部分結果正在整理發表中。整體而言,本計畫 執行結果符合原先第一年所提的計畫內容。
Fig. 1. U0126 inhibited-Pal/NRF-1-increased neurite outgrowth. IMR-32 cells were transfected with pCMS-GFP vector (1g) or pCMS--Pal/NRF-1-FL (1 g) and grown in 10% FBS medium containing DMSO or U0126 (10 or 25 M) for 1, 2, or 3 days. Quantification of neurite outgrowth was determined as described above (n = 4). ***, p < 0.001 compared with vehicle; ##, p < 0.01; ###, p < 0.001 compared with the -Pal/NRF-1 samples; &, p < 0.05; &&, p < 0.01 compared with the vehicle samples.
Fig. 2. (A) U0126 inhibited -Pal/NRF-1-increased IAP promoter activity. IMR-32 cells (1.5 105) were co-transfected with IAP promoter construct pGL3-232 (50 ng) and pcDNA3.1 vector (0.5 g) or pcDNA3.1-NRF-1 (0.5 g). After transfection, IMR-32 cells were treated with serum-free medium containing DMSO or U0126 (10 or 25 M) for 24 h. Luciferase assay was used to measure the IAP promoter activity (n = 6). ***, p < 0.001 compared with the controls; ##, p < 0.01; ###, p < 0.001 compared with the -Pal/NRF-1 samples. (B) U0126 inhibited serum deprivation-increased IAP promoter activity. IMR-32 cells (1.5 105) were cultured on 6-well plate and transfected with the IAP promoter construct pGL3-232 (50 ng) for 12 h. After transfection, the cells were cultured in medium containing 10% FBS, serum-free medium, or serum-free medium plus 10 or 25 M of U0126 for 8 and 16 h. Luciferase assay was used to measure the IAP promoter activity. The luciferase activity was normalized with the concentration of the total protein in each sample. Values are presented as mean SEM (n = 4-5). *, p < 0.05;
***, p < 0.001, compared with the controls; ###, p < 0.001, compared with the serum-free groups.
Fig. 3. U0126 decreased the DNA binding activity of -Pal/NRF-1 increased by serum deprivation. (A) Effects of serum deprivation and U0126 on DNA binding activity of
-Pal/NRF-1. IMR-32 cells (8 105) were first cultured in a 6-cm dish in medium containing 10% serum for 2 days. The medium was then replaced with fresh medium containing 10%
serum (Control), serum-free medium or serum-free medium plus 25 M of U0126. Cells were harvested and nuclear proteins were extracted at different time points. (B) Mean of optical density of DNA--Pal/NRF-1 complex. The optical density of individual shifted bands was quantitated and shown as a percentage of that of the controls. Values are presented as mean SEM (n = 4-5). **, p < 0.01, compared with the controls; #, p< 0.05; ##, p< 0.01, compared with the serum-free groups.
Fig. 4. U0126 inhibited the increase of the IAP mRNA level after serum deprivation. IAP mRNA levels were determined using semi-quantitative RT-PCR. At each time point after treatment, two
g of total RNA was obtained from serum-free (SF) samples, serum-free samples treated with 10
M of U0126 (SF+U), or control (Con) samples. Individual bands were quantitated, normalized by the intensity of their own 28S rRNA, and shown as a percentage of the control. Values are presented as meanSEM (n = 4). *, p < 0.05; **, p < 0.01; compared with the controls; #, p <
0.05; ###, p < 0.001; compared with the serum-free samples.