NATIONAL TAIWAN UNIVERSITY
COLLEGE OF LIFE SCIENCE
INSTITUTE OF ECOLOGY AND EVOLUTIONARY BIOLOGY
&
UNIVERSITÉ MONTPELLIER II
SCIENCES ET TECHNIQUES DU LANGUEDOC
DOCTORAL SCHOOL: SYSTÈMES INTÉGRÉS EN BIOLOGIE, AGRONOMIE, GÉOSCIENCES, HYDROSCIENCES, ENVIRONNEMENT
Doctoral Dissertation
Colonization and adaptations of Ficus in Taiwan
Presented and defended by:
Anthony BAIN (
邊安台)
January 2012
JURY
Lien-Siang CHOU, Professor, National Taiwan University Advisor Martine HOSSAERT-McKEY, Directeur de recherche CNRS, CEFE Advisor Philippe GRANDCOLAS, Directeur de recherche CNRS, MNHN Reviewer Hsy-Yu TZENG, Professor, National Chung Hsing University Reviewer
Jer-Ming HU, Professor, National Taiwan University Examiner
Finn KJELLBERG, Directeur de recherche CNRS, CEFE Examiner
Wen-Jer WU, Professor, National Taiwan University Examiner
II
III
Acknowledgements
Like most of the people who have done a PhD thesis, I will not forget this part of my life. This thesis brought me far from my home, my friends and my family in a far far-away country. I’ve been living in Taipei for four years now, the first months in Asia were everyday discoveries. The Taiwanese people are one of the most welcoming people I have met, I feel like home now in this country. Further than the scientific maturation that the work I have done gave me, the life experience I get was far from what I could expect: I have seen a different culture, different point of view on the world and life. I am really glad that I have made the choice to move to the other side of the planet.
The common urban legend says that a butterfly flying in the forest can be the first air movement that will start a hurricane. In my case, this dissertation could not be without the encounter of two peoples. They met in 2007 at the fig symposium in China and I think it is because they appreciate each other that they were interested to work together making me travel to Taiwan few months later and then move to Taipei.
Lien-Siang Chou and Martine Hossaert-McKey are my advisors for this work, I thank them gratefully for the support and the help during this Ph.D. work.
Secondly, I could not live and study in Taiwan without funding. I really want to thank the Taiwan Ministry of Education and the Bureau de Représentation de Taipei en France to give the opportunity to French students like me to do their PhD in Taiwan. Furthermore I would like to acknowledge the Agence Nationale pour la Recherche (ANR) and the National Science Council (NSC) for the funding of my research work.
I would like to thank also the Committee of my oral defense for its attendance and advices: Dr. Chou Lien-Siang, Dr. Hossaert-McKey Martine, Dr. Grandcolas Philippe, Dr. Hu Jer-Ming, Dr. Kjellberg Finn, Dr. Tzeng Hsy-Yu and Dr Wu Wen- Jer.
I was welcomed in the lab in National Taiwan University by a full cohort of nice students that help me to do my first steps in Taiwan and in the culture. I would give special and sincere thanks to Hui-Wen ( 惠雯) who helps me so much for anything from the administrative maze to the phenology work during the summer.
IV
Without her, this thesis would be much lighter. Big thanks too for the students who help me with the phenology during the summers when I was back in France, so thank you (for that and far more) Minson (黃銘), Yun-Peng (允芃), Guan-Yi (冠誼), I- Chiao (伊喬), Yu-Ting(于庭), Wen-Hsuan (玟璇). My labmates in Dr. Chou’s lab were the first to endure my endless questions about anything, so thank you Harry (子 皓), Chun-Chieh (俊傑), Ula (欣怡), Inge (瑩), Ming-Ching (明慶) and Weilung (維 倫). But I don’t forget the others members of the lab: Thank you Ivy (志慧), Meng- Chen (孟辰), Taco (明章), Odin(祥廈), Sun-Young (上揚), Damien (達勉), Athelan (毓璞), Yi-Ting (依婷), Zhong-Bin (忠斌) and the two Miss Yang (楊姐和雪蘭姐). I am very happy to know you all.
Outside the NTU lab, I was also welcomed in National Chung Hsing University. I wanted to cheerfully thank Dr. Tzeng who helped me a lot to become familiar with the Taiwanese fig trees and to bring me to Orchid Island and to travel with me to China. Thank you for your knowledge and for your kindness. Thank you to introduce me to the people at the Forestry Institute Recreation Area at Kenting. The people there were also very welcoming (as all the Taiwan people I think) so thanks to Ju-Yi (如怡), Shu-Hui (淑惠) and Ms Ha.
Thank you to the Philippines people who helped us when we were there. The warmest welcome I ever received. Thank you to Lilian. Thank you to Daniel and Leonardo who have both leave us here one year ago.
My thesis was a cooperation work between the Taiwanese and the French teams, I would like to come back to France to thank some peoples. Firstly my thesis would be tasteless without her work and my days in the French lab would be less happy if she was not there. I want to thank deeply and sincerely Marjorie for being a part of this thesis. She became quickly a very good friend. Secondly I would like to thank Florian for passing a lot of time with me using microscopes. For your sight loss, I want to thank you. My insect knowledge will be nothing without Jean-Yves Rasplus, thank you for the hours using microscope and telling me what is what. For the good atmosphere in the French lab I would like to say thank you also to Nicolas, Thomas, Delphine, Ben, Roxanne (and thanks for your travels to the university for me), Marie, Julien, Noppolito, Laurinette and Bruno (at CBGP).
V
I would like to give one of the biggest thanks to Finn Kjellberg. He has been my field partner for all my trips outside France or Taiwan. We had good and bad experiences together and he’s always been there for my rush work. As Martine, since my beginning in lab science he is there. If I am able to defend this work it is because of his help, of the long conversations about fig trees, the digs into the data. If I understand the topic I am working on, it is because of Finn. I would like to thank you for the hours you passed with me to talk about figs or on my uncertain drafts, for the kilometers we walk, we drive looking for figs. Thank you, Finn.
Concerning my personal life I would like to thank my mother to let me follow my own path without doubting one moment about what I was doing. I’m sure that when you have a kid living in a faraway country that does not help to sleep well some nights. Thank you for everything. Thank you my little sister Rachel to make my parents discover the new technologies and be more in contact together. I am sorry that I am far and don’t see you growing up. Thank you Yoann and Adélaïde to be what you are: both crazy, both funny but so different. Thank you. Thank you Michel for all you’ve done for us all these years.
Thank you to all my friends in France to be still my friends after all these years far from you. Thank you Guilhem, Katia, Valérian, Julia, Raphaël, Audrey, Zahia, Jim, François, Damien, Bastien, Jacko, Julien, Thierry, Karima, Cécile, Guilhem, Gael, Marina, Lucie, Thomas, Pascal, theVernet and the Familia Gladiatori.
Thank you to my friends in Taiwan. The one I especially would like to thank is Martin. He makes me discover Taipei and all my friends today was his before.
Thank you to let me enter your universe and made me discover its treasure. Thank you to the everyday Pierre, to Thomas, Gemma, Matthieu, Aka, Jon, Mike, Celena, Andy, Fiona, Pauline, Cacia, Yeah, Brian, Kenji,Claudi-a and to the two “vieux de la vieille” Bruno and Fabien. Thank you for these crazy hours. Thank you to the picnic team where I wrote this work, I felt home.
The final thanks to my Emma: Thank you to be.
THANK YOU
VI
VII
致謝 致謝 致謝 致謝
就像多半念唸過博士的人一樣,而我不會忘記這次的旅程。這本論文,
它帶我離家數千里遠。住在台北四年了,剛開始,每個月都有很多很多新的發 現,台灣人是我遇過最友善的人群之一,因為她他們的友善,漸漸的讓我感 覺,台灣,似乎就像是我自己的國家。我在這裡累積的生活經驗,早就超越了 我的工作:我看見了不同的文化,對於人生、對於世界的各種角度。我很高興 我當時選擇飛到世界的另一頭–台灣。
首先,我想要說的是,如果沒有下面這兩位,這本論文可能就不存在 了。2007 年,他她們在中國的 榕樹座談會榕樹及榕小蜂學術研討會相遇,而 我相信我能夠來到台灣旅行,以及到台北生活的原因,是因為他們兩個人的關 係非常密切。周蓮香和 Martine Hossaert-McKey 是我這份工作的指導教授,
我想感謝他們的支持與幫助。再次感謝 Martine,謝謝妳當在我還在念讀碩士 的時候,當我的指導教授,也謝謝妳在我念唸博士的時候給予的寶貴意見。
再者,如果沒有資金,我也不可能在台灣生活及學習。我想感謝台灣教 育部和駐法國台北代表處贊助像我一這樣的法國學生,提供在台灣念唸博士的 機會。
當我到達台灣大學的實驗室時,有一大群非常熱情的學生歡迎、幫助 我。他們教我在台灣怎麼生活、並且帶我安穩的走入下一個階段。我想特別感 謝惠雯在暑假期間,不僅僅是在行政、物候上的幫助。沒有她的話,這本論文 就薄很多。也要感謝在每個夏季裡,當我回法國時,還在台灣幫我做物候工作 的學生: 黃銘、允芃、冠誼、伊喬、于庭和玟璇。也感謝在周教授實驗室裡,
第一個幫我解決問題的人:子皓、俊傑、欣怡、瑩、明慶和維倫。但然而我也 不會忘記這實驗室裡,其他有幫助過我的人:謝謝志慧、孟辰、明章、祥下 廈、上揚、達勉、毓璞、依婷, 忠斌,以及兩位楊小姐,真的很高興認識你 們。
在台大的實驗室外面,也很多人歡迎我。我想感謝曾喜育教授博士,讓我了解 對台灣的榕樹的了解,也感謝他帶我去蘭嶼和中國旅行。謝謝你的知識與貼 心。謝謝你介紹墾丁林試所恆春熱帶植物園國家風景區的人給我。那邊的人也 很熱情(就像我所熟悉的台灣人一樣),所以謝謝 如怡、 淑惠和阿好姐。
謝謝我們待在菲律賓的時候所幫助我們的菲律賓人。那是我生平受過最 熱情的歡迎。謝謝 Lilian。謝謝現在在天堂的 Daniel 跟 Leonardo。
我的論文是台灣法國兩團合作的結果,我現在想感謝法國的一些人。沒 有她的話,我的論文將會是白紙一張,我在法國的實驗室裡,沒有她,我也不 這麼快樂。我想特別感謝 Marjorie,她是我在法國,一起研究環境時, 與寫這 本 論 文 時 , 佔 了 很 重 要 的 角 色 。 我 們 很 快 的 成 為 了 好 朋 友 。 我 也 想 謝 謝 Florian , 花 這 麼 多 時 間 把 眼 睛 放 在 顯 微 鏡 上 , 而 那 樣 幾 乎 等 於 一 個 瞎
VIII
子...。也感謝 Jean-Yves Rasplus,沒有他,我對於昆蟲這種東西,就會 一蓋概不知,謝謝你在顯微鏡旁邊,花那麼多時間跟我解釋什麼是什麼。謝謝 在法國的實驗室裡帶給我很多好氣氛的 Nicolas, Thomas, Delphine, Ben, Roxanne(還要謝謝你每一次都陪我去大學), Marie, Noppolito, Laurinette 跟 Bruno(在 CBGP)。
我想給 Finn Kjellberg 最大的感謝。他是我在台灣、法國野外旅行的 原野考察良師益友。我們一起分享的經驗,無論是好的、壞的 ,每次我必須趕 工作,他都會在我身邊。就像 Martine 在實驗室陪我一樣,他從頭到尾是都參 與著的。因為他的幫助、因為我們討論很多有關榕樹的事、因為我們不斷的在 分析資訊,而我才有辦法答辯這本論文。我想感謝你花這麼多時間,陪我擬我 還很 不確定的草 稿、陪我繼 續聊榕樹、跟我走很長很長的 路、跟我坐 車尋 找...。謝謝你,Finn。
有關我個人的生活,我想謝謝我媽媽,讓我從來都沒有懷疑過我到底在 做什麼,而且勇敢的走下去。我知道,當你的孩子在另一個國度獨自生活著,
做媽 媽的,有時 一定很難入眠。媽媽,謝謝你給我的這一 切。謝謝我 妹妹 Rachel ,讓我父母接觸新科技之外,也更接觸彼此。我好抱歉我離妳很遠,無 法看著妳長大....。謝謝 Yoann 和 Adélaïde 你們這兩個神經病,不只很好 笑,又很不一樣。謝謝你們。謝謝 Michel,謝謝你這麼多年來為我們做的一 切。
謝謝我在法國的所有朋友,即使過了這麼多年沒見面,你們還是把我當 成 好 朋 友 : Guilhem, Katia, Val é rian, Julia, Raphaël, Audrey, Zahia, Jim, François, Damien, Bastien, Jacko, Julien, Thierry, Karima, C é cile, Guilhem, Gael, Marina, Lucie, Thomas, Pascal, the Vernet 和 the Familia Gladiatori。
謝謝我在台灣的朋友。要特別感謝的是 Martin。是他讓我發現台北的,
我身邊每一個朋友都是他介紹的朋友。謝謝你讓我進入你的宇宙,讓我發現這 宇 宙 裡 的 寶 物。 謝 謝 天 天 Pierre, Thomas, Gemma, Matthieu, Aka, Jon, Mike, Celena, Andy, Fiona, Pauline, Cacia, Yeah, Brian, Kenji 小客。
謝謝你們給我這麼多瘋狂的時間。謝謝野餐咖啡裡的每一個人。我是在野餐咖 啡寫這本論文的,它是我的家。
最後一份感謝要給我的 Emma:謝謝妳存在。
謝謝謝謝 謝謝謝謝
IX
Remerciements
Comme la plupart des gens qui ont sont passés par une thèse, c’est une aventure que je n’oublierais pas. Cette thèse m’a amené loin de chez moi, de mes amis et de ma famille dans un lointain pays. Je vis à Taipei depuis maintenant quatre ans, les premiers mois asiatiques furent emplis de découvertes journalières. Les taiwanais sont des gens parmi les plus accueillants que j’ai rencontrés, je me sens maintenant comme chez moi dans ce pays. Au-delà la maturation scientifique que le travail accompli m’a permis d’obtenir, les expériences de vie de ces quatre années sont loin de tout ce que j’aurais pu imaginer d’une autre culture, vision du monde et de la vie. Je suis vraiment heureux d’avoir fait ce choix et d’être parti à l’autre bout de la planète.
Il est souvent dit qu’un battement d’aile de papillon peut déclencher un cyclone a l’autre bout du monde. Pour moi c’est la rencontre de deux personnes qui fut à l’ origine du cyclone. Elles se sont rencontrées en 2007 au colloque international sur les figuiers en Chine et je pense que c’est parce qu’elles se sont appréciées l’une l’autre que j’ai posé le pied à Taiwan quelques mois plus tard puis déménager à Taipei. Lien-Siang Chou et Martine Hossaert-McKey furent mes directrices pour cette thèse, Je voudrais vraiment les remercier chaleureusement pour l’aide et le soutien pendant cette thèse. Merci aussi tout particulièrement Martine qui fut ma directrice de stage pendant mon master.
Ensuite, je n’aurais pas pu vivre et étudier à Taiwan sans un soutien financier.
Je voudrais vraiment remercier le Ministère taiwanais de l’Education et le Bureau de Représentation de Taipei en France de donner l’opportunité à des étudiants français de vivre et faire leur thèse à Taiwan. Enfin je remercie l’Agence Nationale pour la Recherche (ANR) et le National Science Council (NSC) pour le financement de mon travail de recherche.
Mes remerciements aussi aux membres de mon jury de thèse : Dr. Chou Lien- Siang, Dr. Hossaert-McKey Martine, Dr. Grandcolas Philippe, Dr. Hu Jer-Ming, Dr.
Kjellberg Finn, Dr. Tzeng Hsy-Yu et le Dr Wu Wen-Jer.
Je fus aussi formidablement bien accueilli dans mon labo à la National Taiwan University par une cohorte d’étudiants bien sympathiques qui m’ont aidé à faire mes
X
premiers pas dans le culture taiwanaise et littéralement à Taipei. Ma première pensée ira à Hui-Wen ( 惠雯) qui m’a tellement aide … que ce soit pour me sortir du labyrinthe administratif que pour mon suivi phénologique Durant mes étés français.
Sans elle, ma thèse serait bien plus légère. De grand mercis aussi aux autres étudiants qui m’ont aidés pendant les étés : Minson (黃銘), Yun-Peng (允芃), Guan-Yi (冠誼), I-Chiao (伊喬), Yu-Ting(于庭) et Wen-Hsuan (玟璇). Mes camarades de bureau dans le labo du Dr. Chou furent les premiers à être soumis à la question sur à peu près n’importe quel sujet alors merci Harry (子皓), Chun-Chieh (俊傑), Ula (欣怡), Inge (瑩), Ming-Ching (明慶) et Weilung ( 維倫). Mais je n’oublie les autres qui ne partageaient pas le bureau : Merci Ivy (志慧), Meng-Chen (孟辰), Taco (明章), Odin (祥廈), Sun-Young (上揚), Damien (達勉), Athelan (毓璞), Yi-Ting (依婷), Zhong- Bin (忠斌) et les deux Madame Yang (楊姐和雪蘭姐). Je suis vraiment content de vous connaitre.
En-dehors de la NTU, je fus aussi sympathiquement accueilli à la Chung Hsing University. Je voudrais remercier chaleureusement le Dr. Tzeng qui m’a beaucoup aide pour me familiariser avec les espèces de figuiers locales, qui m’a amené à Orchid Island et avec qui je suis allé en Chine. Merci pour votre savoir et votre gentillesse. Merci aussi de m’avoir fait découvrir le Forestry Institute Recreation Area à Kenting et ses gens tout aussi accueillants. Merci Ju-Yi (如怡), Shu-Hui (淑惠) et Madame Ha.
Merci aux philippins qui nous aidèrent pendant nos séjours là-bas. L’accueil asiatique le plus caliente que j’eu. Merci Lilian. Ainsi qu’à Daniel et Leonardo qui nous ont quitté tous les deux il y a un peu plus d’un an.
Ma thèse fut une coopération entre une équipe taiwanaise et une équipe française, je voudrais revenir en France pour parler de la part française de ce travail.
Tout d’abord ma thèse serait insipide sans son travail et les journées au CEFE moins marrantes si elle n’avait pas été là. Merci profondément Marjorie pour avoir fait partie de cette thèse. Elle fut rapidement une très bonne amie. Ensuite je voudrais remercier Florian pour s’être bien détruit les yeux à trier des insectes avec moi. Mon savoir entomologique déjà limité ne serait rien sans mes moments passes avec Jean-Yves Rasplus, merci pour m’avoir dit ce qu’étaient ces choses minuscules sous nos yeux.
Pour la bonne ambiance dans les locaux français merci à Nicolas, Thomas, Delphine,
XI
Ben, Roxanne (et merci pour les allers retours jusqu’à la fac), Marie, Julien, Noppolito, Laurinette et Bruno (du CBGP).
Je voudrais dire un très grand merci à Finn. Il a été mon compagnon d’expédition pour tous mes voyages en dehors de la France et de Taiwan. Nous avons eu des expériences plus ou moins bonnes ensemble et il a toujours été là pour le travail de dernière minute. Comme Martine, depuis mes débuts dans le monde de la science, il est là. Si je fus capable de soutenir ma thèse c’est Grace à lui, grâce aux longues conversations sur les figuiers et à l’exploration des données. Si aujourd’hui je comprends le sujet sur lequel je travaille c’est grâce à lui. Je voudrais te remercier pour toutes ces heures à discuter ou sur mes versions douteuses, pour ces kilomètres parcourus à pied, en voiture a la recherche de figuiers. Merci, Finn.
Plus personnellement je voudrais dire merci à ma mère pour m’avoir laissé suivre ma voie sans douter un moment de ce que je voulais faire. Je pense qu’avoir un enfant qui vit à l’autre bout de la planète ne doit pas aider à avoir le sommeil serein certaines nuits. Merci pour tout. Merci à ma petite sœur Rachel pour avoir fait découvrir a mes parents les nouvelles technologies qui nous permettent d’être plus en contact qu’auparavant. Je suis désolé d’être loin et de ne pas te voir grandir. Merci à mon frère et ma sœur, Yoann et Adélaïde pour être ce qu’ils sont : à la fois tarées et drôles mais si différents. Merci. Merci Michel pour ce que tu as fait pour nous toutes ces années.
Merci à mes amis en France pour l’être resté alors que j’étais loin de vous.
Merci Guilhem, Katia, Valérian, Julia, Raphaël, Audrey, Zahia, Jim, François, Damien, Bastien, Jacko, Julien, Thierry, Karima, Cécile, Guilhem, Gaël, Marina, Lucie, Thomas, Pascal, les Vernet et la Familia Gladiatori.
Merci à mes amis à Taiwan aussi. J’aimerais remercier particulièrement Martin. Il me fit découvrir Taipei et mes amis d’aujourd’hui furent d’abord les siens.
Merci de m’avoir laissé entrer dans ton univers et ou j’y ai trouvé un trésor. Merci au journalier Pierre, Thomas, Gemma, Matthieu, Aka, Jon, Mike, Celena, Andy, Fiona, Pauline, Cacia, Yeah, Brian, Kenji, Claudi-a et aux deux vieux de la vieille Bruno et Fabien. Merci pour toutes ces heures de folie. Merci à l’équipe du Picnic Café ou j’ai écrit cette thèse, je m’y suis senti comme chez moi.
Enfin merci à ma Emma : Merci d’être là.
MERCI
XII
XIII Abstract:
The Island of Taiwan appeared some two million years ago. We investigated the Ficus species of Taiwan in order to understand how the island was colonized and how insular conditions affected the colonizing species. Genus Ficus is characterized by its mutualistic association with highly dispersive pollinating wasps. Further figs, the closed inflorescences of Ficus, are consumed by a large number of highly efficient seed dispersers. We developed a set of portable genetic markers in order to analyze several species using a single set of markers. We then analyzed genetic structuring within Taiwan in a set of four species. One species presented clear genetic differentiation in the warm south Taiwan, suggesting the possibility of incipient ecological speciation in a highly dispersive species. Another species presented indications of ongoing selective processes also in south Taiwan. On a broader scale we show that microsatellite data and notably R statistics provide useful insights into Ficus phylogeography. In three species, continental Asia seems to correspond to a single genetic background despite strong genetic differentiation among populations.
For Ficus variegata, we show that it colonized Taiwan from the Philippines and we detect no trace of gene flow from the continent. An investigation of Ficus reproductive phenology suggests that only species producing frequent fig crops have established on the island. This work opens up a series of perspectives on the response of Ficus to insular conditions.
Key words: Ficus, phylogeography, insularity, Taiwan
XIV 摘要 :
臺灣島約在 200 萬年前形成,此論文研究臺灣的榕屬植物,探討物種於新興島 嶼的拓殖和島嶼環境對物種分佈的影響。榕屬植物其互利共生的授粉小蜂擅於 長途播遷,榕屬植物的隱頭花序──榕果──也被許多種高效率的種子傳播者 取食。在此論文中,我們建立一套屬內通用的基因標記,以同一套分子標記分 析不同物種。接著,利用四個榕樹種分析臺灣的遺傳結構。其中雀榕在溫暖的 南臺灣呈現明顯的遺傳分化,顯示此高度變異種類可能處於生態種化的初期;
而另有徵兆顯示在南臺灣的稜果榕正在進行選汰過程。在更大的尺度下,此研 究顯示微衛星體DNA資料,特別是 R 統計量,可針對榕屬的親緣地理學提供 有用的理解,在許多的物種上,亞洲大陸似乎提供了單一的遺傳背景,儘管在 族群間有強烈的基因分化;然而在幹花榕是由菲律賓拓殖而來,未值測到任何 與大陸的基因交流。最後,針對X種榕樹於臺灣南北兩處的物候調查顯示,唯 有可頻繁結果的榕樹種可在此島建立族群。本研究為榕樹在島嶼環境的反應開 啟了一系列的觀點。
關鍵詞:榕屬;親緣地理;孤立;台灣
XV Résumé :
L’ile de Taiwan est apparue, il y a deux millions d’années. Nous avons étudié les figuiers de Taiwan afin de comprendre comment l’ile fut colonisée et comment l’insularité a pu affecter les espèces colonisatrices. Le genre Ficus est caractérisé par son association mutualiste avec des insectes pollinisateurs extrêmement dispersifs. De plus les figues, inflorescences fermées des Ficus, sont consommés par un grand nombre d’espèces qui elles aussi dispersent efficacement les graines. Nous avons développé une série de marqueurs génétiques afin de pouvoir étudier plusieurs espèces avec les mêmes outils. Ensuite nous avons analysé la structure génétique dans Taiwan d’un groupe de quatre espèces. Une espèce présenta une différenciation génétique claire dans le Sud de Taiwan au climat plus chaud, suggérant la possibilité d’un début de spéciation écologique chez une espèce hautement dispersive. Une seconde espèce a montré des indices de processus de sélection en cours dans le Sud de Taiwan. A une échelle plus étendue, nous avons montre que les données microsatellites et spécifiquement les R statistiques donnent un premier aperçu de la phylogéographie des figuiers. Pour plusieurs espèces, l’Asie continentale semble être la trame de fond génétique de ces espèces malgré une forte différenciation parmi les populations. Pour Ficus variegata, nous avons montré que la colonisation de Taiwan a pour origine les Philippines et nous n’avons pas détecté de trace de flux génétique venant du continent. L’étude de la phénologie reproductive des Ficus suggère que seulement des espèces produisant de fréquentes récoltes ont colonisé l’ile. Ces travaux ouvrent de nombreuses perspectives sur la réponse des figuiers à l’insularité.
Mots-clés : Ficus, phylogéographie, insularité, Taiwan
XVI
XVII Résumé substantiel:
L’ile de Taiwan est apparue, il y a deux millions d’années. Elle est positionnée entre deux entités floristiques distinctes : Le continent asiatique et les Philippines, ainsi la colonisation de Taiwan est partie de ces deux zones. De plus, Taiwan a un climat subtropical avec un fort gradient de température et de précipitations du nord au sud de l’ile. Via deux aspects principaux (génétique des populations et phénologie), nous avons étudié les figuiers de Taiwan afin de comprendre comment l’ile fut colonisée et comment l’insularité a pu affecter les espèces colonisatrices. Le genre Ficus est caractérisé par son association mutualiste avec des insectes pollinisateurs extrêmement dispersifs. De plus les figues, inflorescences fermées des Ficus, sont consommées par un grand nombre d’espèces qui elles aussi dispersent efficacement les graines. Nous avons développé une série de marqueurs génétiques microsatellites sur un large échantillon d’espèces de figuiers venant du monde entier et représentant la diversité génétique du genre afin de pouvoir étudier plusieurs espèces avec les mêmes outils. Ensuite nous avons analysé la structure génétique dans Taiwan d’un groupe de quatre espèces. Une espèce, Ficus subpisocarpa, présenta une différenciation génétique claire dans le Sud de Taiwan au climat plus chaud, suggérant la possibilité d’un début de spéciation écologique chez une espèce hautement dispersive. Une seconde espèce, Ficus septica a montré des indices de processus de sélection en cours dans le Sud de Taiwan. Contrairement à Ficus subpisocarpa, F. benguetensis a montré une différentiation claire dans le nord de l’ile.
Quant à Ficus erecta, elle ne présenta pas de différentiation génétique sur toute l’étendue de l’ile. Cette espèce étant particulièrement abondante à une altitude plus élevée que les autres figuiers de Taiwan, elle n’a vraisemblablement pas subi de
XVIII
pression de sélection amenant à une différentiation d’un quelconque niveau entre les populations géographiques. A une échelle plus étendue, nous avons montre que les données microsatellites et spécifiquement les R statistiques donnent un premier aperçu de la phylogéographie des figuiers. Pour les trois espèces présentes sur le continent, l’Asie continentale semble être la trame de fond génétique malgré une faible différenciation parmi les populations. Plus précisément, pour Ficus variegata, nous avons montré que la colonisation de Taiwan a pour origine les Philippines et nous n’avons pas détecté de trace de flux génétique venant du continent. Alors que ces traces sont évidentes pour F. erecta et F. subpisocarpa. De plus la colonisation de Taiwan vient vraisemblablement des iles Matsu et/ou du Fujian en Chine. L’étude de la phénologie reproductive des Ficus a montré que les populations portent constamment des figues sur leurs branches. F. erecta faisant exception mais lorsque qu’il croit dans l’extrême Sud de Taiwan, sa croissance a tendance à aller vers un mode continu plutôt que saisonnier. La phénologie des figuiers de Taiwan sont donc en opposition avec celle des espèces continentales qui est particulièrement saisonnière.
Cela suggère que seulement des espèces produisant de fréquentes récoltes ont pu coloniser l’ile. Enfin le suivi régulier des espèces taiwanaises a amené à découvrir une interaction myrmécophytique entre F. subpisocarpa et plusieurs espèces de fourmis.
Des structures spécifiques dans les branches ont été mises en évidence et ont été confirmé dans des échantillons d’herbier provenant du sud-est asiatique. Pour conclure, ces travaux ouvrent de nombreuses perspectives sur la réponse des figuiers à l’insularité.
XIX
Colonization and adaptations of Ficus in Taiwan
Contents
Validation sheet I
Acknowledgements III
致謝 VII
Remerciements IX
Abstract XIII
摘要 XIV
Résumé XV
Résumé substantiel XVII
General introduction 1
XX
Review: Ficus in Taiwan 35
Article One: Portable microsatellite primers for Ficus (Moraceae) (accepted in American Journal of Botany Notes & Protocols)
55
Article Two: Four Ficus species in Taiwan: is genetic differentiation possible despite long distance dispersal?
69
Article Three: A genetic and phylogeographic study of five Ficus species occurring in Taiwan and Continental China or the Philippines: insights on diversification in a long distance dispersal genus
99
Article Four: Adaptations and constraints on dioecious Ficus phenology
127
Article Five: A new case of ants nesting within branches of a fig tree: the case of Ficus subpisocarpa in Taiwan (accepted in Sociobiology)
163
General Conclusion 193
Annexes 201
1
GENERAL INTRODUCTION
2
3 General introduction:
The current distribution of species is the consequence of past events that have considerably changed the environment. During the Last Glacial Maximum, the size of the continents was greater, the geography and the climate were quite different from today. Changes occurred shaping the current world. Species ranges expanded, and retracted leading to the distributions we observe today. Analysis of the spatial structure of genetic diversity has permitted, for example, inference of pathways of colonization, such as the tracks of post-glacial expansion of many species from refugia after the Last Glacial Maximum (Hewitt, 2004; Muellner et al., 2005) or inference of inter-continental plant migrations (Dick et al., 2007; Erkens et al., 2009;
Muellner et al., 2006). However in some regions of the world, the history of the latest expansions is unclear. For example in Australasia, numerous studies have tried to delimit biological regions and limits for the floras and the faunas, such as the Wallace line, but the limits are still debated today as they are species dependent (Cox, 2001;
Esselstyn et al., 2010; van Welzen et al., 2011). Southeast Asia is at the center of these debates, firstly because of the incredible biodiversity of these regions (Myers, 2000) and secondly the diversity is specific to certain areas such as Japan, or the Philippines.
In this region, contact zones between different biogeographic provinces may provide a unique key to determine patterns of species occurrence and distribution. Indeed, what preserves the identities of biotas in contact zones between long-separated floras and faunas? Do competition and pre-emption of space limit the spread of ecologically redundant species into the other biota? In this prospective, Taiwan presents a fascinating opportunity to analyse ecological and evolutionary processes in sites of
4
active biotic exchange: it is a recent contact zone [less than 2.5 million years old (Malavieille et al., 2002)] between two different biotas. It formed as a totally new island, which may have been connected to the mainland but if so, only at glacial maxima. Its biogeographic neighbours are the Asian continent and the Philippines, an old isolated group of islands that has never been part of a continent (Sibuet and Hsu 2004). This allows monitoring how two floras and faunas may mix. Hence Taiwan is a model system to investigate processes occurring during the early stages of such contacts. The situation is thus also of relevance to questions about biological invasion, an important aspect of contemporary global change.
Figure 1: The geographic situation of Taiwan. From the map it is apparent that Taiwan can be colonised from the continent and from the Philippines. The link between the Philippines and New Guinea over the Moluccas is also conspicuous. A continental plateau with shallow waters is joining Borneo, Java, Sumatra and the Asian mainland and forms Sundaland. The waters separating Borneo from the Philippines are much deeper, so that the latter islands were never in contact with Sundaland (Heaney et al. 2005). Hence during warm climatic episodes, a moist tropical flora could have colonised Taiwan from the Philippines, but also from the Indochinese peninsula.
5
Moreover Taiwan is a relatively small island. Islands are generally depauperate in species relative to mainland regions, yet they are home to some of nature’s most striking adaptive radiations. The low species richness of islands can be attributed both to their limited size, which increases extinction rates, and to their isolation (MacArthur and Wilson 1967). At the same time, some clades present substantial ecological and phenotypic diversity on islands, a diversification which probably results from evolutionary radiation into empty niches associated with low species numbers (Algar and Losos, 2011) in agreement with the ecological theory of adaptive radiation (Schluter, 2000). Investigating early stages of small island colonization by highly dispersive species may allow to detect early phases of ecological niches expansion and specialization into these ecological niches, a process that could lead to speciation.
Taiwan is a unique territory in Asia. With a high rate of endemism, the Taiwan flora displays a surprising mix of the Philippines and Chinese floras (Hsieh, 2002). These two floras are most different. Indeed numerous authors have tried to analyze the specificities of the Philippine flora (Simpson, 1977). The past links with the continent (including Japanese Islands) have been highlighted in the studies of closely related species such as Cunninghamia konishii which is directly derived from its continental sister species C. lanceolata (Lu et al., 2001; Hwang et al., 2003; Chung et al., 2004), similarly with Lilium formosanum and the Japanese L. longiflorum (Hiramatsu et al., 2001) or again for Rhododendron species (Brown et al., 2006; Chung et al., 2007). At the species level, Taiwan species display important local differentiation from the populations of the areas of origin (Chiang et al., 2001; Huang et al., 2002; Huang et al., 2004; Cheng et al., 2005; Huang and Lin, 2006). In Taiwan, the Central Mountain Range (CMR over 2,000m high) represents an insurmountable barrier for plant
6
species (Lin, 2001; Huang et al., 2002; Cheng et al., 2005) but few studies have described the effect of the latitude on the genetic structure. Indeed the island of Taiwan is a subtropical territory (Yen and Chen, 2000; Chen et al., 2005): while in the north the climate is cold, but no frost at sea level, the southern part of Taiwan is warmer with one yearly rainy season. Studies have shown either no population structuring throughout Taiwan (Chung et al., 2004; Cheng et al., 2005; Wu et al., 2006; Shih et al., 2007) or north-south structuring of the populations, a feature which has always been attributed to post glaciations expansion from refugia for plants (Huang et al., 2004; Cheng et al., 2005; Liao et al., 2010) as well as for terrestrial mammals (Yuan et al., 2006). Nevertheless for Cyclobalanopsis glauca (tropical affinity), the inferred refuge location was located in the Southeast of Taiwan (Huang et al., 2002) while the inferred refuge location for Trochodendron aralioides (temperate affinity) is in the North of Taiwan (Huang et al., 2004).
To go further in our understanding on the current biogeographic patterns of species and on community diversity, we decided to study a group of interacting organisms that has become a model system in community ecology: figs, their specific pollinators and associated insect communities (Herre et al. 2008).
Fig trees belong to genus Ficus. With about 800 species worldwide, this is the most speciose tree genus (Berg, 1989). There are 26 Ficus taxa in Taiwan. Their distribution areas have been established recently (Tzeng, 2004). The fig trees in Taiwan present a suite of more or less unique characters. First, as opposed to other plant genera in Taiwan (Hsieh, 2002), there are only few endemic species (two endemics, for 26 taxa). Second, many of the Taiwanese Ficus have widespread distributions. An excess of widespread species is actually an excess of species with
7
strong dispersal capacity. These special characters (few endemic species and an important number of widely distributed species) support the hypothesis that Taiwan is a territory recently open to colonization by plants, a situation which is quite different from other well-studied old island such as New Caledonia, although colonisation is more recent than previously asumed (Grandcolas et al., 2008) and which present 76%
endemic plant species (Jaffre et al., 1998). Finally, the non-endemic Ficus of Taiwan may originate from two sources: the continent (including Japan) and the Philippines Archipelago. Of the 26 fig species, only five occur both on the Asian continent and the Philippines. Comparatively to the global tropical plant flora of Taiwan which includes a number of species present both in the Philippines and on the continent (Hsieh, 2002), the fig tree flora is clearly separated in two entities (Table 1). It is important to note that even some species of tropical origin thrive in northern Taiwan (e.g. Ficus septica, F. benguetensis and F. variegata) and some more temperate species thrive in the subtropical part of Taiwan in the south (e.g. Ficus erecta var.
beecheyana and F. subpisocarpa). To summarize, the Ficus flora of Taiwan is a mixture of the Philippine and continental Asia species. They present mainly tropical climate affinity. Ficus is the only ubiquitously locally diverse genus in tropical lowland rainforest and this is achieved by a large portion of the regional pool of species being present locally (Harrison, 2005). However on islands, diversity may be reduced. This is the case in Taiwan. For instance there are only four species of monoecious hemiepiphytic Ficus species in Taiwan (Corner, 1965). Of these two belong to section Conosycea (Ficus microcarpa and F. benjamina var. bracteata) and two belong to section Urostigma (F. subpisocarpa and F. caulocarpa).
8
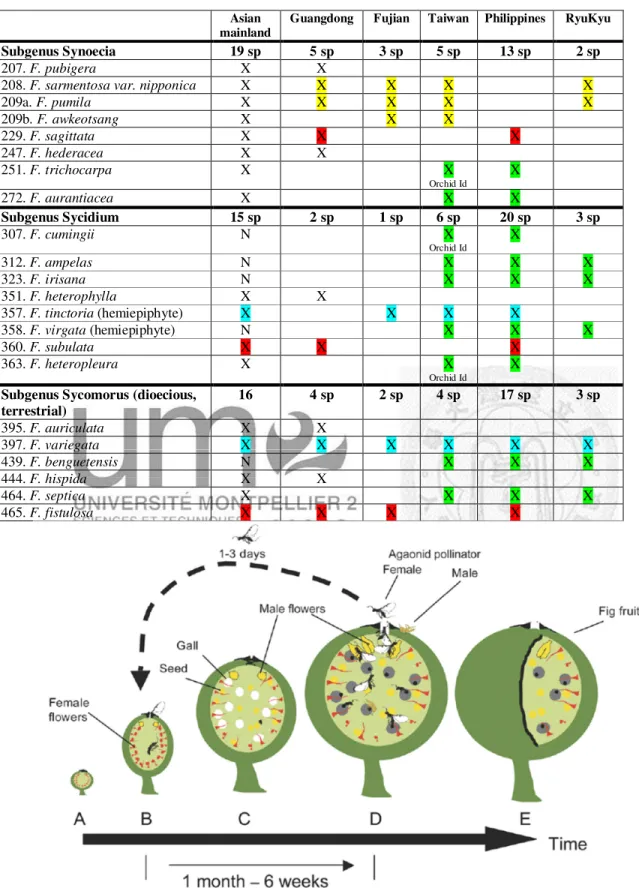
Table 1: Complete list of Ficus species recorded in Taiwan, and the continental provinces of Fujian and Guangdong. Presence of these species on the Asian mainland, RyuKyu and the Philippines is indicated. For presence in Taiwan, Orchid Island is indicated when the species does not occur on the main island of Taiwan but only on Orchid Island.
Species are numbered according to Corner’s (1965) checklist and named according to the latest publications (Berg and Corner, 2005).
Asian mainland
Guangdong Fujian Taiwan Philippines RyuKyu
Subgenus Urostigma Subsection Urostigma (monoecious hemiepiphytic)
15 sp 3 sp 2 sp 2 sp 5 sp 1 sp
5. F. subpisocarpa X X X X X
7. F. concinna X X X X
10. F. virens X X X X
12. F. caulocarpa X X X
Subsection Conosycea (monoecious hemiepiphytic)
48 sp 7 sp 2 sp 2 sp 21 sp 1 sp
22. F. annulata X X X
27. F. drupacea X X X
33. F. altissima X X X
34. F. pubilimba X X X
47. F. glaberrima X X X
68. F. benjamina X X X X
72. F. microcarpa X X X X X X
Section Oreosycea (monoecious terrestrial)
6 sp 3 sp 1 sp 2 sp 6 sp 0 sp
104. F. callosa X X X
105. F. vasculosa X X
114. F. pubinervis N X
Orchid Id.
X
117. F. nervosa X X X X
Subgenus Ficus (dioecious, mostly terrestrial)
Subgenus Ficus Section Ficus 17 sp 13 sp 7 sp 5 sp 6 sp 1 sp F. ruyuanensis (species incerta
sedis)
X X
157. F. pyriformis X X X
159. F. variolosa X X X
160. F. pedunculosa N X X
161. F. erecta X X X X X
162. F. trivia X X
164. F. heteromorpha X X X
165. F. gasparriniana X X X
166. F. pandurata X X X
167. F. stenophylla X X X
169 F. formosana X X X
F. sinociliata (species incerta sedis) X X F. undulata (species incerta sedis) X X
170. F. tannoensis N X (SE)
171. F. vaccinioides N X (SE)
175. F. abellii X X X
Subgenus Ficus Section Eryosycea
13 sp 3 sp 3 sp 2 sp 4 sp 0 sp
181. F. esquiroliana X X X X
182. F. hirta X X X
190. F. ruficaulis N X (S) X
196. F. langkokensis X X X
9 Table 1: continued
Asian mainland
Guangdong Fujian Taiwan Philippines RyuKyu
Subgenus Synoecia 19 sp 5 sp 3 sp 5 sp 13 sp 2 sp
207. F. pubigera X X
208. F. sarmentosa var. nipponica X X X X X
209a. F. pumila X X X X X
209b. F. awkeotsang X X X
229. F. sagittata X X X
247. F. hederacea X X
251. F. trichocarpa X X
Orchid Id
X
272. F. aurantiacea X X X
Subgenus Sycidium 15 sp 2 sp 1 sp 6 sp 20 sp 3 sp
307. F. cumingii N X
Orchid Id
X
312. F. ampelas N X X X
323. F. irisana N X X X
351. F. heterophylla X X
357. F. tinctoria (hemiepiphyte) X X X X
358. F. virgata (hemiepiphyte) N X X X
360. F. subulata X X X
363. F. heteropleura X X
Orchid Id
X Subgenus Sycomorus (dioecious,
terrestrial)
16 4 sp 2 sp 4 sp 17 sp 3 sp
395. F. auriculata X X
397. F. variegata X X X X X X
439. F. benguetensis N X X X
444. F. hispida X X
464. F. septica X X X X
465. F. fistulosa X X X X
Figure 2: The successive fig development phases in a monoecious species (from Harrison, 2005).
10
Moreover, Ficus are part of an obligate pollination mutualism. Due to the closed urn- shaped inflorescence and a very important number of available ovules (to feed the pollinator’s larva), the pollination of figs depends on obligatory and specific pollinating wasps (Hymenoptera, Agaonidae sensu Cruaud et al., 2010). Together they are the actors of an obligate and specific mutualistic relationship with special co- adaptations. The specific fig-fig wasp mutualism seems to follow locally the one-to- one rule (one Agaonidae species for one Ficus species). This idea was basically established by Janzen in 1979 but recent studies have shown a trend that Ficus could often have more than one pollinator species (Lopez-Vaamonde et al., 2001; Cook and Rasplus 2003; Molbo et al., 2003; Machado et al., 2005; Haine et al., 2006; Lin et al., 2011; Moe et al., 2011; Cornille et al. in press). Nevertheless, the fig and fig wasp association remains highly specific (Jousselin et al., 2008). The female pollinator has to find the receptive fig (Phase B sensu Galil and Eisikowitch, 1969 and Figure 2) in the middle of the vegetation. At the receptivity phase, the figs emit volatiles compounds to attract the pollinators (Grison-Pigé et al., 2002; Proffit et al., 2008;
Hossaert-McKey et al. 2010). The insects will approach preferentially the odor of its host tree. Proffit et al. (2008) show that the fig trees produce specific bouquets of odors different among species; nevertheless, it is important to note that the variation among trees of the same species is important, too.
Beyond the pollinating wasps, a whole community of specific chalcidoid wasps (Hymenoptera, Chalcidoidea) also develops within the figs. These non-pollinating fig wasps (NPFW) species are poorly known in Taiwan as in other parts of the world except Africa (van Noort and Rasplus, 1997). Only one complete study has been done on Ficus microcarpa (Chen et al., 1999) establishing the presence of 20 species.
These species are external parasites: they oviposit through the fig wall to reach the
11
ovules they aim to parasitize. When their ovipositor is inside the fig, the NPFW are exceptionally vulnerable to predators. Former studies have shown that ants may thrive in fig trees, feeding on NPFW (Schatz et al., 2006). Pollinating wasps are also predated by ants living on fig trees in the tropics (Schatz et al., 2008) and in temperate areas (Schatz and Hossaert-McKey, 2003). Ants may be numerous on fig trees in the tropics (Schatz et al., 2008; Bain, pers. obs.) but their presence is poorly documented.
With the exception of the work by Schatz, there have been few studies on ant-fig trees interactions. Some studies have investigated ant-Hemipteran interactions on figs (Compton and Robertson, 1988; Dejean et al., 1997; Cushman et al., 1998) and a study has shown that Ficus pisifera (also known as F. obscura var. borneensis; Berg and Corner, 2005) is a myrmecophytic plant (Maschwitz et al., 1994). The path followed by ants to colonise Taiwan is unknown. We may wonder if the strong associations with ants which sometimes prevail in the tropics have reached as far north as Taiwan as associations with ants seem to become less strong outside the tropics.
Using this quite unique model system in community ecology, the main objectives of the present work was to analyse colonisation patterns of this recent island, starting by the fig species. This will be the first step in our understanding of the origins and subsequent evolution of community structure and functioning of interacting species.
In order to perform such studies we selected sufficiently abundant species that include species that have most probably colonized Taiwan from the continent, and species that have colonized Taiwan from the Philippines.
0 10 20 30 40 50 60
Apocrypta Sycophaga
Sycophila
Platyneura
Sycoscapter Platyneura
Inquiline - parasite Gall-maker
Fig Diameter (mm)
Pre-receptive Receptive ---Post-receptive--- Maturity Pollinator
13
Figure 3: The five phases of Ficus erecta var. beecheyana fig development. Prereceptive figs (A phase) are greenish, the ostiole is tightly closed. At the beginning of the receptivity period (B phase), the ostiole becomes looser, allowing female pollinating wasps to enter inside the fig. The longest phase is the interfloral phase (C phase). The figs are hard and have lost their shine and the ostiole is tightly closed. Here birds have peaked at the C phase fig.
During the wasp emergence phase for male figs (D phase) and the final ripening stage for female figs (E phase), the figs swell and become soft. Male D phase figs become brownish yellow, while female figs become purplish white to dark purple.
Ficus erecta Thunb. (Figure 3) belongs to section Ficus of subgenus Ficus. It is a dioecious shrub or a treelet and is found in Japan, Korea, Ryukyu Islands, and in continental China as far north as Jiangsu and inland to Yunnan. It is thus a species adapted to cold conditions (for a Ficus species) with a marked winter. Two varieties and two forms were retained by Corner (1965). The variety occurring in Taiwan is beecheyana (Hook. et Art.) King and the variety erecta on the continent.
14 Figure 4: Ficus variegata figs.
Ficus variegata Blume (Figure 4) belongs to subgenus Sycomorus section Sycomorus, subsection Neomorphe. It grows, often as a pioneer, into a large free standing dioecious tree. The species is extremely widespread, from China to Northern Australia (Queensland). In China it is present from south Yunnan to Fujian. Thus it reaches even less far north than F. subpisocarpa but it occurs further inland. It could have colonized Taiwan either from the Philippines or from the continent.
15
Figure 5: Ficus subpisocarpa figs. Ficus subpisocarpa is monoecious and cauliflorous. The figs grow on the branches and trunk, with strong synchrony.
Ficus subpisocarpa Gagnep. (Figure 5) belongs to subgenus Urostigma, section Urostigma, subsection Urotigma. It is a relatively small monoecious hemiepiphyte often growing on rocks. It is found in Southern Japan, Eastern continental China (northern limit, south east Zhejiang), Taiwan, Vietnam, Cambodia, Northeast Thailand and might be present in the Mollucas (Eastern Indonesia archipelago) (Berg et al., 2011)). Hence it is a species that thrives in relatively cool climates along the seacoast, but it is not found in the colder climates and at high altitudes where Ficus
16
erecta var. beecheyana can occur. Moreover it is often observed in urban environments.
Figure 6: The five phases of Ficus septica fig development. Prereceptive figs (A phase) are light green and shiny, the ostiole is tightly closed. At the beginning of the receptivity period (B phase), the ostiole becomes looser, allowing female pollinating wasps to enter inside the fig. The longest phase is the interfloral phase (C phase). The figs are hard and have lost their shine and the ostiole is tightly closed. During the wasp emergence phase for male figs (D phase) and the final ripening stage for female figs (E phase), the figs swell and become soft.
Their color changes to greenish yellow. Nevertheless while D phase male figs characteristics are very similar with those of C phase figs, E phase female figs keep swelling and finally burst open.
Ficus septica Burm.f. (Figure 6) belongs to section Sycocarpus of subgenus Sycomorus. It is usually a small dioecious tree but may reach 25m. It is common from the Ryukyu Islands and Taiwan to the Vanuatu and throughout the islands of Malesia, but it is absent from continental Asia. As such it is a species of equatorial and tropical distribution, and Taiwan represents an extreme in its distribution.
17
Figure 7: The Ficus benguetensis figs. This species grows figs in cluster on the trunk of by pair on the branches. Often ants are living between the figs and the trunk inside the clusters.
The E phase female figs burst open as F. septica figs.
Ficus benguetensis Merr. (Figure 7) also belongs to section Sycocarpus of subgenus Sycomorus. It is a dioecious shrub of tree up to 15 m tall. It is limited to the Ryukyu Islands, Taiwan and the Philippines. According to Berg (2011) it belongs to the species group of F. ixoroides (Borneo), F. lepicarpa (Malesia in general) and F.
ternatana (Moluccas) and is hence not closely related to F. septica.
18
Figure 8: The five phases of Ficus pedunculosa var. mearnsii fig development. A phase figs are greenish purple and shiny. The receptive figs are hairy and still greenish purple. At the beginning of the C phase, the ostiole becomes very tight. The final phases of fig development are very similar in male and female figs: they become dark purple.
Ficus pedunculosa Miq. var. mearnsii (Merr.) Corner (Figure 8) belongs to subgenus Ficus section Ficus subsection Frutescentia. The distribution of F. pedunculosa extends from Taiwan to New Guinea. It has very large seeds for a Ficus and belongs to a group of species including other species with very large seeds such as F.
deltoidea, which is either epiphytic or grows on very poor sandy soil (e.g. var.
arenaria), and indeed F. pedunculosa var. pedunculosa lives on harsh rocky environments (Bain, pers. obs.). This suggests that the affinity of the species is rather with the Malesian species of subsection Frutescentia, i.e. it belongs to an equatorial group. The form present in Taiwan is var. mearnsii. It is a prostrate shrub on sea- coasts and mountains, but in Taiwan it is absent on mountains. The variety is to be
19
observed in Luzon (Philippines) and in the islands stretching from Luzon to the main island of Taiwan (Babuyan and Batanes Islands, Orchid Island).
Figure 9: The five phases of Ficus tinctoria subsp. swinhoei fig development. Pre- receptive figs are green to brownish green. Receptive figs are greenish yellow, the ostiole is shinny and darker. At the beginning of the interfloral phase (C phase), the ostiole becomes tightly closed and the fig color gets darker. The final phase of the fig development is very different between male and female figs: male figs become yellow and swollen while the female figs become red to purple and are less swollen.
Ficus tinctoria G. Forst. subsp. swinhoei (King) Corner (Figure 9) belongs to subgenus Sycidium, section Palaeomorphe (Berg and Corner, 2005), a section that includes all the hemiepiphytic species of dioecious Ficus. Subspecies swinhoei is a scandent shrub on rocks and in Taiwan it is mainly restricted to sea-shores. It is also found in Mindanao. Berg and Corner (2005) included the subspecies within F.
20
tinctoria subspecies tinctoria, i.e. within the subspecies of equatorial distribution. On the Asian continent the form present is subspecies gibbosa. Hence subspecies swinhoei has an equatorial affinity.
In order to provide a comparative analysis of the genetic structure of the fig species present in Taiwan throughout their range, we had to develop a unique set of genetic markers that would be portable among any Ficus species on a worldwide basis. We developed a set of 20 highly portable microsatellite primers. The markers were both easy to amplify, and easy to read in 24 Ficus species from the six subgenera occurring in Taiwan. (article 1). This will facilitate our investigation of the genetic structure of complete regional floras using a single set of marker.
During the period allotted to this thesis we were able to examine the following questions :
1) What is the spatial genetic structuration of four common species of figs within Taiwan? In this part, we will test the hypothesis that even in highly dispersive species such as Ficus, insular conditions may result in niche enlargement and facilitate incipient speciation, as evidenced by strong genetic structuring within Ficus species (article 2).
2) How did the processes of colonization take place in this newly emerged island?
Through phylogeographic studies, we will contrast a series of species representing what we hypothesise to be a gradient in dispersal capacity (article 3) in order to investigate the degree of isolation of Taiwan comparatively to the regions the fig trees came from. We also wanted to analyze the differentiation between islands and on the continent, and also for one species present both on the Continent and in the
21
Philippines, whether Taiwan had been colonized from one single origin or whether populations were of mixed origin.
3) We will then analyse how much of the plants reproductive phenology is constrained by their biogeographic origin (article 4). For that purpose we will contrast the growth and flowering and fruiting phenology of species for which we have analysed (or are in the process of analysing) genetic structuring. This study will provide important insights into whether flowering phenology could limit northward expansion of species originating from the Philippines and vice-versa.
4) Our goal at the beginning of this thesis was to provide a unique set of data on wasp community (pollinating and non-pollinating fig wasps) composition including data on the ecology of the species, and to combine this data with knowledge on the phylogenetic origin of the different species. The idea was to answer simple but often unanswered questions on the origins of community complexity, such as the importance of species redundancy (direct competition) versus ecology niche differentiation (processes allowing to limit competition) processes. During this study we, in fact, collected a large set of samples that will allow to perform such a study (500 vials, each vial containing the wasps of 10 figs) but the wasp identifications and data analysis are still in process and will not be presented here.
However, we present here our results on the interaction between ants and one monoecious Ficus species. Indeed while there is strong evidence on dioecious fig species suggesting that figs wasps, pollinators and NPFW are a sufficient resource that ensures colonisation of male trees by ants, no such evidence is available for monoecious species. Indeed all published data suggests that ants on monoecious figs are always dependent on sap sucking insects for their subsistence (Dejean et al., 1997;
Zachariades et al., 2009). We therefore carefully investigated figs on ants in Taiwan.
22
While there is ongoing work within the team on ants on Ficus benguetensis, a dioceious species, we developed during the PhD work new insights into ant-fig interactions brought about by this discovery of a new case of ants inhabiting structures in a fig tree. The situation was all the more surprising as the fig tree involved is Ficus subpisocarpa, a monoecious species (article 5).
References:
Algar AC and Losos JB (2011) Evolutionary assembly of island faunas reverses the classic island-mainland richness difference in Anolis lizards. Journal of biogeography 38:1125-1137.
Badeck F-W, Bondeau A, Böttcher K, Doktor D, Lucht W, Schaber J and Sitch S.
(2004) Responses of spring phenology to climate change. New Phytologist 162:295-309.
Barnaud A, Deu M, Garine E, McKey D, and Joly H (2007) Local genetic diversity of sorghum in a village in northern Cameroon: structure and dynamics of landraces.
Theoretical and Applied Genetics 114:237-248.
Berg CC (1989) Classification and distribution of Ficus. Experimentia 45:605-611.
Berg CC (2011) Corrective notes on the Malesian members of the genus Ficus (Moraceae). Blumea 56: 161-164.
Berg CC, Pattharahirantricin N, and Chantarasuwan P (2011) Moraceae. Flora of Thailand, Vol 10, pp 475-675.
Brouat C, Loiseau A, Kane M, Bâ K, and Duplantier J-M (2007) Population genetic structure of two ecologically distinct multimammate rats: the commensal