Cloning and expression of the a-amylase gene and oxytetracycline production
in Streptomyces rimosus
Chun-Wen Cheng1, Jin-Seng Lin1, Yu-Tien Liu2and Shang-Shyng Yang1,*
1Department of Agricultural Chemistry, National Taiwan University Taipei 10617, Taiwan
2Institute of Microbiology and Immunology, National Defense Medical Center, Taipei 10617, Taiwan
*Author for correspondence: Tel.: +886-2-236 21 519, Fax: +886-2-236 79 827, E-mail: [email protected] Received 5 April 1999; accepted 15 January 2000
Keywords: Amylase activity, amylase gene, oxytetracycline production, Streptomyces rimosus Summary
An 8.4 kb Sau3AI DNA fragment containing the Streptomyces rimosus TM-55 a-amylase gene (amy) was ligated to a vector pIJ702, named pCYL01, and cloned into amylase de®cient mutant S. lividans M2 (amy)). Subcloning study
showed that the amy gene was localized in 3.3 kb KpnI-PstI fragment. The molecular weight of the puri®ed a-amylases of S. lividans M2/pCYL01 and S. rimosus TM-55 were estimated to be 65.7 kDa. Dierent sizes of recombinant plasmids carrying the amy gene had been retransferred into the parental strain of S. rimosus TM-55. Among these S. rimosus transformants, TM-55/pCYL01, TM-55/pCYL12 and TM-55/pCYL36 showed amylase activity 1.36- to 2.05-fold at the seventh day (1.61 to 2.42 units vs 1.18 units), and oxytetracycline (OTC) production 2.00- to 2.50-fold at the ninth day (approximate 140 to 170 lg ml)1vs 72 lg ml)1), higher than that of S. rimosus
TM-55 alone, respectively. These results showed that industrial microorganisms could be improved by genetic and metabolic engineering.
Introduction
Streptomyces are important soil actinomycetes because of their ability to produce diverse secondary metabo-lites and are useful in biological and medical applica-tions. Several genes encoding polysaccharide-degrading enzymes in Streptomyces species have been studied (Bahri & Ward 1993; Tsao et al. 1993). a-Amylase (EC 3.2.1.1), an extracellular enzyme, cleaves the a-1,4-glycosidic linkage in starch and generates shorter chains of oligosaccharide and even maltose. a-Amylase genes have been cloned and characterized, included S. hygroscopicus (Hoshiko et al. 1987), S. lividans (Tsao et al. 1993) and S. venezuelae (Virolle et al. 1988). S. rimosus TM-55 is an oxytetracycline (OTC)-producing strain. Our previous studies have shown that S. rimosus excretes high a-amylase activity into the medium during OTC production in submerged and solid-state fermentations (Wang & Yang 1995; Yang & Cheng 1996; Yang & Wang 1996, 1999). To investigate the application of the amy gene in OTC production, we cloned the S. rimosus TM-55 amy gene and retrans-ferred the dierent sizes of recombinant plasmids harbouring the gene into S. rimosus TM-55. The a-amylase activity and OTC production in the parental strain of S. rimosus and its amy retransformants were also evaluated.
Materials and Methods Test organisms
An amylase de®cient mutant of S. lividans M2 (amy))
used as a cloning host was obtained from Professor W.H. Hsu (National Chung-Hsing University, Taiwan). S. rimosus TM-55 (CCRC 940061) which expressed both amylase activity and OTC production was provided by Dr. Thomas H.H. Ku (Cyanamid Taiwan Corporation). Bacillus subtilis ATCC 6633 was purchased from the American Type Culture Collection. The plasmid pIJ702 was kindly provided by Professor D.A. Hopwood (John Innes Institute, UK) which has two selected markers of black pigment (mel) and thiostrepton resistance (tsrr)
genes.
Culture media and growth conditions
Streptomyces lividans M2 was cultivated in R2YE medium or YEME broth at 28 °C (Hopwood et al. 1985). The modi®ed R2YE/starch medium replaced the sucrose with 1% (w/v) of soluble starch for secreening amylase activity. Yeast extract/starch (YS) medium containing (g l)1) yeast extract, 5 and soluble starch,
10 was used for the expression of Streptomyces a-amylase. The medium used for antibiotic production contained (g l)1): soluble starch, 20; corn steep liquor,
10; (NH4)2SO4, 6; CaCO3, 8; NaCl, 5 and soybean oil, 2
at pH 6.8 to 7.2. If necessary, 50 lg ml)1of thiostrepton
was also added for Streptomyces harbouring plasmid. Isolation of chromosomal DNA and plasmids
Streptomyces mycelia were grown at 28 °C for 24 to 30 h and harvested by centrifugation. The cell pellet was resuspended in 2.5 ml STE buer (containing 340 g sucrose l)1, 25 mM Tris±HCl and 10 mM Na
2EDTA)
with 2 mg lysozyme ml)1, and followed by the addition
of 10% (w v)1) sarcosyl solution. The mixture was
vortexed softly until it became clear. Afterwards, 9 g CsCl, 0.3 ml ethidium bromide and 0.5 ml TE buer (containing 10 mM Tris±HCl and 1 mM EDTA at pH 8.0) were added to the previously lysed solution. The chromosomal DNA was centrifuged at 80,000 ´g for 48 h at 20 °C, and then the eluted DNA was extracted with n-butanol. Finally, the DNA was dialysed against TE buer at 4 °C overnight.
DNA digestion, plasmid isolation, protoplast prepa-ration and plasmid transformation of Streptomyces were performed according to the methods described by Hopwood et al. (1985).
Cloning of the a-amylase gene
Streptomyces rimosus TM-55 chromosomal DNA was partially digested with restriction enzyme Sau3AI. Six to 9 kb DNA fragments were collected from the 5 to 25% region of the NaCl linear gradient obtained after centrifugation at 68,000 ´g for 5 h at 25 °C. Sau3AI DNA fragment was compatibly ligated to the BglII-cleaved plasmid pIJ702 which had been pretreated with calf intestinal alkaline phosphatase (CIAP). The above Sau3AI genomic libraries were introduced into protop-lasts of S. lividans M2 (amy)). These transformants were
cultivated on R2YE medium containing thiostrepton. All white (insertion inactivation of pIJ702 in mel gene) and thiostrepton-resistant (tsrr) colonies were
retrans-ferred onto R2YE/starch medium by replica plating and incubated at 30 °C for 2 days. The plates were sprayed with iodine solution (0.127% I2and 1.66% KI in water),
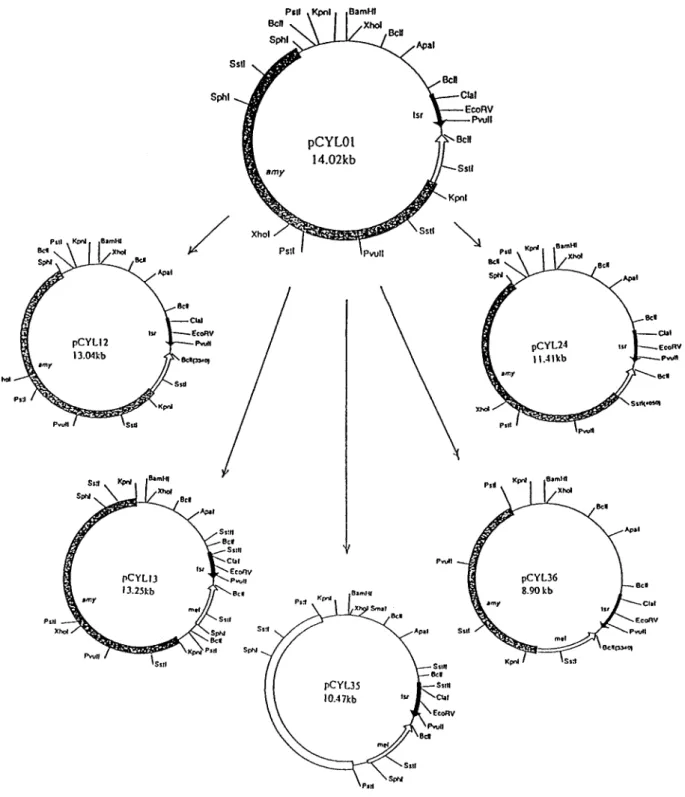
and the transformant with recombinant plasmid carry-ing an amy gene showed a clear zone. The host strain S. lividans M2 was used as a negative control in this screening. ApaI, BamHI, BglII, ClaI, EcoRI, KpnI, HpaI, HindIII, PstI, SalI, SacI, SphI and XhoI were used to construct the physical map of the recombinant pCYL01 with the amy gene from S. rimosus TM-55 as shown in Figure 1.
Puri®cation of a-amylase
The a-amylase of the S. lividans M2/pCYL01 transfor-mant was puri®ed according to the procedures described in the previous paper (Yang & Cheng 1996). Mycelia were removed by centrifugation at 1500 ´g for 15 min. The supernatant was subjected to protein precipitation
with ammonium sulphate between 30 and 70% of saturation. The precipitate was dissolved and dialysed in 20 mMphosphate buffer, pH 6.8 at 4 °C. It was further puri®ed by gel ®ltration on a DEAE-Sephacryl column (1 cm i.d. ´ 28 cm) (Pharmarcia, Sweden) and eluted with 0 to 0.8 M of a-NaCl linear gradient in 0.02M phosphate buffer at 0.14 ml min)1. Fractions with
enzyme activity were pooled and then passed through a Sephadex G-100 column (Pharmarcia, Sweden). Active fractions were concentrated with a Centricon microconcentrator (Amicon, USA).
a-Amylase activity assay
a-Amylase activity was measured as described by the Wilson & Ingledew (1982). One millilitre of enzyme solution and 2.0 ml of 0.2% soluble starch in 0.05M K2PO4±NaOH buer pH 6.0 were incubated at 40 °C
for 10 min. The absorbance of reaction mixture 0.2 ml and iodine reagent (0.05% I2 and 5% KI in water)
5.0 ml were measured at 620 nm against the blank. One unit of enzyme activity was de®ned as the amount of enzyme hydrolysing 1.0 mg of soluble starch in 10 min under the assay conditions.
Determination of oxytetracycline
OTC was determined by the paper disc method in antibiotic medium I (Merck No. 1 A-5272, Germany) with strain Bacillus subtilis ATCC 6633 at 30 °C (Yang & Ling 1989). Total OTC equivalent potency was calculated from the clear zone of a standard curve of OTC in the range of 1 lg ml)1 to 10 mg ml)1. The
regression study indicated that Y = 0.1322X ) 0.8917 and r2= 0.9912, where Y is the log value of
concen-tration of OTC, and X is the diameter of inhibition zone. Chemical analysis
Total protein was determined by the Lowry method. Reducing sugar content was measured by the dinitro-salicylic acid (DNS) method.
Results
Cloning of an a-amylase gene from S. rimosus TM-55 About 3000 white (melanin defective) and thiostrepton-resistant S. lividans M2 transformants were collected. These colonies were transferred onto YS medium by replica plating, incubated for 48 h and then sprayed with I2/KI solution. One of the S. lividans M2
transfor-mants showed a clear zone around the colony under the purple-blue background. Plasmid was isolated from this colony and then tested for amylase activity. All of the S. lividans M2 retransformants showed amylase activity. The recombinant plasmid with the amylase activity was designated as pCYL01 (Figure 1).
A series of subcloning and complementation experi-ments were performed with several restriction endonuc-leases. The DNA insert containing KpnI to SphI fragment had the a-amylase activity in S. lividans M2, included pCYL01, pCYL12 and pCYL13. The 1.84 kb KpnI-SstI deletion in pCYL24 showing weakly positive enzyme activity and no a-amylase activity was detected in the 2.87 kb KpnI-PstI deletion in pCYL35 (Figure 2). Finally, a plasmid with a 3.30 kb of KpnI-PstI fragment, named pCYL36, had the amylase activity as did the
plasmid pCYL01 in S. lividans M2 (Figure 2). Thus, it can be concluded that the cloned amy gene from S. rimosus was located within this 3.3 kb insert. Characterization of the a-amylase from S. rimosus The biochemical properties of the a-amylase puri®ed from the amy gene clone from S. lividans M2/pCYL01 were found to be same as S. rimosus TM-55 a-amylase. Puri®ed a-amylase from both S. lividans M2/pCYL01 Figure 1. Subcloning strategies of the a-amylase gene from S. rimosus TM-55. Solid bar with tsr indicates thiostrepton-resistant; open bar with mel indicates tyrosinase and dot bar with amy indicates a-amylase, respectively. The recombinant plasmids of pCYL01, pCYL12, pCYL13 and pCYL36 showed a-amylase activity. pCYL24 showed weakly a-amylase and pCYL35 had no detectable amylase activity.
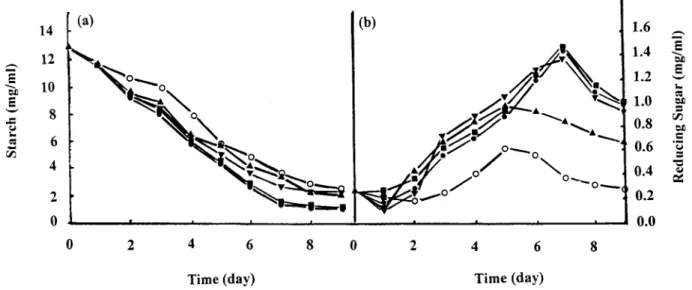
and S. rimosus TM-55 showed the same electrophoretic pattern on SDS-PAGE. The molecular weight of S. rimosus TM-55 a-amylase was 65.7 kDa. Maltose and soluble starch were found to be better carbon sources than glucose for amylase production in the parental strain TM-55 as well as in its transformants carrying the amy gene. Starch concentration decreased with incuba-tion time, whereas reducing sugar concentraincuba-tion de-creased slightly on the ®rst day, then inde-creased gradually and reached the maximal value on the ®fth to the seventh day. Starch consumption was found to be highest in S. rimosus TM-55/pCYL12 and S. rimosus TM-55/pCYL36, followed by S. rimosus TM-55/ pCYL01, S. rimosus TM-55, and S. rimosus TM-55/ PIJ702 respectively (Figure 3a). However, the reducing sugar concentration was maximal in S. rimosus TM-55/
pCYL12 and S. rimosus TM-55/pCYL36, followed by S. rimosus TM-55/pCYL01, S. rimosus TM-55, and S. rimosus TM-55/pIJ702 had the lowest (Figure 3b). a-Amylase production was observed to be growth-associated in the medium containing 2% of carbon source. The optimal reaction pH values were between 5.5 and 6.5, and activity was stable at pH 6±8 for 24 h. Both activities had optimum reaction temperature between 37 and 40 °C. The enzyme activity was found to be about 40% when the reaction temperature was over 50 °C, and it was completely inactivated at 70 °C after 10 min incubation.
a-Amylase activity and oxytetracycline production a-Amylase activities of S. rimosus TM-55/pCYL01, S. rimosus TM-55/pCYL36 and S. rimosus TM-55/ pCYL12 were 1.36-, 1.44- and 2.05-fold higher than that of the parental strain S. rimosus TM-55 on the seventh day, respectively (Figure 4a). The enzyme ac-tivity in S. rimosus TM-55/pIJ702 was found to be same as that expressed in S. rimosus TM-55. a-Amylase activity of S. lividans M2 (amy)) carrying pIJ702 was
below 0.3 unit ml)1and was used as a negative control.
The oxytetracycline production in the submerged fermentation is presented in Figure 4b. Transformants of S. rimosus TM-55/pCYL01, S. rimosus TM-55/ pCYL12 and S. rimosus TM-55/pCYL36 had oxytetra-cycline potency about 2.00- to 2.50-fold higher than that of the parent strain S. rimosus TM-55 after 9 days of incubation. Oxytetracycline potency of S. rimosus TM-55/pIJ702 was almost undetected.
Discussion
The a-amylase gene of S. rimosus TM-55 localized in a 3.3 kb KpnI-PstI DNA fragment was cloned in Figure 2. Restriction cleavage maps of the S. rimosus amy subclones.
a-Amylase activities of the S. lividans M2 transformants harbouring each recombinant plasmid are indicated on the right side with positive (+) or negative ()). The solid bar represents the corresponding location of the DNA inserts. Abbreviations of enzyme cutting site represented as follows: K: KpnI, Ss: SstI, Pv: PvuII, X: XhoI, Ps: PstI, and Sp: SphI.
Figure 3. Concentration of starch and reducing sugar in S. rimosus TM-55 and its transformants. (a) Starch concentration, (b) reducing sugar concentration. m m, S. rimosus TM-55; s s, S. rimosus TM-55/pIJ702; d d, S. rimosus TM-55/pCYL01; j j, S. rimosus TM-55/ pCYL12; and . ., S. rimosus TM-55/pCYL36.
S. lividans M2. The apparent molecular weight of puri®ed a-amylase from S. lividans M2/pCYL01 har-bouring amy was estimated by SDS-PAGE to be 65 kDa. This is close to the molecular weight of a-amylase of S. limosus (63 kDa) (Long et al. 1987) and somewhat larger than that of a-amylase of S. griseus (59.7 kDa) (Vigal et al. 1991; Garcia-Gonzalez et al. 1991), S. hygroscopicus (52 kDa) (Hoshiko et al. 1987), S. thermoviolaceus (49 kDa) (Bahri & Ward 1993) or S. venezulae (59 kDa) (Virolle et al. 1988), but smaller than that of a-amylase of S. lividans (100 kDa) (Tsao et al. 1993).
We have successfully retransferred the previously cloned plasmids carrying amy into S. rimosus TM-55. The length of pCYL01, pCYL12 and pCYL36 are dierent, but the transformants of S. rimosus TM-55 can express higher levels of a-amylase activity than that of the parental strain. It implied that the amy gene in S. rimosus TM-55 was ampli®ed by using the high copy number of vector pIJ702. These transformants were also found to have higher OTC-producing potencies than that of S. rimosus TM-55. During cultivation, the starch concentration decreased and biomass and a-am-ylase activity increased after 7 days of incubation. In this study, we used starch as the sole source of carbon in submerged fermentation. S. rimosus TM-55 trans-formants harbouring amy could show both a-amylase activity and OTC production higher than that of their parent strain. Upon digestion of starch, the reducing sugar increased gradually. The presence of higher reducing sugar could thus be related to the higher a-amylase activity. Furthermore, the highest a-amylase activity in S. rimosus TM-55/pCYL12 may be due to the moderate length, among the three recombinant plasmids. S. rimosus TM-55/pCYL12 would have more amy gene products with consumption of the same
nucleotide materials in stimulating a-amylase activity than that of pCYL01. Surprisingly, S. rimosus TM-55/ pIJ702 could use soluble starch, however, its optimal reducing sugar content was about 35% lower than that of S. rimosus TM-55. The reason for the lower reducing sugar content in S. rimosus TM-55/pIJ702 cannot be explained at present and further investigation is needed. In addition, S. rimosus TM-55/pCYL01 and S. rimosus TM-55/pCYL36 had higher OTC potencies than that of S. rimosus TM-55/pCYL12 after 9-day incubation, and all of them had higher OTC production than that of S. rimosus TM-55. The correlation between a-amy-lase activity (growth-associated) and OTC production (secondary metabolite) in S. rimosus TM-55 is still unknown. Evidence has demonstrated that during the cultivation of S. rimosus TM-55, the OTC potency could be stimulated by maltose supplementation and inhibited by glucose addition (Yang & Yuan 1990). It was reported that the prevention of glucose repression and the reduction of energy charge could enhance the production of amylase and secondary metabolites (Demain 1972; Vining & Doull 1988; Cheng & Yang 1995). In solid state fermentation, maltose could enhance the production of tetracycline in S. viridifac-iens and OTC in S. rimosus, respectively, whereas galactose or glucose inhibited the OTC production in S. rimosus (Yang & Ling 1989; Yang & Yuan 1990). These ®ndings indicate that the production of enzymes and antibiotics can be regulated by dierent carbon sources.
Whether the production of OTC of these S. rimosus TM-55 transformants is aected by the percentages of starch, oligosaccharide and maltose which are produced by the cloned a-amylase secretion, or by any other factors in the medium (Demain 1989; Ali et al. 1993) during fermentation needs further study.
Figure 4. a-Amylase activity and oxytetracycline production in S. rimosus TM-55 and its transformants. (a) a-Amylase activity, (b) oxytetracycline production. m m, S. rimosus TM-55; s s, S. rimosus TM-55/pIJ702; d d, S. rimosus TM-55/pCYL01; j j, S. rimosus TM-55/pCYL12; and . ., S. rimosus TM-55/pCYL36.
A novel ®nding is that no OTC titre was detected in the transformant of S. rimosus TM-55/pIJ702. It was known that melanin could be oxidized by tyrosinase, a monooxygenase, which is located on the intact pIJ702 (Hopwood et al. 1985). The functional group of OTC may be undergo deactivation by the tyrosinase (mel) oxidation of the transformant S. rimosus TM-55 carry-ing pIJ702 alone durcarry-ing OTC fermentation process. However, when we inserted the amy gene into the mel gene, the tyrosinase was found to be inactivated. Consequently, the OTC production in S. rimosus TM-55 transformants harbouring the amy gene could be prevented from oxidation by tyrosinase.
In summary, we report the cloning and characteriza-tion of the a-amylase gene from S. rimosus TM-55, and retransfer of this gene into its parental strain of S. rimosus TM-55. The amy transformants of S. rimosus TM-55 had both higher a-amylase activity and OTC production than those of the parental strain. Construc-tion of such genetically and metabolically engineered microorganisms is going to gain much importance in industrial production of enzymes in the future.
Acknowledgement
The authors thank the National Science Council and Council of Agriculture of Republic of China for their ®nancial support [NSC 82-0409-B002-195, NSC 83-0409-B002-064, and 87 SCITECH-1.1-AGRIC-17(5)]. References
Ali, A.Z., Abdelrahman, N. & Baghlaf, A. 1993 Use of date products in production of oxytetracycline by Streptomyces rimosus. Bioscience Biotechnology and Biochemistry 57, 987±988.
Bahri, S.M. & Ward, J.M. 1993 Sequence of the Streptomyces thermoviolaceus CUB74 a-amylase encoding gene and its tran-scription analysis in Streptomyces lividans. Gene 127, 133±137. Cheng, C.W. & Yang, S.S. 1995 Amylase production of Streptomyces
rimosus TM-55 and their 2-deoxyglucose resistant mutants. Chinese Journal of Microbiology and Immunology 28, 109±116. Demain, A.L. 1972 Cellular and environmental factors aecting the
synthesis and expression of metabolites. Journal of Applied Chemistry and Biotechnology 22, 345±362.
Demain, A.L. 1989 Functions of secondary metabolites. In Genetics and Molecular Biology of Industrial Microorganisms, eds. Hersh-berger, C.L., Queener, S.W. & Hegeman, G. pp. 1±11. Washington D.C.: American Society for Microbiology. ISBN 1-55581010-1.
Garcia-Gonzalez, M.D., Martin, J.F., Vigal, T. & Liras, P. 1991 Characterization, expression on Streptomyces lividans and pro-cessing of the amylase of Streptomyces griseus IMRU 3570: two dierent amylases are derived from the same gene by an intracel-lular processing mechanism. Journal of Bacteriology 173, 2451± 2458.
Hopwood, D.A., Bibb, M.J., Chater, K.F., Bruton, C.J., Kieser, H.M., Lydiate, D.J., Smith, C.P., Ward, J.M. & Schrempf, H. 1985 Genetic Manipulation of Streptomyces: a Laboratory Manual. The John Innes Foundation, Norwich. ISBN 0-70840336-0. Hoshiko, S., Makabe, O., Nojiri, C., Katsumata, K., Satoh, E. &
Nagaoka, K. 1987 Molecular cloning and characterization of the Streptomyces hygroscopicus a- amylase gene. Journal of Bacterio-logy 169, 1029±1036.
Long, C.M., Virolle, M.J., Chang, S.Y. & Chang, S. 1987 a-Amylase gene of Streptomyces limosus: Nucleotide sequence, expression motifs, and amino acid sequence homology to mammalian and invertebrate a-amylase. Journal of Bacteriology 169, 5745± 5754.
Tsao, L.S., Lin, L.L., Chen, J.C. & Hsu, W.H. 1993 Cloning and characterization of an a-amylase gene from Streptomyces livi-dans. Biochimical et Biophysica Acta 1171, 255±262.
Vigal, T., Gell, J.A., Daza, A., Garcia-Gonzalez, M.D. & Martin, J.F. 1991 Cloning, characterization and expression of an a-amylase gene from Streptomyces griseus IMRU 3570. Molecular Gene and Genetics 225, 278±288.
Vining, L.C. & Doull, J.L. 1988 Catabolite repression of secondary metabolism in Actinomycetes. In Biology of Actinomycetes '88. pp. 406±411. Tokyo: Japan Scienti®c Societies Press. ISBN 4-76221552-X.
Virolle, M.J., Long, C.M., Chang, S. & Bibb, M.J. 1988 Cloning, characterization and regulation of an a-amylase gene from Streptomyces venezuelae. Gene 74, 321±334.
Wang, J.Y. & Yang, S.S. 1995 Morphogenesis, biomass and oxytet-racycline production of Streptomyces rimosus in submerged cultivation. Chinese Journal of Microbiology and Immunology 28, 21±31.
Wilson, J.J. & Ingledew, W.M. 1982 Isolation and characterization of Schwanniomyces alluvius amylolytic enzymes. Applied and Envi-ronmental Microbiology 44, 301±307.
Yang, S.S. & Cheng, C.W. 1996 Production, puri®cation and characterization of a-amylase by Streptomyces rimosus. Journal of the Chinese Agricultural Chemical Society 34, 649±656. Yang, S.S. & Ling, M.Y. 1989 Tetracycline production of sweet potato
residue by solid state fermentation. Biotechnology and Bioengi-neering 33, 1021±1028.
Yang, S.S. & Wang, J.Y. 1996 Morphogenesis, ATP content and oxytetracycline production by Streptomyces rimosus in solid substrate cultivation. Journal of Applied Bacteriology 80, 545±550.
Yang, S.S. & Wang, J.Y. 1999 Protease and amylase production by Streptomyces rimosus in submerged and solid substrate cultivation. Botanical Bulletin of Academia Sinica 40, 259±265.
Yang, S.S. & Yuan, S.S. 1990 Oxytetracycline production with sweet potato residue by solid state fermentation. World Journal of Microbiology and Biotechnology 6, 236±244.