Taipei Medical University Institutional Repository:Item 987654321/4250
86
0
0
全文
(2) 誌謝 兩年的歲月,如涓涓流水,緩慢,但卻是無法停滯。每每案牘勞形,很累,卻 也無庸置疑地充實。只有在此刻提起寸管之時,才有那種— 如釋重負,終於熬 過來的感覺。多少回在寂靜的夜裡內省吾身,我到底學習到了什麼?多少次我問 我自己,這是我想要的嗎?在幾乎看不到光明的深淵裡,還好有貴人相助,才使 得我能撥雲見日,豁然開朗。 感謝我的指導教授 陳建和老師的提攜與指導,讓我這株小草有個安穩的環境 下成長茁壯。感謝 梁有志老師,在我這學海生涯中,給予我多次的指導與解惑, 給予我正確的邏輯觀念之誘導與建立。感謝我的口試委員 黃嘯谷教授、吳和生 主任、許明照老師所給予的指教與建議,也特別感謝 劉俊仁老師悉心的指導我 撰寫這篇英文論文與救火般地協助,讓本篇論文能更加地完整及正確。 感謝所有的老師在這些日子以來的激勵與幫助,讓學生能開闊視野、不只侷限於 一角。 特別感謝我一些博學多聞的朋友們,遠在英國牛津大學唸博班的 志豪,幫我 找一些我無法找到的paper之外,也總是會給我一些現在國外最新的研究資訊, 並且也會很有耐心的聆聽我的問題並針對問題點來點醒我,雖然他無法即時的在 我身邊伸出援手,但都還是很努力的扮演好安慰我的角色,因為有你的鼓舞,就 算我們遠在地球的兩端,我們依然朝著自己的夢想努力、打拼。還有在中研院唸 博班及國衛院同事過的學長學姐們 彥淞、世婷,幫忙找我無法下載的paper給我 及實驗設計上的引導與啓發,並且也很感謝他們聽我訴說不悅,因為有你們的關 懷,在怎麼辛苦都能咬牙地撐下去。還有感謝 顯裕給予我實驗上技術的支持及 觀念的建立。感謝 照君的指導與傾聽…………。因為有你們的存在,讓我知道 我不是孤獨的。 另外感謝國衛院老師們及榮總的醫師老板們,謝謝他們的栽培 及愛護,讓我得以有勇氣地往前一步去考取博班。謝謝實驗室的伙伴們 柯小白、 QOO、琇丹、藍絜,有你們的陪伴,讓我在研究室不只是侷限於實驗之外,還 II.
(3) 得到許多的酸甜苦辣點滴在心頭,讓我在實驗室的生活得以多姿多采。感謝可愛 的學弟妹們-氶弘、千翔、詩芸在我廢寢忘食的做實驗之時,他們能夠幫我買晚 餐,讓我得以溫飽之外還溫暖了我的心,讓我體會到天上最美的是星星,人生最 美的是溫情。 並且感謝梁家的學姐們給予實驗上的協助,及其學弟妹同學的相助,有了你們 的談笑風生,兩方的實驗室多了那麼點的歡樂氣氛。還有三樓實驗室的一些同 學、學弟妹們 徹堃、鋒哥、凱玄,我不會忘記有你們的陪伴及謝謝你們傾聽我 所講的笑話。 最後我要感謝我的父母及姐妹們,謝謝你們的支持,讓我無後顧之憂的打拼我 自己的學業,一圓求學的夢想。有你們,真好。就是要這麼一輩子的在一起。. 奕璇 謹致於 台北醫學大學 醫學科學研究所 醫學檢驗暨生物技術系 民國九十七年七月二十一日 III.
(4) IV.
(5) 摘要 退化性關節炎為好發於年紀大的老年人,其特徵是因軟骨發炎導致軟骨的 結構破壞,進而瓦解軟骨產生關節疼痛感。近來,對於退化性關節炎的治療包括 服用非類固醇抗發炎藥物或服用葡萄糖胺來減緩疼痛,此外亦有使用外科關節置 換手術。然而有些病患對於藥物反應不佳,且又礙於年紀而無法做關節置換手 術,因此發展新的治療方式頗為重要。近年來研究指出,注射玻尿酸至關節腔內, 不僅可潤滑關節腔及包覆軟骨,並且能修復軟骨細胞外基質成份如 aggrecan、 typeⅡ collagen 等保護作用。先前文獻指出高分子量玻尿酸具有降低由 IL-1β誘 發所引起的 TNF-α、MMPs 及 iNOS 等,與退化性關節炎相關的發炎物質基因 表現。而在本研究中加入高分子量玻尿酸至軟骨肉瘤細胞的結果發現,高分子量 玻尿酸可抑制環氧化酶-2 (COX-2)及血紅素氧化酶 1 (HO-1)的表現,證實了 HO-1 在不同組織的特異性。除此之外,高分子量玻尿酸也會提升軟骨肉瘤細胞核接受 器 peroxisome proliferator-activated receptor (PPARγ)的表現。由於 PPARγ的增 加,表示此藥物有著抗發炎的作用,亦即證實高分子量玻尿酸具抗發炎的作用。 深入探討發現,高分子量玻尿酸是經由抑制 JNK 磷酸化或經由提升 Akt 的磷酸 化而抑制了 NF-κB 的訊息傳遞路徑來抑制這些發炎物質的產生。本研究中,我 們也同時探討低分子量玻尿酸的作用,結果發現低分子量玻尿酸的作用卻是增加 環氧化酶-2 ( COX-2 )及血紅素氧化酶 1 (HO-1 )的表現,並且抑制 PPARγ的表現 量。從分子機制證實,低分子量玻尿酸是藉由提升 p38 及抑制 Akt 的磷酸化,以 提升 NF-κB 的訊息傳遞路徑,進而增加發炎性物質的產生。. V.
(6) Abstract Osteoarthritis (OA), which is a primarily inflammatory and degenerative joint disease characterized by progressive loss of articular cartilage, is the most common form of arthritis occurred in the elder people. Loss of cartilage cushion causes friction between the bones, will lead to pain of joint. Current treatments of OA are taking Non-steroidal anti-inflammatory drugs (NSAIDs), Glucosamine and chondrotin sulfate, or Total Knee Arthroplastry (TKA). Because of poor response to medicine and too older for TKA, a new treatment for OA is important. Previous report showed that injection of Hyaluronic acid in knee joint was helpful for OA, which could provide a backbone for the attachment of aggrecan side chains by link proteins in chondrocyte. Present study aims to investigate the effects of HMW-HA on the gene expression of OA- associated cytokines and enzymes, including IL-1β, TNF-α MMP-1, MMP-13, and inducible NOS (iNOS) in primary chondrocyte from OA patients. In this study, the expression of COX-2 and HO-1 were down regulated by HMW-HA treatment in chondrosarcoma cell (SW1353). Besides, up-regulation of PPARγ could also be found in SW1353 after HMW-HA treatment. It indicated that treatment of HMW-HA had anti-inflammatory effect. Further examination showed that the regulation of MMPs、 COX-2、HO-1 and PPARγ expression were through the Akt phosphorylation and NF-κB inhibition. In contrast to HMW-HA, LMW-HA would. induced inflammatory. mediators, including activated MMPs, COX-2 and HO-1 expression as well as down regulated PPARγ expression. These results were proved lately by in vitro studies that through activation of NF-κB and inhibition of Akt phosphorylation. From all the results obtained from this study, the HMW- and LMW-HA have opposite mechanism on the chondrosarcoma cell, SW1353.. VI.
(7) 關鍵詞: 間質金屬蛋白酶-1( MMP-1 ) 間質金屬蛋白-13( MMP-13 ) 軟骨肉瘤 ( Chondrosarcoma ) 高分子量玻尿酸 ( HMW-HA ) 低分子量玻尿酸 ( LMW-HA ) 退化性關節炎 ( Osteoarthritis, OA ) 血基質氧化酶-1 ( Heme oxygenase-1 ) 環氧化酵素( COX-2 )、 過氧化體增殖劑活化受器 γ ( PPARγ). Key words : Matrix metalloproteinase 1 (MMP-1) Matrix metalloproteinase 13 (MMP-13) Chondrosarcoma cell, SW1353 High molecular weight Hyaluronaic acid ( HMW-HA) Low molecular weight Hyaluronaic acid ( LMW-HA) Osteoarthritis ( OA ) Heme oxygenase-1 ( HO-1) Cyclooxygenase-2 ( COX-2) Peroxisome proliferator-activated receptor ( PPARγ). VII.
(8) Contents. Page. 1. Introduction 1.1 What is Osteoarthritis (OA)………………………………………....…….…1 1.2 ECM and MMPs……………………………………………….......……...…1 1.3 OA and COX-2……………………………………….…….………………..3 1.4 OA and HO-1…………………………………………………………….…..3 1.5 MMPs and signaling pathways………………………………….………..….4 1.6 Hyaluronan-CD44 and signaling pathways………………………………….5 1.7 OA and PPARγ……………………………………………………….…….6 1.8 Cell cycle and apoptosis………………………………………………....…..7. 2. Materials and Methods 2.1. Reagents………………………………………………………..……....…..…9 2.2 Culture of chondrosarcoma and preparation of cell lysates………………......9 2.3 WST-1 assay……………………………………………………………...…..10 2.4 RT-PCR…………………………………………………………….……..…..10 2.5 SDS PAGE and Western blotting………………………………….……...….12 2.6 Flow Cytometry (FACS)…………………………………………..………....12 2.7 Statistics analysis…………………………………………………...…….….13 3. Results 3.1 Dose effect of HMW-HA and LMW-HA on SW1353 cell viability………...14 3.2 Dose dependent effect of HMW-HA on IL-1β induced MMP-1 and MMP -13 expression…………………………..……………………………………...…..14 3.3 Dose dependent effect of LMW-HA on IL-1β induced MMP-1 and MMP-13 expression……………………………………….…………………...…….…..15 3.4 Dose dependent effect of HA on COX-2 xpressio………………………...…15 VIII.
(9) 3.5 Treatment of SW1353 with dose dependent manner of HMW-HA increase the level of HO-1 gene and protein expression………………….....…….……......16 3.6 Effects of HA on the expression of Nuclear receptor, PPAR.………………..16 3.7Effect. of. HMW-HA on. the. Akt. phosphorylation. and. I κ B α. expression…..………………………………………………………….………17 3.8Effect of HMW-HA on phosphorylation of MAPKs induced by IL-1 β………………..…………………………………………….……………..…18 3.9 Effect of HMW-HA on the protein expression of cyclin D1 and dk4…….….19 3.10 Effect of LMW-HA on the-Akt phosphorylation and IκBα expression………………………………………………………………….......19 3.11 Effect of LMW-HA on phosphorylation of MAPKs induced by IL-1β…………………………………………………………………..….……20. 4. Discussion………………………………………..…………………………….….21. 5. Reference………………………………………………………………………….67. IX.
(10) Figure Figure 1 Effect of HMW-HA and LMW-HA on SW1353 cell viability in chondrosarcoma cell, SW1353………………………………….…..…...29 Figure 2. Effects of HMW-HA on IL-1β induced MMPs gene expression in chondrosarcoma cell, SW1353…………………………………….….....30. Figure 3. Effects of LMW-HA on IL-1β induced MMPs gene expression in chondrosarcoma cell, SW1353…………………………………….….…32. Figure 4. Effects of HMW-HA on IL-1β induced MMPs protein prodution in chondrosarcoma cell,SW1353……………………………….………..…34. Figure 5. Effects of LMW-HA on IL-1β induced MMPs protein expression in chondrosarcoma cell, SW135………………………………….…..…….36. Figure 6. Effects of HMW-HA on IL-1β induces COX-2 gene and protein expression in chondrosarcoma cell, SW1353………………….…….…..38. Figure 7. Effects of LMW-HA on IL-1β induces COX-2 gene and protein expression in chondrosarcoma cell, SW1353………………….………...40. Figure 8. Effects of HMW-HA on IL-1β induces HO-1 gene and protein expression in chondrosarcoma cell, SW1353………………………...……………...42. Figure 9. Effects of LMW-HA on IL-1β induces HO-1 gene and. protein. expression in chondrosarcoma cell, SW1353……….…………….……..44 Figure 10 Effects of HMW-HA on IL-1β induces PPARγgene and protein expression in chondrosarcoma cell, SW1353………...……………….…46 Figure 11 Effects of LMW-HA on IL-1β induces PPARγgene and protein expression in chondrosarcoma cell, SW1353………….………….……..48 Figure 12 Effect of HMW-HA on IL-1β induced phosphorylation of Akt and IκBα expression in chondrosarcoma cell, SW1353………………….50 X.
(11) Figure13 Effect of HMW-HA on IL-1β induced phosphorylation of ERK, JNK, p38 in chondrosarcoma cell, SW1353………………………….…..…...52 Figure 14 Effect of HMW-HA on IL-1β induced cyclin D1 ans Cdk4 expression in chondrosarcoma cell, SW1353……………………………………...…..54 Figure 15 Effect of LMW-HA on IL-1β induced phosphorylation of Akt and IκBα expression in chondrosarcoma cell, SW1353………….……....56 Figure 16 Effect of LMW-HA on IL-1β induced phosphorylation of ERK, JNK, p38 in chondrosarcoma cell, SW1353……………………………….….58 Figure 17 Effect of LMW-HA on IL-1β induced cyclin D1 ans Cdk4 expression in chondrosarcoma cell, SW1353………………………...………………...61 Figure 18 HMW-HA signaling pathway in SW1353………………………………62 Figure 19 Proliferator-activated receptor γ (PPARγ) induction by HMW-HA signaling pathway in SW1353………………………………….……….63 Figure 20 LMW-HA signaling pathway in SW1353……………………………….64 APPENDIX 1………………………………………………………………………..65 LMW-HA induces cell apoptosis. XI.
(12) 1. Introduction. 1.1 What is Osteoarthritis (OA). Osteoarthritis (OA) is a gradually progressing disorder of mammalian joints, characterised by the destruction of articular cartilage, which results in discomfort and dysfunction of the affected joint [1, 2]. The pathologic changes during the development of OA are remarkably similar and include proteoglycan degradation at the early stage, followed by type II collagen degradation, leading eventually to localized or complete loss of cartilage matrix [3].. 1.2 ECM and MMPs. Degradation of extracellular matrix (ECM) is an important event in a variety of physiological processes especially in development, tissue remodeling and wound healing. ECM remodeling is also an essential component of cell proliferation, metastasis, and invasion of malignant tumors [1, 4]. The Matrix metalloproteinases (MMPs) play important roles in the degradation of ECM in various physiological and pathological conditions. MMPs constitute a family of enzymes with more than 20 members identified, which are extracellular endopeptidases requiring Zn2+ for their enzymatic activity that degrade the extracellular matrix [5]. MMPs are calssified to a protein family because of their sequence kinship and consequent structural and enzymatic similarities. MMPs activity is tightly coordinated at several levels: transcriptional regulation, activation of latent zymogen, and interaction with endogenous inhibitors [6]. Most MMPs are expressed at low level and can be induced or activated by numerous cytokines and growth factor stimuli. 1.
(13) MMPs play roles in pathological conditions involving untimely and accelerated turnover of ECM, including immune response, inflammation, angiogenesis, metastsis, proliferation and apoptosis [6-9]. Among the MMPs, attention has been focus on a 52kDa MMP-1(also called collagenaseⅠ) and 60kDa MMP-13 (Collagenase-Ⅲ). MMP-1 has been shown to be a prognostic factor in different malignancies such as nasopharyngeal carcinomas, knee cartilage degeneration and non-small cell lung cancer [10-12]. Human chondrocyte cell lines express a variety of MMPs, especially the MMP-1, the studies to assess the inflammatory of chondrocytes have demonstrated a strong correlation between chondrocytes proinflammatory and high level of MMP-1 expression [3, 13]. It has been reported that the levels of matrix-destructive enzymes such as MMP-1, MMP-3, MMP-13 increased and those of TIMP-1, a proteinase inhibitor, decreased in OA cartilage [14, 15] and reported the effect of estrogen on MMP-1, MMP-3, MMP-13 and TIMP in OA chondrocytes [16, 17]. In cartilage lesion of human OA for evidence of mRNA expression for three MMPs (1, 3, 9) that the level of expression has been examined in four histologically distinct stages [18, 19]. The regulation of MMP-1,3,13 is regarded as the most important component for mRNA expression [20-25]. MMP-1,3 and 13 are inducible genes that can be induced by IL-1 proinflammtory cytokine in normal articular human chondrocytes [20, 21]. Interleukin-1 (IL-1), tumor necrosis factor-α (TNF-α), and epidermal growth factor (EGF) can increase MMP-1 and MMP-13 transcription level in both chondrocytes and synovial cells. MMP-1 gene contains a novel promoter region, which can interact with inflammation responsive transcriptionn faction SAF-1 in chondrocytes [26, 27], These extracellular stimuli result in activation of SAF-1and nuclear AP-1 transcription factor complex, which then binds to MMP-1 promoter and activates transcription [28, 29]. Synovial fibroblasts are activated to produce MMPs, 2.
(14) the effects of microparticles were investigated, microparticles occur abundantly in the synovial fluid in rheumatoid arthritis (RA) and it is increased during activation and apoptosis [30].. 1.3 OA and COX-2. Pro-inflammatory cytokines such as IL-1β and TNF-α are produced by activated synoviocytes and articular chondrocytes and play a pivotal role in the pathogenesis of osteoarthritis (OA) [31]. IL-1β and TNF-α also activate other mediators of inflammation such as cyclo-oxygenase-2 (COX-2) which increase prostaglandin E2 (PGE2)-production, joint pain and further synovial inflammation. IL-1β and TNF-α also suppress the expression of cartilage-specific extracellular matrix components. Many of the biological effects of IL-1β and TNF-α in many cells (i.e. up-regulation of MMPs, COX-2 and inducible nitric oxide synthase) are also mediated by NF-κB. NF-κB appears to be a common downstream target of multiple converging catabolic signaling pathways (e.g. those mediated by IL-1β and TNF-α). In pervious papers, Curcumin, COX-2 inhibitor and HDAC inhibitor have already entered clinical trials because of its potent anti-inflammatory, anti-carcinogenic and free radical scavenger properties [32-34].. 1.4 OA and HO-1 Heme oxygenase (HO) activity catabolizes heme to biliverdin, which is reduced to bilirubin by biliverdin reductase, carbon monoxide, and iron. HO-1 is induced by a variety of stimuli or agents that cause oxidative stress, such as cytokines, reactive oxygen species, nitric oxide, heat shock, ultraviolet radiation, hypoxia, and hyperoxia 3.
(15) [35]. In pervious many papers found that heme oxygenase-1 (HO- 1) is down-regulated by pro-inflammatory cytokines and up-regulated by IL-10 in OA chondrocytes [36]. But, in the murine collagen-induced arthritis (CIA) model [37]. Treatment with SnPP significantly reduced the severity of CIA, with inhibition of joint inflammation and cartilage destruction. SnPP was more effective than CoPP in preventing the development of CIA (prophylactic administration). HO-1 overexpression cannot slow the progression of the chronic inflammatory disease, whereas treatment with SnPP, which inhibits HO-1, exerts prophylactic and therapeutic effects [38].. 1.5 MMPs and signaling pathways. Inhibition of MAPK and NF-κB pathway resulted in inhibition of MMP-1 expression suggesting these signaling pathways are involved in the regulation of MMP-1 expression [26].The MMPs inhibitors have undergone extensive development in the last decade and have proven effectiveness against inflammatory, invasion and angiogenesis of cell carcinoma both in vitro and in vivo [39-41]. Thus, in clinical setting, inhibition of MMPs can be applied in the treatment of OA or post-surgically as an adjuvant therapy to prevent recurrence [42]. This and the fact that HA also may be induce chondrocytes proliferation and collagen repair prompt us to further investigate the mechanism by which HA exert these effects. On the other hand, HA oligosaccharide inhibits chondrocytes proliferation and induces transcription of MMPs , collagen disruption and cell invasiveness [43, 44].. 4.
(16) 1.6 Hyaluronan-CD44 and signaling pathways. Hyaluronic acid (HA) is a high molecular weight naturally biopolymer, GlcNAcβ(1→4)GlcUAβ(1→3) of the ECM, which is a broadly used in anti-inflammation. Recently researches used HA in clinical applications as diverse as dermal scaffolds, cartilage defects, and ECM regeneration, such as wound healing [45]. CD44 is a primary receptor of HA, providing cells for sensing and attachment to HA and intracellular signaling pathway [46]. HA–CD44 signaling can activate several pathways, through activation of specific intermediates. HA binding to CD44 activated NF-κB signal transduction pathway through Ras and PKC. HA-CD44 interation was found to activate mitogen-activated protein kinase (MAPK) in tumor cells. Three mitogen-activated protein kinase (MAPK) pathways are known to regulate MMP-1, MMP-13 and other MMPs expression, p38 kinase, c-Jun N-terminal kinase (JNK) and extracellular signal-regulated kinase (ERK) [47-52]. Recently, ERK/MAP kinase was found to play an important role in caseine production and regulate the production of AP-1 as a result of extracellular stimuli by human fibroblasts and chondrocytes [53]. ERK and JNK have been implicated in the regulation of collagenase gene expression in cultured fibroblasts [54]. Hyaluronan has been shown to inhibit the activity of JNK, p38 and NF-κB in rheumatoid synovial fibroblasts (RSF) through ICAM . This show CD44 mediates HA effects only partially, and investigated the role of intercellular adhesion molecular (ICAM-1) is another cell surface receptor for HA [21]. Hyaluronan-CD44 promotes phospholipase C-mediated Ca2+ signaling and cisplatin resistance in head and neck cancer [25]. MCF-7 cell line treated with Hyaluronan increased ERK1/2 activation was stimulated by CD44-RHAMM receptor [55]. Hyaluronan-binding protease (HABP) also activates ERK1/2 and PI3K/Akt signaling pathways in fibroblasts and stimulates cell proliferation and migration [56]. 5.
(17) The first report of HA oligomer involvement in angiogenesis appeared in 1985, in which HA fragments limited to the 6–20 size range were shown to be angiogenic [57]. These HA fragments are not only mitogenic for endothelial cells, but also enhance endothelial cell migration and induce multiple signaling pathways [58]. Such HA fragments induce tyrosine kinase cascades [59]. The small oligosarccharides such as hexzmers to inhibit tumor growth can be attributed to PTEN, a phosphatase that degrades PIP3, inhibiting growth by inducing pro-apoptotic mediatros [60]. Hyaluronan oligosaccharides also activate a NF-κB /I-κBα auto-regulatory loop [61], inducing transcription of metalloproteases MMP-9 and MMP-13 [43]. The induction of nitric-oxide synthase by such saccharides also occurs through a nuclear NF-κB-dependent mechanism [62], and up-regulated c-Jun Nterminal kinase (JNK) and p38 mitogen-activated protein kinase expression. Soluble RHAMM, another HA receptor also induces G2/M arrest by suppressing the expression of Cdc2/Cyclin B1[63].. 1.7 OA and PPARγ. Peroxisome proliferator-activated receptors (PPARs) are a family of ligand-activated transcription factors belonging to the nuclear receptor superfamily. So far, three PPAR subtypes have been identified: PPARα, PPARβ/δ, and PPARγ. PPARα is present mostly in the liver, heart, and muscle, where it is the target of the fibrate class of drugs and is believed to function in the catabolism of fatty acid [64]. IL-1β treatment of OA chondrocytes downregulated PPARγ1 expression in a dose- and time-dependent manner. PPARγ1 is downregulated in OA cartilage. The pro-inflammatory cytokine IL-1β may be responsible for this downregulation via a mechanism involving activation of the MAPKs (p38 and JNK) and NF-κB signaling 6.
(18) pathways. The IL-1β- induced downregulation of PPARγ expression might be a new and additional important process by which IL-1β promotes articular inflammation and cartilage degradation [65].. 1.8 Cell cycle and apoptosis. MAPK cascades induce specific cell cycle responses to extracellular stimuli. Sustained ERK activity is a potent promoter of G1 to S phase progression and increases transcription of c-Fos, which results in increased levels of AP-1 and cell proliferation [66]. HMW-HA inhibit the mitogen-activated proliferation of a variety of cells, including mesenchymal cells, it can inhibits the mitodependent induction of cyclin D1 and down-regulation of p27kip1 and skp2 gene expression and block G1 to S phase entry in vascular SMCs [67]. on the other hand, Hyaluronan Oligosaccharides also can inhibit tumorigenicity of osteosarcoma cell line [68]. HMW-HA activates cell motility via PI3K/Akt and Ras kinase induction in tumor cell, it also up-regulates ERK1/2 and PI3k/Akt signaling pathways in fibroblast and stimulates cell proliferation, migration and anti-apoptotic effect [56, 69]. But our data reveal HMW-HA can inhibit cell proliferation through up-regulate PI3K/Akt signal, and inhibit apoptosis in chondrosarcoma cell. In the present papers, we propose to further investigate that hyaluronan can be a potential therapeutic agent for osteoarthritis (OA). We have been investigate that treatment of human chondrocytes and synovial fibroblasts with HA inhibits MMP-1, MMP-13 activity and ECM repair. Our study also investigate the mechanism by which HA decreases MMP-1, MMP-13 activity in human chondrocytes and human synovial fibroblasts. We also examine whether NF-kB reduction by HA plays a role in regulation of MMP-1, MMP-13 activity. In addition, we will determine whether CDK 7.
(19) inhibitors play a role in mediating HA-induced cell cycle arrest to inhibit MMPs production in human chondrocytes and human synovial fibroblasts.. 8.
(20) 2. Materials and Methods 2.1. Reagent. Leibovitz’s L-15 medium (L-15), fetal bovine serum (FBS), L-glutamine, penicillin/streptomycin, and amphotericin were purchased from Life Technologies (Invitrogen Corp, American). Antibodies specific for MMP-1, MMP-13, pAkt , pJNK , CD44, IκBα, apoptosis antibodies and GAPDH were purchased from cell signaling technology. Horseradish peroxidase-conjugated anti-rabbit (mouse) IgG antibody was purchased from Santa Cruz biotechnology, Inc (Santa Cruz, CA). Recombiant Human IL-1β was purchased from R&D System, Inc. Hyaluronan sodium was purchased from SEIKAGAKU (ARTZDispo, Japan), Oligosaccharide Hyaluronan sodium 6 mer was from pruchaed COSMOBIO. Co, Ltd. Other chemicals were purchased from Sigma (St Louis, MO). We thank Dr. Yu-Chih Liang, Ph. D kindly providing antibodies of COX-2, HO-1 PPARγ, ICAM-1, VCAM-1.. 2.2 Culture of chondrosarcoma and preparation of cell lysates. Chondrosarcoma cells were cultured in L-15 medium supplemented with 2 mM L-glutamine, 10% heat-inactivated FBS, and penicillin (100 U/mL)/streptomycin (100 mg/mL). Cells were attached to a Petri dish after 24h-48h incubation. Cells were plated at a concentration of 1 x 105 cells/mL and used for the experiment when they reached 80% confluency. Cultures were maintained in a humidified incubator without CO2 at 37oC. After reaching confluence, cells were treated with various concentrations of drug for indicated time intervals and incubated in a humidified 9.
(21) incubator at 37 ℃. In our experiments, cells were cotreatd with specific IL-1β for different time course. After incubation, cells were lysed by adding lysis buffer containing 10 mM Tris HCl (pH 7.5), 1 mM EGTA, 1 mM MgCl2, 1 mM sodium orthovanadate, 1 mM DTT, 0.1% mercaptoethanol, 0.5% Triton X-100, and the protease inhibitor cocktails (final concentrations: 0.2 mM PMSF, 0.1% aprotinin, 50 μg/mL leupeptin).. Cells adhering. to the plates were scraped off using a RUBBER POLICEMAN and stored at –70 oC for further measurements.. 2.3 WST-1 assay. The assay is based on the cleavage of the tetrazolium salt WST-1 to formazan by cellular mitochondrial dehydrogenases. Expansion in the number of viable cells resulted in an increase in the activity of the mitochondrial dehydrogenases, which leads to the increase in the amount of formazan dye formed. The formazan dye produced by viable cells can be quantified by multi-well spectrophotometer (microtiter plate reader) from pruchaed QIAGEN by measuring the absorbance of the dye solution at 450 nm.. 2.4 RT-PCR. Total RNA was extracted (REzol) from chondrosarcoma cells or chondrocytes treated with either vehicle or HMW-HA for 24 h or LMW-HA for 1, 3, 6, 12 and 24 h. cDNA was synthesized from 0.5 μg of total RNA with TaqMan reverse transcription reagents Applied Biosystems). Reverse transcription was performed in the Creacon technology PCR System beginning with an incubation period of 10 min at 25 °C 10.
(22) followed by a reverse transcription period of 30 min at 48 °C and ending with reverse transcription inactivation by 95 °C for 5 min. RT-PCR was carried out in the Creacon technology PCR System thermal cycler/detector in the following sequence: 5 min at 95 °C, 30 sec at 94 °C and 30 cycles consisting of 40 sec at 50 °C, 30 sec at 72 °C, 7 min at 72 °C after cooling down at 4 °C . The following primers were used: MMP-1 Forward : 5’-GTCAGGGGAGATCATCGG-3’ Reverse : 5’-GCCCAGTACTTATTCCCT-3’ MMP-13 Forward : 5’-GCTTAGAGGTGACTGGCAA-3’ Reverse : 5’-CCGGTGTAGGTGTAGATAGGAA-3’ COX-2 Forward : 5’-TTCAAATGAGATTGTGGGAAAT-3’ Reverse : 5’-AGATCATCTCTGCCTGAGTATCTT-3’ HO-1 Forward : 5’-CAGGCAGAGAATGCTGAGTTC-3’ Reverse : 5’-GCTTCACATAGCGCTGCA-3’ PPARγ Forward : 5’-GTATGACTCATACATAAAGT-3’ Reverse : 5’-TCTCGTGGACTCCATATTTG-3’ GAPDH Forward : 5’-CTGCCGTCTAGAAAAACC-3’ Reverse : 5’-CCAAATTCGTTGTCATACC-3’ 11.
(23) GAPDH was amplified as an internal control. The samples were loaded in triplicate, and the results of each sample were normalized to GAPDH.. 2.5 SDS PAGE and Western blotting. Electrophoresis was ordinarily carried out using different percentages of SDS-polyacrylamide. electrophoresis. (SDS-PAGE).. Following. electrophoresis,. proteins on the gel were electrotransferred onto a polyvinyldifluoride (PVDF) membrane. After transfer, the PVDF membrane was washed once with PBS and twice with PBS plus 0.1% Tween 20. The PVDF membrane was then blocked with blocking solution containing 3% bovine serum albumin in PBS containing 0.1% Tween 20 for 1 h or 5% milk blocking buffer in TBST at room temperature. The PVDF membrane was incubated with a solution containing primary antibodies in the blocking buffer. Finally, the PVDF membrane was incubated with peroxidase-linked anti-mouse IgG antibodies for 1 h and then developed using a LumiGLO chemiluminescence kit (Amersham, UK).. 2.6 Flow Cytometry (FACS). AnnexinⅤ has a high affinity in a Ca2+-dependent manner to negatively charged phospholipid phophatidylserine, which is found at the outer cell membrane early during apoptosis. The FITC conjugated AnnexinⅤ and PI stain. As the cell membrane becomes increasingly permeable during the later stage of apoptosis, PI can readily move across the cell membrane and bind to DNA. This combination allows the differentiation among 3 populations of cell in two-color flow cytometry, using 488 nm excitation and a 515 nm band pass filter for fluoresce in detection and a filter with 12.
(24) band pass over 600 nm for PI detection. 2.7 Statistics analysis For sample in which n=3, statistical analyses were performed using the analysis of variance (ANOVA). Our western band were be quantified to density mean by “Image-Pro Plus” (density-meter) software, and an illustration of quantification is used by SigmaPlot and Excel software.. 13.
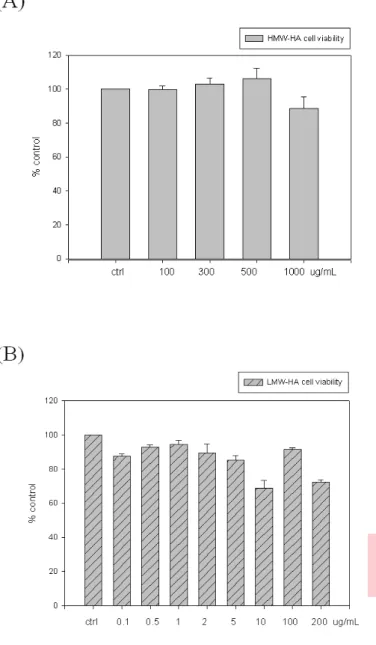
(25) 3. Results. 3.1 Dose effect of HMW-HA and LMW-HA on SW1353 cell viability. In order to know the effect of HA on viability of chondrosarcoma cell SW1353, various doses of HMW-HA or LMW-HA were used. As the data showed in the figure (Figure 1A), there were no significant difference on SW1353 viability after 0, 100, 300, 500, and 1000 μg/mL of HMW-HA treatment. However, in contrast to HMW-HA treatment, SW1353 cell treated with higher doses of LMW-HA (0, 0.1, 0.3, 0.5, 1, 3, 5, 10, 100 and 200 µg/mL) would lead to SW1353 cell death (Figure 1B). All HMW-HA and LMW-HA were concentration base on Molecular concentration.. 3.2 Dose dependent effect of HMW-HA on IL-1β induced MMP-1 and MMP-13 expression To evaluate the anti-inflammatory effect of HMW-HA, chondrosarcoma SW1353 was pretreated with HMW-HA and then activated by IL-1β. As shown in Figure 2, as a positive control, IL-1β could increase MMP-1 and MMP-13 mRNA level after 24 hour treatment (Figure 2A and 2B). However, pretreated with HMW-HA would decrease the mRNA expression level of MMP-1 and MMP-13 that induced by IL-1β. The mRNA level would decrease as HMW-HA dose increase. The inhibitory effect of HMW-HA could also be found in the protein level. As data showed in the Figure 4, protein expression of both MMP-1 and MMP-13 induced by IL-1β could be inhibited by HMW-HA pretreatment. Our findings are similar to the data reported previously that HA can regulate MMPs and other proteins expression in many cell [44, 70]. 14.
(26) 3.3 Dose dependent effect of LMW-HA on IL-1β induced MMP-1 and MMP-13 expression Although HMW-HA could inhibit the induction effect of IL-1β on MMP-1 and MMP-13 expression, effects of LMW-HA on anti-inflammatory was not clear. Therefore, SW1353 was pretreated with various concentrations (0, 0.1, 5, 10 μg/ml) of LMW-HA following with IL-1β treatment. However, not like HMW-HA, mRNA level of MMP-1 and MMP-13 were further increase after LMW-HA treatment (Figure 3). These finding could also be detected at protein level (Figure 5). The results indicated that LMW-HA may enhance the expression of inflammatory response mediator (MMPs) and induce cartilage matrix loss.. 3.4 Dose dependent effect of HA on COX-2 expression. In the previous study, IL-1β was showed to activate clooxygenase-2 (COX-2) and increase prostaglandin E2 (PGE2) production., These would subsequently increase joint. pain. and. further. synovial. inflammation[31].. Since. HMW-HA. has. anti-inflammatory effect, the ability of HMW-HA on IL-1β activation was evaluated. As shown in Figure 6, pretreated with HMW-HA would abolish the COX-2 expression that activated by IL-1β. This inhibitory effect could be detected in both RNA (Figure 6A) and protein level (Figure 6B). In the pilot study, LMW-HA was showed to enhance the inflammatory mediator (i.e MMP-1 and MMP-13) production. It indicated that LMW-HA may enhance COX-2 expression at disease site. In order to figure out the effects of LMW-HA on COX-2, SW-1353 cells was pretreated with LMW-HA and then activated by IL-1β. As shown in Figure 7, the COX-2 expression level would be induced by IL-1β. Further induction could be detected via LMW-HA pretreatment. 15.
(27) 3.5 Treatment of SW1353 with dose dependent manner of HMW-HA increase the level of HO-1 gene and protein expression HO-1 is induced by a variety of stimuli or agents that cause oxidative stress, such as cytokines, reactive oxygen species, nitric oxide, heat shock, and hypoxia. HO-1 induction has also been shown to down-regulate the inflammatory response in animal models of acute inflammation [35]. But recently several reports have identified the heat shock protein 32 (Hsp32)/heme oxygenase-1 (HO-1) as a BCR/ABL-dependent survival molecule in CML cells and murine collagen-induced arthritis (CIA) model [37] [71]. Treatment with SnPP, which an inhibitor of HO-1, could significantly reduced the severity of CIA through inhibition of joint inflammation and cartilage destruction. We herein pretreated Chondrosarcoma SW1353 with HMW-HA to evaluate its effect on anti-inflammation. As data shown in Figure 8, SW-1353 express moderate amount of HO-1 and IL-1β treatment would not further inducing HO-1 expression.. However, pretreated with HMW-HA would down regulate HO-1. expression at both RNA and protein level (Figure 8A and 8B). In previous papers shown, HO-1 overexpression cannot slow the progression of the chronic inflammatory disease, whereas treatment with SnPP, which inhibits HO-1[38]. To examine the effects of LMW-HA on HO-1 expression, SW-1353 was pretreated with LMW-HA. Unlike the results obtained by HMW-HA treatment, insteated of down regulation, LMW-HA would enhance HO-1 expression in SW1353 cell (Figure 9). 3.6. Effects of HA on the expression of Nuclear receptor, PPARγ. Others have previously shown that PPARγ activators display anti-inflammatory and chondroprotective properties in vitro. However, the expression and regulation of PPARγ expression in cartilage are poorly defined. To determine the influence of 16.
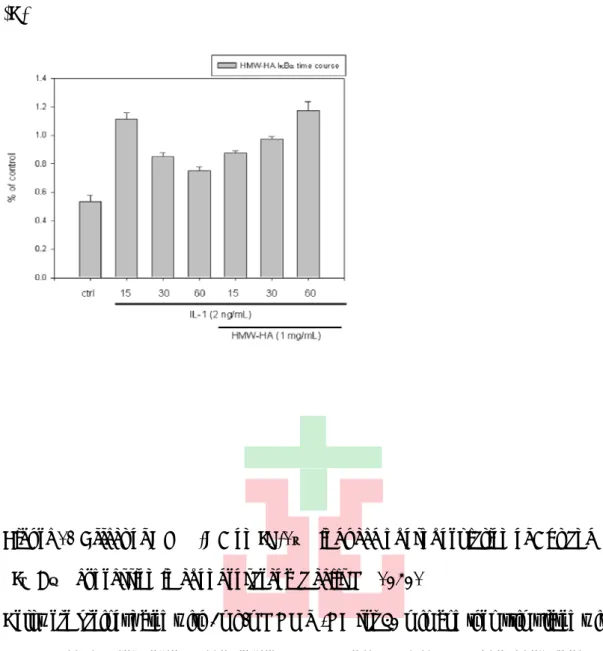
(28) HMW-HA on the synthesis of. MMP-13、MMP-1、COX-2、HO-1 and IL-1β in. SW1353 of osteoarthritis (OA). As show in Figure 10(A) Figure 10 (B), Cell were preincubated with various concentrations of HMW-HA ( 0, 0.1, 0.3, 0.5 and 1 mg/mL ) for 30 minutes and then stimulated IL-1β 2 ng/mL for 24 hrs, After 24 hours, cell lysates were prepared and analyzed for peroxisome proliferator-activated receptor γ (PPARγ) protein expression by Immunoblotting. These data strongly suggest that HMW-HA with 1mg/mL increased PPARγ expression. According to Figure 10(A) Figure 10 (B) shown, to assess the contribution of these pathways in the HMW-HA-mediated upregulation of PPARγ, we found the MAPKs JNK and p38, but not ERK, are involved in HMW-HA-mediated upregulation of PPARγ. Taken together, Figure 12 shown, this data suggested that Mediation of HMW-HA-mediated upregulation of PPARγ by NF-κB inhibition. As show in Figure 11(A), Figure 11(B). Cell were preincubated with various concentrations of LMW-HA ( 0, 0.1, 5, 10 μg/mL ) for 30 minutes and then stimulated IL-1β 2 ng/mL for 24 hrs. After 24 hours, cell lysates were prepared and analyzed for peroxisome proliferator-activated receptor γ (PPARγ) protein expression by Immunoblotting. These data suggest that LMW-HA decreased PPARγ expression through promoting phosphorylation of p38 MAPK pathway.. 3.7 Effect of HMW-HA on phospho-Akt phosphorylation and IκBαexpression. According to pervious study, Glucosamine can induce p-Akt activation. To examine that HMW-HA can active pAkt the same as Glucosamine, so I used the maximum dose of HMW-HA, 1 mg/mL for time course. Figure 12(A) shown, we found HMW-HA can active IL-1β -induced pAkt activation. To base on HMW-HA up-regulated Akt phosphorylation, we examine whether the 17.
(29) HMW-HA reduces inflammatory elements through up-regulated p-Akt and inhibited p- IκBα phosphorylation. we used the maximum dose of HMW-HA, 1 mg/mL for time course. Figure 12(B) shown, we found HMW-HA can inhibit IL-1β-induced pIκBα phosphorylation after 30 minutes and inhibit total-IκBα degradation efficiently. Figure 12(C). 3.8. Effect of HMW-HA on phosphorylation of MAPK pathways induced by IL-1β. In pervious papers shown, HA suppresses IL-1β-enhanced MMP-1 and MMP-3 synthesis in RSF (rheumatoid synovial fibroblasts) via ICAM-1 through down-regulation of NF-κB and p38 [21]. To examined the phosphorylation effect of HMW-HA, cells were pretreated HMW-HA 1 mg/mL for 30 minutes and then stimulated with IL-1β 2 ng/ml for different time course ( 0, 15, 30 and 60 min ) . Cell lysates is analyzed by immunoblotting. Figure 13(A) result that IL-1β is able to activate the phosphorylation of ERK starting at 15 min. Pretreatment with HMW-HA for 30 min, phosphorylation of ERK seems to slightly inhibited and lead to cell proliferation inhibited by HMW-HA at 15 min. As in Figure 13(B) shown, this result the level of inflammatory protein secretion was decrease by treated HMW-HA and phosphorylation of JNK MAPK pathway, which pro-inflammtroy pathway was inhibited by HMW-HA at 60 minutes. This result was demonstrated that HMW-HA reduces inflammatory mediator through JNK MAPK pathway.. 18.
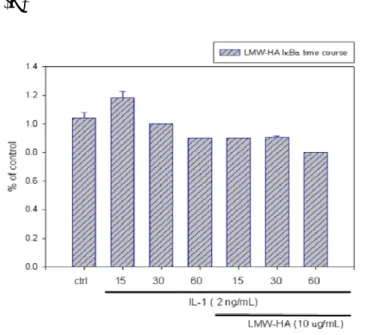
(30) 3.9. HMW-HA regulates cyclin D1/cdk4 protein expression of cell cycle. As in Figure 13(A), result that IL-1β is able to activate the phosphorylation of ERK starting at 15 min. Pretreatment with HMW-HA for 30 min, phosphorylation of ERK seems to slightly inhibited by HMW-HA as the same time as stimulated by IL-1β. Hyaluronic acid were used in the clinical treatment of osteoarthritis, The result shown that HMW-HA had inhibitory effect of cyclin D1 and Cdk4 in dose-dependent manner and time-course. (Figure 14A&B). 3.10. Effect of LMW-HA affect phospho-Akt phosphorylation and IκBα expression. Recently papers shown, LMW-HA treatment decreased tumor cell proliferation, increased apoptosis, and downregulated activation of Akt and the expression of BCRP in glioma cells and treatment-resistant glioma stem cells [72]. To examine the effect of LMW-HA affect p-Akt in SW1353, I used the maximum dose of LMW-HA, 10 μg/mL for time course. Figure 15(A) shown, we found LMW-HA can down-regulate IL-1β-induced p-Akt activation and induced apoptosis in SW1353 the same as in glioma cells. To determine whether LMW-HA induced p- IκBα phosphorylation and induced TIκBα degradation. I used the maximum dose of LMW-HA, 10 μg/mL for time course. Figure 15(B)(C) shown, we found LMW-HA can induce inflammatory elements through down-regulate p-Akt and induce NF-κB signaling pathway, after 15 minutes.. 19.
(31) 3.11. Effect of LMW-HA on phosphorylation of MAPK pathways induced by IL-1β. In pervious papers findings suggest that hyaluronan- CD44 interactions affect matrix metabolism via activation of NF-κB and p38 MAP kinase in C-28/I2 cell [73]. To determining that pro-inflammatory cytokine IL-1β as well as LMW-HA may be responsible for this up-regulation via a mechanism involving activation of the MAPKs (p38 and JNK) and NF-κB signaling pathways. In my poilt study konwn, LMW-HA can produce proinflammatory mediators, so we use different dose of LMW-HA ( 0, 0.1 5, 10 μg/mL ) treat chondrosarcoma SW1353 and detected by immunoblotting for different time course ( 0, 15, 30, 60 min ). As shown in Figure 16, LMW-HA is a strong inflammatory inducer for increasing p38 phosphorylation at 15 min.. 20.
(32) 4. Discussion. Osteoarthritis (OA) is a gradually progressing disorder of mammalian joints, characterised by the destruction of articular cartilage, which results in discomfort and dysfunction of the affected joint [1, 2]. The pathologic changes during the development of OA are remarkably similar and include proteoglycan degradation at the early stage, followed by type II collagen degradation, leading eventually to localized or complete loss of cartilage matrix [3]. Hyaluronan (HA) is a high molecular weight nonsulphated glycosaminoglycan (GAG) component of the extracellular matrix (ECM) present in many tissues. HA has many structural, rheological and physiological functions in tissues, including ECM and cellular interaction, growth factor intercaton, the regulation of osmosis and wound healing,. immune response, re-epithelialixation stages [45]. Some HA plays. several roles in the anti-inflammatory response, including the anti oxidant scavenging of ROS and other sources [21], these data suggest that, during oxidative stress and inflammation. Previously studies used high molecular weight HA and the opposite result was obtained. It is hypothesized that the inhibition of NF-κB DNA binding to the nucleus is probably the consequence of HA reduced ROS and MMPs production in the fibroblasts. NF-κB activation requires sequential phosphorylation, and degradation of IκBα, that, in the end, disappears from the cytoplasm. According to these finding we use of HMW-HA (0.1-1 mg/mL) may reduce cell damage by inhibiting NF-κB as well as protecting cells from inflammatory mediators and MMP-1 and MMP-13 attack. Hyaluronan oligosaccharides also activate NF-κB /I-κB α auto-regulatory loop [61], inducing transcription of metalloproteases MMP-9 and MMP-13 [43]. The induction of nitric-oxide synthase by such saccharides also occurs through a nuclear 21.
(33) NF-κB-dependent mechanism [62], and up-regulated JNK and p38 mitogen-activated protein kinase expression. According to these findings we also use HA oligosaccharides (0.1-10μg/mL) found it could induce MMP-1, MMP-13 and other inflammatory mediators, including COX-2 and HO-1 through induction of p38 MAP kinase and NF-κB pathway.. It has been proposed that production of VEGF after HO-1 overexpression in murine CIA ( Collagen-induced arthritis ) would promote angiogenesis. In the murine collagen-induced arthritis (CIA) model, HO-1 overexpression cannot slow the progression of the chronic inflammatory disease [38]. Our studies showed HO-1 expression during OA, whereas treatment with HMW-HA, which inhibits HO-1 expression [38]. It is interesting to note that recently observed a therapeutic effect of HO-1 inhibition via HMW-HA (0.1-0.5 mg/mL) in my study. The data presented here provide evidence to support the essential role of HO-1 in the negatively regulated anti-inflammatory function. It was previously reported that the LMW-HA may induce injury by MMPs and others inflammatory mediators in both in vitro and in vivo [43]. According to these data, we suggest that LMW-HA (0.1-10 μg/mL) also could induce inflammatory mediators, including COX-2 and HO-1, the same as induce NF-κB pathway. The data also presented here provide evidence to support the essential role of HO-1 in the negatively regulated induce inflammatory function by injury.. PPARs are a family of nuclear hormone receptors that function as ligand-activated transcription factors that upon heterodimerization with RXR, function as transcriptional regulators of glucose and lipid metabolism. PPARβ/δ is involved in development, wound healing, fatty-acid metabolism and repression of the 22.
(34) inflammatory response. It has recently been discovered that PPARs are also strongly linked to inflammatory reaction. Inflammation inducers such as lipopolysaccharide (LPS) and tumor necrosis factor (TNFα) induce activation of NF-κB, a major transcription factor in the inflammatory process, and promote the secretion of a series of inflammatory cytokines in various cells. PPARα and PPARγ ligands can block the NF-κB pathway, modulating inflammatory reaction. We herein demonstrate that PPARγ exists expression by treatment HMW-HA (0.1-0.5 mg/mL), the same as increased anti-inflammatory after stimulation with IL-1β in SW1353. The data suggest that the ligand for PPARγ inhibit COX-2 and suppress inflammatory mediators. On the other hand, we also demonstrate that induction of inflammatory response by treatment with LMW-HA (0.1-10 μg/mL), and induction of NF-κB pathway. It was previously reported that chondroitin-sulfate (CS) inhibited TNF-α-induced NF-κB activation and inducle nitric oxide synthase expression by blocking Akt/NF-κB signals in JB6 cells [74].. It has recently been discovered that the PI3K/Akt signaling pathway has been shown to play an important role in negatively regulating LPS-induce acute inflammatory responses in vitro and in vivo. Recently paper shown that α-Lipoic acid attenuates LPS-induced inflammatory responses by activating the PI3K /Akt signaling pathway in cultured human monocytic cells [75]. The PI3K/Akt pathway has been shown to act both positively and negatively on NF-κB-dependent pathway. These differences may be due to the use of different cell types and / or different stimulations. For example, in human monocytic cels, TLR-2-dependent PI3K/Akt pathway was shown to positively regulate the transactivation potential of p65. However, PI3K /Akt negatively regulated p65 23.
(35) transactivation induced by TLR4 activation. According to these data, we suggest that HMW-HA (1 mg/mL) time-course was able to activate the Akt phosphorylation and Inhibit NF-κB phosphorylation. We also demonstrate LMW-HA (10 μg/mL) time-course resulted from an enhance activation of NF-κB and p38 MAPK signaling cascade and a reduced activation of the PI3K/Akt pathway following LMW-HA treatment. Taken together, these data suggest that HA plays a critical role negatively regulating NF-κB activity by activation of the PI3K/Akt pathway. In the current study, Peroxisome proliferator-activated receptor γ (PPARγ) ligands inhibit cell proliferation and induce apoptosis in cancer cells. Recently paper shown, troglitazone treatment, applied in a dose-dependent manner, cause a marked decrease in pRb, cyclin D1, cyclin D2, cyclin D3, Cdk2, Cdk4 and Cdk6 expression as well as significant increase in p21 and p27 expression in MDA-MB-231 cells [76]. In our study, we demonstrated that HMW-HA (0.1-1mg/mL) dose-dependent manner and time-course , resulted decrease in cyclin D1 and Cdk4 expression the same as promote cell cycle arrest in SW1353. Moreover, the PPARγ receptor has been shown to be implicated in carcinogenesis and inflammation. We herein suggested that the HMW-HA (0.1-0.5 mg/mL) inhibits in a dose-dependent manner IL-1β-mediated proinflammatory response by interfering with the phosphorylation of ERK1/2 inactivation.. CD44 is encoded by a single gene, but multiple isoforms of CD44 are generated by alternative RNA splicing. The gene for CD44 contains 20 exons. The nonvariant exons encode for an extracellular domain, a transmembrane domain, and an intracellular domain. Isoforms of CD44 are generated by the insertion of alternative exons (V1–V11) at a single site within the membrane-proximal portion of the extracellular domain The predominant 72–amino acid cytoplasmic domain can also be 24.
(36) replaced by an alternatively spliced shorter form. Differential posttranslational modifications, including glycosylation and the attachment of glycosaminoglycans, generate additional structural diversity of CD44. The regulation of the affinity of cell adhesion molecules is prerequisite for regulating cell–cell and cell–matrix interactions mediated by broadly expressed receptors that are exposed continuously to their ligands. Most primary cells express CD44 but in a low affinity state that does not exhibit a capacity to bind to HA. Cellular activation can induce a transition of CD44 to a high affinity state that mediates binding to HA. Transition from the “inactive” low affinity state to the “active” high affinity state of CD44 on leukocytes can be induced by the ligation of antigen receptors, and on leukocytes and epithelial and other mesenchymal cells by soluble factors including cytokines [77, 78]. A variety of mechanisms have been implicated in the transition from inactive to active forms of CD44, including variant exon usage, receptor oligomerization, glycosylation, and sulfation [79]. However, to date, no data are available to indicate how these posttranslational modifications alter either the configuration of the receptor, its three-dimensional structure, or its molecular interactions with other moieties to modify the affinity of the receptor for HA. In our study, we could not demonstrate that CD44 has high or low affinity state mediates binding to HMW-HA and / or LMW-HA, could not demonstrate that CD44 splicing form in SW1353, and could not demonstrate that CD44 receptor conformation change or posttranslation control (glycosylation, sulfation) by interacting with HMW-HA and /or LMW-HA. In contrast to normal primary cells, many tumor-derived cells express CD44 in a high affinity state with capacity to mediate constitutive binding to HA. In addition to being a receptor for HA, CD44 can interact with several ECM proteins, such as 25.
(37) fibronectin and collagens, growth factors, cytokines and chemokines, as well as metalloproteinases [79], but less is known about the regulation of the interactions of these ligands with CD44. Transmembrane CD44 serves multiple roles, including mediating the metabolism of HA [80], in the regulation of tumor invasiveness and in the modulation of inflammatory cell function. Alterations in CD44 expression and structure have been documented in many types of cancer and are related to tumor dissemination [79]. Moreover, targeted deletion of CD44 prevented dissemination of some tumors [81]. Most of the known effects of CD44 on cell adhesion, migration, and metastasis are intimately associated with its capacity to promote cell attachment to HA. Recent findings suggest that CD44 might also promote metastasis through its association with other molecules. For example, CD44 provides a docking site for MMP-9 on the surface of melanoma and carcinoma cells [82] and thus can indirectly contribute to pericellular proteolysis to regulate tumor cell motility, growth factor activation, angiogenesis, as well as survival mechanisms. In our study, may think that variable exons of CD44,or COOH terminal of CD44 or different amino acid binding site of CD44 variable exons, or HMW/LMW-HA interact with CD44 binding affinity might provide docking site with MMP-1, MMP-13 surface and LMW-HA of SW1353 and regulate proinflammatory response. And in certain circumstances HMW-HA, that inhibited proinflammatory cytokine and LMW-HA inductuion of HA binding to CD44 in SW1353. It has recently been discovered that the Ras signaling pathway is important in both cell proliferation and tumor progression. Alternatively spliced isoforms of CD44 containing variable exon 6 (v6) can serve as coreceptors for growth factor receptors that activate Ras. This paper shown that use v6-specific small interfering RNA (siRNA) to investigate the role of CD44 alternative splicing in Ras signaling. Authors identify a positive feedback loop in which Ras signaling promotes CD44v6 splicing, 26.
(38) and CD44v6 then sustains late Ras signaling, which is important for cell cycle progression. These results are the first demonstration of a positive feedback loop linking signaling-dependent alternative splicing to mitogenic progression. The production of CD44 variants through alternative splicing is regulated by splicing factors, such as Sam68 and SRm160, and stimulated by Ras/MAPK (mitogenactivated protein kinase) signaling (Ras–Raf–MEK–ERK). These splicing factors depend upon exon splicing enhancers in the CD44 variable exons. In our study, we were not demonstrate that CD44 variants by some splincing factors in SW1353, and we do not know that splicing factors stimulated by Ras/MAPK signaling. Their activity in promoting the inclusion of CD44 variable exons is controlled by Ras/MAPK signaling, at least in part through modification of splicing factors at the level of phosphorylation. However, the signaling pathway between Ras/MAPK activation and stimulation of alternative splicing in the nucleus is not well defined. Interestingly, CD44 variants containing variable exon 6 (v6) have been shown to be important for Ras activation through formation of a coreceptor complex with the hepatocyte growth factor HGF and its receptor tyrosine kinase Met, whose phosphorylation activates Ras signaling [83]. Addition of growth factors to quiescent cells results in biphasic activation of the Ras/PI3k/MAPK signaling pathways [84], with an early rapid increase in MAPK activation from 5 to 60 min, followed by a late second wave of MAPK activation of lower amplitude beginning at ~7–10 h. In our study, we demonstrate that HMW-HA and /or LMW-HA through CD44 inactive JNK and activate p38 of MAPKs from 15-60 min. This second wave of MAPK activity is sustained for hours after growth factor stimulation and is critical for cells to progress from G1 into S phase [85]. Despite its importance, however, mechanisms underlying the sustained activity of Ras/MAPK 27.
(39) signaling have remained elusive. Here we demonstrate HMW-HA inhibit ERK pathway the same as inhibit cyclin D1/Cdk4 of cell cycle, result in cell cycle arrest. In conjunction with growth factors and their receptor tyrosine kinases, specific CD44v6 variants produced by alternative splicing subsequently enhance the late wave of Ras/ MAPK activation to facilitate cell cycle progression. In our study, as in Figure 13(B) shown, this result the level of inflammatory protein secretion was decrease by treated HMW-HA and phosphorylation of JNK MAPK pathway, which pro-inflammatory pathway was inhibited by HMW-HA at 60 minutes. This result was demonstrated that HMW-HA reduces inflammatory mediator through JNK MAPK pathway. HMW-HA suggest to inhibit the activity of JNK and NF-κB in SW1353 through CD44. As shown in Figure 16, we suggest that LMW-HA is a strong inflammatory inducer for increasing p38 phosphorylation of MAPKs.. 28.
(40) Figure 1. Effect of HMW-HA and LMW-HA on cell viability in human chondrosarcoma cells SW1353 (A) Cell were treated with various concentration of HMW- HA (100, 300,500,1000 μg/mL) and (B) cell were treated with various concertration of LMW-HA (0.1-200 μg/mL) for 24 hrs. The extent of cell viability was determined by WST-1 as described in “Methods”. Data are expressed as mean ± SD.. 29.
(41) (A). 30.
(42) Figure 2. Effects of HMW-HA on IL-1β induced MMPs gene expression in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of HMW-HA ( 0.1, 0.3, 0.5, 1 mg/mL) for 30 min and then stimulated with 2 ng/mL IL-1β for 24 hrs. Cell lysates were collected at end of study and analyzed with (A) MMP-1, (B) MMP-13. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments.. 31.
(43) (A). (B). 32.
(44) Figure 3. Effects of LMW-HA on IL-1β- induced MMP-1 and MMP-13 gene expression in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of LMW- HA ( 0.1, 5, 10 μg/mL) for 30 min and then stimulated with IL-1β 2 ng/mL for 24 hrs. Cell lysates were collected at end of study and analyzed with (A) MMP-1 mRNA, (B) MMP-13 mRNA. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 33.
(45) (A). *. *. (B). *. 34.
(46) Figure 4. Effects of HMW-HA on IL-1β-induced MMPs protein prodution in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of HMW-HA (0.1, 0.3, 0.5, 1 mg/mL) for 30 min and then stimulated with IL-1β 2 ng/mL for 24hrs. Cell lysates were collected at end of study and analyzed with (A) MMP-1, (B) MMP-13. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 35.
(47) (A). *. (B). 36.
(48) Figure 5. Effects of LMW-HA on IL-1β induced MMPs protein expression in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of LMW- HA (0.1, 5, 10 μg/mL) for 30 min and then stimulated with IL-1β 2 ng/mL for 24 hrs. Cell lysates were collected at end of study and analyzed with (A) MMP-1, (B) MMP-13. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 37.
(49) (A). *. (B). 38. *. *.
(50) Figure 6. Effects of HMW-HA on IL-1β induces COX-2 gene and protein expression in chondrosarcoma cells SW1353. Cell were preincubatedwith various concentration of HMW- HA ( 0.1, 0.3, 0.5, 1 mg/mL) for 30 min and then stimulated with IL-1β 2 ng/mL for 24 hrs. Cell lysates were collected at end of study and analyzed with (A) COX-2 mRNA, (B) COX-2 protein. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 39.
(51) (A). *. (B). *. 40.
(52) Figure 7. Effects of LMW-HA on IL-1β induces COX-2 gene and protein expression in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of LMW- HA (0.1, 5, 10 μg/mL) for 30 min and then stimulated with IL-1β 2 ng/mL for 24 hrs. Cell lysates were collected at end of study and analyzed with (A) COX-2 RNA , (B) COX-2 protein. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 41.
(53) (A). *. *. (B). *. 42.
(54) Figure 8. Effects of HMW-HA on IL-1β induces HO-1 gene and protein expression in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of HMW- HA (0.1, 0.3, 0.5, 1 mg/mL) for 30 min and then stimulated with IL-1β 2 ng/mL for 24 hrs. Cell lysates were collected at end of study and analyzed with (A)HO-1 mRNA and (B) HO-1 protein. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as Mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 43.
(55) (A). (B). * *. 44.
(56) Figure 9. Effects of LMW-HA on IL-1β induces HO-1 gene and protein expression in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of LMW- HA ( 0.1, 5, 10 μg/mL) for 30 min and then stimulated with IL-1β2 ng/mL for 24 hrs. Cell lysates were collected at end of study and analyzed with (A) HO-1 mRNA and (B) HO-1 protein. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 45.
(57) (A). *. *. (B). * *. 46.
(58) Figure 10.. Effects of HMW-HA on IL-1β-induces PPARγgene and protein. expression in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of HMW- HA ( 0.1, 0.3, 0.5, 1 mg/mL) for 30 min and then stimulated with IL-1β 2 ng/mL for 24 hrs. Cell lysates were collected at end of study and analyzed with (A) PPARγ mRNA and (B) PPARγ protein. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 47.
(59) (A). *. *. *. (B). * *. 48.
(60) Figure 11. Effects of LMW-HA on IL-1β−induces PPARγgene and protein expression in chondrosarcoma cells SW1353. Cell were preincubated with various concentration of LMW- HA ( 0.1, 5, 10 μg/mL) for 30min and then stimulated with IL-1β 2 ng/mL for 24 hrs. Cell lysates were collected at end of study and analyzed with (A) PPARγ mRNA, (B) PPARγ protein. Equal loading in each lane was demonstrated by the similar intensities of GAPDH. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 49.
(61) (A). (B). 50.
(62) (C). Figure 12 Effect of HMW-HA on IL-1β induced phosphorylation of Akt and IκBα expression in chondrosarcoma cells SW1353. Cell were preincubated with 1mg/mL HMW-HA for 30 min and then stimulated with IL-1β 2 ng/mL for various time periods ( 15, 30 and 60 min). (A) phosphorylated-Akt, (B)phosphorylated-IκBα and (C)Total-IκBα were detected by western blot as described in “Method”. Equal loading in each lane was demonstrated by the similar intensities α-tubulin. Data are expressed as mean ± SD from experiments.. 51.
(63) (A). (B). 52.
(64) (C). Figure13. Effect of HMW-HA on IL-1β−induced phosphorylation of ERK, JNK, p38 in chondrosarcoma cells SW1353. Cell were preincubated with 1mg/mL HMW-HA for 30 min and then stimulated with IL-1β 2 ng/mL for various time periods ( 15, 30 and 60 min). (A) phosphorylated-ERK, (B) phosphorylated-JNK, or (C) phosphorylated-p38 was detected by western blot as described in “Method”. Equal loading in each lane was demonstrated by the similar intensities α-tubulin. Data are expressed as mean ± SD from experiments.. 53.
(65) % of control. (A). IL-1 (2 ng/mL) HMW-HA (1 mg/mL). -. + -. + 0.1. + 0.3. HMW-HA cdk4 protein. (B). 54. + 0.5. + 1.
(66) Figure 14 Effect of HMW-HA on IL-1β− induced cyclin D1 ans Cdk4 expression in chondrosarcoma cells SW1353 Cell were preincubated for 30min with various concentration of HMW- HA ( 0.1, 0.3, 0.5, 1 mg/mL) and then stimulated with IL-1β 2 ng/mL for 24 hrs.(A) Cyclin D1/Cdk4 protein. Cell were preincubated with 1mg/mL HMW-HA for 30 min and then stimulated with IL-1β 2 ng/mL for various time periods ( 15, 30 and 60 min) (B) cyclin D1 time course. Equal loading in each lane was demonstrated by the similar intensities of GAPDH and α-tubulin. Data are expressed as mean ± SD from experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 55.
(67) (A). (B). 56.
(68) (C). Figure 15 Effect of LMW-HA on IL-1β−induced phosphorylation of Akt and IκBα expression in chondrosarcoma cells SW1353. Cell were preincubated for 30 min with 1mg/mL LMW-HA and then stimulated with IL-1β2 ng/mL for various time periods (15, 30, 60 min). (A) phosphorylated-Akt,(B)phosphorylated-IκBα and (C)Total- IκBα were detected by western blot as described in “Method”. Equal loading in each lane was demonstrated by the similar intensities α-tubulin. Data are expressed as mean ± SD from experiments.. 57.
(69) (A). (B). 58.
(70) Figure 16 Effect of LMW-HA on IL-1β−induced phosphorylation of ERK, JNK, p38 in chondrosarcoma cells SW1353. Cell were preincubated for 30 min with 1mg/mL LMW-HA and then stimulated with IL-1β 2 ng/mL for various time periods (15, 30, 60 min) and (A) phosphorylated-ERK, phosphorylated-JNK, and (B) phosphorylated-p38 was detected by western blot as described in “Method”. Equal loading in each lane was demonstrated by the similar intensities GAPDH. Data are expressed as mean ± SD from three independent experiments.. 59.
(71) 120. 100. % of contro;\l. 80. 60. 40. 20. 0. IL-1 (2 ng/mL) LMW-HA (10 ug/mL). -. + -. + 0.1. + 5. + 10. LMW-HA cdk4 protein. 120. 100. % of control. 80. 60. 40. 20. 0. IL-1 (2 ng/mL) LMW-HA (10 ug/mL). -. + -. + 0.1. + 5. LMW-HA cyclin D1 protein. 60. + 10.
(72) Figure 17 Effect of LMW-HA on IL-1β−induced cyclin D1 ans Cdk4 expression in chondrosarcoma cells SW1353 Cell were preincubated for 30 min with various concentration of HMW- HA ( 0.1, 5, 10 μg/mL) and then stimulated with IL-1β2 ng/mL for 24 hrs and (A) Cyclin D1, (B ) Cdk4 protein. Equal loading in each lane was demonstrated by the similar intensities of α-tubulin. Data are expressed as mean ± SD from three independent experiments. *, P<0.05 as compared with the positive control group with IL-1β treatment.. 61.
(73) Figure 18 HMW-HA signaling pathway in SW1353. 62.
(74) Figure 19 Proliferator-activated receptor γ (PPARγ) induction by HMW-HA signaling pathway in SW1353. 63.
(75) Figure 20 LMW-HA signaling pathway in SW1353. 64.
(76) APPENDIX 1 LMW HA induces cell apoptosis. LMW-HA (HA6). apoptotic cell (% of control). 3 2.5 2 early apoptotic cell late apoptotic cell. 1.5 1 0.5 0 control. IL-1. 10. 100. 200. ug/mL. 65.
(77) 66.
(78) 5.Reference. 1.. 2. 3.. 4. 5.. 6.. 7.. 8. 9.. 10.. 11.. 12.. Cahue, S., et al., The ratio of type II collagen breakdown to synthesis and its relationship with the progression of knee osteoarthritis. Osteoarthritis and Cartilage, 2007. 15(7): p. 819-823. Yelin, E., Arthritis: The cumulative impact of a common chronic condition. Arthritis and Rheumatism, 1992. 35(5): p. 489-497. Homandberg, G.A., Cartilage damage by matrix degradation products: Fibronectin fragments. Clinical Orthopaedics and Related Research, 2001(391 SUPPL.). Ala-Aho, R. and V.M. Ka?ha?ri, Collagenases in cancer. Biochimie, 2005. 87(3-4 SPEC. ISS.): p. 273-286. Sternlicht, M.D. and Z. Werb, How matrix metalloproteinases regulate cell behavior. Annual Review of Cell and Developmental Biology, 2001. 17: p. 463-516. Sounni, N.E. and A. Noel, Membrane type-matrix metalloproteinases and tumor progression. Biochimie, 2005. 87(3-4 SPEC. ISS.): p. 329-342. Chang, C. and Z. Werb, The many faces of metalloproteases: Cell growth, invasion, angiogenesis and metastasis. Trends in Cell Biology, 2001. 11(11). Stamenkovic, I., Extracellular matrix remodelling: The role of matrix metalloproteinases. Journal of Pathology, 2003. 200(4): p. 448-464. Young, D.A., et al., Histone deacetylase inhibitors modulate metalloproteinase gene expression in chondrocytes and block cartilage resorption. Arthritis research & therapy., 2005. 7(3). Heist, R.S., et al., Matrix metalloproteinase polymorphisms and survival in stage I non-small cell lung cancer. Clinical Cancer Research, 2006. 12(18): p. 5448-5453. Nasr, H.B., et al., Matrix metalloproteinase-1 (-1607) 1G/2G and -9 (-1562) C/T promoter polymorphisms: Susceptibility and prognostic implications in nasopharyngeal carcinomas. Clinica Chimica Acta, 2007. 384(1-2): p. 57-63. Qi, C. and H. Changlin, Levels of biomarkers correlate with magnetic resonance imaging progression of knee cartilage degeneration: A study on canine. Knee Surgery, Sports Traumatology, Arthroscopy, 2007. 15(7): p. 869-878. 67.
數據
相關文件
The method of least squares is a standard approach to the approximate solution of overdetermined systems, i.e., sets of equations in which there are more equations than unknowns.
Center for Faculty Development, Department of medical education, Taichung Veterans General hospital 1 School of Medicine, National Yang-Ming University, Taipei, Taiwan 2.
臺大機構典藏NTUR (National Taiwan University 二 Repository, http://ntur.lib.ntu.edu.tw) 經驗與協助推 動臺灣學術機構典藏TAIR (Taiwan Academic Institutional Repository,
The resulting color at a spot reveals the relative levels of expression of a particular gene in the two samples, which may be from different tissues or the same tissue under
We do it by reducing the first order system to a vectorial Schr¨ odinger type equation containing conductivity coefficient in matrix potential coefficient as in [3], [13] and use
由於 A 為方陣, 其 row 的個數和 column 的個數皆為 n, 此時很自然地可以將 Theorem 3.4.2 和 Theorem 3.4.6 相連結得到 invertible
introduction to continuum and matrix model formulation of non-critical string theory.. They typically describe strings in 1+0 or 1+1 dimensions with a
1 Department of Neurology and Stroke Center, Taipei Medical University-Shuang Ho Hospital; Taipei Neuroscience Institute; School of Medicine, College of Medicine, Taipei
