Urban & Fischer Verlag
http://www.urbanfischer.de/journals/jpp
Paraquat toxicity is reduced by nitric oxide in rice leaves
Kuo Tung Hung, Chin Jung Chang, Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
Received July 2, 2001 · Accepted October 1, 2001
Summary
The role of nitric oxide (NO) in plants has been the object of intensive research. In the present work, we evaluated the protective effect of NO against paraquat (PQ) toxicity of rice (Oryza sativa) leaves. PQ toxicity in rice leaves was determined by the decrease of protein content. PQ toxicity of rice leaves was reduced by free radical scavengers such as thiourea, sodium benzoate, ascorbic acid and sodium azide. NO-releasing compounds [N-tert-butyl-α-phenylnitrone (PBN), sodium nitroprus-side (SNP), 3-morpholinosydnonimine (SIN-1), and ascorbic acid+NaNO2] were effective in
reduc-ing PQ toxicity in rice leaves. SIN-1 and ascorbic acid+ NaNO2 prevented both the PQ-induced
increase in content of malondialdehyde and the PQ-induced decrease in activities of active oxygen species-detoxifying enzymes. The protective effect of SIN-1 or ascorbic acid+NaNO2on PQ toxicity,
PQ-induced lipid peroxidation and PQ-decreased antioxidative enzyme activities was reversed by 2-(4-carboxy-2-phenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide, a NO-specific scavenger, sug-gesting that the protective effect of NO-releasing compounds is attributed to NO released. Reduction of PQ toxicity by NO in rice leaves is most likely mediated through increase in antioxidative enzyme activities and decrease in lipid peroxidation.
Key words:lipid peroxidation – nitric oxide – Oryza sativa – paraquat
Abbreviations:Asc ascorbate. – APOD ascorbate peroxidase. – CAT catalase. – c-PTIO 2-(4-car-boxy-2-phenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide. – GR glutathione reductase. – MDA malondialdehyde. – NO nitric oxide. – PBN N-tert-butyl-α-phenylnitrone. – POD peroxidase. – PQ paraquat. – SIN-1 3-morpholinosydonimine. – SNP sodium nitroprusside. – SA sodium azide. – SB sodium benzoate. – SOD superoxide dismutase
Introduction
Paraquat (PQ), also known as methyl viologen, is a herbicide widely used in agriculture. It has long been known to exert its toxic effects by catalyzing the transfer of electrons from
pho-* E-mail corresponding author: [email protected]
tosystem I of chloroplast membrane to molecular oxygen, producing oxygen radicals that cause lipid peroxidation and membrane damage (Calderbank 1968, Farrington et al. 1973, Cha et al. 1982). A normally operated photosynthetic electron transport is usually required for inducing PQ toxicity (Mees 1960, Chang and Kao 1995). Superoxide can serve as a source to generate more active hydroxyl radicals by
Weiss and Fenton reactions (Naqui and Chance 1986). Tran-sition metals, such as iron and copper, are able to accelerate Haber-Weiss and Fenton reactions (Gutteridge et al. 1981). It has been demonstrated that iron or copper ions are essential mediators in PQ toxicity in bacterial cells (Kohen and Chevion 1985), in mammalian cells in tissue culture (Sandy et al. 1987) and mice (Kohen and Chevion 1986). It has been shown that PQ toxicity in pea and rice leaves is enhanced by iron and reduced by iron chelators (Zer et al. 1994, Chang and Kao 1997 a). Free radical scavenging properties of polyamines have been documented (Drolet et al. 1986). It was found that polyamines were effective in reducing PQ toxicity of rice and sunflower leaves (Chang and Kao 1997b, Benavides et al. 2000).
Nitric oxide (NO) is a labile free radical and is produced from L-arginine by NO synthase in mammalian cells (Sakuma et al. 1988, Rees et al. 1989). Over the last decade, extensive research about the role of NO in animals has demonstrated that it is a key signal molecule involved in a wide variety of processes including vasorelaxation, neurotransmission, and the innate immune response (Schmidt and Walter 1994). In contrast, the role of NO in plants is less well understood. Evi-dence is emerging to support that NO may also play some roles in the plant kingdom. Antibodies raised against rabbit NO synthase cross-reacted with proteins from pea embryonic axes and wheat germ (Sen and Cheema 1995). In senescing pea leaves, NO emission was promoted by the addition of ethylene precursors and NO-releasing compounds inhibited leaf expansion (Leshem and Haramaty 1996). Leshem et al. (1998) provided evidence to show that NO is a natural senes-cence-delaying plant growth regulating agent. NO-releasing compounds were found to promote phytoalexin accumulation in potato tuber tissues (Noritake et al. 1996), inhibit coniferyl alcohol peroxidase activity in the xylem of Zinnia elegans (Ferrer and Bacelo 1999), inhibit cytochrome oxidase activity in isolated soybean cotyledon mitochondria (Millar and Day 1996), and inhibit the activity of tobacco aconitase, catalase (CAT), ascorbate peroxidase (APOD) and salicylic acid-induced protein kinase (Clark et al. 2000, Kumar and Klessig 2000, Navarre et al. 2000). The participation of NO as a signal in plant disease resistance has also been reported (Delle-donne et al. 1998, Durner et al. 1998, Klessig et al. 2000). Scherer and Holk (2000) also observed that NO could be an intermediate in cytokinin signaling.
Since NO is a toxic free radical, most of the work on NO function in plant cells has focused on its ability to act in the same direction as active oxygen species. In animal systems, several reports convincingly demonstrate a protective role for NO in oxidative stress (Rubbo et al. 1984, Wink et al. 1993, Rauhala et al. 1996, Goss et al. 1997, O’Donnell et al. 1997, d’Ischia et al. 2000). In a series of work conducted by Lamat-tina’s group, it was found that NO protects against cellular damage produced by diquat, PQ and Phytophthora infestans in potato leaves (Laxalt et al. 1997, Beligni and Lamattina 1999 a, b). It is not known whether a similar protective role for
NO is also active in other leaf systems. For this reason, we in-vestigated the effect of NO on the toxicity of PQ, a well known free-radical generating chemical, in rice leaves.
Materials and Methods
Materials and chemicals
Rice (Oryza sativa L., cv. Taichung Native 1) was cultured as pre-viously described (Lin et al. 1999). The apical 3-cm segments excised from the third leaves of 12-day-old seedlings were used. A group of 10 segments was floated in a Petri dish containing 10 mL of test solution. Incubation was carried out at 27˚C in the light (40µmol m– 2s–1).
Test solutions included PQ, NO-releasing compounds, a NO scav-enger (c-PTIO), and antioxidant compounds. PBN, SIN-1 and SNP were used as NO-releasing compounds. We also used a solution con-taining ascorbic acid (Asc) and NaNO2as another NO-releasing
com-pound. Asc, sodium azide (SA), sodium benzoate (SB), and thiourea were used as antioxidants. All chemicals were purchased from Sigma Co. (St. Louis, MO, USA).
Chlorophyll, protein and MDA determinations
Chlorophyll was determined according to Wintermans and De Mots (1965) after extraction in 96 % (v/v) ethanol. For protein extraction, leaf segments were homogenized in 50 mmol L–1sodium phosphate buffer
(pH 6.8). The extracts were centrifuged at 17,000×g for 20 min, and the supernatants were used for determination of both protein by the method of Bradford (1976) and enzyme activities. Malondialdehyde (MDA) was extracted with 5 % (w/v) trichloroacetic acid and deter-mined according to Heath and Packer (1968).
Enzyme assays
Peroxidase (POD) activity was measured using a modification of the procedure of MacAdam et al. (1992). Activity was calculated using the extinction coefficient [26.6 (mmol L–1)–1cm–1at 470 nm] for
tetraguaia-col. CAT activity was assayed by measuring the initial rate of disap-pearance of H2O2 (Kato and Shimizu 1987). The decrease in H2O2
was followed as the decline in absorbance at 240 nm, and activity was calculated using the extinction coefficient [40 (mmol L–1)–1cm–1 at
240 nm] for H2O2 (Kato and Shimizu 1987). Superoxide dismutase
(SOD) was determined according to Paoletti et al. (1986). APOD was determined according to Nakano and Asada (1981). The decrease in Asc concentration was followed as the decline in optical density at 290 nm and activity was calculated using the extinction coefficient [2.8 (mmol L–1)–1cm–1at 290 nm] for Asc. Glutathione reductase (GR)
was determined by the method of Foster and Hess (1980). One unit of activity for CAT, POD, SOD, APOD, and GR was defined as the amount of enzyme which degraded 1µmol H2O2per min, caused the
formation of 1µmol tetraguaiacol per min, inhibited 50 % the rate of NADH oxidation observed in control, degraded 1µmol of Asc per min, and decreased 1 A340per min, respectively.
Experimental design
Protein and MDA contents were expressed per g fresh weight (FW). Enzyme activities were expressed as units g–1FW or units mg–1
pro-tein. Absolute contents of each measurement varied among experi-ments because of seasonal effects. However, the patterns of re-sponses to PQ, PQ plus NO-releasing compounds or PQ plus antioxi-dants were reproducible. For biochemical analysis, four independent extractions were performed for each treatment. All experiments de-scribed here were repeated at least three times. Similar results and identical trends were obtained each time. The data reported here are from a single experiment.
Results and Discussion
Changes in chlorophyll, protein, and MDA contents in de-tached rice leaves treated with 25
µmol L
–1PQ in the light are shown in Figure 1. The decrease in protein and increase inFigure 1.Changes in contents of chlorophyll, protein and MDA in rice leaves treated with PQ. Detached rice leaves were treated with either water or 25µmol L–1PQ in the light. Vertical bars represent standard
errors (n=4). Only those standard errors larger than the symbol are shown.
Figure 2.Effect of antioxidants (Asc, SB, SA, and thiourea) on protein content in rice leaves treated with PQ. The concentration of PQ was 25µmol L–1. Protein was determined 24 h after treatment in the light.
The white columns mean that detached rice leaves were treated with water only. Vertical bars represent standard errors (n=4).
MDA were evident at 4 h after PQ treatment. However, slight decrease in chlorophyll was only observed at 24 h after PQ treatment. Clearly, PQ is more effective in decreasing protein content than chlorophyll content. Thus, PQ toxicity in de-tached rice leaves was judged by the decrease in protein content in the following experiments. It is generally believed that the primary action of PQ is a result of the deviation of photosynthetic electron transport in chloroplasts (Calderbank 1968). Previously, we demonstrated that considerable reduc-tion of PQ toxicity in rice leaves occurred in the presence of photosynthetic electron transport inhibitors in the light and that PQ was unable to induce toxicity in detached rice leaves under dark condition (Chang and Kao 1995). These results suggest that light is essential for PQ response in rice leaves, although the light intensity (40
µmol m
– 2s–1) used in thepres-ent investigation is quite low compared to sunlight. It appears that detached rice leaves are highly sensitive to PQ.
PQ treatment resulted in a marked increase in MDA con-tent, indicating that PQ brings about lipid peroxidation (Fig. 1). Lipid peroxidation is caused by active oxygen species (Foster and Hess 1980). Free radical scavenging properties of polyamines have been documented (Drolet et al. 1986) and it was found that polyamines were effective in reducing PQ toxicity of rice leaves (Chang and Kao 1997b). Transition
met-als, such as iron and copper, are able to accelerate Haber-Weiss and Fenton reactions (Gutteridge et al. 1981). Evidence was also provided to show that PQ toxicity was reduced by iron chelators in rice leaves (Chang and Kao 1997 a). These results support the possibility that PQ-induced toxicity in rice leaves is mediated through oxidative stress. In the present in-vestigation, when free radical scavengers such as thiourea, SB, SA and Asc were used together with PQ, it was found that they partially prevented the toxicity of PQ in rice leaves (Fig. 2). However, Asc, SB, SA, and thiourea alone had no effect on protein content (data not shown). These results support the involvement of free radicals as the chemical species inducing PQ toxicity in rice leaves.
Beligni and Lamattina (1999 a, b) reported that NO counter-acts cellular damage caused by diquat in potato leaves. Based on their evidence, they propose that NO acts as an antioxidant in reducing cellular damage. Thus, it is of great in-terest to know whether a similar protective role of NO is also active in the rice leaf system. Consequently, detached rice leaves were treated with PQ in the presence or absence of NO-releasing compounds, such as PBN, SIN-1, SNP and a mixture Asc and NaNO2for 24 h in the light. As indicated in
Table 1, all NO-releasing compounds used are effective in re-ducing PQ toxicity in rice leaves. We also observed that 100
µmol L
–1 SIN-1 and 100µmol L
–1 Asc plus 200µmol L
–1 NaNO2 are more effective in reducing PQ toxicity than PBNand SNP.
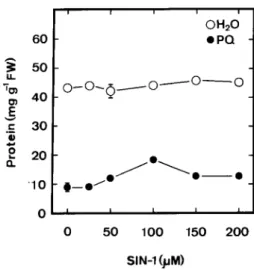
SIN-1 alone had no effect on protein content (Fig. 3). When applied together with PQ, SIN-1 concentrations between 50 and 200
µmol L
–1produced a clear protection against protein loss (Fig. 3). The optimal concentration of SIN-1 in reducing PQ toxicity was observed to be 100µmol L
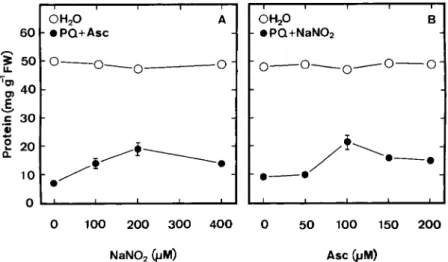
–1. It was also ob-served that NaNO2alone (Fig. 4 A) or Asc alone (Fig. 4 B) hadno effect on protein content in detached rice leaves. Both Asc and NaNO2are required in reducing PQ toxicity of detached
rice leaves (Figs. 4 A and 4 B). To be effective in reducing PQ toxicity, the optimal concentrations of Asc and NaNO2were
Table 1.Effect of NO-releasing compounds on protein content in rice leaves treated with PQ
Treatment Protein, mg g-1FW H2O 53.6±1.0 PQ 8.7±0.6 PQ+PBN 17.1±1.5 PQ+SIN-1 20.2±1.7 PQ+SNP 16.1±1.3 PQ+Asc+NaNO2 22.9±1.9
The concentrations of PQ, PBN, SIN-1, SNP, Asc, and NaNO2were 25,
100, 100, 100, 100, and 200µmol L–1, respectively. Protein content was
determined 24 h after treatment in the light, data are the means± standard errors (n=4).
Figure 3.Effect of SIN-1 concentrations on protein contents in rice leaves treated with either water or PQ. The concentration of PQ was 25µmol L–1. Protein was determined 24 h after treatment in the light.
Vertical bars represent standard errors (n=4). Only those standard errors larger than the symbol are shown.
100 and 200µmol L–1, respectively (Figs. 4 A and 4 B). The
pro-tective effect of SIN-1 or Asc+NaNO2on PQ toxicity was
ob-served to be evident at 4 h after treatment (data not shown). To investigate whether the protective effect induced by treatment with NO-releasing compounds (SIN-1 and Asc + NaNO2) was the result of the production of NO, 100
µmol L
–1c-PTIO, a NO-specific scavenger, was applied along with 100
µmol L
–1SIN-1 or 100µmol L
–1Asc+200µmol L
–1NaNO2.
The protective effect of SIN-1 or Asc+NaNO2on PQ-induced
toxicity could be reversed by c-PTIO (Fig. 5). Clearly, the pro-tective effect on PQ toxicity by NO-releasing compounds is due to NO.
The addition of NO to rat brain homogenates caused a de-crease in lipid peroxidation (d’Ischia et al. 2000). PQ induces lipid peroxidation in several leaf systems (Calderbank 1968, Farrington et al. 1973, Cha et al. 1982, Chang and Kao 1997a). Until now it was not known whether NO-releasing compounds can confer protection against lipid peroxidation in plant sys-tems. Figure 6 shows the effect of SIN-1 and Asc+NaNO2on
MDA content of detached rice leaves treated with PQ. PQ alone resulted in a marked increase in MDA content (Figs. 1 and 6). Although SIN-1 or Asc+NaNO2alone had no effect
on MDA content (data not shown), treatment with SIN-1 or Asc +NaNO2caused a reduction of PQ-induced increase in MDA
content (Fig. 6). The effect of SIN-1 or Asc+NaNO2on
PQ-induced MDA content could also be reversed by c-PTIO (Fig. 6).
The protective effect of NO-releasing compounds on PQ toxicity is clearly associated with the decrease in lipid peroxi-dation (Fig. 6) and protein loss (Fig. 5). Plant cells are equipped with several free radical detoxifying enzymes to protect them against oxidative damage. These enzymes
in-Figure 4. (A) Effect of NaNO2 concentrations on
protein content of rice leaves treated with water or 25µmol L–1PQ+100µmol L–1Asc; (B) Effect of
Asc concentrations on protein content in rice leaves treated with water or 25µmol L–1 PQ +
200µmol L–1NaNO
2. Protein was determined 24 h
after treatment in the light. Vertical bars represent standard errors (n=4). Only those standard errors larger than the symbol are shown.
Figure 5. Effect of SIN-1 and Asc+ NaNO2 on PQ toxicity in rice
leaves in the presence (+) or absence ( – ) of c-PTIO. The concentra-tions of SIN-1, Asc, NaNO2, PQ, and c-PTIO were 100, 100, 200, 25,
and 100µmol L–1, respectively. Protein was determined 24 h after
treatment in the light. The white columns mean that detached rice leaves were treated with water only. Vertical bars represent standard errors (n=4).
clude SOD, APOD, GR, CAT and POD (Foyer et al. 1997, Na-kano and Asada 1981). The striking changes in lipid peroxida-tion seen in leaves treated with PQ alone may be a reflecperoxida-tion of the inability of the scavenging enzymes to cope with the in-creased levels of oxidant species and/or their degradation by these species. Treatment with PQ alone decreases POD, CAT, SOD, APOD and GR activities (Fig. 6). The PQ-induced loss of antioxidative enzyme activities was found to be protected by SIN-1 or Asc+NaNO2(Fig. 6). Treatment with c-PTIO alone was
found to have no effect on MDA content and antioxidative en-zyme activities (data not shown). However, c-PTIO arrested NO-releasing compounds-mediated enzyme protection (Fig. 6). It should be noted that when antioxidative enzymes were expressed as specific activities (units mg–1 protein), PQ
in-creased specific activities of antioxidative enzymes, NO-releasing compounds counteracted PQ-increased specific ac-tivities of antioxidative enzymes and c-PTIO reversed the effect of NO-releasing compounds-mediated enzyme protection (Ta-ble 2).
Ferrer and Barcelo (1999) found that the NO donor SNP (5 mmol L–1) and NO (55
µmol L
–1s–1) itself were able to inhibitTable 2.Effect of SIN-1 and Asc+NaNO2on the specific activities of antioxidative enzymes in PQ-treated rice leaves in the presence or absence
of c-PTIO.
Treatment Units mg– 1protein H
2O PQ PQ+SIN-1 PQ+Asc PQ+SIN-1 PQ+Asc+ +NaNO2 +c-PTIO NaNO2+c-PTIO
CAT 0.11±0.01 0.23±0.02 0.16±0.03 0.18±0.01 0.24±0.01 0.24±0.02 POD 0.68±0.02 2.25±0.17 1.71±0.16 1.80±0.22 3.44±0.22 3.60±0.44 SOD 1.01±0.03 4.03±0.16 2.11±0.05 1.28±0.09 4.51±0.19 4.35±0.15 APOD 0.77±0.01 1.56±0.08 1.21±0.16 1.17±0.07 1.95±0.15 2.19±0.23 GR 0.18±0.02 0.60±0.05 0.44±0.07 0.42±0.03 0.94±0.09 1.03±0.15 The concentrations of PQ, SIN-1, Asc, and NaNO2were 25, 100, 100, and 200µmol L– 1, respectively. Enzyme specific activities were
Figure 6.Effect of SIN-1 and Asc+NaNO2
on the content of MDA and the activities of antioxidative enzymes in PQ-treated rice leaves in the presence (+) or absence ( – ) of c-PTIO. The concentrations of SIN-1, Asc, NaNO2, PQ, and c-PTIO were 100,
100, 200, 25, and 100µmol L–1,
respec-tively. MDA content and enzyme activities were determined 24 h after treatment in the light. The white columns mean that de-tached rice leaves were treated with water only. Vertical bars represent standard er-rors (n=4).
POD activity in the xylem of Zinnia elegans. Clark et al. (2000) also demonstrated that NO donor (0.8 mmol L–1) inhibited the
activities of tobacco CAT and APOD, heme-containing en-zymes. However, our results show that neither SIN-1 nor Asc+ NaNO2 treatment effected the activities of POD, CAT, and
APOD in rice leaves (data not shown). Why is SIN-1 or Asc+ NaNO2able to counteract PQ toxicity, but not able to inhibit
the activities of POD, CAT, and APOD in rice leaves? In the present investigation, we used
µmol L
–1concentration range of SIN-1 and Asc+NaNO2and did not measure theconcen-tration of NO released by SIN-1 or Asc + NaNO2 in rice
leaves. We do not know the exact concentration of NO inside the rice leaf tissue. It is possible that the concentration of NO released by SIN-1 or Asc+ NaNO2is high enough to
scav-enge free radicals produced by PQ, but not sufficient to in-hibit the activities of heme-containing enzymes.
In rice leaves, involvement of active oxygen species in wa-ter stress-, methyl jasmonate-, excess Cu-, and excess
Cd-promoted senescence has been demonstrated (Hung and Kao 1998, Chen and Kao 1999, Lin and Kao 2000, Chien et al. 2001). Work is under way to determine whether NO counter-acts this promotion of senescence in rice leaves. Preliminary results show that the promotion of rice leaf senescence caused by polyethylene glycol, which induces lipid peroxida-tion, can be counteracted by NO-releasing compounds, but that caused by sorbitol, which has no effect on lipid peroxida-tion, cannot be counteracted by NO-releasing compounds (unpublished data).
In conclusion, the results presented in this paper suggest that reduction of PQ toxicity by NO in rice leaves is most likely mediated through increase in antioxidative enzyme activities and decrease in lipid peroxidation.
Acknowledgements. This work was supported by grant NSC 90-2313-B-002-267 from the National Science Council of the Republic of China.
References
Beligni MV, Lamattina L (1999 a) Nitric oxide counteracts cytotoxic processes mediated by reactive oxygen species in plant tissues. Planta 208: 337– 344
Beligni MV, Lamattina L (1999 b) Nitric oxide protects against cellular damage produced by methyl viologen herbicides in potato plants. Nitric Oxide 3: 199 – 208
Benavides MP, Gallego SM, Comba ME, Tomaro ML (2000) Relation-ship between polyamines and paraquat toxicity in sunflower leaf discs. Plant Growth Regul 31: 215 – 224
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248 – 254
Calderbank A (1968) The bipyridylium herbicides. Adv Pest Cont Res 8: 127– 235
Cha LS, McRae DG, Thompson JE (1982) Light-dependence of para-quat-initiated membrane deterioration in bean plants. Evidence for the involvement of superoxide. Physiol Plant 56: 492 – 499 Chang CJ, Kao CH (1995) Paraquat-induced chlorophyll loss in rice
leaves: Evidence for the involvement of light and photosynthetic electron transport. Chinese Agron J 5: 287– 293
Chang CJ, Kao CH (1997a) Paraquat toxicity is reduced by metal che-lators in rice leaves. Physiol Plant 101: 471– 476
Chang CJ, Kao CH (1997b) Paraquat toxicity is reduced by polyamines in rice leaves. Plant Growth Regul 22: 163 –168
Chen L-M, Kao CH (1999) Effect of excess copper on rice leaves: evi-dence for involvement of lipid peroxidation. Bot Bull Acad Sin 40: 283 – 287
Chien H-F, Wang J-W, Lin CC, Kao CH (2001) Cadmium toxicity of rice leaves is mediated through lipid peroxidation. Plant Growth Regul (in press)
Clark D, Durner J, Navarre DA, Klessig DF (2000) Nitric oxide inhibi-tion of tobacco catalase and ascorbate peroxidase. Mol Plant – Microbe Interact 13: 1380 –1384
d’Ischia M, Palumbo A, Buzzo F (2000) Interactions of nitric oxide with lipid peroxidation products under aerobic conditions: Inhibitory ef-fects on the formation of malondialdehyde and related thiobarbitu-ric acid-reactive substances. Nitthiobarbitu-ric Oxide 4: 4 –14
Delledonne M, Xia Y, Dixon RA, Lamb C (1998) Nitric oxide functions as a signal in plant disease resistance. Nature 394: 585 – 588 Drolet G, Dumbroff EB, Legge RL, Thompson JE (1986) Radical
scav-enging properties of polyamines. Phytochemistry 25: 367– 371 Durner J, Wendehenne D, Klessig DF (1998) Defense gene induction
in tobacco by nitric oxide, cyclic GMP, and cyclic ADP-ribose. Proc Natl Acad Sci USA 95: 10328 –10333
Farrington JA, Ebert M, Land EJ, Fletcher K (1973) Bipyridylium qua-ternary salts and related compounds. V. Pulse radiolysis studies of the reaction of paraquat radical with oxygen, implication for the mode of action of bipyridyl herbicides. Biochim Biophys Acta 314: 372 – 381
Ferrer MA, Bacelo AR (1999) Differential effects of nitric oxide on per-oxidase and H2O2production by the xylem of Zinnia elegans. Plant
Cell Environ 22: 891– 897
Foster JG, Hess JL (1980) Responses of superoxide dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol 66: 482 – 487 Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen
per-oxide- and glutathione-associated mechanism of acclimatory stress tolerance and signaling. Physiol Plant 100: 241– 254
Goss SPA, Hogg N, Kalyanaraman B (1997) The effect of nitric oxide release rates on the oxidation of human low density lipoprotein. J Biol Chem 272: 21647– 21653
Gutteridge JMC, Rowley DA, Halliwell B (1981) Superoxide-dependent formation of hydroxyl radicals in the presence of iron salts. Bio-chem J 199: 263 – 265
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Bio-chem Biophys 125: 189 –198
Hung KT, Kao CH (1998) Involvement of lipid peroxidation in methyl jasmonate-promoted senescence in detached rice leaves. Plant Growth Regul 24: 17– 21
Kato M, Shimizu S (1987) Chlorophyll metabolism in higher plants. VII. Chlorophyll degradation in higher plants. VII. Chlorophyll degrada-tion in senescing tobacco leaves: Phenolic-dependent peroxi-dative degradation. Can J Bot 65: 729 –735
Klessig DF, Durner J, Noad R, Navarre DA, Wendehenne D, Kumar D, Zhou JM, Shah J, Zhand S, Kachroo P, Trifa Y, Pontier D, Lam E, Silva H (2000) Nitric oxide and salicylic acid signaling in plant de-fense. Proc Natl Acad Sci USA 97: 8849 – 8855
Kohen R, Chevion M (1985) Transition metals potentiate paraquat tox-icity. Free Radic Res Commun 1: 79 – 88
Kohen R, Chevion M (1986) Paraquat toxicity is enhanced by iron and reduced by desferrioxamine in laboratory mice. Biochem Pharma-col 34: 1841–1843
Kumar D, Klessig DF (2000) Differential induction of tobacco MAP ki-nases by the defense signals nitric oxide, salicylic acid, ethylene, and jasmonic acid. Mol Plant – Microbe Interact 13: 347– 351 Laxalt AM, Beligni MV, Lamattina L (1997) Nitric oxide preserves the
level of chlorophyll in potato leaves infected by Phytophthora
infes-tans. Eur J Plant Pathol 103: 643 – 651
Leshem YY, Haramaty E (1996) Plant aging: the emission of NO and ethylene and effect of NO-releasing compounds on growth of pea (Pisum sativum) foliage. J Plant Physiol 148: 258 – 263
Leshem YY, Wills RBH, Ku VV-V (1998) Evidence for the function of the free radical gas – nitric oxide (NO) – as an endogenous maturation and senescence regulating factor in higher plants. Plant Physiol Biochem 36: 825 – 833
Lin J-N, Kao CH (2000) Involvement of lipid peroxidation in water stress-promoted senescence of detached rice leaves. Biol Plant 43: 141–145
Lin J-N, Wang J-W, Kao CH (1999) Effect of abscisic acid and water stress on the senescence of detached rice leaves. Biol Plant 42: 313 – 316
MacAdam JW, Nelson CJ, Sharp RE (1992) Peroxidase activity in the leaf elongation zone of tall fescue. Plant Physiol 99: 872 – 878 Mees GC (1960) Experiment on herbicidal action of
1,1-ethylene-1,2-dipyridylium dibromide. Ann Appl Biol 48: 601– 612
Millar AH, Day DA (1996) Nitric oxide inhibits the cytochrome oxidase but not the alternative oxidase of plant mitochondria. FEBS Lett 398: 155 –158
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascor-bate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22: 867– 880
Naqui A, Chance B (1986) Reactive oxygen intermediates in biochem-istry. Annu Rev Biochem 55: 127–166
Navarre DA, Wendehenne D, Durner J, Noad R, Klessig DF (2000) Nitric oxide modulates the activity of tobacco aconitase. Plant Physiol 122: 573 – 582
Noritake T, Kawakita K, Doke N (1996) Nitric oxide induces phytoalexin accumulation in potato tuber tissues. Plant Cell Physiol 37: 113 –116 O’Donnell VB, Chumley PH, Hogg N, Bloodsworth A, Darley-Usmar
VM, Freeman BA (1997) Nitric oxide inhibition of lipid peroxidation: kinetics of reaction with lipid peroxyl radicals and comparison with
α-tocopherol. Biochemistry 36: 15216 –15223
Paoletti F, Aldinucci D, Mocali A, Capparini A (1986) A sensitive spec-trophotometric method for the determination of superoxide dismu-tase activity in tissue extracts. Anal Biochem 154: 536 – 541 Rauhala P, Sziraki I, Chiueh CC (1996) Peroxidation of brain lipids in
vitro: Nitric oxide versus hydroxyl radicals. Free Radic Biol Med 21:
391– 394
Rees DD, Palmer RM, Hodson HF, Moncada S (1989) A specific inhib-itor of nitric oxide formation from L-arginine attenuates endothe-lium-dependent relaxation. Br J Pharmacol 96: 418 – 424
Rubbo H, Radi R, Trujillo M, Telleri R, Kalyanaraman B, Barnes S, Kirk M, Freeman BA (1984) Nitric oxide regulation of superoxide and peroxinitrite-dependent lipid peroxidation. J Biol Chem 269: 26066 – 26075
Sakuma I, Stuehr DJ, Gross SS, Nathan C, Levi R (1988) Identification of arginine as a precursor of endothelium-derived relaxing factor. Proc Natl Acad Sci USA 85: 8664 – 8667
Sandy MS, Moldeus P, Ross D, Smith MT (1987) Cytotoxicity of the re-dox cycling compound diquat in isolated hepatocytes: Involvement of hydrogen peroxide and transition metals. Arch Biochem Biophys 259: 29 – 37
Scherer GFE, Holk A (2000) NO donors mimic and NO inhibitors in-hibit cytokinin action in betalaine accumulation in Amoranthus
can-datus. Plant Growth Regul 32: 345 – 350
Schmidt HHWH, Walter U (1994) NO at work. Cell 78: 919 – 925 Sen S, Cheema IR (1995) Nitric oxide synthase and calmodulin
im-munoreactivity in plant embryonic tissue. Biochem Arch 11: 221– 227
Wink DA, Hanbauer I, Krishna MC, DeGraff W, Gamson J, Mitchell JB (1993) Nitric oxide protects against cellular damage and cyto-toxicity from reactive oxygen species. Proc Natl Acad Sci USA 90: 9813 – 9817
Wintermans JFGM, De Mots A (1965) Spectrophotometric characteris-tics of chlorophylls a and b and their pheiphytins in ethanol. Bio-chim Biophys Acta 109: 448 – 453
Zer H, Peleg I, Chevion M (1994) The protective effect of desferriox-amine on paraquat-treated pea (Pisum sativum). Physiol Plant 92: 437– 442