Regulation of K12 Ker atin Gene in Cor neal Epithelial Cells
I-Jong Wang†, Chia-Yang Liu¶, Carlson Eric¶, Fung-Rong Hu†, Winston W.-Y. Kao¶
†Department of Ophthalmology, National Taiwan University Hospital, 7, Chung-Shan South Road, Taipei 100, Taiwan
¶Department of Ophthalmology, University of Cincinnati Medical Center, Health Professions Building, Suite 350, ML0527, Eden and Bethesda Avenues, Cincinnati,
Ohio 45267-0527, USA,
Running title: K12 Keratin Gene in Corneal Epithelial Cells
Correspondence and reprint requests should be addressed to W. W. -Y. Kao.
Department of Ophthalmology, University of Cincinnati Medical Center, Health
Professions Building, Suite 350, ML0527, Eden and Bethesda Avenues, Cincinnati,
Ohio. TEL: (513) 558-5151, FAX: (513) 558-3108 E-mail: [email protected]
INTRODUCTION
Keratins are a group of water-insoluble proteins that form 10 nm intermediate filaments in
all epithelial cells. 1
About 30 different keratin molecules have been identified and can be
divided into acidic and basic neutral subfamilies in tissue-specific manners.1-4
In vivo, a
basic keratin is usually co-expressed and "paired" with a particular acidic keratin. 1;3-5
According to their mode of expression and their association with progressively more
complicated structures, the keratin pairs can be divided into several categories: (1) keratins
of the simple epithelia include the K8/K18 pair and several small MW keratins; (2) keratins
expressed by the basal cells of all stratified squamous epithelia, K5/K14 pair; (3) kertins
expressed by the suprabasal cells of all stratified epithelia that become hyperplastic due to
diseased or wound conditions or tissue culture stimuli, the K6/K16 pair; and (4)keratins
expressed by suprabasal cells of normally differentiating stratified squamous epithelia.6
For
example, expression of the K12/K3 pair has been regarded as a marker for cornea-type
epithelial differentiation.5;7;8
The K12/K3 intermediate filaments are vital for corneal
epithelial cell integrity, phsycal stability and rigidity. Mutations of K12 and K3 genes in
human, and ablation of K12 gene via gene targeting in mice result in fragile corneal
epithelium, a clinical manifestation characteristics of Meesmann’s corneal dystrophy.{9
}10
Although K12/K3 pair is considered as the cornea-type differentiation marker, recent
evidence indicates that K3 can be expressed in several tissues other than the corneal
epithelium.7
}11
demonstrated that the expression of K12 is restricted to the corneal epithelium.7;12-17
The expression of tissue-specific keratin was dependent on the on and/or
turn-off of a variety of genes.18-20
Many of these keratinocytes genes are regulated at the
transcriptional levels in a coordinate manner during keratinocyte differentiation.21;22
Most
of these genes are also regulated in an independent manner by at least one regulatory agent
and the gene regulatory regions do not appear to be conserved. 21
The fine-tuned regulation
of certain keratin genes has been shown in pairwise (e.g., K1-K10, K3-K12, K5-K14, and
K6-K16) and some of these pairs are expressed reciprocally (e.g., K3-K12 and
K5-K14).23-26
There are several transcriptional factors known for the regulation of keratin gene
expression. For example, AP-2 and SP-1have been shown to be able to regulate the
expression of K1, K3, K5, K6, K12, and K14.6;27-29
Furthermore, the regulation of keratin
gene expression always needs the combination of several transcription factors to express
tissue specificity. 6;27-29
We have attempted, but failed to identify the cornea epithelial, K12,
cell-specific promoter using conventional in vitro transfection of cultured corneal epithelial
cells and in vivo transgenic mice.30
But recently, we successfully used Gene Gun,
particle-mediated gene transfer, to deliver K12 transgenes to corneal epithelial cells in vivo.30
We
observed that 2.5 kb DNA fragment 5’ flanking Krt1.12 possibly contained corneal
epithelial cell-specific regulatory cis-DNA elements and PAX 6 is an important
constructs with Gene Gun analysis as well as DNase I footprinting and electrophoretic
motility shift assays (EMSA) were performed to elucidate the regulation of tissue
MATERIALS AND METHODS Plasmids.
Two plasmid DNA constructs i.e., pCMVβ and pNASSβ (Clontech, Palo Alto, CA ), were used as control reporter genes. A series of keratin 12 promoter fragments were
prepared and cloned into pNASSβ vector as shown in Fig. 1. The 2.5 kb Krt1.12 promoter construct was derived by Nru/XhoI cut from a 5.0 kb Krt1.12 promoter
construct and ligated to Ehe/XhoI cut pNASSβ. The 2.0 kb Krt1.12 promoter was derived by SacI/XhoI cut from a 2.5 KB Krt1.12 promoter and ligated into EcoRI/XhoI
cut pNASSβ using an Eco/Asc/Sac adaptor. A 4.5 kb EcoRI/HindIII fragment from 0.6 KB containing the β-gal reporter driven by the 3’ 600 bp of the K12 promoter was ligated to a 3.7 kb EcoRI/HindIII fragment from 2.0 kb Krt1.12 constructs containing
the vector backbone of pNASSβ and 5’ 1000 bp of the K12 promoter. The resulting a 2 kb 0.4 kb vector is identical to 2.0 kb with the exception that is missing 400 bp of the
K12 promoter starting at the 975 bp EcoRI site and extending 400 bp 3’ toward the 3’
end of the promoter. The 2 kb 0.6 kb and 2 kb 0.8 kb construct were produced by
the same method as 2 KB 0.4 kb construct. The 3’ ends of these constructs were
created by digestion with XhoI and corresponded to position +40 on the transcriptional
initiation site. The fragments were cloned in the sense orientation about the β
-galactosidase gene, and constructs were numbered according to the sequence positions
from 2.5 kb Krt1.12 constructs, and all constructs were confirmed by DNA sequencing.
All of these construct and primers used for polymerase chain reaction were shown in
Table1.
In vivo Gene Tr ansfer by Gene Gun.
Plasmid DNA purified by Qiagen columns (Qiagen, Chatsworth, CA) was coated onto gold particles of 0.6 µm, 1.0 µm or 1.6 µm (5 µg DNA per mg of gold particles) according to the procedures recommended by the manufacturer of HeliosTM
Gene Gun
System (Biorad, Hercules, CA). The tubing coated with gold was cut into 0.5-inch
segments. Thus, each segment contains 0.5 mg gold and 2.5 µg of reporter DNA. All animal experiments were performed according to the ARVO resolution on
the use of animals in vision research. New Zealand white rabbits (about 2 kg) were
anesthetized with ketamine (30 mg/kg) and xylazine (3 mg/kg). One drop of 0.5 %
proparacaine/HCl was applied to cornea.30
Back hair was clipped and residual hair was
removed by treatment with NAIR (Carter-Wallace, NY). The Gene Gun was held against corneas, conjunctivas and skin to bombard gold particles into tissues, 1 delivery
per individual cornea, 2 deliveries onto opposite sites of individual conjunctiva, and up
to 12 sites on the skin of individual rabbits. Samples were collected 48 h after delivery
and subjected to further experiments.
Excised tissue specimens were minced with a razor blade, and 0.5 ml of extraction
buffer (0.25 M Tris-HCl, pH 7.4, 0.1 % Tween 20) was added. The samples were
subjected to 3 freeze-thaw cycles, 5 min on dry ice and 5 min at 37°C. The supernatants were collected by centrifugation at 13,ooo x g, 4°C for 10 min.
Assays of β-galactosidae Activity and Whole mount β-galactosidase Histochemical Staining
Aliquots of supernatant were incubated in a 0.3 ml mixture containing 50 mM
2-mercaptoethanol, 1 mM MgCl2, 1.33 mg/ml o-nitrophenyl β-galactopyranoside and 0.1 M phosphate buffer, pH 7.0 at 37°C for 1 to 5 h. 0.7 ml of 1 M Na2CO3 was added to terminate the reaction. The enzyme activity was determined by comparing the optical
density at 460 nm to that of purified β-galactosidase (Boehringer and Mannheim, Indianapolis, IN). The promoter activity of KB reporter gene constructs was calculated
as a fold-increase of the promoter-less pNASSβ construct.
The whole eye ball was enucleated, fixed immediately with 4%
paraformaldehyde in PBS at 4°C for 2hr, and then washed three times with PBS at 4°C. Staining was carried out at 30°C for 16 hr in a solution of
5-bromo-4-chloro-3-indolyl-β-galactopyronoside (X-gal, Sigma) at a final concentration of 0.4 mg/ml made from a 40 mg/ml stock in dimethylformamide, with 4 mM K3Fe(CN)6, 4 mM
K4Fe(CN)6.6H2O, 2 mM MgCl2 in PBS. After staining, eyeballs were rinsed with PBS and photographed as whole mounts.
Prepar ation of Nuclear Extr acts
Nuclear extracts were prepared as described by Andrews and Faller.31
Briefly, bovine
corneal epithelialcells were scraped into ice-cold PBS, and homogenized in lysis buffer
(0.1% Titon X-100). Then homogenized solution was purified through 1.8 M sucrose
cushion at 24,000 rpm for 45 mins, and the pellet was resuspended in cold buffer (20
mM HEPES-KOH, pH 7.9, 25% glycerol, 420 mM NaCl, 1.5 mM MgCl2, 0.2 mM
EDTA, 0.5 mM dithiothreitol, 0.2 mM PMSF). Then, it was homogenized with
dounce-glass homogenizer, and incubated on ice for 20 min. Cellular debris was
removed by centrifugation for 2 min at 4 °C. Then the supernatant fraction, containing
nuclear DNA-binding proteins, was precipitated by 0.39 g/ml of NH4 OH at 34,000 rpm
for 25 mins and stored in aliquots at 70 °C.
DNase I Footpr inting Aanalysis.
Four DNA probes (-332 bp to –109 bp, -394 bp to –131 bp, -599 bp to –375 bp, -1415
bp to –1394 bp) were prepared with polymerase chain reaction by four pairs of primers
(Table 2), and labeled with [γ-32P] ATP by T4 polynucleotide kinase at the 5'-end. They were incubated with crude nuclear extracts (75 µg) from bovine corneal epithelial cells as previously described.32 A 50 µl mixture containing 10 mM Tris (pH 8), 5 mM MgCl2, 5 mM CaCl2, 50 mM KCl, 500 nM dithiothreitol, 0.05 mg/ml bovine serum albumin,
15 minutes. The mixture was incubated for additional 45 minutes with 100,000 CPM of
DNA probe before DNase I digestion for 1 minutes and 30 seconds at 25 o C with
variable amount of DNase I. The DNase I digestion was stopped with 50 µl of stop buffer (0.2 M NaCl, 30 mM ethylenediaminetetraacetic acid, 1% sodium dodecyl
sulfate, and 0.1 mg/ml yeast tRNA). Digested DNA probes were purified by phenol
extraction and ethanol precipitation and separated on 6.5% denatured polyacrylamide
gels. Dried gels were exposed to Kodak XAR film with intensifying screens for 24
hours at –80 o
C. G and G+A sequencing reactions were performed to determine the
positions of the protected regions. Negative control reactions (probe) were performed
in the absence of nuclear protein extracts. Region protected by nuclear proteins and
numbers indicating nucleotide positions relative to the transcription start site of the K12
gene.
Electrophoretic Mobility Shift Assays.
Gel-shift analysis of different region of Krt1.12 promoter was performed with 20000
CPM of 32
P-labeled double-stranded synthetic oligonucleotide probe (-182 bp to –111
bp, -256 bp to –293 bp, –942 bp to –913 bp, -1661 bp to –1624 bp, and –1887 bp to –
1858 bp) (Table 3), and crude nuclear extracts prepared from bovine corneal epithelial
cells as described. Twenty microliters of binding reaction mixtures contained 10 µg of
mM HEPES (pH 7.9), 10% glycerol, 4 mM Tris (pH 8.0), 1 mM
ethylenediaminetetraacetic acid, 0.3 mg/ml bovine serum albumin, 1mM dithiothreitol,
0.1 mg/ml poly(dAdT), and 60 mM KCl. Each labeled probe was competed with an
excess of the same unlabeled, double-stranded synthetic oligomer. Binding mixtures
were separated on a nondenaturing 5% polyacrylamide gel and exposed to Kodak XAR
Results
Functional Analysis of the 5' -Flanking Region of the Krt1.12 Gene
To identify cis-acting regulatory elements in the 5'-flanking region of the Krt1.12 gene,
we constructed a series of 5' deletionβ-gal expression vectors and transiently transfected them into rabbit conneal, conjunctival, and cutaneous epithelia by gene
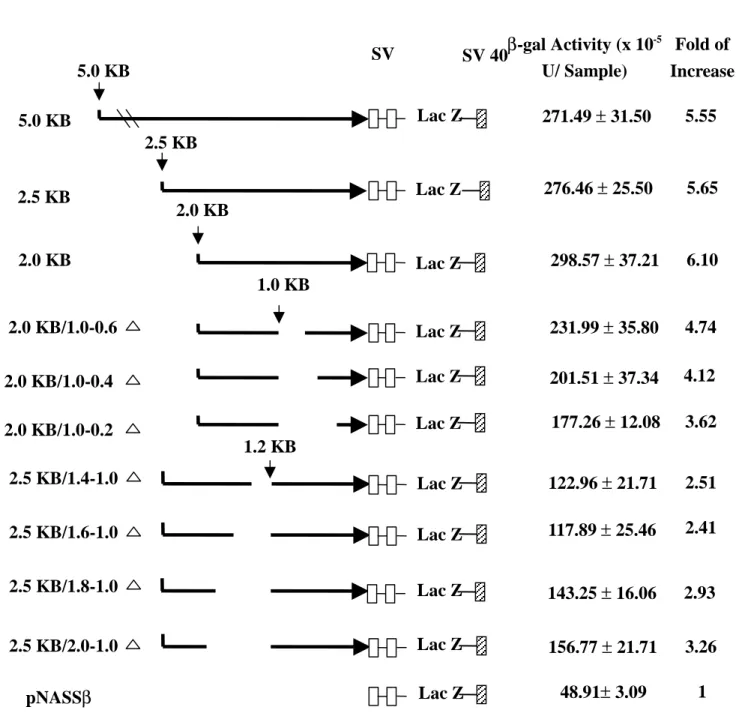
guns. The promoter activities of the various constructs obtained from 6 independent
experiments in rabbit corneal epithelial cells are summarized in Fig. 1. Data obtained
from β-gal activities of skins and conjunctivas (the same constructs as in Fig.1) were not significantly different from the activity of pNASSβ in conjunctival and cutaneous epithelia (data not shown).
The activity of pNASSβconstruct was assigned a relative level of 1.0. As described above, 5.0 kb, 2.5 kb, and 2.0 kb fragments displayed similar β-gal activities, and were significantly more than the activity of pNASSβ (Student’s t-test, p<0.05). Deletions of 445, 645, and 845 bp from –975 bp to 3’ end respectively did significantly
lower β-gal activities of 2.0 kb constructs (Student’s t-test, p<0.05). 5' deletion of 402 and 600 bp from –1014 bp to 5’end in 2.5 kb constructs resulted in further decreasing in
β-gal activity compared with (Student t-test, p<0.05), suggesting the presence of a enhancer-like element(s) between 1613 bp and 1014 bp, especially from –1416 bp
from –2015 bp to –1014 bp, rescued some β-gal activities (Sudent’s t-test, p<0.05). These results suggested that the region between 2015 bp and 1613 bp contains a
silencer-like element(s).
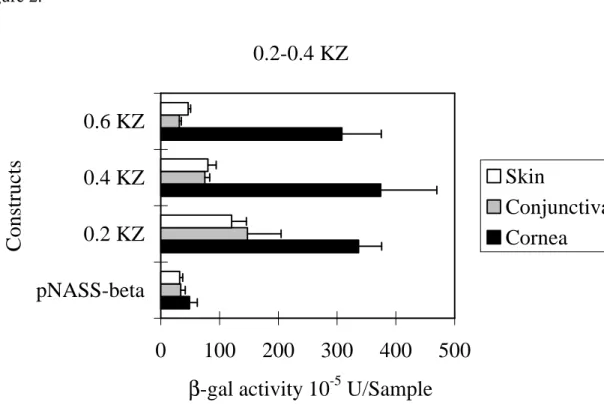
We further tested three constructs, 527 bp to +40 bp, -327 bp to +40 bp and
127 bp to +40 bp (Fig. 2), to determine whether the presence of tissue-specific elements
existed between –527 bp to +40 bp of Krt1.12 promoter. We found that the 0.2 kb
construct, -127 bp to +40 bp, did not express the tissue specificity (Fig 2. and Fig. 3A).
However, the tissue specificity could be found in 0.4 kb (-327 bp to +40 bp) and 0.6 kb
constructs (- 527 bp to +40 bp) (Fig. 2, Fig 3B. and Fig 3C). We suggested that there
might be a tissue-specific element(s) in the region from –327 bp to +40 bp.
DNase I Footpr inting Assay
Four probes (-1415 bp to –1167 bp , -599 to-375 bp, -394 bp to –131 bp, and –331 bp
to –109 bp) were used to identify the possible transcriptional factor binding sites
according to results functional analysis in the promoter of krt1.12 gene. There was two
continuous binding regions from –182 bp to –110 bp and from –256 bp to –193 bp
could be found in the DNase I binding assays (Fig.4). There was no binding site could
be found in another two probes (~-1415 bp to –1167 bp, -599 bp to-375 bp). From these
for some fundamental promoter activities of Krt1.12 gene, but this region was not for
the tissue specificity. However, the fragment form –256 bp to –193 bp might play an
important role of tissue specificity, and could also be found the constructs longer than
0.4 KB (-375 bp to +40 bp).
Electrophoretic Mobility Shift Assays (EMSA)
To characterize transcription factors that interact with the binding sites in the Krt1.12
promoter, EMSAs was performed. Synthetic oligonucleotides used in EMSAs (Table 3)
were designed from the regions protected by nuclear proteins. EMSA of site from –182
to -111 and –256 to –193 showed binding complex with nuclear proteins from nuclear
proteins of corneal epithelial cells, but the band could also be detected with nuclear
proteins from retinal cells (Fig. 5) This complex was competed and eliminated by a
400- or 800-fold molar excess of the unlabeled oligonucleotides termed self-competitor.
These results suggested that there were several transcriptional factors acting with
oliogonucleotides and regulating the tissue specificity of Krt1.12 promoter.
In the regard of PAX-6 elements, three segments of synthetic probe
corresponding to the –942 bp to –913 bp, -1661 bp to –1624 bp, and -1887 bp to –1858
bp respectively, were used to test the EMSAs. We observed that the first segment gave
a nonspecific binding with nuclear extracts, the second segment could bind the nuclear
Discussion
A 0.4 kb Kr t1.12 Pr omoter can expr ess Cor neal Specificity
In this paper, we showed those constructs including 5.0 kb, 2.5 kb, 2.0 kb, 0.6 kb, 0.4 kb,
and 0.2 kb of Krt1.12 promoter displayed similar activities on corneal epithelial cells
(Fig. 1 and Fig. 2). Except the 0.2 kb construct, all of them expressed minimal activities
on conjunctival and cutaneous epithelial cells. This result strongly suggests that a 0.2
kb of Krt1.12 promoter can efficiently express the promoter activities in epithelial cells
and is consistent with our previous findings.30
However, this 0.2 kb construct could not
display corneal specificity (Fig. 2 and Fig. 3). The corneal specificity could only occur
in constructs longer than 0.2 kb. We further used deletion constructs to confirm the
tissue specificity. When we deleted 0.4 kb, 0.6 kb, and 0.8 kb from 1.0 kb in a 2.0 kb of
Krt1.12 construct, we observed that the promoter activities decreased gradually and the
corneal specificity disappeared in the 0.8 kb deletion construct (Fig. 1 and Fig 3). This
further confirmed the region from 0.2 kb to 0.4 kb could regulate the expression of
corneal specificity and implied there also exists some enhancer elements from 0.2 kb to
1.0 kb.
DNase I Footpr inting and EMSAs
To further elucidate the actual transcriptional binding sites in these regions, DNase I
to –193 bp and –182 bp to –111 bp) could be protected by nuclear extracts and these
two fragments could also be confirmed by EMSAs. However we also found that retinal
nuclear extracts could also bind both segments. Combining above data, we suggested
that these two segments might be responsible for the expression of corneal tissue
specificity and the sequence proximal to 3’ end of these regions played a role in the
regulation of Krt1.12 promoter activity in epithelial cells.
By searching the Gene Bank (NCBI GenBank database, Blast web client
software), we found that there were eight possible transcriptional factor binding sites
including AP-1 (-240 bp to –250 bp), VPB (-229 bp to -238 bp), CP-2 (-211 bp to -221
bp), S8 (-158 bp to -174 bp), GFI1 (-114 bp to -138 bp), AP1 (-124 bp to -133 bp),
DELTAFF1 (-125 bp to -136 bp), AP4 (-113 bp to -122 bp) binding site in sense
direction of these areas. In the antisense orientation, we also found there were eleven
possible transcriptonal binding sites including AP4 (-113 bp to -122 bp), PADS (-123
bp to -132 bp), OCT1 (-128 bp to -153 bp), CEBPB (-133 bp to -147 bp), OCT1 (-119
bp to -133 bp), CATA-1 218 bp to -231 bp), CEBP 226 bp to -243 bp), CEBP B
(-229 bp to -242 bp), GKLF (-234 bp to -248 bp), BARBIE (-236 bp to -251 bp), and
GKLF (-239 bp to -253 bp) binding sites. From these binding sites, we speculated that
the regulation of Krt1.12 promoter in the expression of corneal specificity is the result
by previous studies of other cytokeratin genes.21;22
Among these transcriptional factors, AP-1 (activation protein 1) was well known
to be a regulator of many cytokeratin genes including K1, K3, K5, K8, K10, and K18.6 33-37
The regulation may be a basal activity or response to stimuli, e.g. calcium, vitamin
D, and steroid hormone, through the protein kinase C (PKC) or mitogen activated
protein kinase (MAPK) pathways.38-42
But the roles of other transcriptional factors in
the regulation of cytokeratin gene were still unknown.
Form the deletion analysis, we suggested that there were enhancer element(s)
from –975 bp to –534 bp and –1416 bp to –1014 bp. Probes (-1415 bp to –1167 bp and
-599 bp to-375 bp) were also performed to study the binding with nuclear extracts, but
we still could not find any protection region in these probes. We don’t know what kind
of transcriptional factors in these areas due to too broad to be analyzed by specific
probes in EMSAs. But from our previous studies, we found that co-transfection of
Pax-6 cDNA with K12 promoter-β-gal constructs result in 4-fold increase of β-gal activities.30
Other studies also demonstrated that Pax-6 played an important role in the
regulation of eye-specific gene in lens, iris epithelium, and retina.43-45
We constructed
three probes as described above for EMSAs. We found that the probe from –1661 bp
to –1624 bp, which contains two Pax-6 binding sites, could specifically bind the corneal
epithelium, but also can be expressed in other ocular tissue.46
We suggested that this
Pax-6 segment enhanced the promoter activity of K12 gene, but might be not involved
in the regulation of corneal specificity.
In conclusion, we found the upstream sequence (-182 bp to –111 bp and –256 bp
to –193 bp) of Krt1.12 promoter contains the capability to regulate the corneal
specificity. A minimal 110 bp (0.2 kb) of Krt1.12 promoter can efficiently drive the
basal activity in corneal and conjunctival epithelial cells. The regulation of corneal
specificity might be from corporation of several transciptional factors. The Pax-6
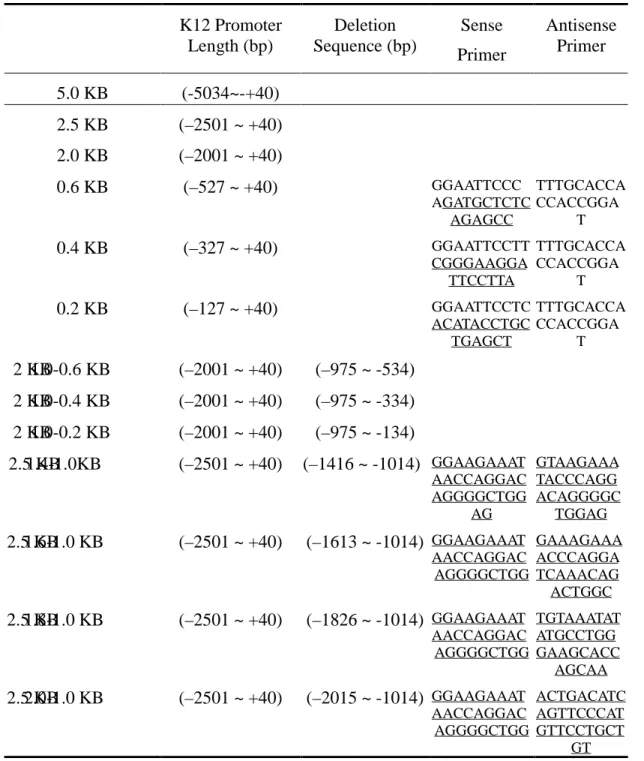
Table 1. Plasmid Constructs Used in Gene Gun Assay and Probes Used for Footprinting K12 Promoter Length (bp) Deletion Sequence (bp) Sense Primer Antisense Primer 5.0 KB (-5034~-+40) 2.5 KB (–2501 ~ +40) 2.0 KB (–2001 ~ +40) 0.6 KB (–527 ~ +40) GGAATTCCC AGATGCTCTC AGAGCC TTTGCACCA CCACCGGA T 0.4 KB (–327 ~ +40) GGAATTCCTT CGGGAAGGA TTCCTTA TTTGCACCA CCACCGGA T 0.2 KB (–127 ~ +40) GGAATTCCTC ACATACCTGC TGAGCT TTTGCACCA CCACCGGA T 2 KB 1.0-0.6 KB (–2001 ~ +40) (–975 ~ -534) 2 KB 1.0-0.4 KB (–2001 ~ +40) (–975 ~ -334) 2 KB 1.0-0.2 KB (–2001 ~ +40) (–975 ~ -134) 2.5 KB 1.4-1.0KB (–2501 ~ +40) (–1416 ~ -1014) GGAAGAAAT AACCAGGAC AGGGGCTGG AG GTAAGAAA TACCCAGG ACAGGGGC TGGAG 2.5 KB 1.6-1.0 KB (–2501 ~ +40) (–1613 ~ -1014) GGAAGAAAT AACCAGGAC AGGGGCTGG GAAAGAAA ACCCAGGA TCAAACAG ACTGGC 2.5 KB 1.8-1.0 KB (–2501 ~ +40) (–1826 ~ -1014) GGAAGAAAT AACCAGGAC AGGGGCTGG TGTAAATAT ATGCCTGG GAAGCACC AGCAA 2.5 KB 2.0-1.0 KB (–2501 ~ +40) (–2015 ~ -1014) GGAAGAAAT AACCAGGAC AGGGGCTGG ACTGACATC AGTTCCCAT GTTCCTGCT GT
Table 2. Primers Used for Dnase I Footprinting
Probe Length (bp) Sense Primer Antisense Primer -1415~-1167 CCCCTGTCCTG GGTATTTCTTA GAGAGTTGGTCTCA AGGTCAGT -599~-375 ACTTCCAAACA GCTGGCTC CGGTTCAGTATAAG CCAACC -394~-131 GGTTGGCTTATA CTGAACCG TGCATCGTGAAAAT TCACTG -331~-109 GGCTTCGGGAA GGATCCTT CCAGCTCAGCAGGT ATGTGA
Table 3. Probes for Electrophoretic Motility Shift Assays
Probe Length (bp) Sequence
-256~-193 ACTGAAGGTGACAGATTCCTTACGACAGCCTA TCTGCTCCACCCAGCCTTCTTTCTTGTGTGTC -182~-111 TGGTAATGGTTATTCGATTATAGCTATATCAGTGAAT TTTCACGATGCATAAATCACATACCTGCTGAGCTG Pax 6 (-1887~-1858) AAATGAGGCAAGTGGATTGCAGACTGTGT Pax 6 (-1661~-1624) TTTTCAAACACTTTCCCAGGGTCAGGAACA Pax 6 (-942~-913) CGTTTTATTAAATTCCTGTGAATTCTTTGG
Figure 1. SV 40 SD/S A SV 40 Poly A+ 2.5 KB 2.0 KB/1.0-0.6 1.0 KB 2.0 KB/1.0-0.4 2.0 KB 2.0 KB/1.0-0.2 2.5 KB 2.0 KB 5.0 KB Lac Z Lac Z Lac Z Lac Z Lac Z Lac Z 5.0 KB KB β-gal Activity (x 10-5 U/ Sample) Fold of Increase 271.49 ± 31.50 276.46 ± 25.50 298.57 ± 37.21 231.99 ± 35.80 201.51 ± 37.34 177.26 ± 12.08 5.55 5.65 6.10 4.74 4.12 3.62 2.5 KB/1.4-1.0 2.5 KB/1.6-1.0 2.5 KB/1.8-1.0 2.5 KB/2.0-1.0 Lac Z Lac Z Lac Z Lac Z 122.96 ± 21.71 117.89 ± 25.46 143.25 ± 16.06 156.77 ± 21.71 2.51 2.41 2.93 3.26 1.2 KB pNASSβ Lac Z 48.91± 3.09 1
Figure 2.
0.2-0.4 KZ
0
100
200
300
400
500
pNASS-beta
0.2 KZ
0.4 KZ
0.6 KZ
Constructs
β
-gal activity 10
-5U/Sample
Skin
Conjunctiva
Cornea
Reference List
1. Moll R, Franke WW, Schiller DL et al. The catalog of human cytokeratins:
patterns of expression in normal epithelia, tumors and cultured cells. Cell 1982;
31:11-24.
2. Hollenberg SM, Sternglanz R, Cheng PF et al. Identification of a new family of
tissue-specific basic helix-loop- helix proteins with a two-hybrid system.
Mol.Cell Biol. 1995; 15:3813-22.
3. Quinlan RA, Schiller DL, Hatzfeld M et al. Patterns of expression and
organization of cytokeratin intermediate filaments. Ann.N.Y.Acad.Sci. 1985;
455:282-306.
4. Steinert PM, Roop DR. Molecular and cellular biology of intermediate filaments.
Annu.Rev.Biochem. 1988; 57:593-625.
5. Cooper D, Sun TT. Monoclonal antibody analysis of bovine epithelial keratins.
Specific pairs as defined by coexpression. J.Biol.Chem. 1986; 261:4646-54.
6. Wu RL, Galvin S, Wu SK et al. A 300 bp 5'-upstream sequence of a
differentiation-dependent rabbit K3 keratin gene can serve as a
7. Chaloin-Dufau C, Sun TT, Dhouailly D. Appearance of the keratin pair K3/K12
during embryonic and adult corneal epithelial differentiation in the chick and in
the rabbit. Cell Differ.Dev. 1990; 32:97-108.
8. Cooper D, Schermer A, Sun TT. Classification of human epithelia and their
neoplasms using monoclonal antibodies to keratins: strategies, applications, and
limitations. Lab Invest 1985; 52:243-56.
9. Kao WW, Liu CY, Converse RL et al. Keratin 12-deficient mice have fragile
corneal epithelia. Invest Ophthalmol.Vis.Sci. 1996; 37:2572-84.
10. Irvine AD, Corden LD, Swensson O et al. Mutations in cornea-specific keratin
K3 or K12 genes cause Meesmann's corneal dystrophy. Nat.Genet. 1997;
16:184-87.
11. Wei ZG, Wu RL, Lavker RM et al. In vitro growth and differentiation of rabbit
bulbar, fornix, and palpebral conjunctival epithelia. Implications on conjunctival
epithelial transdifferentiation and stem cells. Invest Ophthalmol.Vis.Sci. 1993;
34:1814-28.
12. Liu JJ, Kao WW, Wilson SE. Corneal epithelium-specific mouse keratin K12
13. Zhu G, Ishizaki M, Haseba T et al. Expression of K12 keratin in alkali-burned
rabbit corneas. Curr.Eye Res. 1992; 11:875-87.
14. Liu CY, Zhu G, Converse R et al. Characterization and chromosomal localization
of the cornea-specific murine keratin gene Krt1.12. J.Biol.Chem. 1994;
269:24627-36.
15. Chen WY, Mui MM, Kao WW et al. Conjunctival epithelial cells do not
transdifferentiate in organotypic cultures: expression of K12 keratin is restricted
to corneal epithelium. Curr.Eye Res. 1994; 13:765-78.
16. Wu RL, Zhu G, Galvin S et al. Lineage-specific and differentiation-dependent
expression of K12 keratin in rabbit corneal/limbal epithelial cells: cDNA cloning
and northern blot analysis. Differentiation 1994; 55:137-44.
17. Moyer PD, Kaufman AH, Zhang Z et al. Conjunctival epithelial cells can
resurface denuded cornea, but do not transdifferentiate to express cornea-specific
keratin 12 following removal of limbal epithelium in mouse. Differentiation 1996;
60:31-38.
18. Fuchs E. Epidermal differentiation: the bare essentials. J.Cell Biol. 1990;
19. Rice RH, Green H. The cornified envelope of terminally differentiated human
epidermal keratinocytes consists of cross-linked protein. Cell 1977; 11:417-22.
20. Stoler A, Kopan R, Duvic M et al. Use of monospecific antisera and cRNA probes
to localize the major changes in keratin expression during normal and abnormal
epidermal differentiation. J.Cell Biol. 1988; 107:427-46.
21. Eckert RL, Welter JF. Transcription factor regulation of epidermal keratinocyte
gene expression. Mol.Biol.Rep. 1996; 23:59-70.
22. Eckert RL, Crish JF, Banks EB et al. The epidermis: genes on - genes off. J.Invest
Dermatol. 1997; 109:501-09.
23. Schermer A, Galvin S, Sun TT. Differentiation-related expression of a major 64K
corneal keratin in vivo and in culture suggests limbal location of corneal epithelial
stem cells. J.Cell Biol. 1986; 103:49-62.
24. Sun TT, Tseng SC, Huang AJ et al. Monoclonal antibody studies of mammalian
epithelial keratins: a review. Ann.N.Y.Acad.Sci. 1985; 455:307-29.
25. Schermer A, Jester JV, Hardy C et al. Transient synthesis of K6 and K16 keratins
in regenerating rabbit corneal epithelium: keratin markers for an alternative
26. Weiss RA, Eichner R, Sun TT. Monoclonal antibody analysis of keratin
expression in epidermal diseases: a 48- and 56-kdalton keratin as molecular
markers for hyperproliferative keratinocytes. J.Cell Biol. 1984; 98:1397-406.
27. Leask A, Byrne C, Fuchs E. Transcription factor AP2 and its role in
epidermal-specific gene expression. Proc.Natl.Acad.Sci.U.S.A 1991; 88:7948-52.
28. Leask A, Rosenberg M, Vassar R et al. Regulation of a human epidermal keratin
gene: sequences and nuclear factors involved in keratinocyte-specific
transcription. Genes Dev. 1990; 4:1985-98.
29. Byrne C, Fuchs E. Probing keratinocyte and differentiation specificity of the
human K5 promoter in vitro and in transgenic mice. Mol.Cell Biol. 1993;
13:3176-90.
30. Shiraishi A, Converse RL, Liu CY et al. Identification of the cornea-specific
keratin 12 promoter by in vivo particle-mediated gene transfer. Invest
Ophthalmol.Vis.Sci. 1998; 39:2554-61.
31. Andrews NC, Faller DV. A rapid micropreparation technique for extraction of
DNA-binding proteins from limiting numbers of mammalian cells. Nucleic Acids
32. Morris TA, Fong WB, Ward MJ et al. Localization of upstream silencer elements
involved in the expression of cone transducin alpha-subunit (GNAT2). Invest
Ophthalmol.Vis.Sci. 1997; 38:196-206.
33. Oshima RG, Abrams L, Kulesh D. Activation of an intron enhancer within the
keratin 18 gene by expression of c-fos and c-jun in undifferentiated F9 embryonal
carcinoma cells. Genes Dev. 1990; 4:835-48.
34. Takemoto Y, Fujimura Y, Matsumoto M et al. The promoter of the endo A
cytokeratin gene is activated by a 3' downstream enhancer. Nucleic Acids Res.
1991; 19:2761-65.
35. Tamai Y, Takemoto Y, Matsumoto M et al. Sequence of EndoA gene encoding
mouse cytokeratin and its methylation state in the CpG-rich region. Gene 1991;
104:169-76.
36. Casatorres J, Navarro JM, Blessing M et al. Analysis of the control of expression
and tissue specificity of the keratin 5 gene, characteristic of basal keratinocytes.
Fundamental role of an AP-1 element. J.Biol.Chem. 1994; 269:20489-96.
37. Lu B, Rothnagel JA, Longley MA et al. Differentiation-specific expression of
human keratin 1 is mediated by a composite AP-1/steroid hormone element.
38. Pulverer BJ, Kyriakis JM, Avruch J et al. Phosphorylation of c-jun mediated by
MAP kinases. Nature 1991; 353:670-74.
39. Boyle WJ, Smeal T, Defize LH et al. Activation of protein kinase C decreases
phosphorylation of c-Jun at sites that negatively regulate its DNA-binding
activity. Cell 1991; 64:573-84.
40. Su B, Jacinto E, Hibi M et al. JNK is involved in signal integration during
costimulation of T lymphocytes. Cell 1994; 77:727-36.
41. Gille H, Sharrocks AD, Shaw PE. Phosphorylation of transcription factor
p62TCF by MAP kinase stimulates ternary complex formation at c-fos promoter.
Nature 1992; 358:414-17.
42. Marais R, Wynne J, Treisman R. The SRF accessory protein Elk-1 contains a
growth factor-regulated transcriptional activation domain. Cell 1993; 73:381-93.
43. Grindley JC, Davidson DR, Hill RE. The role of Pax-6 in eye and nasal
development. Development 1995; 121:1433-42.
44. Cvekl A, Kashanchi F, Sax CM et al. Transcriptional regulation of the mouse
alpha A-crystallin gene: activation dependent on a cyclic AMP-responsive
45. Gopal-Srivastava R, Cvekl A, Piatigorsky J. Pax-6 and alphaB-crystallin/small
heat shock protein gene regulation in the murine lens. Interaction with the
lens-specific regions, LSR1 and LSR2. J.Biol.Chem. 1996; 271:23029-36.
46. Koroma BM, Yang JM, Sundin OH. The Pax-6 homeobox gene is expressed
throughout the corneal and conjunctival epithelia. Invest Ophthalmol.Vis.Sci.