Changes of protein expression in testes of L2 strain Taiwan country
chicken in response to acute heat stress
Shih-Han Wang1, Chung-Yu Cheng1, Chao-Jung Chen2, Hsin-Hsin Chen3, Pin-Chi
Tang1,4,5, Chih-Feng Chen1,4,5, Yen-Pai Lee1, H. Sunny Sun6, and San-Yuan Huang1,4,5,7, *
1 Department of Animal Science, National Chung Hsing University, Taichung 40227,
Taiwan
2 Department of Medical Research, China Medical University, Taichung 40227,
Taiwan
3 Department of Veterinary Medicine, National Chung Hsing University, Taichung
40227, Taiwan
4 Agricultural Biotechnology Center, National Chung Hsing University, Taichung
40227, Taiwan
5 Center for the Integrative and Evolutionary Galliformes Genomics, iEGG Center,
National Chung Hsing University, Taichung, 40227, Taiwan
6 Institute of Molecular Medicine, National Cheng Kung University, Tainan 701,
Taiwan
7 Center of Nanoscience and Nanotechnology, National Chung Hsing University,
Taichung, 40227, Taiwan.
*Corresponding author:
Dr. San-Yuan Huang, Department of Animal Science, National Chung Hsing University, 250 Kuo-Kuang Road, Taichung 40227, Taiwan
E-mail: [email protected] 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Tel: +886-4-22870613 ext. 245 1
Abstract
Heat stress causes a decrease of fertility in roosters. Understanding the molecular response to heat stress in avian testes will benefit the improvement of thermotolerance in roosters. Nevertheless, the protein expression in response to acute heat stress is still limited. The purpose of this study was to investigate the differential protein expressions in testes of L2 strain Taiwan country chicken after acute heat stress. Twelve 45-wk-old roosters were allocated into four groups, including control roosters kept at 25 °C, roosters subjected to 38 °C acute heat stress for 4 h without recovery, with 2 h recovery, and with 6 h recovery, respectively. Testis samples were collected for morphological assay and protein analysis. The morphology and apoptosis of testes after heat stress were evaluated by microscopy and TUNEL assay. The differential protein expressions in chicken testis after heat stress, the testes from control group and heat stress group were analyzed by two-dimensional difference gel electrophoresis. Some of the differentially expressed proteins were validated by Western blotting and immunohistochemistry. The results showed that abnormal and apoptotic spermatogenic cells increased quickly at 2 h of recovery after acute heat stress, especially for the spermatocytes. Two-dimensional difference gel electrophoresis revealed that there were 119 protein spots representing 92 distinct proteins differentially expressed in chicken testis after heat stress. Gene ontology analysis showed that these proteins majorly participated in protein folding and proteolysis. The heat shock proteins, chaperonin containing t-complex, and proteasome subunits were downregulated and glutathione S-transferanse, transgelin, and DJ-1 were upregulated in chicken testis after heat stress, respectively. The results of this study revealed that acute heat stress impaired the processes of translation, protein folding, and protein degradation and thus results in apoptosis and destructure 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
of spermatogenesis. On the other hand, the expression of antioxidant enzymes that attenuate heat induced damage, including glutathione S-transferase and DJ-1, increased. The mechanism of acute heat stress response in avian testis can be the basic foundation to the future study.
Keywords: Chicken testis; Acute heat stress; Protein expression; 2-D difference gel electrophoresis (2-D DIGE)
1. Introduction
In tropical and subtropical areas, including Taiwan, elevated temperature along with high humidity in summers affects semen quality and result in decreasing reproductive performance of male chickens [1,2]. Even though the effects of heat stress on cocks were more serious than on hens, the studies majorly focused on the direct effect of heat stress on layers or broilers [2-4]. Although Taiwan country chickens (TCCs) show better thermotolerance than that of foreign breeds [5], semen quality and fertility of male TCCs also decreased in summer [6] and after treatment [7].
Two-dimensional gel electrophoresis (2-DE) has been applied to investigate the global protein expressions in biological samples [8,9]. Nevertheless, the gel-to-gel variation seems a challenge for quantitative comparison by 2-DE [10]. Thus, 2-D difference gel electrophoresis (2-D DIGE) has been established as an improvement method for protein quatification to avoid gel-to-gel variation [11,12]. Comparative proteomics by 2-D DIGE has been used to explore male infertility in human and mouse [13,14].
The operating temperature of the testis is equivalent to the core body temperature 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
(40-41 °C) in chickens; therefore, there may be compensatory or alternate mechanisms that allow for normal spermatogenesis in domestic fowl [15]. The global gene expression analysis showed that some genes associated with thermotolerance were differentially expressed in the testes of L2 strain of TCC after acute heat stress [16]. It has been shown that proteins associated with the processes of spermatogenesis and spermiogenesis were differentially expressed by modulating the expression of proteins related to proliferation, differentiation, apoptosis and cell survival pathways in heat-stressed mouse and human testis [8,9]. However, the global protein expression in chicken testes in response to acute heat stress is still undefined. This study investigated the effect of acute heat stress on protein expressions in the testes of TCC by 2D-DIGE and mass spectrometer as base for exploring the mechanism of acute heat stress response in chickens.
2. Materials and methods
2.1. Experimental animals
A total of 12 45-wk-old roosters of L2 strain TCC selected by National Chung Hsing University were used in this study. L2 strain TCC is selected for egg production [17]. The experiments were reviewed and approved by The Institutional Animal Care and Use Committee (IACUC) of National Chung Hsing University, Taichung, Taiwan. The roosters were kept in climate chamber for more than 2 weeks as an adaptation period [L:D=14:10, ad libitum, 25°C, 55% relatively humidity (RH)].
2.2. Condition for acute heat stress and sample collection
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
The roosters were allocated into four groups (three roosters in each group) after the adaptation period. The control group was remained at 25 °C and 55% RH. The other three groups included roosters received an acute heat stress at 38 °C and 55% RH for 4 h without recovery (R0), recovered at 25 °C for 2 h (R2), and recovered at 25 °C for 6 h (R6). After recovery, roosters were sacrificed. The testis samples were collected. The samples for 2-D DIGE analysis were immersed in liquid nitrogen immediately for further 2-D DIGE analysis. The samples for histological analysis were slide into small pieces and fixed in Bouin’s solution for 24 h and the transferred to 70% ethanol before preparing paraffin-embedded blocks [18]. The testis samples were then dehydrated and embedded in paraffin for preparing tissue sections for further analysis.
2.3. Sample preparation for 2-D DIGE analysis
The procedure for preparating soluble protein of testes is followed previous method [19]. Briefly, testis samples were homogenized in homogenization buffer (0.3M sucrose, 0.5M Tris/HCl pH6.8, 1.67 mM Pefabloc SC PLUS, and protease inhibitor cocktail tablet) at 4°C. Homogenate was collected and centrifuged at 4°C (12,000×g for 5 min) and the supernatant was further centrifugated (90,000×g) at 4°C for 3 h. The supernatant was dialyzed against 100 mM ammonium bicarbonate at 4°C overnight. Protein concentration was determined by modified Bradford assay using bovine serum albumin as a standard [20]. For 2-D DIGE, every individual testis sample was pooled together according their treatment. Briefly, equal amount protein of three testis samples from the same group were pooled together and then dried by speed vacuum (EYELA CVE-200D, Tokyo, Japan). All protein samples were resolved in 2-DE lysis buffer (4% w/v CHAPS, 7 M urea, 2 M thiourea, 10 mM Tris-1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
HCl, pH8.3, 1 mM EDTA) for 2-D DIGE analysis.
2.4. 2-D DIGE and Image analysis
The procedure of 2-D DIGE analysis followed the method of Lin et al. [21]. Protein samples were labeled with N-hydroxy succinimidyl ester-derivatives of the cyanine dyes Cy2, Cy3 and Cy5 (GE Healthcare, Uppsala, Sweden). In brief, 100 µg protein was labeled with 250 pmol of either Cy3 or Cy5 for comparison on the same gel. In order to facilitate image matching and cross-gel statistical comparison, a pool of all samples in each condition was labeled with Cy2 (2.5 pmol/ μg protein) as an internal standard for all gels. The labeling reactions were performed in dark on ice for 30 min and then stopped with excess of free L-lysine for 10 min. The differentially Cy3-, Cy5-, and Cy2-labeled samples were mixed well, and the distribution of samples is listed in Table 1. All samples were reduced with dithiothreitol (DTT) for 10 min, and IPG buffer, pH 3–10 nonlinear (2% (v/v), GE Healthcare) was added. The final volume was subjected to adjust to 350 µl with 2D-lysis buffer for isoelectric focusing (IEF). The immobilized non-linear pH gradient (IPG) strips (pH 3–10, 24 cm) were rehydrated by CyDye-labeled samples in dark for at least 12 h, and IEF was carried out using a Multiphor II apparatus (GE Healthcare) for a total of 62,500 Vhr at 20 °C. Strips were equilibrated in equilibration buffer [6 M urea, 30% glycerol, 1% SDS (w/v), 100 mM Tris–HCl (pH 8.8)] containing 65 mM DTT for 15 min, followed by equilibrating in equilibration buffer with 240 mM iodoacetamide for another 15 min. The equilibrated strips were layered on top of 12.5% SDS-polyacrylamide gels and sealed with 0.5% (w/v) low melting point agarose containing bromophenol blue. For the second dimension separation, electrophoresis was performed at 10°C with a condition of 2 watt per gel until the dye front completely run off the bottom of the 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
gels by using the Ettan Twelve gel tank (GE Healthcare). After electrophoresis, the gels between low fluorescent glass plates were scanned by using an Ettan DIGE Imager (GE Healthcare). Protein spots on all gels were detected, normalized, and analyzed using DeCyder 2-D Differential Analysis Software v7.0 (GE Healthcare). Spots with 1.5 average fold-change in abundance as well as P value < 0.05 were selected to further protein identification.
2.5. Gel stain
The method of gel stain followed the procedure of Wu et al. [12]. Briefly, gels were fixed in fix solution (30% ethanol and 2% phosphoric acid) at room temperature overnight. After washing with ddH2O for 30 min three times, gels were incubated in
staining solution (34% methanol, 17% ammonium sulfate, 3% phosphoric acid) for 1 h and then added 0.5 g/L colloidal Coomassie blue G-250. The gels were stained on shaker for 3 days.
2.6. In-gel digestion
Differentially expressed protein spots were excised from the stained gels for further in-gel digestion [12]. Gel spots were washed in 50% acetonitrile with 10 mM ammonium bicarbonate three times and then dehydrated in pure acetonitrile. Gel spots were then dried by speed vacuum and subject to in-gel digestion. For in-gel digestion, gel plugs were reswollen with 20 ng/µl trypsin (Promega, Madison, WI, USA) in 10 mM ammonium bicarbonate at 4°C for 30 min. The gel plugs were subject to overlay with 10 mM ammonium bicarbonate, and digestion was allowed to proceed for 16 h at 37°C. After digestion, peptide extracts were recovered by 50% ACN and 1% TFA 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
with sonication twice. Peptide extracts from same gel plug were pool together and stored at -20°C for mass spectrometery analysis.
2.7. Protein identification
Matrix-assisted laser desorption/ionization time-of-flight mass spectrometer (MALDI-TOF MS) analysis followed the procedure of Lu et al. [22] with some modification. Digested samples were spotted directly onto a 600um/384well AnchorChip sample target (Bruker Daltonics, Bremen, Germany), and then add with an equal volume of 1mg/ml solution of alpha-cyano-hydroxycinnamic acid in 0.1% TFA/50% ACN. MALDI mass spectra were obtained using a Bruker UltraFlex TOF mass spectrometer equipped with a 384 sample Scout source (Bruker Daltonics). An external peptide calibration standard containing Angiotensin II ([M+H]+ 1046.54), Angiotensin I ([M+H]+ 1296.68), Substance P ([M+H]+ 1347.74), Bombesin ([M+H]+ 1619.82), ACTH clip 1-17([M+H]+ 2093.09), ACTH clip 18-39([M+H]+ 2465.20) and Somatostatin 28 ([M+H]+ 3147.47) (Bruker Daltonics) were used to calibrate the instrument. Spectra were acquired in reflectron mode. Masses were processed using the FlexAnalysisTM 2.4 software (Bruker Daltonics). Peptide masses were searched
against a comprehensive nonredundant protein sequence database (NCBInr 20130524 version with 25805290 sequences and 8915431356 residues; SwissProt 2013_05 version with 540052 and 191770152 residues) employing BioTools 3.0 software (Bruker Daltonics) in combination with the Mascot program for protein identification. The search conditions were taxonomy of bony vertebrates, fixed modification of carbamidomethyl modification, variable modifications of oxidation modification, trypsin as enzyme, peptide mass tolerance 100 ppm, and significant threshold P < 0.05. The protein identification was further confirmed by MALDI TOF/TOF MS/MS 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
analysis.
The AnchorChip target was subjected to acquire MALDI TOF/TOF spectra, using a Bruker UltraFlex III MALDI-TOF/TOF MS (Bruker Daltonics) equipped with a delayed extraction ion source. Metastable ions generated by laser induced decomposition (LID) in the LIFT mode (Bruker Daltonics) were analyzed. The precursor ion and the corresponding fragment ions were selected in a time gate followed by further acceleration in the LIFT cell with 19.0 kV. The fragment ions were accelerated into the second field-free region and separated in the two-stage gridless reflectron. The reflectron voltage were set at 27.4 kV. Masses were processed using the FlexAnalysisTM 2.4 software (Bruker Daltonics). The proteins were
identified by searching MS/MS spectra against NCBInr database (NCBInr 20130524 version with 25805290 sequences and 8915431356 residues; SwissProt 2013_05 version with 540052 and 191770152 residues) using BioTools 3.0 software (Bruker Daltonics) in combination with the MASCOT program. The search criteria were as follows: taxonomy for bony vertebrates, fixed modification of carbamidomethyl modification, variable modifications of oxidation modification, 100 ppm for precursor ion, and 0.5 Da for fragment ions. Positive identification was achieved with the score whose higher than minimal significant individual ion score (P<0.05).
2.8. Bioinfomatic analysis
The differentially expressed proteins among the treatments were annotated for their subcellular distribution, biological processes, and molecular function by Gene Ontology (GO) database (http://www.geneontology.org/). The unique protein names were uploaded and searched for their cellular component and molecular function. To simplify the classification, the second or third level of tree browser in molecular function was used.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
2.9. Hematoxylin–eosin staining
Bouin-fixed and paraffin-embedded blocks of testis tissues were sectioned, deparaffinized, and rehydrated with EZ-DeWax™ (HK584-5K, BioGenex, San Ramon, CA, USA) following the manufacturer's recommendations. Sections were stained with hematoxylin and eosin and examined by Olympus BX51 Microscope and DP70 Digital Camera System (Olympus, Shinjuku-ku, Tokyo, Japan).
2.10. Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay
The TUNEL assay for identifying apoptotic cells were performed using the In
Situ Cell Death Detection Kit, POD (Anti-fluorescein antibody, Fab fragment from
sheep, conjugated with peroxidase) (Roche, Germany). Paraffin-embedded testis tissues were sectioned, deparaffinized, and rehydrated with EZ-DeWax™. The sections were then subjected to TUNEL assay according to the standard protocol recommended by demonstration of the kit.
2.11. Western blotting
The procedure of Western blotting was performed according to previous study [23]. Soluble proteins from chicken testes were lysed in sample buffer (62.5mM Tris-HCl (pH6.8), 2% SDS, 10% glycerol, 5% β-mercaptoethanol, and 0.002% bromophenol blue) and separated by 10% SDS-PAGE. After electrophoresis, proteins were transferred to nitrocellulose membrane (Amershan Biosciences AB, Uppsala, Sweden). The membranes were blocked in TTBS (20 mM Tris-HCl (pH7.4), 0.5 M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
NaCl, and 0.05% Tween 20) containing 3% gelatin for 1 h. Then the membranes were immunoblotted with a 1:10000-diluted anti-heat shock protein 90 α (HSP90α) mouse monoclonal antibody (NB110-96872, Novus Biologicals, Littleton, CO, USA) for 1 h, a 1:500-diluted anti-polyadenylate-binding protein 1 (PABPC1) rabbit polyclonal antibody (ABE40, Millipore, Temecula, CA, USA) overnight, a 1:1000-diluted anti-chaperonin containing t-complex β (CCTβ) rat monoclonal antibody (ADI-CTA-202, EnZo Life Science, Farmingade, NY, USA) overnight, a 1:400-diluted anti-transgelin (TAGLN) rabbit polyclonal antibody (ab14106, Abcam, Cambridge, MA, USA), a 1:200-diluted anti-heat shock protein 25 (HSP25) mouse polyclonal antibody (sc-51956, Santa Cruz Biotechnology, Inc, Dallas, Texas, USA) or a 1:10000-diluted anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mouse monoclonal antibody (NB300-221, Novus Biologicals, Littleton, CO, USA) overnight. After washing in TTBS three times (5 min each), the membranes were incubated with 1:5000-diluted goat anti-mouse, goat anti-rabbit, or goat anti-rat IgG conjugated with alkaline phosphatase (Sigma, St. Louis, MA, USA) for 1 h matching the primary antibody used. Then, the membranes were washed in TTBS for four times and subsequently developed using a buffer containing nitro blue tetrazolium and 5-bromo-4-chloro-3-indoyl phosphate (BioRad, Hercules, CA, USA). The density of immunoblotted protein bands were determined by Ultra-Lum TotalLab software (Nonlinear Dynamics Ltd., Newcastle, NE1, UK). The expression levels of protein were obtained by normalized density of protein bands to the density of GAPDH band.
2.12. Immunohistochemistry
Bouin-fixed and paraffin-embedded blocks of testis tissues were sectioned, deparaffinized, and rehydrated with EZ-DeWax™. The testis sections were boiling-1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
heated in sodium citrate buffer (0.05% Tween 20 in 0.01 M sodium citrate, pH 6.0) for 10 min to enhance antigen retrieval. Endogenous peroxidase activity was quenched by 3% H2O2 for 10 min, and then nonspecific binding was block by super
block solution (ScyTek Laboratories, West Logan, UT, USA) for 20 min. Sections were incubated with primary antibodies against target proteins. In brief, sections were incubated with antibody against HSP90α (NB110-96872, Novus Biologicals; dilution 1:500) at room temperature (RT) for 1 h, TAGLN (ab14106, Abcam, Cambridge, MA, USA; dilution 1:200) at RT for 1 h, PABPC1 (ABE40, Millipore, Temecula, CA, USA; dilution 1:25) at 4°C overnight, CCTβ (ADI-CTA-202, EnZo Life Science, Farmingade, NY, USA; dilution 1:200) at RT for 1 h. After washing in PBST (0.05% Tween 20 in PBS) 5 min for three times, sections were subject to incubate with horseradish peroxidase (HRP) conjugated anti-rabbit IgG (Millipore, Temecula, CA, USA), anti-mouse IgG, or anti-rat IgG (GeneTex, InC. San Antonio, Texas, USA) for 30 min. After washing with PBST for three times, positive cells were stained as brown color using 3,3′-diaminobenzidine as the substrate. The nuclei were stained with hematoxylin. To confirm the specificity of antibodies, negative controls were performed by the same procedure, except for that the primary antibody was replaced by mouse, rat, or rabbit IgG (Vector Laboratories, Burlingame, CA, USA).
2.13. Statistical analysis
The relative values of proteins on 2D-DIGE gels or on membranes of Western blotting were analyzed using the t-test procedure in Statistical Analysis System software [24]. 3. Results 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
3.1. Morphology and apoptosis of chicken testis after acute heat stress
To investigate the effect of acute heat stress on the testis protein expression of L2 strain TCCs, chickens were treated with 38°C for 4 h and then recovered at 25°C for 0 h, 2 h, and 6 h. Observation of the testicular cell morphology showed that several multinucleated giant cells and spermatocytes with hyperchromatic nuclei at 2 h of recovery after heat exposure (Figure 1, panel C). However, there was no significant change at 6 h of recovery after heat stress. Results of TUNEL assay also showed that TUNEL-positive cells were only increased at 2 h of recovery after heat stress, and most of them were spermatocytes (Figure 2).
3.2. Comparison of protein profiles and identification of differentially expressed proteins in chicken testis after acute heat stress
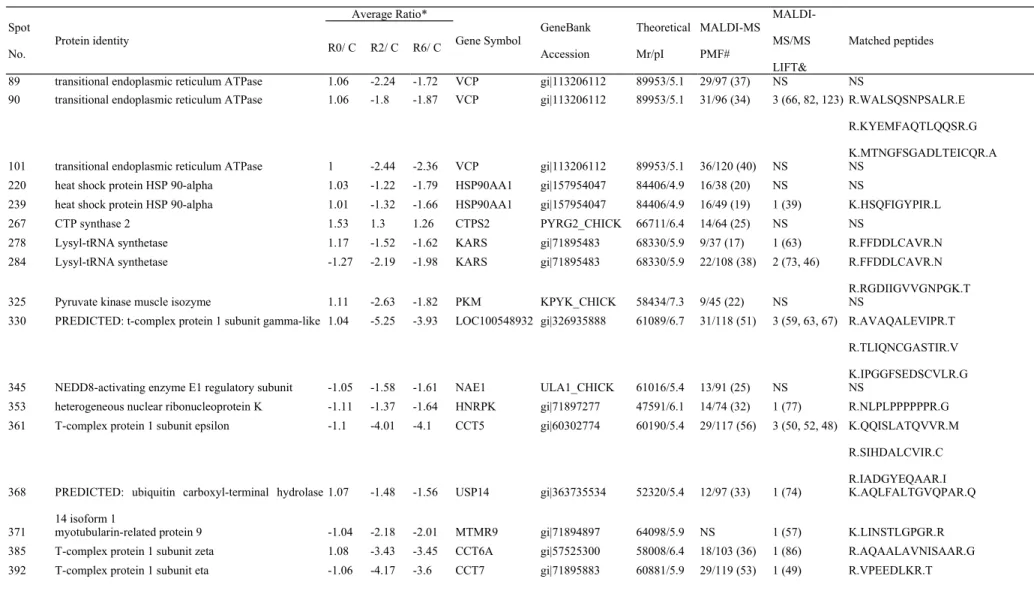
Chicken testes from heat-stressed and non-heat stressed chickens were used to evaluate the effect of acute heat stress on the global change of protein expression. More than 1,500 protein spots were detected on 2D-DIGE protein profiles. Comparing the control group, there were 119 protein spots differentially expressed (≧1.5-fold; P < 0.05) in the heat-stressed groups (Figure 3). The identities of the differentially expressed protein spots were indentified by MALDI-TOF and MALDI-TOF/TOF (Table 2). These proteins represented 92 distinct proteins.
3.3. Bioinformatic analysis of the differentially expressed proteins
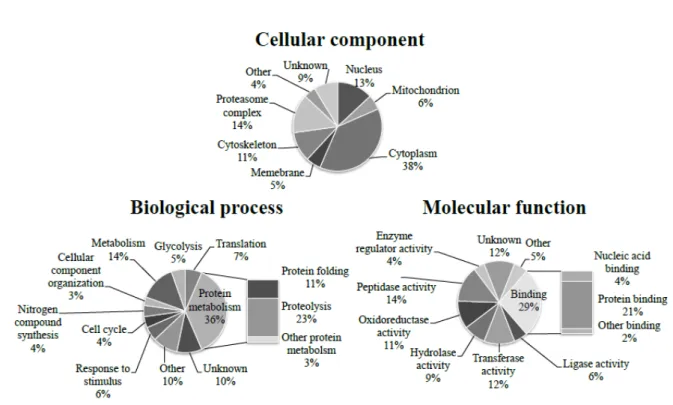
The differentially expressed proteins were mostly localized in cytoplasm (38%), 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
proteasome complex (14%), and nucleus (13%), while some of differentially expressed proteins were localized in cytoskeleton (11%), mitochondrion (6%), and membrane (5%) (Figure 4). These proteins majorly took part in metabolism (14%), protein folding (11%), and proteolysis (23%), and most of them, such as heat shock protein (HSP) 90-alpha (HSP90α), chaperonin containing t-complex (CCT), and subunits of protease complex, were downregulated (P<0.05) in testes of heat-stressed chickens (Figure 4, Table 2).
3.3. Validation of protein expression in heat-stressed chicken testis
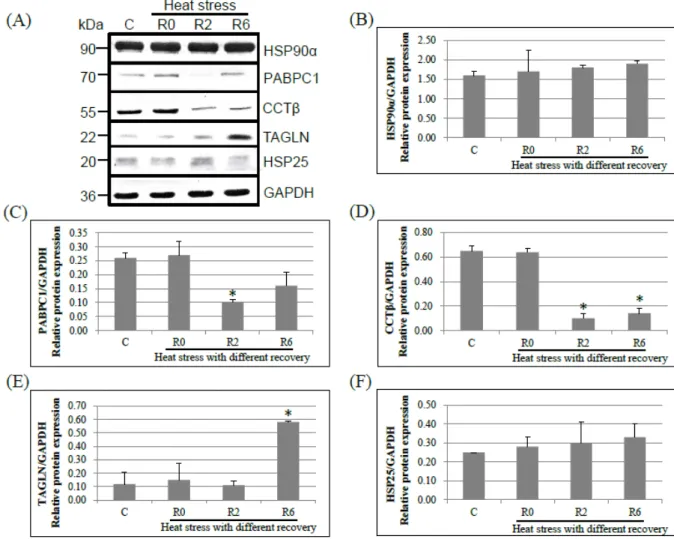
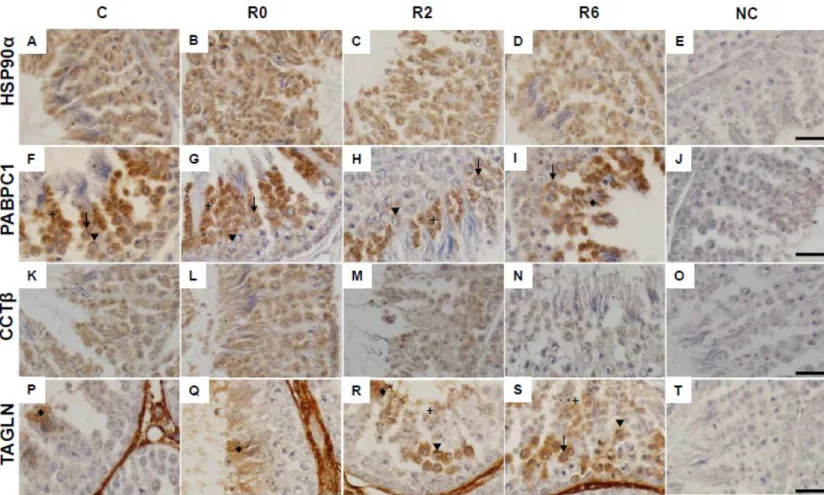
To validate the expressions of differentially expressed proteins identified by 2D-DIGE, Western blot analysis was performed to detect some important proteins including HSP90α, transgelin, CCT-β, PABPC1, and HSP25. The results revealed that the expression of CCT-β, PABPC1, and TAGLN showed the same pattern with the observation in 2D-DIGE analysis (Figure 5). However, the protein level of HSP90α and HSP25 were not different among treatment by the immunoblot analysis. The locations of protein expression, including HSP90α, transgelin, CCT-β, and PABPC1, in chicken testis were defined by using IHC (Figure 6). The HSP90α expressed in all spermatogenic cells in testes of control and heat-stressed chickens (Figure 6, A to D). The CCTβ also expressed in all spermatogenic cells in testes of non heat stressed chickens, but its level reduced in all spermatogenic cells after heat stress (Figure 6, K to N). In testes of control chickens, PABPC1 was expressed in secondary spermatocytes and round spermatids, and a lower level in primary spermatoctyes. The expression of PABPC1 was reduced in spermatocytes and round spermatids at 2 h of recovery after heat stress, and it was increased in elongated spermatids at 6 h of recovery (Figure 6, F to I). TAGLN was strongly expressed in the 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
basement membrane and interstitial cells and also expressed in the elongated spermatids of chicken testes. After 2 h of recovery, the expression increased in primary spermatocytes in the testes of heat-stressed chickens. The expression of TAGLN in spermatocytes and round spermatids were also increased at 6 h of recovery after heat stress (Figure 6, P to S).
4. Discussion
4.1. The change of morphology and protein levels in chicken testis responded to acute heat stress
The molecular mechanism of the effect of acute heat stress on male fertility in chickens is still not completely understood. Our previous study analyzed the global mRNA expression in response to acute heat stress in the testis of L2 strain TCCs [16]. The present study further analyzed the testicular morphology and protein expression in heat-stressed chicken testes. The results showed that spermatocytes were more sensitive to acute heat stress compared to other germ cells (Fig. 2) and it was consistent with observations in previous studies [25,26]. The apoptosis accompanied with autophagy in mouse testes significantly increased at 12 h after heat stress and leads to destruction of spermatogenesis [25]. The initiation of apoptosis in chicken testis occurred earlier than that in mouse during recovery after heat stress (Fig. 2). The result implicated that heat stress induces apoptosis very quickly and may thus destruct spermatogenesis in chicken testes. The result of microarray analysis showed that the expression of 309 genes altered at transcription level in chicken testes after acute heat stress [16]. Nevertheless, only 92 distinct proteins changed in heat-stressed chicken testes during recovery. Most of the differentially expressed proteins were 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
downregulated after heat stress. When mouse testes are treated with 43°C for 15 min for 5 days, the unfolded protein response leads translation arrest and ultimately apoptosis in spermatocytes [26]. The unfolded protein response is considered to induce both cell survival and apoptosis when unfolded proteins accumulated under endoplasmic reticulum stress in mammals [27,28]. During the unfolded protein response, transcriptional induction of chaperones is increased and thus the protein degradation and protein folding are enhanced to dissolve misfolded or unfolded proteins. At the same time, the translation is suppressed to prevent further unfolded protein accumulation and preserve energy [27,28]. However, once protein-folding defect could not be resolved by protein degradation and protein folding, the unfolded protein response is turn to apoptosis for protecting the organism [28]. We inferred that the unfolded protein response, including increased transcription and halting translation, was also happened in the testes of heat-stressed chickens. However, the homeostasis was destructed and thus resulting in apoptosis of the chicken testicular cells.
After acute heat stress, the protein expression of glutathione S-transferase increased in chicken testis as well as the mRNA expression observed in previous report [16]. However, the protein expression of HSP90α, HSP25, CCTβ, PABPC1, and subunits of proteasome deceased in heat-stressed chicken testes while the mRNA expressions of these genes increased or not changed after acute heat stress. The mRNA expression of TAGLN in chicken testes decreased at 0 and 2 h after heat stress [16], but the protein expression increased at 6 h of recovery after heat stress. There were inconsistency in the expression of some genes in chicken testes between the transcription and translation levels in response to acute heat stress. The TAGLN, also called smooth muscle protein of 22kDa, is an actin binding protein and as is a marker of differentiation in smooth muscle [29]. It has been reported that mRNA level of 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
TAGLN increased in senescent fibroblasts or stress-induced premature senescence
fibrobalsts [30-32]. In various tumor cell lines, overexpression of TAGLN significantly inhibited cell proliferation and accelerated apoptosis while depletion of TAGLN promoted cell survival [33,34]. Although the mRNA expression of TAGLN declined at 0 h and 2 h of recovery in heat-stressed chicken testis [16], the protein level of TAGLN increased in spermatocytes and round spermatids at 2 h and 6 h of recovery after heat stress (Table 2, Figure 5). It seems that elevated protein level of TAGLN in chicken testes might enhance apoptosis and interfere spermatogenesis.
4.2. Protein biosynthesis is impaired by heat stress
Protein biosynthesis is a complex process for cellular homeostasis, cell generates new protein by transcription and translation while loss of protein via degradation and export [35]. Although the transcription is not impaired in heat-stressed chicken testis [16], we found that proteins associated with translation and protein degradation were reduced after heat stress, involving PABPC1, lysyl-tRNA synthetase, aspartyl-tRNA synthetase, eukaryotic translation elongation factor 1 gamma, elongation factor 1-alpha 1, 40S ribosomal protein S12, protein disulfide-isomerase A3 precursor, and subunit of proteasome (Table 2). The function of PABPC1 is to regulate initiation of translation and stabilize mRNA by binding to the poly(A) tail of mRNAs, and it is prominent expressed in spermatocytes and round spermatids [36,37]. The decline of PABPC1 suppresses translation induced by proteasome inhibitor in apoptotic cells [38]. Overexpression of PABPC1 protects cells against oxidative stress by inhibiting apoptosis signaling pathway [39]. The PABPC1 also expresses in the cytoplasm of spermatogenic cells and can enhance translation in mouse testes [40]. Heat stress causes a reduction of PABPC1 in chicken testes, this may imply that translation is 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
inhibited in spermatocytes and round spermatids (Figure 6F to 6I). Lysyl-tRNA synthetase and aspartyl-tRNA synthetase are the aminoacyl-tRNA synthetases that attach amino acid onto their cognate tRNAs to form an aminoacyl-tRNA during protein synthesis [41]. The subunits of elongation factor-1 (α, β, γ, and δ) interacts aminoacyl-tRNA synthetases and transfer aminoacyl-tRNA to ribosome [42,43]. The nascent proteins which are formed from ribosomes are released into cytoplasm or endoplasmic reticulum for folding in the active structures. In cytoplasm, HSP90 works with other chaperones and co-chaperones governs protein folding [44]. A chicken homologue of human HSP27, HSP25, plays the roles in inhibition of actin polymerization and dissociation the misfolded protein complex protein aggreasomes [45,46]. In endoplasmic reticulum, the folding and quality control of glycoproteins are performed by protein disulfide-isomerase A3, a thiol-disulfide oxidoreductase that catalyzes the formation of disulfide bonds [47]. The inconsistency of mRNA and protein expression in chicken testes after acute heat stress may infer that acute heat stress causes a reduction of translation while the transcription is not interfered.
The proteasomes are protein complex that degrade misfolded proteins through ubiquitin–proteasome system (UPS) or ubiquitin–independent degradation in eukaryotes [48,49]. The process of UPS is mediated by many factors, including valosin-containing protein (VCP) and reactive oxygen species (ROS) [50-52]. Interfering VCP inhibits the function of UPS and increases apoptosis [53,54]. In this study, heat stress downregulated proteasome subunits and VCP, and thus might cause a reduction of protein degradation. Proteasomes have been detected at sperm tail connecting piece and acrosomal region in bull spermatozoa, and proteasomes are related to acrosomal reaction, capacitation process, and zygotic development [55]. However, some of the proteasome subunits are suggested to correlate with subfertility and expressed high abundantly in low fertility bull and human sperm [56,57]. On the 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
other hand, several studies reported that inhibition of proteasome alone or with HSP90 inhibitor causes an increase of apoptosis and an activation of autophagy in cancer cells [58-61]. Furthermore, heat stress induces spermatogenic cell death via apoptosis and autophagy in mouse testes [25]. In this study, it showed that the expressions of proteasome subunits and HSP90 were downregulated at 2 h and 6 h of recovery in heat-stressed chicken testes (Table 2). The observations suggest that the reduction of proteasomes may result in cell death through apoptosis accompanied with autophagy to clean abnormal cells in chicken testis after acute heat stress.
4.3. Acute heat stress causes a dramatic decrease of chaperonin containing t-complex
The chaperonin containing t-complex (CCT complex) is a eukaryotic chaperone containing eight subunits (CCTα, CCTβ, CCTγ, CCTδ, CCTε, CCTζ, CCTη, and CCTθ). The CCT complex assists the folding of cytoskeleton proteins such as actin and tubulin which are involved in sperm motility and cell junction [62-67]. The CCT complex participates in cell cycle, cell growth, and post-translational modification [68-69]. In male mouse germ cells, the CCT complex is localized at the centrosome, manchette, and condensing chromatin and is associated with spermatogenesis [70]. In human, mouse, and bull spermatozoa, CCT complex is localized on the surface of spermatozoa and participates in sperm-zona pellucida interaction in an indirectly way by mediating zona pellucida receptor [71-73]. The upregulated CCT complex plays an important role in the recovery of mammalian cell lines from protein damage that caused by stress [74]. However, the expression levels of CCT subunits have different functions. The CCTβ, one of the t-complex proteins in germ cells, is declined after ischemia-reperfusion of the mouse testis, and mutation of CCTβ in yeast exhibits heat sensitivity and cold sensitivity [75,76]. High levels of 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
CCTε and CCTθ were found in low fertility Holstein bulls and implicated incomplete spermiogenesis [56]. Moreover, cells with the CCTη mutation result in impaired stress
response [75].Result of this study revealed that CCT complex is downregulated at 2 h of recovery after heat stress in chicken testes (Table 2, Figure 5, Figure 6 K to N), and it may implicate that spermatogenesis may be interfered through disrupting cell cycle of spermatogenetic cells.
4.4. Antioxidant enzymes is upregulation after heat stress
Oxidative stress resulting from elevation of ROS can be induced by many factors, including heat stress, radiation, and ischemia [77-79]. In male mouse, scrotal hyperthermia carried out in water bath at 43°C for 15 min induces antioxidant enzymes to protect testicular cells from oxidative stress [80]. However, the testicular antioxidant system is diminished in apoptotic germ cells when rat testes are immersed in water bath at 43°C for 30 min once daily for 6 consecutive days [77]. In the present study, 4 h of heat stress also caused an increase of antioxidant enzymes, including glutathione S-transferase [80] and DJ-1 [81], in chicken testes (Table 2). One of the roles of DJ-1 is to act as an antioxidant enzyme and responses to oxidative stress [81,82]. Upregulation of DJ-1 protects cells against cell death from various stresses in mouse and chicken cell lines [83,84]. This is the first observation that DJ-1 upregulated in heat-stressed chicken testis. These results suggest that the upregulation of antioxidant enzymes like, including glutathione S-transferase and DJ-1 may attenuate heat-induced oxidative stress and cell death in chicken testes.
4.5. Possible mechanism of testicular response to acute heat stress in L2 strain Taiwan country chicken
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
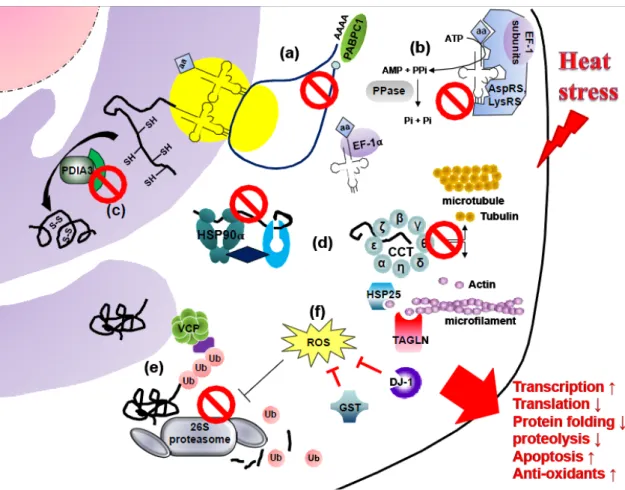
According to the findings of this study and the reported functions of differentially the expressed proteins, a possible model is proposed to elucidate the mechanism of acute heat stress response in chicken testes (Figure 7). Proteins associated with initiation of translation (PABPC1), tRNA aminoacylation (lysyl-tRNA synthetase, aspartyl-(lysyl-tRNA synthetase, eukaryotic translation elongation factor 1 gamma, elongation factor 1-alpha 1, and inorganic pyrophosphatase), disulfide bond formation (protein disulfide-isomerase A3 precursor), protein folding (HSP90α, HSP25, and CCT), and protein degradation (valosin-containing protein and proteasome complex) were downregulated after acute heat stress in chicken testes. The decrease of CCT impaired folding of actin and tubulin and followed then disrupted cell cycle. In addition, upregulated transgelin TAGLN after heat stress might inhibit cell proliferation. However, the increase of antioxidant enzymes (glutathione S-transferase and DJ-1) may diminish the injury caused by acute heat stress. The effect of acute heat stress on chicken testes included a decrease of protein translation and degradation as well as an enhancement of ROS production, thus resulting in accelerating apoptosis and interfering spermatogenesis. Meanwhile, antioxidant enzymes were increased to protect testicular cells against oxidative stress induced by heat stress.
5. Conclusion
Morphological observations of the present study showed that apoptosis was induced very quickly in chicken testes after acute heat stress. The analysis of protein expression revealed that there were 92 distinct proteins differentially expressed after 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
acute heat stress. The possible mechanism of acute heat stress response in chicken testes was inhibiting translation, protein degradation, and spermatogenesis. However, the antioxidant enzymes were elevated to attenuate the damage caused by acute heat stress.
Acknowledgements
The authors would like to thank the National Science Council (NSC99-2321-B-005-016-MY3) and the Ministry of Education (under the ATU plan) of the Republic of China for financial support of this research. Computational analyses and data mining were performed using the system provided by the Bioinformatics Core at the National Cheng Kung University, supported by the National Science Council, Taiwan.
1 2 3 4 5 6 7 8 9 10 11 12
References
[1] Karaca AG, Parker HM, Yeatman JB, McDaniel CD. Role of seminal plasma in heat stress infertility of broiler breeder males. Poult Sci 2002;81:1904–9.
[2] McDaniel CD, Bramwell RK, Wilson JL, Howarth B, Jr. Fertility of male and female broiler breeders following exposure to elevated ambient temperatures. Poult Sci 1995;74:1029–38.
[3] Mashaly MM, Hendricks GL, 3rd, Kalama MA, Gehad AE, Abbas AO, Patterson
PH. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult Sci 2004;83:889–94.
[4] Mujahid A, Yoshiki Y, Akiba Y, Toyomizu M. Superoxide radical production in chicken skeletal muscle induced by acute heat stress. Poult Sci 2005;84:307–14. [5] Yeh CC. Effect of acute heat stress on the blood characteristics of Taiwan country
chickens and broilers. J Chin Soc Anim Sci 1992;21:57–66 [in Chinese with English abstract].
[6] Lee, Y.P. The Taiwan country chicken. / National Chung Hsing University, Taichung, Taiwan; 1992.
[7] Yan, Q.C. Effect of temperature on semen characteristics and sperm heat shock protein 70 in males of Taiwan country chicken. Master thesis. Department of Animal Science, National Chung Hsing University, Taichung, Taiwan; 2001.
[8] Zhu YF, Cui YG, Guo XJ, Wang L, Bi Y, Hu YQ, et al. Proteomic analysis of effect of hyperthermia on spermatogenesis in adult male mice. J Proteome Res 2006;5:2217–25.
[9] Zhu H, Cui Y, Xie J, Chen L, Chen X, Guo X, et al. Proteomic analysis of testis biopsies in men treated with transient scrotal hyperthermia reveals the potential targets for contraceptive development. Proteomics 2010;10:3480–93.
[10] Wu WW, Wang G, Baek SJ, Shen RF. Comparative study of three proteomic quantitative methods, DIGE, cICAT, and iTRAQ, using 2D gel- or LC-MALDI TOF/TOF. J Proteome Res 2006;5:651–8.
[11] Azimzadeh O, Scherthan H, Sarioglu H, Barjaktarovic Z, Conrad M, Vogt A, et al. 1
2 3
Rapid proteomic remodeling of cardiac tissue caused by total body ionizing radiation. Proteomics 2011;11:3299–311.
[12] Wu CL, Chou HC, Cheng CS, Li JM, Lin ST, Chen YW, et al. Proteomic analysis of UVB-induced protein expression- and redox-dependent changes in skin fibroblasts using lysine- and cysteine-labeling two-dimensional difference gel electrophoresis. J Proteomics 2012;75:1991–2014.
[13] Yamakawa K, Yoshida K, Nishikawa H, Kato T, Iwamoto T. Comparative analysis of interindividual variations in the seminal plasma proteome of fertile men with identification of potential markers for azoospermia in infertile patients. J Androl 2007;28:858–65.
[14] Li E, Guo Y, Ning Q, Zhang S, Li D. Research for the effect of octylphenol on spermatogenesis and proteomic analysis in octylphenol-treated mice testes. Cell Biol Int 2011;35:305–9.
[15] Li C, Wang X, Wang G, Li N, Wu C. Expression analysis of global gene response to chronic heat exposure in broiler chickens (Gallus gallus) reveals new reactive genes. Poult Sci 2011;90:1028–36.
[16] Wang SH, Cheng CY, Tang PC, Chen CF, Chen HH, Lee YP, et al. Differential gene expressions in testes of L2 strain Taiwan country chicken in response to acute heat stress. Theriogenology 2013;79:374–82
[17] Yang KT, Lin CY, Huang HL, Liou JS, Chien CY, Wu CP, et al. Expressed transcripts associated with high rates of egg production in chicken ovarian follicles. Mol Cell Probes 2008;22:47–54.
[18] Rinchard J, Ciereszko A, Dabrowski K, Ottobre J. Effects of gossypol on sperm viability and plasma sex steroid hormones in male sea lamprey, Petromyzon
marinus. Toxicol Lett 2000;111:189–98.
[19] Huang SY, Lin JH, Chen YH, Chuang CK, Lin EC, Huang MC, et al. A reference map and identification of porcine testis proteins using 2-DE and MS. Proteomics 2005;5:4205–12.
[20] Bradford MM. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem
1976;72:248–54
[21] Lin ST, Chou HC, Chang SJ, Chen YW, Lyu PC, Wang WC, et al. Proteomic
analysis of proteins responsible for the development of doxorubicin resistance in human uterine cancer cells. J Proteomics 2012;75:5822–47.
[22] Lu JJ, Tsai FJ, Ho CM, Liu YC, Chen CJ. Peptide biomarker discovery for identification of methicillin-resistant and vancomycin-intermediate Staphylococcus
aureus strains by MALDI-TOF. Anal Chem 2012;84:5685–92.
[23] Huang SY, Tam MF, Hsu YT, Lin JH, Chen HH, Chuang CK, et al. Developmental changes of heat-shock proteins in porcine testis by a proteomic analysis. Theriogenology 2005;64:1940–55.
[24] SAS. 2010. SAS/STAT User's Guide: Version 9.2 ed. SAS Institute Inc., Cary, NC, USA
[25] Zhang M, Jiang M, Bi Y, Zhu H, Zhou Z, Sha J. Autophagy and apoptosis act as partners to induce germ cell death after heat stress in mice. PloS One 2012;7:e41412.
[26] Kim JH, Park SJ, Kim TS, Park HJ, Park J, Kim BK, et al. Testicular hyperthermia induces Unfolded Protein Response signaling activation in spermatocyte. Biochem Biophys Res Commun 2013;434:861–6.
[27] Kaufman RJ. Stress signaling from the lumen of the endoplasmic reticulum: coordination of gene transcriptional and translational controls. Genes Dev 1999;13:1211–33.
[28] Kaufman RJ, Back SH, Song B, Han J, Hassler J. The unfolded protein response is required to maintain the integrity of the endoplasmic reticulum, prevent oxidative stress and preserve differentiation in β-cells. Diabetes Obes Metab 2010;12:99–107. [29] Zhang JC, Kim S, Helmke BP, Yu WW, Du KL, Lu MM, et al. Analysis of
SM22-deficient mice reveals unanticipated insights into smooth muscle cell differentiation and function. Mol Cell Biol 2001;21:1336–44
[30] Chainiaux F, Magalhaes JP, Eliaers F, Remacle J, Toussaint O. UVB-induced premature senescence of human diploid skin fibroblasts. Int J Biochem Cell Biol 2002;34:1331–9.
Induction of replicative senescence biomarkers by sublethal oxidative stresses in normal human fibroblast. Free Radical Biol Med 2000;28:361–73.
[32] Frippiat C, Chen QM, Zdanov S, Magalhaes JP, Remacle J, Toussaint O. Subcytotoxic H2O2 stress triggers a release of transforming growth factor-beta 1,
which induces biomarkers of cellular senescence of human diploid fibroblasts. J Biol Chem 2001;276:2531–7.
[33] Li Q, Shi R, Wang Y, Niu X. TAGLN suppresses proliferation and invasion, and induces apoptosis of colorectal carcinoma cells. Tumor Biol 2013;34:505–13. [34] Thompson O, Moghraby JS, Ayscough KR, Winder SJ. Depletion of the actin
bundling protein SM22/transgelin increases actin dynamics and enhances the tumourigenic phenotypes of cells. BMC Cell Biol 2012;13:1.
[35] Cooper GM. Cell: a molecular approach. Massachusetts, USA: Sinauer Associates 4th ed.; 2007.
[36] Gorgoni B, Gray NK. The roles of cytoplasmic poly(A)-binding proteins in regulating gene expression: a developmental perspective. Brief Funct Genomic Proteomic 2004;3:125–41.
[37] Ozturk S, Guzeloglu-Kayisli O, Demir N, Sozen B, Ilbay O, Lalioti MD, et al. Epab and Pabpc1 are differentially expressed during male germ cell development. Reprod Sci 2012;19:911–22.
[38] Marissen WE, Triyoso D, Younan P, Lloyd RE. Degradation of poly(A)-binding protein in apoptotic cells and linkage to translation regulation. Apoptosis 2004;9:67–75.
[39] Nagano-Ito M, Banba A, Ichikawa S. Functional cloning of genes that suppress oxidative stress-induced cell death: TCTP prevents hydrogen peroxide-induced cell death. FEBS Lett 2009;583:1363–7.
[40] Kimura M, Ishida K, Kashiwabara S, Baba T. Characterization of two cytoplasmic poly(A)-binding proteins, PABPC1 and PABPC2, in mouse spermatogenic cells. Biol Reprod 2009;80:545–54.
[41] Mirande M. Aminoacyl-tRNA synthetase family from prokaryotes and eukaryotes: structural domains and their implications. Prog Nucleic Acid Res Mol Biol 1991;40:95–142.
[42] Reed VS, Wastney ME, Yang DC. Mechanisms of the transfer of aminoacyl-tRNA from aminoacyl-tRNA synthetase to the elongation factor 1 alpha. J Biol Chem 1994;269:32932–6.
[43] Sang Lee J, Gyu Park S, Park H, Seol W, Lee S, Kim S. Interaction network of human aminoacyl-tRNA synthetases and subunits of elongation factor 1 complex. Biochem Biophys Res Commun 2002;291:158–64.
[44] Wiech H, Buchner J, Zimmermann R, Jakob U. Hsp90 chaperones protein folding
in vitro. Nature 1992;358:169–70.
[45] Katoh Y, Fujimoto M, Nakamura K, Inouye S, Sugahara K, Izu H, et al. Hsp25, a member of the Hsp30 family, promotes inclusion formation in response to stress. FEBS Lett 2004;565:28–32.
[46] Wieske M, Benndorf R, Behlke J, Dölling R, Grelle G, Bielka H, et al. Defined sequence segments of the small heat shock proteins HSP25 and alphaB-crystallin inhibit actin polymerization. Eur J Biochem 2001;268:2083–90.
[47] Oliver JD, Roderick HL, Llewellyn DH, High S. ERp57 functions as a subunit of specific complexes formed with the ER lectins calreticulin and calnexin. Mol Biol Cell 1999;10:2573–82.
[48] Glickman MH, Ciechanover A. The ubiquitin-proteasome proteolytic pathway: destruction for the sake of construction. Physiol Rev 2002;82:373–428.
[49] Jariel-Encontre I, Bossis G, Piechaczyk M. Ubiquitin-independent degradation of proteins by the proteasome. Biochim Biophys Acta 2008;1786:153–77.
[50] Carrard G, Bulteau AL, Petropoulos I, Friguet B. Impairment of proteasome structure and function in aging. Int J Biochem Cell Biol 2002;34:1461–74.
[51] Whatley BR, Li L, Chin LS. The ubiquitin-proteasome system in spongiform degenerative disorders. Biochim Biophys Acta 2008;1782:700–12.
[52] Meyer H, Bug M, Bremer S. Emerging functions of the VCP/p97 AAA-ATPase in the ubiquitin system. Nat Cell Biol 2012;14:117–23.
[53] Wójcik C, Rowicka M, Kudlicki A, Nowis D, McConnell E, Kujawa M, et al.
Valosin-containing protein (p97) is a regulator of endoplasmic reticulum stress and of the degradation of N-end rule and ubiquitin-fusion degradation pathway substrates in mammalian cells. Mol Biol Cell 2006;17:4606–18.
VCP mutations causing frontotemporal lobar degeneration disrupt localization of
TDP-43 and induce cell death. J Biol Chem 2009;284:12384–98.
[55] Rawe VY, Diaz ES, Abdelmassih R, Wojcik C, Morales P, Sutovsky P, et al. The role of sperm proteasomes during sperm aster formation and early zygote development: implications for fertilization failure in humans. Hum Reprod 2008;23:573–80.
[56] D'Amours O, Frenette G, Fortier M, Leclerc P, Sullivan R. Proteomic comparison of detergent-extracted sperm proteins from bulls with different fertility indexes. Reproduction 2010;139:545–56.
[57] Martinez-Heredia J, de Mateo S, Vidal-Taboada JM, Ballesca JL, Oliva R. Identification of proteomic differences in asthenozoospermic sperm samples. Hum Reprod 2008;23:783–91.
[58] Crawford LJ, Walker B, Irvine AE. Proteasome inhibitors in cancer therapy. J Cell Commun Signal 2011;5:101–10.
[59] Peron M, Bonvini P, Rosolen A. Effect of inhibition of the ubiquitin-proteasome system and Hsp90 on growth and survival of rhabdomyosarcoma cells in vitro. BMC Cancer 2012;12:233.
[60] Yang Y, Yu X. Regulation of apoptosis: the ubiquitous way. FASEB J 2003;17:790–9.
[61] Zhu K, Dunner K Jr, McConkey DJ. Proteasome inhibitors activate autophagy as a cytoprotectiveresponse in human prostate cancer cells. Oncogene 2010; 29:451–62. [62] Aumuller G, Seitz J. Immunocytochemical localization of actin and tubulin in rat
testis and spermatozoa. Histochemistry 1988;89:261–7.
[63] Kubota H, Hynes G, Carne A, Ashworth A, Willison K. Identification of six Tcp-1-related genes encoding divergent subunits of the TCP-1-containing chaperonin. Curr Biol 1994;4:89–99.
[64] Kubota H, Hynes G, Willison K. The chaperonin containing t-complex polypeptide 1 (TCP-1). Multisubunit machinery assisting in protein folding and assembly in the eukaryotic cytosol. Eur J Biochem 1995;230:3–16.
[65] Lewis SA, Tian G, Vainberg IE, Cowan NJ. Chaperonin-mediated folding of actin and tubulin. J Cell Biol 1996;132:1–4.
al. Eukaryotic cytosolic chaperonin contains t-complex polypeptide 1 and seven related subunits. Proc Natl Acad Sci USA 1993;90:11975–9.
[67] Bourke GJ, El Alami W, Wilson SJ, Yuan A, Roobol A, Carden MJ. Slow axonal transport of the cytosolic chaperonin CCT with Hsc73 and actin in motor neurons. J Neurosci Res 2002;68:29–35.
[68] Yokota SI, Yanagi H, Yura T, Kubota H. Upregulation of cytosolic chaperonin CCT subunits during recovery from chemical stress that causes accumulation of unfolded proteins. Eur J Biochem 2000;267:1658–64.
[69] Yokota S, Yanagi H, Yura T, Kubota H. Cytosolic chaperonin is up-regulated during cell growth. Preferential expression and binding to tubulin at G(1)/S transition through early S phase. J Biol Chem 1999;274:37070–8.
[70] Soues S, Kann ML, Fouquet JP, Melki R. The cytosolic chaperonin CCT associates to cytoplasmic microtubular structures during mammalian spermiogenesis and to heterochromatin in germline and somatic cells. Exp Cell Re 2003;288:363–73. [71] Dun MD, Smith ND, Baker MA, Lin M, Aitken RJ, Nixon B. The chaperonin
containing TCP1 complex (CCT/TRiC) is involved in mediating sperm-oocyte interaction. J Biol Chem 2011;286:36875–87.
[72] Redgrove KA, Anderson AL, Dun MD, McLaughlin EA, O'Bryan MK, Aitken RJ,
et al. Involvement of multimeric protein complexes in mediating the capacitation-dependent binding of human spermatozoa to homologous zonae pellucidae. Dev Biol 2011;356:460–74.
[73] Byrne K, Leahy T, McCulloch R, Colgrave ML, Holland MK. Comprehensive mapping of the bull sperm surface proteome. Proteomics 2012;12:3559–79.
[74] Yokota SI, Yanagi H, Yura T, Kubota H. Upregulation of cytosolic chaperonin CCT subunits during recovery from chemical stress that causes accumulation of unfolded proteins. Eur J Biochem 2000;267:1658–64.
[75] Amit M, Weisberg SJ, Nadler-Holly M, McCormack EA, Feldmesser E, Kaganovich D, et al. Equivalent mutations in the eight subunits of the chaperonin CCT produce dramatically different cellular and gene expression phenotypes. J Mol Biol 2010;401:532–43.
torsion alters the presence of specific proteins in the mouse testis as well as the phosphorylation status of specific proteins. J Androl 2006;27:285–93.
[77] Kanter M, Aktas C, Erboga M. Heat stress decreases testicular germ cell proliferation and increases apoptosis in short term: an immunohistochemical and ultrastructural study. Toxicol Ind Health 2013;29:99–113.
[78] Thornalley PJ, Vasak M. Possible role for metallothionein in protection against radiation-induced oxidative stress. Kinetics and mechanism of its reaction with superoxide and hydroxyl radicals. Biochim Biophys Acta 1985;827:36–44.
[79] Yang CS, Tsai PJ, Chou ST, Niu YL, Lai JS, Kuo JS. The roles of reactive oxygen species and endogenous opioid peptides in ischemia-induced arrhythmia of isolated rat hearts. Free Radic Biol Med 1995;18:593–8.
[80] Paul C, Teng S, Saunders PT. A single, mild, transient scrotal heat stress causes hypoxia and oxidative stress in mouse testes, which induces germ cell death. Biol Reprod 2009;80:913–9.
[81] Lev N, Ickowicz D, Melamed E, Offen D. Oxidative insults induce DJ-1 upregulation and redistribution: implications for neuroprotection. Neurotoxicology 2008;29:397–405.
[82] Mitsumoto A, Nakagawa Y. DJ-1 is an indicator for endogenous reactive oxygen species elicited by endotoxin. Free Radic Res 2001;35:885–93.
[83] Inberg A, Linial M. Protection of pancreatic beta-cells from various stress conditions is mediated by DJ-1. J Biol Chem 2010;285:25686–98.
[84] Fan Z, Hu X, Zhang Y, Yu C, Qian K, Qin A. Proteomics of DF-1 cells infected with avian leukosis virus subgroup J. Virus Res 2012;167:314–21.
Figure 1. Testicular morphology of heat-stressed L2 strain Taiwan country chickens with different recovery periods. Testes were collected at 0 h (B), 2 h (C), and 6 h (D) after 4 h of acute heat stress, and testes from non-heat-stressed chickens were also collected as control (A). The testicular sections were stained by Hematoxylin and eosin. Arrow, aberrant multinucleated giant cells; arrowhead, hyperchromatic nuclei. Magnification: 400X. Bar=20 µm. 1 2 3 4 5 6 7
Figure 2. TUNEL assay of testes from L2 strain Taiwan country chickens during recovery after acute heat stress. The testes were collected for TUNEL assay at 0 h (C, D) 2 h (E, F), and 6 h (G, H) after acute heat stress. Testes from non-heat-stressed chickens were used as control group (A, B). Bar=100 µm (for A, C, E, and G). 1
2 3 4 5
Bar=20 µm (for B, D, F, and H). Arrow, apoptotic cell. 1
Figure. 3. 2D-DIGE protein profiles of chicken testis without or with/4 h of 38°C acute heat stress and different recovering time. Protein samples were labeled with Cy-dyes, separated with pH 3-10, 24 cm non-linear IPG strips and followed by 12.5% SDS-PAGE. Control, non heat-stressed group; R0, heat-stressed group without recovery; R2, heat-stressed group with 2 h of recovery; R6, heat-stressed group with 6 h of recovery. Yellow circle, differentially expressed protein spots with fold-change higher than 1.5. 1 2 3 4 5 6 7 8 9
Figure 4. Gene ontology (GO) analysis of differentially of the differentially expressed proteins (92 proteins) after acute heat stress. The original GO terms of cellular component, molecular function, and biological process annotations were downloaded from the NCBI Entrez gene database and further analyzed. The percentages are the total hits divided by the number of annotated proteins for the category.
1 2 3 4 5 6
Figure 5. Protein expression levels of HSP90α, PABPC1, CCTβ, and TAGLN in chicken testes after acute heat stress. (A) Western blot analysis of HSP90α, PABPC1, CCTβ, TAGLN, and GAPDH. GAPDH was used as an internal control. (B-F) Relative levels of each protein expression are shown. Data was presented as mean±standard error. *P < 0.05 compared to control. C, control; R0, heat stress without recovery; R2, heat stress with 2 h of recovery; R6, heat stress with 6 h of recovery. 1 2 3 4 5 6 7 8
Figure 6. Immunohistochemical analysis of the expression of HSP90α (A-D), PABPC1 (F-I), CCTβ (K-N), and TAGLN (P-S) in chicken testis after acute heat stress. C, non-heat stressed; R0, 0 h of recovery after heat stress; R2, 2 h of recovery after heat stress; R6, 6 h of recovery after heat stress; NC, negative control. Arrowhead, 1
2 3 4
primary spermatocyte; arrow, secondary spermatocyte; +, round spermatid; ♦, elongated spermatid. Bar=20 μm. 1
Figure 7. Possible mechanism of acute heat stress response in chicken testes. (a) Heat stress causes a reduction of PABPC1 and results in destabilization of mRNA translation. (b) Under normal condition, tRNA aminoacylation is performed by aminoacyl-tRNA synthetases associated with elongation factor 1 subunits (EF-1 subunits) and inorganic pyrophosphatase (PPase). After that, elongation factor 1-alpha 1 (EF-1α) carries aminoacyl-tRNA to ribosome. In testes of heat-stressed chickens, aspartyl-tRNA synthetase (AspRS), lysyl-tRNA synthetase (LysRS), eukaryotic translation elongation factor 1 gamma (one of EF-1 subunits), PPase, EF-1α, and ribosome subunits are decreased. (c) In endoplasmic reticulum, protein disulfide-isomerase A3 precursor (PDIA3) participated in the formation of disulfide bond in proteins reduced after heat stress. (d) In cytoplasm, HSP90α and CCT assist folding of proteins, especially actin and tubulin, involved in cell cycle and cell adhesion. Nevertheless, HSP90α and CCT are abated after heat stress and subsequently 1 2 3 4 5 6 7 8 9 10 11 12 13 14
interrupted the folding of actin and tubulin. (e) The valosin-containing protein (VCP) modulates ubiquitin protease system. Misfoled proteins are ubiquinted and then degraded by 26S proteasome complex. After heat stress, the VCP and subunit of 26S proteasome are downregulated. (f) The antioxidant enzymes, glutathione S-transferase (GST) and DJ-1 are upregulated to attenuate the ROS induced after acute heat stress. 1
2 3 4 5
Table 1. Arrangement of samples in the different 2-D DIGE gels#.
Cy2 (100 µg) Cy3 (100 µg) Cy5 (100 µg)
Gel1 Pool* Control R0
Gel2 Pool R2 R6
Gel3 Pool R6 Control
Gel4 Pool Control R2
Gel5 Pool R0 R2
Gel6 Pool R6 R0
# Testis samples were collected from L2 strain Taiwan Country chicken with acute heat stress followed by different
recovery time (R0, R2, and R6) or without heat stress (Control).
*Pool: mixture of all samples in each condition (Control, R0, R2, and R6). The final amounts of protein in gels 4, 5, and 6 are 500 µg.
1
2 3 4 5
Table 2. Differentially expressed protein spots in the testis of L2 strain Taiwan country chicken after heat stress. Spot
No.
Protein identity
Average Ratio*
Gene Symbol GeneBank Accession Theoretical Mr/pI MALDI-MS PMF# MALDI-MS/MS LIFT& Matched peptides R0/ C R2/ C R6/ C
89 transitional endoplasmic reticulum ATPase 1.06 -2.24 -1.72 VCP gi|113206112 89953/5.1 29/97 (37) NS NS
90 transitional endoplasmic reticulum ATPase 1.06 -1.8 -1.87 VCP gi|113206112 89953/5.1 31/96 (34) 3 (66, 82, 123) R.WALSQSNPSALR.E R.KYEMFAQTLQQSR.G K.MTNGFSGADLTEICQR.A
101 transitional endoplasmic reticulum ATPase 1 -2.44 -2.36 VCP gi|113206112 89953/5.1 36/120 (40) NS NS
220 heat shock protein HSP 90-alpha 1.03 -1.22 -1.79 HSP90AA1 gi|157954047 84406/4.9 16/38 (20) NS NS
239 heat shock protein HSP 90-alpha 1.01 -1.32 -1.66 HSP90AA1 gi|157954047 84406/4.9 16/49 (19) 1 (39) K.HSQFIGYPIR.L
267 CTP synthase 2 1.53 1.3 1.26 CTPS2 PYRG2_CHICK 66711/6.4 14/64 (25) NS NS
278 Lysyl-tRNA synthetase 1.17 -1.52 -1.62 KARS gi|71895483 68330/5.9 9/37 (17) 1 (63) R.FFDDLCAVR.N
284 Lysyl-tRNA synthetase -1.27 -2.19 -1.98 KARS gi|71895483 68330/5.9 22/108 (38) 2 (73, 46) R.FFDDLCAVR.N
R.RGDIIGVVGNPGK.T
325 Pyruvate kinase muscle isozyme 1.11 -2.63 -1.82 PKM KPYK_CHICK 58434/7.3 9/45 (22) NS NS
330 PREDICTED: t-complex protein 1 subunit gamma-like 1.04 -5.25 -3.93 LOC100548932 gi|326935888 61089/6.7 31/118 (51) 3 (59, 63, 67) R.AVAQALEVIPR.T R.TLIQNCGASTIR.V K.IPGGFSEDSCVLR.G
345 NEDD8-activating enzyme E1 regulatory subunit -1.05 -1.58 -1.61 NAE1 ULA1_CHICK 61016/5.4 13/91 (25) NS NS
353 heterogeneous nuclear ribonucleoprotein K -1.11 -1.37 -1.64 HNRPK gi|71897277 47591/6.1 14/74 (32) 1 (77) R.NLPLPPPPPPR.G
361 T-complex protein 1 subunit epsilon -1.1 -4.01 -4.1 CCT5 gi|60302774 60190/5.4 29/117 (56) 3 (50, 52, 48) K.QQISLATQVVR.M
R.SIHDALCVIR.C R.IADGYEQAAR.I 368 PREDICTED: ubiquitin carboxyl-terminal hydrolase
14 isoform 1
1.07 -1.48 -1.56 USP14 gi|363735534 52320/5.4 12/97 (33) 1 (74) K.AQLFALTGVQPAR.Q
371 myotubularin-related protein 9 -1.04 -2.18 -2.01 MTMR9 gi|71894897 64098/5.9 NS 1 (57) K.LINSTLGPGR.R
385 T-complex protein 1 subunit zeta 1.08 -3.43 -3.45 CCT6A gi|57525300 58008/6.4 18/103 (36) 1 (86) R.AQAALAVNISAAR.G
392 T-complex protein 1 subunit eta -1.06 -4.17 -3.6 CCT7 gi|71895883 60881/5.9 29/119 (53) 1 (49) R.VPEEDLKR.T
396 Aspartyl-tRNA synthetase 1.08 -1.77 -1.19 Dars SYDC_RAT 57546/6.0 13/78 (39) NS NS
410 T-complex protein 1 subunit theta 1.06 -1.49 -2.13 CCT8 gi|52138673 60017/5.4 17/76 (40) 2 (34, 26) K.HYSGLEEAVYR.N
K.LFVTNDAATILR.E
415 T-complex protein 1 subunit theta 1.04 -1.62 -1.88 CCT8 gi|52138673 60017/5.4 13/41 (23) NS NS
416 myotubularin-related protein 9 1.26 -1.8 -1.18 MTMR9 gi|71894897 64098/5.9 NS 1 (52) K.LINSTLGPGR.R
419 T-complex protein 1 subunit alpha -1.08 -4.03 -4.07 TCP1 gi|57530301 61056/5.6 24/109 (38) 3 (34, 37, 26) R.EQLAIAEFAR.S
K.FATEAAITILR.I
K.YFVDAGAMAVR.R + Oxidation (M)
420 protein disulfide-isomerase A3 precursor -1.07 -4.12 -4.13 PDIA3 gi|45383890 56546/5.7 30/104 (58) 1 (48) K.FIGDKDASVVGFFR.D
422 PREDICTED: zinc finger protein 229 1.28 -1.56 -1.01 ZNF229 gi|345785654 95403/9.1 14/56 (18) NS NS
429 T-complex protein 1 subunit delta 1.01 -3.22 -2.77 CCT4 gi|46048906 58271/9.0 22/82 (44) 1 (49) R.SIHDALCVIR.C
437 T-complex protein 1 subunit delta -1.02 -4.51 -3.29 CCT4 gi|46048906 58271/9.0 21/72 (45) 1 (89) R.SIHDALCVIR.C
438 T-complex protein 1 subunit delta 1.09 -2.87 -2.39 CCT4 gi|46048906 58271/9.0 16/92 (32) NS NS
441 inosine-5’-monophosphate dehydrogenase 2 -1.33 -2.34 -2.8 IMPDH2 gi|71895387 55923/6.6 15/100 (32) 1 (69) K.NLIDAGVDALR.V
442 T-complex protein 1 subunit delta -1.02 -2.24 -2.24 CCT4 gi|46048906 58271/9.0 18/78 (33) NS NS
445 T-complex protein 1 subunit beta 1.02 -5.07 -4.02 CCT2 gi|60302718 57737/5.9 30/102 (62) 2 (47, 64) R.GATQQILDEAER.S
K.HGINCFINR.Q
458 26S protease regulatory subunit 4 -1.07 -2.01 -1.77 PSMC1 gi|45384432 49344/5.8 19/122 (35) NS NS
474 uridine 5’-monophosphate synthase 1.01 1.3 1.53 UMPS gi|71895223 52250/6.2 20/119 (52) 1 (84) R.AELPGVHPVAAR.L
502 NEDD4-binding protein 1 -1.13 -1.97 -1.81 N4BP1 N4BP1_CHICK 104952/5.9 12/46 (15) NS NS
505 PREDICTED: similar to FLJ20699 protein 1.12 -1.62 -2.14 LOC426193 gi|118083181 52821/5.8 NS 1 (49) K.AVDLLYPIR.Y
506 PREDICTED: beta-Ala-His dipeptidase -1.43 -1.5 -1.87 CNDP1 gi|363730532 54809/5.7 12/61 (30) NS NS
510 ruvB-like 1 -1.16 -2.76 -2.34 RUVBL1 gi|57524964 50504/6.0 19/115 (48) 1 (83) R.ALESSISPIVIFASNR.G
530 aspartyl aminopeptidase 1.03 -1.62 -1.59 DNPEP gi|61098378 52569/6.6 16/50 (34) NS NS
565 elongation factor 1-alpha 1 -1.37 -1.52 -1.62 EEF1A1 gi|54020687 50449/9.1 NS 1 (66) K.YYVTIIDAPGHR.D
566 tubulin beta-3 chain -1.21 -1.66 -1.26 TUBB2C gi|153792017 50285/4.8 37/137 (70) 2 (68, 94) R.YLTVAAVFR.G
R.FPGQLNADLR.K
585 eukaryotic translation elongation factor 1 gamma -1.01 -1.65 -1.9 EEF1G gi|311307329 50324/6.1 NS 1 (50) R.SPFVLDEFKR.K
599 26S protease regulatory subunit 7 -1.11 -2.01 -2.04 PSMC2 gi|57525333 49030/5.7 30/162 (62) 1 (54) K.QTLQSEQPLQVAR.C
607 PREDICTED: l-lactate dehydrogenase [cytochrome]-like
633 PREDICTED: 26S protease regulatory subunit 8-like -1.31 -1.71 -2.13 LOC100539380 gi|326933955 46314/7.1 11/40 (29) NS NS
635 tubulin beta-3 chain -1 -1.53 -1.77 TUBB2C gi|153792017 50285/4.8 21/90 (37) 2 (71, 67) R.YLTVAAVFR.G
R.FPGQLNADLR.K
644 lissencephaly-1 homolog -1.24 -1.7 -1.26 PAFAH1B1 gi|45383504 47204/7.2 14/71 (39) 1(43) K.EWIPRPPEK.Y
646 adenylosuccinate synthetase isozyme 2 -1.04 -1.84 -1.57 ADSS gi|71895783 49823/5.9 NS 1 (62) R.TFDELPVNAQNYVR.F
647 PREDICTED: secernin-1 1.69 2.24 1.82 SCRN1 gi|363730132 46696/4.7 17/102 (39) 1 (53) K.IDAEHPELR.S
648 PREDICTED: similar to KIAA0193 (SCRN1) -1.46 -1.74 -1.35 LOC420635 gi|118086002 46638/4.5 17/66 (38) 1 (70) K.VECTYIEIEQVPK.T
657 polyadenylate-binding protein 1 -1.03 -2.4 -3.09 PABPC1 gi|71896197 71123/9.5 25/99 (32) 1 (66) K.FSPAGPILSIR.V
658 poly(A) binding protein, cytoplasmic 1, isoform CRA_d
1.13 -2 -2.39 PABPC1 gi|119612224 53531/9.4 21/89 (36) 1 (72) K.FSPAGPILSIR.V
663 polyadenylate-binding protein 1 1 -2.75 -3.47 PABPC1 gi|383417509 51817/9.4 17/68 (38) NS NS
670 Phosphoglycerate kinase -1.14 1.68 1.81 PGK PGK_CHICK 45087/8.3 15/104 (39) 1 (109) K.LGDVYVNDAFGTAHR.A
671 phosphoglycerate kinase 1.05 1.7 1.7 PGK gi|45384486 45087/9.2 16/83 (37) 1 (48) K.LGDVYVNDAFGTAHR.A
674 phosphoglycerate kinase -1.18 1.63 1.82 PGK gi|45384486 45087/9.2 21/111 (48) 2 (97, 140) K.LGDVYVNDAFGTAHR.A
K.ACANPADGSVILLENLR.F
678 phosphoglycerate kinase -1.19 1.73 2.05 PGK gi|45384486 45087/9.2 19/114 (44) 2 (85, 113) K.LGDVYVNDAFGTAHR.A
K.ACANPADGSVILLENLR.F
679 isocitrate dehydrogenase [NADP], mitochondrial 1.09 -1.3 -2.33 IDH2 gi|71896205 51007/9.8 16/70 (38) NS NS
682 phosphoglycerate kinase 1.07 -1.79 -2.56 PGK gi|45384486 45087/9.2 16/83 (38) 1 (80) K.LGDVYVNDAFGTAHR.A
684 phosphoglycerate kinase 1.16 -2 -3.38 PGK gi|45384486 45087/9.2 15/62 (42) 2 (152, 113) K.LGDVYVNDAFGTAHR.A
K.ACANPADGSVILLENLR.F
688 phosphoglycerate kinase 1.25 -1.75 -2.93 PGK gi|45384486 45087/9.2 22/131 (52) 2 (121, 68) K.LGDVYVNDAFGTAHR.A
K.ACANPADGSVILLENLR.F
692 26S protease regulatory subunit S10B -1.1 -1.67 -1.77 PSMC6 gi|57529874 44431/6.7 15/52 (38) 2 (52, 46) K.LSDGFNGADLR.N
711 methylosome protein 50 1.06 1.31 1.53 WDR77 gi|71895697 39176/4.7 11/121 (33) 1 (49) K.DTIFLSCAEDER.I
722 serpin B6 -1.03 -1.11 2.23 SERPINB6 gi|57530448 43243/5.1 23/111 (55) 1 (57) K.TSSILFCGR.F
752 PREDICTED: 26S proteasome non-ATPase regulatory subunit 13-like
-1.06 -2.95 -3 LOC422990 gi|363734044 43353/5.4 18/80 (48) 2 (66, 67) K.GSIDEVDKR.V
K.DLPVSEQQER.A 780 Chain A, Structure Of Farnesyl Pyrophosphate
Synthetase
1.17 -1.56 -1.35 gi|157834058 42447/6.2 12/61 (28) 2 (39, 39) K.VDLSHFSEER.Y
R.AAFQQYEESSYR.R