J. Exp. Mar. Biol. Ecol., 173 (1993) 57-69
0 1993 Elsevier Science Publishers B.V. All rights reserved 0022-0981/93/$06.00
57
JEMBE 02019
The effects of flow on feeding of three gorgonians from
southern Taiwan
Chang-Feng Dai
and
Ming-Chao LinInstitute of Oceanography, National Taiwan University. Taipei, Taiwan, ROC (Received 30 January 1992; revision received 7 June 1993; accepted 11 June 1993)
Abstract: The feeding performances of three gorgonians, Subergorgia suberosa, Melithaea ochracea, Acan- thogorgia vegae, in different flow regimes were studied. The three gorgonians expanded their polyps and fed in varying ranges of flow velocities. Differences in the feeding range are possibly related to polyp morpholo- gies. S. suberosa, which has tall polyps and encounters higher drag, is more easily deformed in currents; it feeds in a narrow range of flow velocities (7-9 cm’s -I). M. ochracea, which has short polyps and encoun- ters lower drag, is less readily deformed in currents; it feeds in a wider range of flow velocities (4-40 cm’s -‘). Polyp heights of A. vegae are intermediate and this coral feeds in currents of 2-22 cm.s-‘. The upper lim- its on the flow ranges are considered to be determined by the balance between the energy gained from feeding and the cost of keeping the polyps expanded. Optimal feeding rates of the three gorgonians were found at moderate flow velocities. Feeding rates increased initially with flow velocity, reached a peak value at 8 cm’s ‘, then decreased with increasing flow velocities. Both the colony feeding effectiveness and the polyp feeding effectiveness at 8 cm’s_’ are ranked: S. suberosa > A. vegae > M. ochracea.
Key words: Gorgonacea; Suspension feeding; Water flow
INTRODUCTION
Feeding is an essential activity for most animals because it provides the basic en- ergy for sustaining life. Passive suspension feeders such as octocorals depend on exogenous current for food delivery and waste disposal; thus their distributions are often related to flow velocity of the environment (Jorgenson, 1955; Riedl, 197 1; Koehl, 1977; Muzik & Wainwright, 1977; Sebens, 1984). Current-induced drag forces can dislodge organisms (Riedl, 1971; Wainwright et al., 1976) and interrupt their particle capture activities (Patterson, 1984; Best, 1988) while low flow velocities may result in food deficiency (Harvell & LaBarbera, 1985). The conflicting demands of maximizing feeding efficiency and minimizing drag forces often exist in passive suspension feed- ers. Habitats where flow is most often within the optimal range offer the greatest ad- vantage of feeding and survival for marine sessile organisms. As a consequence, ma- rine sessile organisms often live and feed in a particular range of current velocities.
Functional constraints arising from the conflicting demands of maximizing feeding Correspondence address: C.-F. Dai, Institute of Oceanography, National Taiwan University, Taipei 10764, Taiwan, ROC.
and Ill~nirnizing drag are often characterized by certain lnorphologic~~~ features ofsessile organisms. The effect of flow velocity on feeding abilities has been the focus of several investigations (Best, 1988; Okamura. 1990; Patterson. 1991). But the combined influ- ences of flow velocities and morphological characters 4x1 feeding abilities has received little aLtention.
~orgonians (Anthozoa: Gorgonacea) are one of the major bcnthic ~lrgan~sln of tropical coral reefs. They display a variety of coionq morphologics and occupy vari- ous habitats on reefs (Bayer, 1981). Colony morphology and orientations of gorgon- ians are related to current regimes (Wainwright & Dillon, 1969; Crigg, 197 1; Leversec. 1976). Colonies occurring in uni- or bidirectional currents are often planar and oriented ~rpendicular to the prevailing current direction. Several studies have demonstrated the effects offlow velocities on feeding abilities ofgorgonians (Leversec. 1976; L,asker et al., 1983; Sponaugle & LaBarbera, 1991). Lasker ( 198 1) compared the feeding abilities of three Caribbean gorgonians in experiments conducted in feeding chambers or in flow tanks under relatively low current speeds (3.5 and 7.3 cm’s ‘I). Sponaugle & LaBar- bcra (1991) studied the feeding rates of two Caribbean gorgonians with similar mor- phalogics in a range of current velocities. However, the relative feeding abilities of morphologically different species in various flow velocities have rarely been studied.
The purpose of this study is twofold: (1) to examine the eRects of flow velocities on the shape and feeding performance of three gorgonians with different colony and polyp morphologies, and (2) to utilize laboratory results to account for the distribution pat- tern of the three species on reefs. Three gorgonian species, S~berg#~gii~ .~iib~r~~~l (Pallas.
1776). ,4 ~~~~~th~g~rgi~/ wgrte Aurivillus, 193 1, and M~~ith~~e~~ u~hr~~e~~ Linnaeus, 1758. tvcrc selected for this study. Colonies of S. .cuhero.wr are often planar or bush) with the plane perpendicular to the current direction. Branches are separated by as much tls 3 cm. They often grow on the lower part of reef slopes or on boulders scattered over a sandy bottom. A. wgae colonies arc flab&la&d and usually grow on the lateral side of blocks or reef fronts. Colonies of M. (~~~~~z~eff are ~~ultiplaxlar and often exist on upper part. of reef fronts or boulders where the current is relatively strong. The plane of the fan is oriented perpendicular to the direction of current. All the three species are widely distributed throughout the fringing reefs of southern Taiwan (Chcn c% Chang,
1991) and on Pacific coral reefs (Muzik & Wainwright, 1977).
MATERIALS AND METHODS
Colonies of the three species, each about 15 cm high, were collected from Nanwan Bay, southern Taiwan 120” 44’ E, 21’ 57’ N). Specimens were collected on the same reef but from different depths and microhabitats. Species identifications followed Chen & Chang (1991) and related references (Muzik & Wainwright, 1977; Zou & Scott,
1980; Bayer, 198 1).
FLOW AND FEEDING OF THREE GORGONIANS 59
acclimation before the experiment. The acclimation tank was furnished with two cur- rent generators which create consistent flow and the coral colonies were fed with
Artemia
nauplii.Morphometric studies of the gorgonians were conducted in lab by using a digitizer (Lab Visions LV-1) associated with a personal computer. Colonies were immersed in a sea-water tank and their projected areas, number of branches, and total lengths of branches were measured. The total number of polyps was estimated by counting polyps in 10 randomly selected 1 cm segments from a colony. Polyp length and oral disk di- ameter were measured under a stereomicroscope when the polyps were fully expanded.
A recirculating flow tank of 75-l capacity (150 cm long, 60 cm wide and 20 cm high) was made with reference to Leversee (1976). The tank was filled with 69 1 filtered seawater before experiment. A laboratory stirrer (Her-Cheng SC-VS35W) with a pro- peller (14 cm in diameter) was used to generate water flow. The flow velocity was regulated by a solid state motor control and measured by an electromagnetic current meter (Kenek VM-401H). The water temperature was maintained at 24 “C.
When flow velocity and water temperature were steady, a coral colony starved for at least 24 h was transferred into the tank and oriented perpendicular to the flow di- rection. The coral colony was allowed 3 h for further acclimation in such steady cur- rent regime. The range of flow velocities in which a coral colony expands its polyps
TABLE I
Morphornetric data (mean 5 1 SD) of colony A and B of the three gorgonians (colony height = 15 cm). Polyps are measured in fully expanded condition.
Colony A
Subergorgia Acanthogorgia
suberosa vegae
Melithaea ochracea Total number of branches
Total length of branches (cm) Projected colony area (cm’) Total number of polyps Polyp density (polys cm _ “) Polyp height (mm) Polyp diameter (mm) 8 63 134 57.5 231.4 416.2 56.3 92.3 167.3 1127k63 4628 k 440 18501t1192 20.0 50.1 110.6 5.32 + 0.26 3.02 k 0.25 0.96 + 0.18 5.12 k 0.24 2.96 k 0.18 0.95 * 0.15 Colony B
Total number of branches 11 84 152
Total length of branches (cm) 61.1 264.5 496.6
Projected colony area (cm2) 65.8 107.4 173.9
Total number of polyps 1354 + 74 4913 + 555 19814k 1242
Polyp density (polys . cm *) 20.6 46.3 113.9
Polyp height (mm) 5.19+0.31 3.07 + 0.31 0.95 f 0.20
was determined by treating each colony to a particular flow velocity for 3 h. Each colony was tested in a range 0 to 40 cm.s-’ of flow velocities and the sequence was randomly assigned to the colony.
Feeding experiments were conducted in the range of flow velocities in which the coral expanded its polyps. Living Artemiu nauplii were harvested from culture 24 h after hatching. 1400 Artemiu nauplii were counted by naked eye using a 10 ml pipette, di- luted to 1 1, and poured evenly into the flow tank. During the following 6 h, the num- ber ofArtemia remaining in the tank was counted at l-h interval. The number ofArtemiu was determined by taking five l-l samples using a l-l beaker. Samples were poured through a plankton net (200 pm) and the Artemiu were counted on the netting. The
Artemih were returned to the tank along with the tittered seawater after counting. Two colonies were tested for each species. Feeding experiments were made 48 h apart to allow complete clearing of Artemb from the polyp guts.
Feeding rates of the three gorgonians were normalized by number of polyps 01 surface area to compare their feeding effectiveness. Two normalization procedures were taken following Hunter (1989): (1) colony feeding effectiveness, a measure of feeding rate per unit area, and (2) polyp feeding effectiveness, a measure of feeding rate pet
100 polyps. Because feeding rates during the first hour were most noticeable, these were used to represent colony feeding abilities.
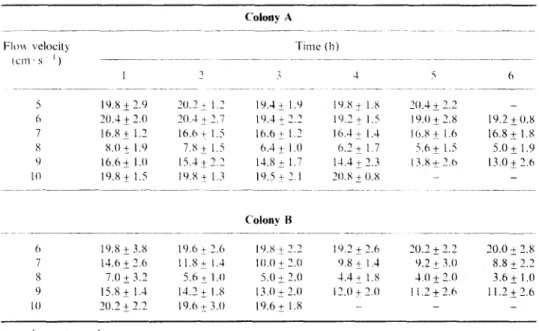
TABLE II
Residual concentration (shrimps I ’ ) or 4rfcwio nauplii (mwn 5 I SD) in a range ul flou velocities in the 6-h period of feeding experiment for Strher~or& suheroscr. Initial concentration was 20 shrimps I ’
Colony H -___
6 19.8 If- 3.8 IO.6 t 2.6 19.X +_ 2.2 19.2 k 2.6 30.2 k 2.2 20.0 * 2.x 7 14.6 k 2.6 11.8? I.4 10.0 k 3.0 9.8 i I .J 9.2 + 3.0 8.8 t 2.7 8 7.0 2 3.2 5.6 I 1 .I) 5.0 & ‘.O 1.4i 1.X 4.0 I2.0 3.6 + I.0 9 15.8 2 1.4 14.7 + 1.x 13.0 i_ 1.0 12.0 + 2.0 Ii.?_t 2.6 I l.Zk 3.6
IO 20.2 + 2.2 19.6 t 3.0 19.6& 1.X _
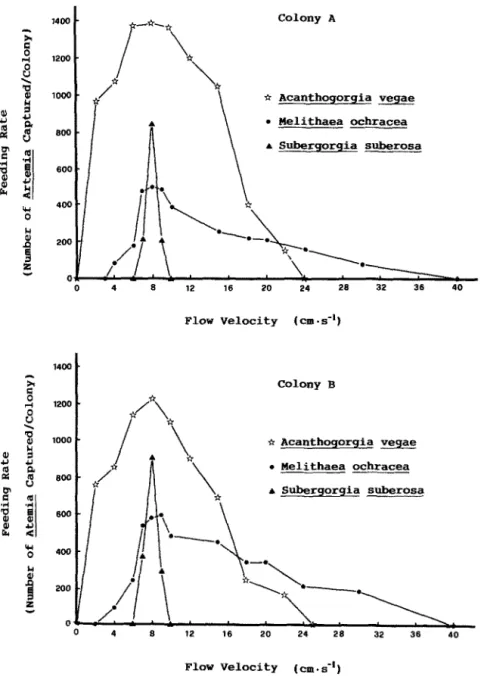
FLOW AND FEEDING OF THREE GORGONIANS 61 Colony A TV Acanthogorgia vegae A Subergorgia suberosa Flow Velocity (cm-s-') Colony B or Acanthogorgia veqae . Melithaea ochracea A Subergorgia suberosa 0 4 8 ‘12 16 20 24 28 32 38 40 Flow Velocity (cm.s-')
Fig. 1. Feeding rates (number of Artemiu captured per colony) during the first hour of exposure to Artemia nauplii of the three gorgonians (Subergorgia suberosa; Acanthogorgia vegae; Melithuea ochracea) in a range
of flow velocities.
To detect the natural mortality and precipitation of
Artemiuduring the experiment,
blank tests were conducted at the flow velocities of 8 and 40 cm-s-’ under the con-
dition that no coral colony was introduced to the tank.
62 C.-D. DA1 AND M.-c‘. LIN
RESULTS
MORPHOMETRICS
Morphometric data of the two colonies of the three gorgonian species are shown in Table I. Among the three species studied, S. mberosa has the largest polyps and the lowest polyp density. M. ochracea has the smallest polyps and the highest polyp den- sity. Polyp size and polyp density of A. vegur were intermediate among the three species. The projected area of the three species were: A. vegue> M. ochraceu> S. .subero.w.
FEEDING EXPERIMENTS
The results of the blank test showed that there was no significant change in num- ber of Artemia nauplii during the 6 h experimental period. The regression slope was not
TABLE 111
Residual concentration (shrimps I ‘) of Arfema naupbi (mean & 1 SD) m a range of flow ~elocitics in the 6-h period of feeding experiment for Accrnthogorgiu vegcrr. initial concentration was 20 shrimps. I ‘.
Colony A 7.2 + 1.0 2.7 i 0.4 0.8 _t 0.2 5.0 IO.8 1.5 + 0.5 0.4 f 0. I 0.3 5 0.0 0.0 * 0.0 0.0 & (1.0 0.0 + 0.0 0.0 + 0.0 (1.0 + 0.0 O.h_+O.l 0.0 + 0.0 0.0 f 0.0 2.7 + 0.4 0.6 t 0.1 0. 1 2 0.0 7.25 I.1 2.6 + 0.4 1.0 t 0.2 14.3 * 1.7 10.3 _t 0.3 7.1 5 0.9 17.0 2 1.7 17.oi 1.7 13.1 & 1.3 ___~__. ~~~. .-..-._~~~. _~~~ ___ Colony B 0.4 t 0.2 0.2 2 0.0 0.0 & 0.0 0.0 f 0.0 0.0 _c 0.0 0.0 & 0.0 0.4 + 0. I 5.9 & 1.0 11.7iO.6 0.2 _c 0. 1 0. I ?_ 0.0 0.0 + 0.0 0.0 2 0.0 0.0 + 0.0 0.0 f 0.0 0.‘_c0.l 5.0 i; 0.Y 10.7 * 1.3 3 0.0 * 7.8
co+-
1.2 4.6 + I .x 1.0 + 2.1) 3.0 i 0.6 2.4 i 0.2 1 7.6 + 2.6 5.X 2 I.8 3.4 i 1.0 2.8 + 0.6 2.6; 0.6 2.0 & 0.8 6 3.6 + I .O 2.2 + 0.6 2.6 0.6 2.2 0.6 2.0 0.8 1.4 0.6 8 2.4 _t 0.6 1.6 I i +_ 2 * I.0 I.0 ) 0.8 1.6_+0.4 0.6 f 0.6 0.2 _+ 0.2 IO 1.2 2 0.6 2.8 + I.3 2.6 t I.0 2.0 + 0.4 2.0 _t 0.4 1.4 i 0.6 I2 7.0 * I .2 5.6 i_ 2.2 1.x i 11.6 3.x + I.1 2.x + 0.6 2.0 If 0.8 15 10.0 + 2.0 5.8 f 0.6 3.2 c 0.6 2.8 + 0.6 2.6 k 1.0 2.4 i 0.218 16.42 1.0 13.0 it 2.8 11.2i_ 1.8 100 20.X Y.2 + I.8 9.2 + 0.6
FLOW AND FEEDING OF THREE GORGONIANS 63
significantly different than 0 (t = 0.57, n = 35, p> 0.05, at 8 cm.s-‘; t = 0.32, n = 35, p>O.O5, at 40 cm’s_‘), so natural mortality and precipitation of Artemiu shrimps during the experiment period can be neglected.
Colonies of S. suberosa expand their polyps at flow velocities from 5 to 10 cm.s-‘, but the feeding appear to occur between 7 and 9 cm.s -’ (Table II). A slight difference was found between the two colonies tested. Only some polyps of colony A were ex- panded at flow velocities of 5 and 6 cm*s -I, but the coral withdrew its polyps after exposure to Artemia for 5 h at 5 cm.s-‘. Polyps of colony B were not expanded at 5 cm.s-‘. Feeding was not detectable at 5 and 6 cm.s-’ for both colonies. At 7, 8, and 9 cm.s-‘, most of the polyps were fully expanded and feeding was detected. Feeding
TABLE IV
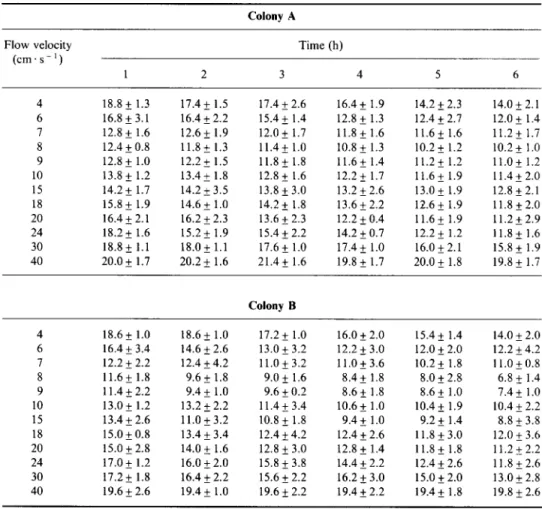
Residual concentration (shrimps. I - ‘) of Artemia nauplii (mean 2 1 SD) in a range of flow velocities in the 6-h period of feeding experiment for Melithaea ochrucea. Initial concentration was 20 shrimps. I ‘.
Colony A
Flow velocity Time (h)
(cm.s-‘) 1 2 3 4 5 6 4 6 8 9 10 15 18 20 24 30 40 18.8 + 1.3 17.4 * 1.5 17.4 + 2.6 16.4 + 1.9 14.2 f. 2.3 14.0 f 2.1 16.8 * 3.1 16.4 k 2.2 15.4+ 1.4 12.8 k 1.3 12.4 f 2.7 12.0 f 1.4 12.8 f 1.6 12.6 k 1.9 12.0 * 1.7 11.8 f 1.6 11.6* 1.6 11.2* 1.7 12.4 k 0.8 11.8 k 1.3 11.4* 1.0 10.8 f 1.3 10.2 f 1.2 10.2 f 1 .o 12.8 k 1.0 12.2 k 1.5 11.8 k 1.8 11.6* 1.4 ll.2* 1.2 11.02 1.2 13.8 t 1.2 13.4 k 1.8 12.8 f 1.6 12.2 f 1.7 11.6* 1.9 11.4* 2.0 14.2 k 1.7 14.2 + 3.5 13.8 f 3.0 13.2 f 2.6 13.0 f 1.9 12.8 + 2.1 15.8 k 1.9 14.6 k 1 .O 14.2 + 1.8 13.6 f 2.2 12.6 f 1.9 11.8k2.0 16.4k2.1 16.2 k 2.3 13.6 + 2.3 12.2 f 0.4 11.6* 1.9 11.2+2.9 18.2 k 1.6 15.22 1.9 15.4 * 2.2 14.2 f 0.7 12.2 f 1.2 11.85 1.6 18.82 1.1 18.0* 1.1 17.6 _I 1.0 17.4* 1.0 16.0 f 2.1 15.8 f 1.9 20.0 2 1.7 20.2 k 1.6 21.4+ 1.6 19.8 + 1.7 20.0 k 1.8 19.8 * 1.7 Colony B 4 18.6 f 1.0 18.62 1.0 17.2 f 1.0 16.0 f 2.0 15.4 f 1.4 14.0 * 2.0 6 16.4 f 3.4 14.6 k 2.6 13.0 f 3.2 12.2 f 3.0 12.0 + 2.0 12.2 + 4.2 7 12.2 f 2.2 12.4 + 4.2 11.0 f 3.2 11.0+3.6 10.2 f 1.8 11.0&0.8 8 11.6+ 1.8 9.6 + 1.8 9.0 f 1.6 8.4 f 1.8 8.0 + 2.8 6.8 + 1.4 9 11.4 + 2.2 9.4 + 1.0 9.6 f 0.2 8.6+ 1.8 8.6 f 1.0 7.4* 1.0 10 13.0 * 1.2 13.2 + 2.2 11.4k3.4 10.6 f 1 .O 10.4 f 1.9 10.4 * 2.2 15 13.4 k 2.6 11.0 k 3.2 10.8 + 1.8 9.4* 1.0 9.2+ 1.4 8.8 + 3.8 18 15.OkO.8 13.4k3.4 12.4 f 4.2 12.4 + 2.6 11.8&3.0 12.0 k 3.6 20 15.0 + 2.8 14.0 2 1.6 12.8 f 3.0 12.8 k 1.4 11.85 1.8 11.222.2 24 17.0 + 1.2 16.0 5 2.0 15.8 f 3.8 14.4 * 2.2 12.4 + 2.6 11.8k2.6 30 17.2 + 1.8 16.4 & 2.2 15.6 f 2.2 16.2 + 3.0 15.0 & 2.0 13.Ok2.8 40 19.6 k 2.6 19.4 + 1.0 19.6 f 2.2 19.4 * 2.2 19.4+ 1.8 19.8 k 2.6
6-l C’.-D. DA1 AND M-C. LIN
rates at 8 cm.s -’ were the highest (Fig. I). At IO cm’s -‘, the coral colonies expanded their polyps in the beginning, but feeding was not detectable. The colonies gradually withdrew their polyps after 3 h of exposure to Artemiu.
Both colonies ofA. vegue expanded their polyps-at flow velocities from 0 to 24 cm.s I and feeding could be detected at velocities between 2 and 22 cm.s’ ’ (Table III). The highest feeding rates occurred at 6-10 cm*s-’ (Fig. I).
Both colonies of M. ochruceu expanded their polyps at flow velocities from 4 to SO cm*s-’ and feeding was detected between 4 and 40 cm.s ~ ’ (Table IV). At flow velocities between 40 and 50 cm‘s --‘, the coral polyps were fully expanded but no feeding effect was detectable.
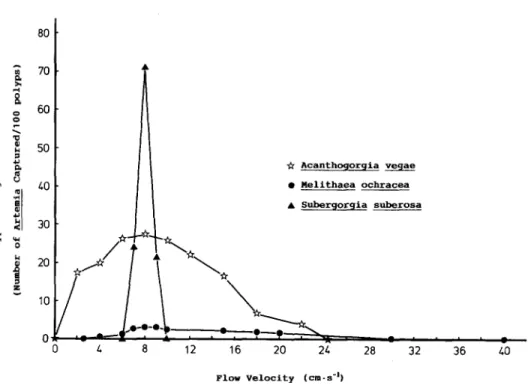
Feeding rates at different flow velocities showed that the highest rate of the three species appeared at =8cm.s.’ (Fig. 1). At 8cm.s~‘. feeding rates of the three spe- cies is ranked A. vegae > S. subemw > N. ochracecl. However, both the normalized col- ony feeding effectiveness (Fig. 2) and the polyp feeding effectiveness (Fig. 3) at 8 cm’s ’ are ranked S. suberosu> A. vegae > M. ochracea.
Acanthcqorgia wxa nelithaea ocRracea ___~ Subergorgia suberosa --~
FLOW AND FEEDING OF THREE GORGONIANS 65 Q Acanthogorgia vegae . Melithaea ochracea A Subergorgia suberosa 0 L a 12 16 20 21 28 32 36 10 Plow Velocity (cm.s-9
Fig. 3. Normalized polyp feeding effectiveness (number of Artemia captured per 100 polyps) for three gor- gonians (Subergorgia suberosa; Acanthogorgia vegae; Melithaea ochracea) in a range of flow velocities during
the first hour of exposure to Artemiu nauplii.
DISCUSSION
Feeding rates of the three species in different flow velocities show a similar trend (Fig. 1). Feeding rates increased initially with flow velocity, reached a peak value at
z8 cm.s -I, then decreased with increasing flow velocities. A similar phenomenon was noted in a sea pen (Best, 1988) and two gorgonians (Sponaugle & LaBarbera, 1991). Sponaugle (1991) suggested that the enhanced feeding success at intermediate flow velocities is typical of some suspension feeders. Feeding rates of a suspension feeder at different flow velocities are determined by the combination of three factors: (1) cap- ture efficiency, (2) rate of encounter with particles, and (3) deformation of the feeding elements (Leonard et al., 1988). Since gorgonians are passive suspension feeders, they must contend with potentially damaging hydrodynamic drag forces while filtering the water for food (Lewis, 1982; Patterson, 1984). In slow currents, volume of flow in- creases with flow velocity and the encounter rate increases (Best, 1988). Up to a point, the high encounter rate compensates for declines in capture efficiency as the feeding elements deform. Beyond this point, food capturing function becomes more impaired and feeding rate decreases. At very high flow velocity, organisms quit feeding and take evasive action by retracting polyps.
hb C.-D. DA1 AND M.-C. LIN
Each of the three gorgonians expanded their polyps and fed in a particular range of flow velocities. The lower velocity limit is likely constrained by feeding success, while the upper velocity limit may be constrained by either feeding success or the probabil- ity of detachment (Wainwright & Koehl, 1976; Harvell & LaBarbera. 1984). The range of flow velocities in which a gorgonian coral expands its polyps can also be viewed as a balance between the energy gained from feeding and the cost of keeping the polyps expanded. When water movement is too low, it may be more costly for the colony to remain expanded and capturing few zooplankton than to contract and capture none (Sebens, 1984). When currents are too strong to allow efficient feeding and the energy cost of remaining expanded is so high, then colonies may retract their polyps. Thus, at both low and high flow velocities, colonies expanded their polyps initially but then retracted them after a few hours of exposure to food items. The energy cost for a coral colony to remain expanded is related to polyp morphology. A tall polyp not only re- ceives greater drag but also is more easily deformed under strong currents (Koehl,
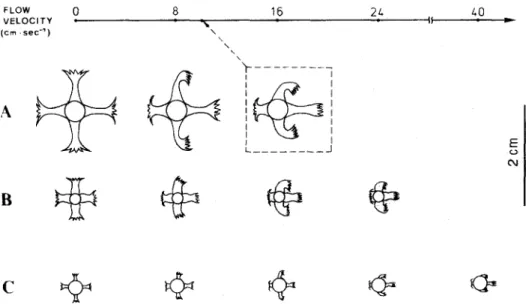
1977). Best (1988) demonstrated that the deformability of organisms strongly affects feeding efficiency and volume of water filtered. Although the branches of the three gorgonians have not been observed to bend in any current (Muzik & Wainwright, 1977; this study), deformation of polyps in high flow veiocities was evident (Fig. 4). Among the three species studied, S. suharosa has the tallest polyps and is more easily deformed at high flow velocities. At 8 cm’s -‘, polyp deformation of S. .suher-ostr was noticeable and the polyps could still feed in downstream eddies. Downstream eddies may result in localized concentrations of prey and the reduced currents in eddies may allow more
FLOW 0 8 16 21 VELOCITY l A 1’ (Cm .sec-‘) \ \
I---:
IGa
I___-____JFig. 4. Deformation of polyps of three gorgonians (Subergorgiu suberosu: Actmrhogorgiu vegue; Melithneu ochruceu) at different ROW velocities. Pictures are redrawn to scale from microscopic photographs.
FLOWANDFEEDINGOFTHREEGORGONIANS 67
efficient feeding (Leversee, 1976). But at 10 cmes-‘, polyps are so severely deformed that they can no longer feed effectively. Because the energy cost of keeping the polyps expanded is so high, the coral contracts its polyps after several hours of exposure to such flow velocity. On the other hand, polyps of M. ochrucea are the shortest and the least easily deformed at high flow velocities. In addition, polyps of M. ochrucea are arranged semi-laterally on the surface of the branches and expanded directly into the downstream zone of reduced currents (Muzik & Wainwright, 1977). These features enable M. ochracea to feed in a wide range of flow velocities. Although S. suberosa feed in a much narrower range of flow velocities, its feeding effectiveness per unit surface area and per polyp are the highest in the optimal flow velocity (Figs 2 and 3).
The feeding abilities of the three gorgonian species reflect the differences in their morphologies and habitats. The currents in the coastal areas of southern Taiwan are mostly dominated by semidiurnal tides. In Nanwan Bay, where the gorgonians were collected, the tidal currents flow westward during flood tide and eastward during ebb tide (Dai, 1991). Current velocities range from 2 to 50 cm’s_’ with an average of
15 cm.s -l. Current velocity is stronger in shallow water at depths between 5 and 15 m, especially in exposed areas (Liang et al., 1978). M. ochracea, which can feed in a wide range of flow velocities, is widely distributed on the reefs of southern Taiwan (Chen & Chang, 1991). Colonies of M. ochracea often grow on the upper part of reef front where currents are relatively strong. This distribution pattern is consistent with its
ability to feed in relatively high flow velocities. S. suberosu, which feeds in a relatively narrow range of flow velocities, has restricted distribution on the reefs of southern Taiwan. If often exists on the lower part of reef slopes or on sheltered boulders scat- tered over sandy bottom at depths between 15 and 25 m. A. vegue, which feed in relatively strong currents, usually grow on semi-exposed reef fronts or the lateral side of boulders .
This study reveals that different gorgonacean corals may feed most successfully in different ranges of flow velocities. Such flow-related feeding capabilities of suspension feeders are likely related to the deformation of their feeding elements in different flow velocities. The feeding performances of passive suspension feeders in different ranges of flow velocities may have significant influences on their distribution in the environ- ment, particularly where flow regimes are more complex such as on coral reefs.
ACKNOWLEDGEMENTS
We wish to express our appreciation to Drs. K.Y. Wei, T.-S. Chiu and C.-P. Chen for their comments on the manuscript. We also thank Mr. Y.-C. Tsay and T.-Y. Fan for their assistance in specimen collection. This study was supported by a grant from the National Science Council, R.O.C. (NSC 79-0418-B-002A-15).
6X C.-D. DA1 AND M.-C. I.lN
Bayer, F. H., 198 1. Key lo genera of Octocorallia exclusive of Pennatulacea (~oehlerala: ‘Anthozoa), wllh diagnosis of new taxa. Prr~c. Viol. Sot. Wush.. Vol. 94, pp. 902-947.
Best. B.A., 1988. Passive suspension feeding in a sea pen: efTects of ambient Ilou- on ~olumc How rate and filtering efficiency. Biol. Bull.. Vol. 175, pp. 332-342.
Chen, C. C. & K. H. Chang, 1991. Gorgonacea (Coelentcrata: .4nthoroa: Octocorallia) of southern Taiwan. Lull. host. Zool. Acad. Sinica. Vol. 30, pp. 149-182.
Dai, C. F., 1991. Reef environment and coral fauna of southern Taiwan. Aroll Rec. Bull.. \ol. 354. pp. i-
Grigg, R., 1972. Orientation and growth form of sea fans. Linrrrol. Oceunogr., Vol. 1’1, pp. 8% 192. Harvell. D. & M. LaBarbera, 1985. Flexibility: a mechanism for control of local velocity in hydroid cc&~-
nits. Biol. BUN., Vol. 168. pp. 3 12-320.
Hunter. T., 1989. Suspension feeding in oscillating tlow: the eff‘ec~ 01‘ colon) morphology and flow regime on plankton capture by the hydroid Oh& bngi.wiww. Biol. Bull.. Vol. 176, pp. 41-59.
Jargenscn, C.B., 1955. Quantitative aspects of filter feeding in invcrtebratcs. Bkjl. Rm, Vol. 30, pp. 391- -154.
Kochl, M.A. R., 1977. Effects of sea anemones on the ilo\\ forces they cncoun~c~-. ./. E.\/I. BI<J/.. Vol. 69. pp. x7-105.
Lasker, H. R., 1981. A comparison ofthe particulate feeding abilities of three specrcs of gorgonian soft COI-al. ~Lfcir. Ecol. Prog. Ser.. Vol. 5, pp. 61-67.
Lasker, H.R., M.D. Goltfried & M.A. Coffroth. 1983. Effects of depth on the feeding capabilities of two nctocorals. Mur. Biol., Vol. 73, pp. 73-78.
Leonard. A. B., J. R. Strickler & N. D. Holland, 1988. Effects of current speed on tiltrauon during suspcn- sion feeding in OIigonterra serripinnrr (Echinodermata: Crinoidea). Mm. BioJ. Vol. 97, pp. 1 I I- 125. Lcversee, G. J.. 1976. Flow and feeding in fan-shaped colonies of the gorgonian coral, Leptogorgia Biol. BUN..
Vol. 151, pp. 344-356.
Lcwlb. J. B., 1982. Feeding behaviour and feeding ecology of the Octocorallia (Cuclenterata: Anthozoa). J. %r,o/.. Land., Vol. 196, pp. 371-384.
Lang. N.K.. S.L. Lien, W.C. Chen and H.T. Chang, 1978. Oc~~rnq~q~hic ~rr~~etrificrtk~~~ ,!I the vicimr~ o/ .~~f~crn.tun-Nai~~ctn BuJ,. Spec. publ. No. IX. Instilute of Oceanography. Nalionai Taiwan Universit?. Taipei, Taiwan, 207 pp.
Murik, K. & S. Wainwright, 1977. Morphology and habitat of tive Fijian sea fans. HUN. blur. Sci.. Vol. 37. pp. 308-337.
Okamura, B., 1990. Particle size, flow velocity, and suspension-feeding by the erect brqozoans Hlcgulukr rwririm and B. stobnif~ra. Mm. Biol., Vol. 105, pp. 33-38.
Patterson, M. R., 1984. Patterns of whole colony prey capture in the octocoral. ~tic:1~o/1itrr,1 .kkrium. Biol. Bull.. Vol. 167, pp. 613-629.
Pauerson. M. R., 1991. The effcc~s of flow on polyp-lcvcl prc) capt urc in an ~,ctocoral. .,JI~.\~YI~III~~~ viderirml. B/r,/. Bull.. Vol. 180, pp. 93- 102
Ricdl, R.. 197 1. Water movemem. In. Muriw ewhgy, edited bq 0. Kinnc, !& ilel-Intcrsciencc. London. pp. 1 123- 1 150.
Scbcns. K. P.. 1984. Water How and coral colony siLe: mtcrhabllal comparisons 01 the octocoral .4k:)~oi1c~r/!r .siderium. Pm. Natl. Acad. Sci. USA, Vol. 81, pp. 5413-5477.
Sponaugle, S.. 1991. Flow patterns and velocities around a suspension-feeding gorgonian polyp: evidence frum physical models. J. Exp. MNY. Biol. Em/., Vol. 148. pp. 135-145.
Sponaugle, S. & M. LaBarbera, 1991. Drag-induced deformation: a functional feeding stratcgq in two species of gorgonians. .I. E.x~. Mar. Biol. Ecol.. Vol. 148, pp. 121-134.
\Vainwright, S. A. & J. R. Dillon. 1969. On the oricnlation of SCB fan5 (Genus (;rq~~~!itr). Hk~l. BlrN.. Vol. 136, pp. 130-139.
FLOW AND FEEDING OF THREE GORGONIANS 69
Wainwright, S. A. & M. A. R. Koehl, 1976. The nature of flow and the reaction of benthic cnidaria to it. In, Coelenterate ecology and behavior, edited by G. 0. Mackie, Plenum Press, New York, pp. 5-21. Wainwright, S. A., W. D. Biggs, J. D. Currey & J. M. Gosline, 1976. Mechanical design in organisms. Princeton
University Press, Princeton, New Jersey, USA, 423 pp.
Zou, R. L. & P. J. B. Scott, 1980. The Gorgonacea of Hong Kong. In, The marineffora undfauna