行政院國家科學委員會專題研究計畫 期中進度報告
脯胺酸異構 pinl 的生物功能及調控機制之探討(2/3)
計畫類別: 個別型計畫
計畫編號: NSC91-2320-B-110-006-
執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日
執行單位: 國立中山大學生物醫學研究所
計畫主持人: 呂佩融
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 92 年 12 月 16 日
一、 中文摘要
本報告為計劃 NSC91-2320-B-110-006 期中報
告,計劃名稱:脯胺酸異構酶Pin1 的生物功能及
調控機制之探討(2);Pin1 是一個具特異性的 phosphorylation-dependent PPIase,可專一性地辨認 phosphorylated Ser/The-Pro motif,並是第一個發現
細胞生長所需的PPIase。在人類及酵母菌細胞中, Pin1 同時具有負面調控細胞進入 mitosis 以及正面 調控離開mitosis 的過程。之前研究指出,在 in vitro 的情況之下,Pin1 會結合及調控一系列 mitotic phosphoproteins ,之後被證明為 MPM2 antigens 本研究主要的目的是為了了解Pin1 功能與結構之 關係,以及Pin1 本身磷酸化及去磷酸化而被調控 的分子機制。在此三年的計畫中,我們有三個主 要的目的:(1)完成 Pin1 structure- function 細節分 析 (2)了解在 in vitro 情況下,Pin1 如何調控其 target protein 的機制 (3)了解 Pin1 在 cell cycle 中的 localization 及 phosphorylation。在第一年研究 (NSC90-2320-B-110-010),我們已利用 site direct mutagenesis 的方法來產生 Pin1 的 mutants,並且
研究各個Pin1 mutants 對於 PPIase 活性及 MPM2
antigen binding 的影響。證明了 WW domain 是和 phosphoSerine -Proline motif binding 有關而 PPIase domain 則是和 prolyl isomerase activity 有關。
第二年(NSC91-2320-B-110-006) 研究我們針
對Pin1 WW domain 之磷酸化機制及其對 Pin1 功
能之調控進行研究,我們證明了Ser16 之 in vitro
及in vivo 的磷酸化位置並猜測此磷酸化可能與細
胞週期調控及PKA- signaling pathway 有關。磷酸
化Pin1 WW domain 也被證明可影響 Pin1 與其受
質之交互作用及Pin1 在微細胞位置,更有趣的是
Pin1 Ser16 位置磷酸化的 WW domain 可以造成 mitotic 的停滯及細胞凋亡(apoptosis) 且增加多核 細胞(multinucleated cells)的數目。由此我們相信, Pin1 磷酸化為調控 WW domain phosphoSerine - binding activity 及細胞生理活性的重要機制,以上 結果已發表於J. Biol. Chem。此外,我們與哈佛大 學合作,Pin1 在小鼠之生理角色的探討亦在第二 及第三年計畫中其間進行。 關鍵字:脯胺酸異構酶,細胞週期,磷酸化,磷 酸激酶 Abstract
Pin1, a unique phosphorylation-dependent PPIase that specifically recognizes phosphorylated Ser/The-Pro motifs, has been demonstrated as the first PPIase that is essential for cell growth. Pin1 both negatively regulates entry into mitosis and positively regulates progression through mitosis in human and yeast cells (1). Moreover, it has been shown that Pin1 binds and regulates a subset of mitotic phosphor-proteins, MPM2 antigens in vitro (1). In addition, our preliminary results showed that Pin1 itself likely is phosphorylated in a cell cycle- regulated manner. A major goal of this grant proposal is to elucidate the molecular mechanism by which Pin1 regulates cell cycle progression and how itself was regulated by phosphorylation and dephosphory- lation. Three specific aims are within this three year proposal: (a) to perform a detailed structure-function analysis on Pin1, (b) to elucidate the mechanism by which Pin1 regulates its target proteins (Pints) in vitro, (c) to examine the localization and phosphory- lation of Pin1 during the cell cycle.
In the second grant year, we have demonstrated that the Pin1 WW domain is phosphorylated on Ser16 both in vitro and in vivo. Furthermore, this phosphorylation regulates the ability of the WW domain to mediate Pin1 substrate interaction and cellular localization. Both Pin1 and WW domain mutants refractory to Ser16 phosphorylation induced mitotic block and apoptosis, and increased multi- nucleated cells with 8N DNA content. Thus, phosphorylation is a new mechanism critical for regulating the WW domain phosphoserine-binding activity and Pin1 function in vitro and vivo.
On the other hand, we have continued the Pin1 structure–function analysis and determined the ability of Pin1 mutants obtained from year 1 to
rescue a temperature-sensitive yeast strain YPM2 in vivo. Taken together with the results of PPIase and localization activities, we have characterized both WW and PPIase domain mutations and provide detailed structure-function analysis of Pin1.
Keywords: prolyl isomerase Pin1, cell cycle, phosphorylation, kinase, apoptosis
二、Introduction
Phsophoserine-binding modules help determine the specificity of signal transduction events. Indeed, Proline- directed protein kinases, such as cyclin- dependent kinases (Cdks), play important roles in regulating eukaryotic cell division. At the G2-to-M phase transition, a rapid activation of Cdc2, the mitotic Cdk, results in phosphorylation of many proteins on serine or threonine residues that are followed by praline (Ser/Thr-Pro sites). These phosphorylation events are thought to trigger many of the structural modifications that occur during mitosis. However, it is not clear what effects the phosphorylation of mitotic proteins actually and how these abrupt changes in phosphorylation state lead to an organized and programmed set of the mitotic events. Pin1, a unique phosphorylation-dependent PPIase that specifically recognizes phosphorylated Ser/ The-Pro motifs (2). It has been demonstrated that Pin1 is essential for mitotic progression in human and yeast cells (3), and is required for the DNA replication checkpoint in Xenopus extracts (4). Pin1 substrates are a defined subset of phosphorylated proteins, including many MPM-2 antigens (1,3,5). Pin1-catalyzed prolyl isomerization regulates the conformation and function of phosphoproteins (3-6) and also facilitates dephosphorylation because of the conformational specificity of some phosphatases (7). Thus, Pin1-dependent peptide bond isomerization is a critical post- phosphorylation regulatory mechanism, allowing cells to turn phosphoprotein function on or
off with high efficiency and specificity (1-6). The consistent level of Pin1 during the cell cycle suggested that Pin1 is likely regulated by a post-translational modification. Given the current interest in aberrant cell cycle regulation in carcinogenesis and the ability to kill cells specifically at mitosis by inhibiting Pin1 function, the results obtained in these studies will be important for understanding the detailed molecular mechanism of the cell cycle control, should shed light on mechanisms of carcinogenesis, and might be eventually used to develop new therapeutic reagents for cancer. In the second grant year, we have aimed to determine the ability of Pin1 mutants to rescue a temperature-sensitive yeast mutant, YPM2, to identify the Pin1 phosphorylation sites, and to examine how the phosphorylation effect on Pin1 function both in vivo and in vitro.
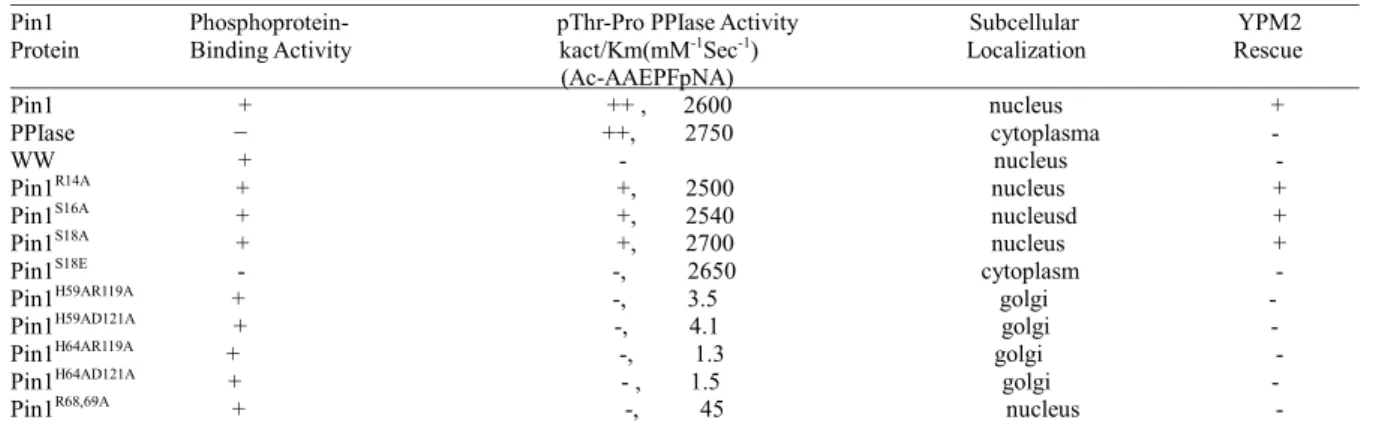
We reported here that single mutation of S16E, Y23A or W34A in pSerPro binding pocket of WW domain reduced the rescue ability of Pin1 in YPM2 yeast strain (Table1 and Figure1). The similar effect can be also observed when H59A, H64A, or R68A single mutation was made in catalytic site of PPIase domain (Table1 and Figure1). Taken together, both WW and PPIase domains are essential for Pin1 function in vivo. In addition, our results have showed that Pin1 was phosphorylated on Ser16 by PKA. The WW domain functions as a pSer/ Thr Pro binding module, with the binding pocket including the side chains of S16, R17, Y23 and W34 (3,4). Interestingly, Ser16 is located at the center of the shadow pSer/ThrPro - binding pocket. We have demonstrated that S16 phosphorylation can regulate the ability of the WW domain to mediate Pin1 substrate interaction and cellular localization that plays an essential role for the WW domain pSer/Thr-binding activity and Pin1 cell cycle function (5,8).
Previously, we have demonstrated Pin1, a unique phosphorylation-dependent PPIase that specifically recognizes phosphorylated Ser/The-Pro
motifs (2) which suggested that Pin1 could provide a new post-translational level of control to allow the
Table 1. Functional Properties of Pin1 Mutants
Pin1 Phosphoprotein- pThr-Pro PPIase Activity Subcellular YPM2 Protein Binding Activity kact/Km(mM-1Sec-1) Localization Rescue
(Ac-AAEPFpNA) Pin1 + ++ , 2600 nucleus +
PPIase − ++, 2750 cytoplasma - WW + - nucleus -
Pin1R14A + +, 2500 nucleus +
Pin1S16A + +, 2540 nucleusd +
Pin1S18A + +, 2700 nucleus +
Pin1S18E - -, 2650 cytoplasm -
Pin1H59AR119A + -, 3.5 golgi -
Pin1H59AD121A + -, 4.1 golgi -
Pin1H64AR119A + -, 1.3 golgi -
Pin1H64AD121A + - , 1.5 golgi -
Pin1R68,69A + -, 45 nucleus -
The phosphoprotein-binding activity was assayed by their ability to bind MPM-2 antigens.+,normal binding. PPIase activity was assayed by their ablity to isomerizes peptide substrates. +, PPIase activity; -, PPIase inactive.
Figure 1
Fig1 (A) genetic munipulation of Pin1 in YPM2
(B) Pin1 protein expression profile in YPM2 .
(C) YPM2 rescue experiments by Pin1 mutants general increase in protein phosphorylation to result in the organized and programmed set of mitotic events. One of my specific aims is to perform a detailed structure-function analysis on Pin1. Previous results demonstrate that Pin1 WW domain binds phosphoproteins and then displays a phosphorylation -dependent PPIase activity by PPIase domain. We here studied whether pSer-binding activity of WW domain or PPIase activity or both are essential for
Pin1 function in vivo. We used one Pin1 deletion yeast strain YPM2 to examine both the pSer-binding dead and PPIase dead Pin1 mutants. We found that the disruption of either pSer-binding activity of WW domain or PPIase activity can abolish the Pin1 ability to sescueYPM2 in vivo (Table1 and Figure1). Taken together, we concluded that both pSer-binding activity of WW domain and PPIase activity are essential for Pin1 function in vivo.
In addition, compelling evidence indicates that phosphory -lation on Ser or Thr residues (pSer/Thr) also promotes the formation of protein-protein (1-3). One well characterized family of such proteins utilizing phosphorylation-dependent interactions are 14-3-3 proteins, which function as adapter proteins. More recently, small pSer/Thr-binding protein modules reminiscent of SH2 domains involved in recognition of phosphorylated Tyr have been described, which include group IV WW domains and FHA domains (1-3). Although the function of these pSer/ Thr-binding proteins or modules depends on phosphorylation of their ligands, the role of phosphorylation of these modules themselves has not been described.
A subset of WW domains is among the first pSer/Thr-binding modules to be characterized (1-4).
WW domains are found in a variety of proteins, and many have been shown to mediate protein-protein interactions at least partly through Pro-rich motifs (1). A subset of WW domains function as pSer/Thr- binding modules (2). The best characterized pSer/ Thr-binding WW domain is the peptidyl- prolyl isomerase (PPIase) Pin1. The first and essential step in Pin1-dependent regulation of targets is the substrate binding activity mediated by its WW domain (1-3). The WW domain functions as a pSer/ Thr-Pro-binding module, with the binding pocket including the side chains of S16, R17, Y23 and W34 (1,3,6). Interestingly, Ser16 is located at the center of the shadow pSer/Thr-Pro-binding pocket.
Fig.2
A
Fig2.
(A) Kd values between Pin mutants and Pintide
(B) MPM2 pull down by Pin1isabolished by PKA phosphorylation
Here we have demonstrated that an essential role for phosphorylation of Ser16 in regulation of the WW domain pSer/Thr-binding activity and subsequent Pin1 cell cycle function. The evidence include the following experimental results (a) Phosphorylation on Ser16 is both necessary and sufficient to regulate the pin1 substrate interaction in
vitro (Fig2). (b) Cell cycle regulated phosphorylation of Pin1 on ser16in cells (8). (c) Phosphorylation on Ser16regulates subcellular localization of Pin1(8). (d) WW domain S16 A but not S16E mutant affects the cell cycle in vivo (8). The role of phosphorylation of Pin1 WW domain described above can be divided into two parts, one has been published in J. Biol.
Chem. (2002) 277, Jan 25, 2381-2384 and one other
was in submission.
四、計畫成果自評
本研究報告包含二部分,第一部分為探討 Pin1 的結構及功能分析,包括測量突變點對於 Pin1 的PPIase 測量及對 phosphoprotein 的結合能力及 對之 in vivo 活性分析,此與三年計畫之第二年specific Aim 相符合;第二部分為探討 Pin1 被磷酸
化之機制及對Pin1 功能之調控,其中第二部分發
表於J.B.C (2002)277, Jan 25, 2381-2384.具有學術
價值及校稿中,第一部分待整理更多數據後,將 發表於國際生化期刊。
五、References
(1) Zhou, X. Z., Lu, P.J., Wulf, G., and Lu, K.P.(1996)
Nature 380, 544-547
(2) Lu, P. J., Zhou, X. Z., Shen, M., and Lu, K. P. (1999)
Science 283, 1325-1328
(3) Lu, K.P., Hanes, S. D., and Hunter, T. (1996) Nature 380, 544-547
(4) Verdecia, M. A., Bowman, M. E.,Lu, K. P., Hunter, T., nd Noel, J. P. (2000) Nat Struct Biol 7, 639-643 (5) Shen, M., Stukenberg, P. T., Kirschner, M. W., and Lu,
K. P(1998) Genes Dev. 12, 706-720
(6) Yaffe, M. B., Schutkowski, M., Shen, M., Zhou, X. Z., Stukenberg, P. T., Rahfeld, J., Xu, ., Kuang, J., Kirschner, M. W., Fischer, G., Cantley, L. C., and Lu, K. P.(1997) Science 278, 1957 – 1960
(7) Zhou, X. Z., Kops, O., Werner, A., Lu, P. J., Shen, M., , Fischer, G.,and Lu, K.P.(2000) Mol Cell 6,873-883 (8) P. J. Lu., X. Z. Zhou., Y. C.Liou., J. P. Noel., K. P.