Jian-Ying Chuang
2, Elisabetta Bianchi
3and Yi-Ren Hong
41Institute of Microbiology and
Biochemistry and Department of Biochemical Science and Technology, National Taiwan University, No.1, Sec. 4, Roosevelt Rd., Taipei, 10617, Taiwan
2Molecular Genetics Laboratory, Department of Microbiology and Immunology, Chang-Gung University, 259, Wen-Hwa 1st Rd., Kwei-Shan, Taoyuan, 333, Taiwan 3Laboratory of Immunoregulation, Department of Immunology, Institut Pasteur, 25 rue du Dr. Roux, 75724 Paris, France 4Graduate Institute of Biochemistry, Kaohsiung Medical University, 100, Shih-Chuan 1st Rd., Kaohsiung, 807, Taiwan Received 7 January 2008; received in revised form 1 April 2008;
accepted 1 April 2008 Available online 10 April 2008
Epstein-Barr virus (EBV) expresses the immediate-early protein Rta to activate the transcription of EBV lytic genes and the lytic cycle. We show that RanBPM acts as a binding partner of Rta in yeast two-hybrid analysis. The binding was confirmed by glutathione-S-transferase pull-down assay. A coimmunoprecipitation experiment and confocal microscopy revealed that RanBPM and Rta interact in vivo and colocalize in the nucleus. The interaction appears to involve the SPRY domain in RanBPM and the region between amino acid residues 416 to 476 in Rta. The interaction promotes the transactivation activity of Rta in activating the transcription of BMLF1 and p21 in transient transfection assays. Additionally, RanBPM interacts with SUMO-E2 (Ubc9) to promote sumoylation of Rta by SUMO-1. This fact explains why the expression of RanBPM enhances the transactivation activity of Rta. Taken together, the present results indicate a new role of RanBPM in regulating a viral protein that is critical to EBV lytic activation.
© 2008 Elsevier Ltd. All rights reserved.
Edited by J. Karn Keywords: Epstein-Barr virus; Rta; RanBPM
Introduction
Epstein-Barr virus (EBV) is a human herpesvirus that infects lymphoid and epithelial cells. This virus is typically maintained under latent conditions after infection and must undergo a lytic cycle to prolif-erate. In the immediate-early stage of the lytic cycle, the virus expresses two immediate-early transcrip-tion factors, Rta and Zta, encoded by BRLF1 and BZLF1, respectively, to activate the EBV lytic cas-cade.1,2Additionally, Rta and Zta often cooperate to activate synergistically many EBV lytic genes, inclu-ding BRLF1 and BMRF1.3,4However, Rta may also
*Corresponding author. E-mail address:
Abbreviations used: EBV, Epstein-Barr virus; SUMO, small ubiquitin-like modifier; E2, ubiquitin carrier protein ligase; E3, ubiquitin-protein isopeptide ligase; TPA, 12-O-tetradecanoylphorbol 13-acetate; GST, glutathione S-transferase; RRE, Rta-response element; MG132, N-carbobenzoxy-leucyl-leucyl-leucinal; GFP, green fluorescent protein; CMV, cytomegalovirus.
activation activity of Rta. The interaction with Rb results in the release of E2F1 and cell-cycle pro-gression.9–11Moreover, Rta interacts with Sp1 via the Sp1-binding protein MCAF1. This interaction enhances the transcription of BRLF1, BNLF1, p16 and p21.12Rta is known to activate the transcription of BZLF1 via the ZII region in the promoter, which contains an ATF1/2 site.13 Since Rta does not bind directly to this region, Rta may indirectly influence the capacity of ATF1/2 to activate the transcription by inducing ATF2 phosphorylation.13 Rta has been demonstrated to interact with TSG101 to activate the expression of EBV late genes.14
RanBPM is a 90 kDa protein that interacts with Ran nuclear-cytoplasmic transport protein to affect pro-tein shuttling between the cytoplasm and the nucleus.15,16This protein contains an SPRY domain, which binds to the TrkA receptor,17 hDectin-1E,18 L119 and Axl/Sky receptor tyrosine kinases.20 Additionally, RanBPM interacts with many cellular receptors, including the androgen receptor, the gluco-corticoid receptor and the thyroid hormone recep-tor,21,22 enhancing their transcriptional activities. RanBPM interacts also with various signaling pro-teins, including the Met receptor, Mirk, p75NTR and integrin LFA-1, to regulate signaling pathways.23–25 By inhibiting the ubiquitination of p73α, RanBPM stabilizes p73α, and increases the proapototic activ-ity of the protein.26 This work demonstrates that RanBPM interacts with Rta to enhance the sumoyla-tion and transactivasumoyla-tion activity of Rta.
Results
Interaction between RanBPM and Rta in yeast The yeast two-hybrid screening assay was per-formed in yeast strain YRG2 with a bait plasmid, pR476,8,27which encodes a fusion protein that con-tains a Gal4 DNA-binding domain that is fused with the N-terminal 476 amino acids of Rta. After the screening of about 6 × 105colonies that were trans-formed with a human testes cDNA library, 56 plas-mids were found to encode a protein that interacts with RN476. One of these plasmids encoded RanBPM. The interaction was further verified in the strain YRG2 that was cotransformed with pR476 and the cDNA clone that encodes RanBPM.
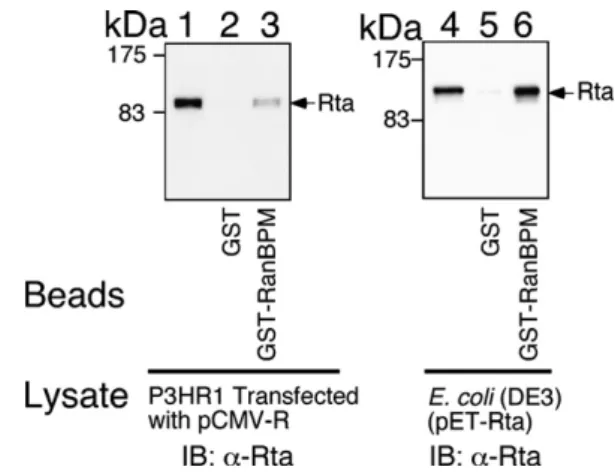
body. The results indicated that Rta in the lysate (Fig. 1, lane 1) was retained by GST-RanBPM- (Fig. 1, lane 3) but not GST-glutathione-Sepharose beads (Fig. 1, lane 2). A similar assay demonstrated that His-tagged Rta in the E. coli lysate (Fig. 1, lane 4) was retained also by GST-RanBPM- (Fig. 1, lane 6) but not by GST-glutathione-Sepharose beads (Fig. 1, lane 5), confirming that Rta interacts directly with RanBPM.
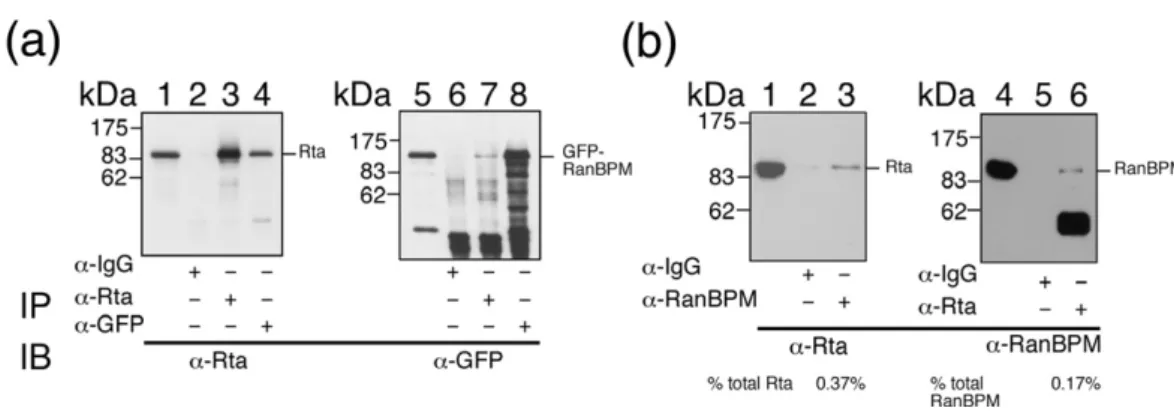
Interaction between Rta and RanBPMin vivo 293T cells were cotransfected with pCMV-R and pEGFP-RanBPM. Cell lysates were then prepared at 24 h after transfection. Immunoblot analysis re-vealed that Rta was present in the lysate (Fig. 2a, lane 1), immunoprecipitated by anti-Rta antibody (Fig. 2a, lane 3) and coimmunoprecipitated with green fluorescent protein (GFP)-RanBPM by anti-GFP antibody (Fig. 2a, lane 4). However, Rta was not immunoprecipitated by anti-IgG antibody (Fig. 2a, lane 2). Additionally, immunoblot analysis with anti-GFP antibody revealed the presence of GFP-RanBPM in the lysate (Fig. 2a, lane 5). Both anti-Rta
Fig. 1. Interaction between Rta and RanBPM in vitro. Lysate prepared from P3HR1 cells that had been trans-fected with pCMV-R (lanes 1–3) or from E. coli BL21(DE3) (pET-Rta) (lanes 4–6) was added to GST- (lanes 2 and 5) and GST-RanBPM- (lanes 3 and 6) glutathione-Sepharose beads. Lanes 1 and 4 were loaded with 5% of the indicated proteins in the cell lysate. Proteins on the beads were finally analyzed by immunoblotting (IB) with anti-Rta antibody (lanes 1–6).
antibody and anti-GFP antibody immunoprecipi-tated GFP-RanBPM (Fig. 2a, lanes 7 and 8). Yet, anti-IgG antibody did not immunoprecipitate GFP-RanBPM (Fig. 2a, lane 6). In addition, P3HR1 cells were treated with 12-O-tetradecanoylphobol-13-acetate (TPA) and sodium butyrate to induce the EBV lytic cycle. After culturing the cells for 24 h, a coimmunoprecipitation study revealed that Rta in the cell lysate was coimmunoprecipitated by anti-RanBPM antibody (Fig. 2b, lane 3); anti-RanBPM, by anti-Rta antibody (Fig. 2b, lane 6), indicating that endogenous Rta and RanBPM interact.
Mapping the Rta-RanBPM interaction region in Rta
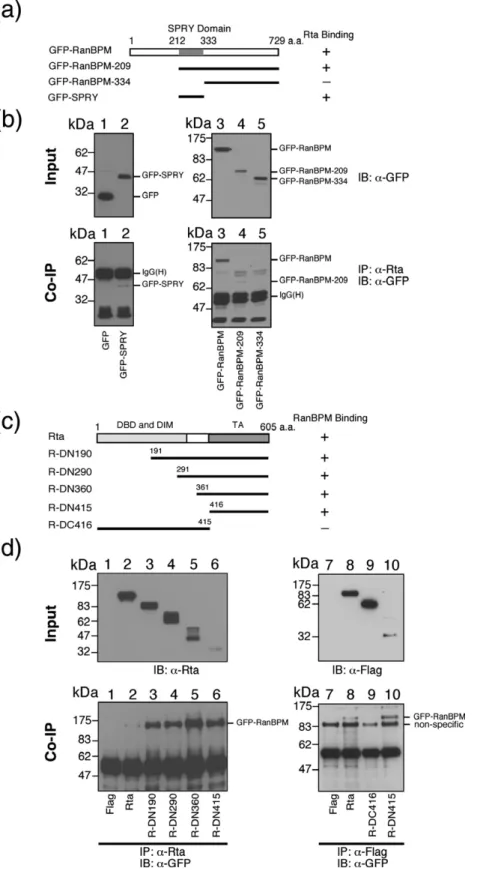
GFP-RanBPM fusion protein was deleted to deli-neate the regions in RanBPM that interact with Rta. Plasmids that encode GFP-RanBPM, GFP-RanBPM-209, GFP-RanBPM-334, and GFP-SPRY (Fig. 3a) were cotransfected with pCMV-R into 293T cells. An empty vector, pEGFP-C1, that expresses GFP was used as a negative control. Immunoblotting revealed that GFP-RanBPM and GFP-RanBPM-209 were present in the lysate and coimmunoprecipitated by anti-Rta antibody (Fig. 3b, lanes 3 and 4). GFP and GFP-RanBPM-334, although detectable in the lysate (Fig. 3b, lanes 1 and 5), were not coimmunoprecipi-tated by anti-Rta antibody (Fig. 3b, lanes 1 and 5), suggesting that the region between amino acid residues 209 and 333 in RanBPM, which contains an SPRY domain,28interacts with Rta. Additionally, this work found that a GFP-fusion protein that contains the SPRY domain could be coimmunopre-cipitated by anti-Rta antibody (Fig. 3b, lane 2), con-firming the interaction between the SPRY domain and Rta. A study was performed also with deleted Rta to define the region in Rta that binds to RanBPM (Fig. 3c). These Rta proteins were expressed in the
cells and were detected in the lysate by immuno-blotting with Rta (Fig. 3d, lanes 2–6) and anti-Flag antibody (Fig. 3d, lanes 7–10). GFP-RanBPM was coimmunoprecipitated with full-length Rta, R-DN190, R-DN290, R-DN360 and R-DN415 by Rta antibody and detected with GFP anti-body (Fig. 3d, lanes 1–6). GFP-RanBPM was coim-munoprecipitated with R-DN415, but not R-DC416, by anti-Flag antibody (Fig. 3d, lanes 7–10), indicating that the region in Rta that binds to RanBPM is located between amino acid residues 416 and 605. Since our yeast two-hybrid screening assay had estab-lished that the N-terminal 476 amino acids of Rta interact with RanBPM, the binding region of RanBPM to Rta is defined as the region between amino acids 416 and 476.
Colocalization of Rta and RanBPM in the nucleus
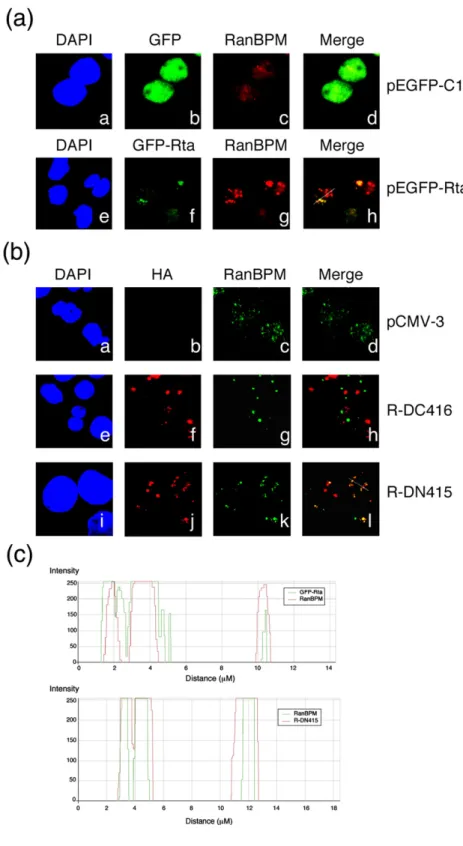
P3HR1 cells were transfected with pEGFP-Rta to study the subcellular location of RanBPM and Rta. A confocal microscopic study revealed that RanBPM was present in the nucleus in P3HR1 cells (Fig. 4a–c) and that transfection of pEGFP-Rta caused an accu-mulation of RanBPM in nuclear dots (Fig. 4a–g), where it colocalized with Rta (Fig. 4a–h). On the other hand, RanBPM did not colocalize with GFP (Fig. 4a–d). In addition, P3HR1 cells were transfected with pR-DC416 and pR-DN415 to examine the colocalization of deleted Rta and RanBPM. RanBPM was colocalized with R-DN415 (Fig. 4b–l), but not with R-DC416 (Fig. 4b–h), in the nucleus. In a negative control, colocalization between RanBPM and Rta was not observed in P3HR1 cells that were transfected with pCMV-3 (Fig. 4b–d). Quantitative analysis of the merged confocal images with Zeiss LSM Image Browser software confirmed the coloca-lization of Rta and RanBPM (Fig. 4c).
Fig. 2. Interaction between Rta and RanBPM in vivo. (a) Anti-Rta and anti-GFP antibody were added to the lysates from 293T cells that were transfected with pCMV-R and pEGFP-RanBPM. Lanes 1 and 5 were loaded with 5% of the cell lysate. Proteins immunoprecipitated (IP) with Rta antibody (lanes 3 and 7), GFP antibody (lanes 4 and 8) or anti-IgG (lanes 2 and 6), were detected by immunoblotting (IB) with anti-Rta antibody (lanes 1–4) and anti-GFP antibody (lanes 5–8). (b) P3HR1 cells were treated with TPA and sodium butyrate for 24 h to induce the EBV lytic cycle. Lanes 1 and 5 were loaded with 3% of the protein in the cell lysate. Proteins in the cell lysate were immunoprecipitated with RanBPM (lane 3), Rta (lane 6) or IgG (lanes 2 and 5) antibodies. Immunoblot analysis was performed with anti-Rta (lanes 1–3) and anti-RanBPM (lanes 4–6) antibodies. The percentages of anti-Rta and RanBPM in the cell lysate that were immunoprecipitated were determined using Image J software (Wayne Rasband, National Institutes of Health, USA) to quantify the proteins in a cell extract coimmunoprecipitated with Rta or RanBPM.
Effect of RanBPM on Rta sumoylation
Since proteins that interact with RanBPM are fre-quently conjugated by SUMO,29–33we investigated whether the interaction between RanBPM and Rta affects the sumoylation of Rta. P3HR1 cells were transfected with pCMV-R, pCR-SUMO1, pEGFP-RanBPM, pEF-PIASxβ, and pcDNA-Ubc9. In input
controls, the level ofα-tubulin, Rta, GFP-RanBPM, PIASxβ and HA-Ubc9 expression was determined by immunoblotting with anti-α-tubulin, anti-Rta, anti-RanBPM and anti-HA antibodies, respectively (Fig. 5a, lanes 1–7). After SUMO-conjugated pro-teins were immunoprecipitated with Flag anti-body, bands of sumoylated Rta were detected by immunoblotting with anti-Rta antibody. Previous
Fig. 3. Mapping the interaction domains of RanBPM and Rta. Plasmids that express deleted GFP-RanBPM (a) including pEGFP-RanBPM (lane 3), pEGFP- pEGFP-RanBPM-209 (lane 4), pEGFP-RanBPM-334 (lane 5), pEGFP-SPRY (lane 2) or pEGFP-C1 (lane 1) were cotrans-fected with pCMV-R into 293T cells. The input, representing 5% of GFP-fusion proteins in the cell lysates, were detected by anti-GFP antibody. Proteins in the cell lysate were coimmunoprecipitated (IP) with anti-Rta antibody and analyzed by immunoblotting (IB) using anti-GFP antibody. Samples in lanes 1, 2 and 3–5 were separated by SDS-PAGE with 9% and 10% polyacryla-mide gel, respectively. (b), Deletion mutants of Rta were used to analyze the region of Rta that interacts with RanBPM (c). 293T cells were cotrans-fected with pEGFP-RanBPM and Rta (lanes 2 and 8), R-DN190 (lane 3), R-DN290 (lane 4), R-DN360 (lane 5), R-DN415 (lanes 6 and10) and R-DN416 (lane 9). Lysate was then prepared and 5% of the indicated protein was loaded as an input con-trol and detected by immunoblotting using anti-Rta and anti-Flag antibo-dies (d). Proteins in the lysate were then immunoprecipitated with anti-Rta or anti-Flag antibodies and de-tected by immunoblotting with anti-GFP antibody (d). Plasmid pTag2A, an empty vector, was used as a ne-gative control (lanes 1 and 7). IgG (H), the heavy chain of IgG; DBD, DNA-binding domain; DIM, dimer-ization domain; TA, transactivation domain.
studies showed that the detectable level of SUMO-1 conjugated proteins increased in the presence of 5– 10 μM of MG132.33–35 Under the conditions, the amount of sumoylated Rta becomes abundant and further enhancement of Rta sumoylation by SUMO E3 ligases was difficult to detect. Therefore, in this study, we decreased the concentration of MG132 to 0.5 μM. Under the experimental conditions, Rta sumoylation was undetected after cotransfecting pCR-SUMO-1 and pCMV-R into P3HR1 cells (Fig. 5a, lane 2). However, over-expression of
GFP-RanBPM increases Rta sumoylation, as shown by the appearance of a sumoylated Rta band by immu-noblotting (Fig. 5a, lane 3). Furthermore, over-expressing Ubc9 or PIASxβ did not cause high levels of Rta sumoylation (Fig. 5a, lanes 4 and 5), which were instead seen when both proteins were over-expressed together (Fig. 5a, lane 7). Similarly, co-expressing Ubc9 and GFP-RanBPM markedly increased the amount of sumoylated Rta (Fig. 5a, lane 6); the level of Rta sumoylation appeared to be higher than that induced by Ubc9 and PIASxβ (Fig.
Fig. 4. Indirect immunofluores-cence analysis. P3HR1 cells were transfected with an empty vector (pEGFP-C1) (a, a–d), pEGFP-Rta (a, e–h), pHA-R-DC416 (b, e–h), pHA-R-DN415 (b, i–l), or pCMV-3 (b, a–d). Cells were incubated with monoclonal anti-RanBPM antibody (a, c and g; b, c, g, and k) or poly-clonal anti-HA antibody (b, b, f and j) and observed under a confocal laser-scanning microscope. Staining with 4′-6-diamidino-2-phenylindole re-vealed the positions of the nucleus. (d, h and l) are merged images. Yellow color in the merged images was analyzed by the generation of a white line scan to quantify the fluorescence intensity with ZEISS LSM Image Browser software (c).
5a, lane 7). In addition, P3HR1 cells were cotrans-fected with pCR-SUMO-1, pCMV-R, and RanBPM deletion mutants including pEGFP-RanBPM209, pEGFP-RanBPM-334, or pEGFP-SPRY. In input con-trol, the expression ofα-tubulin, Rta and GFP-fusion proteins were detected with anti-α-tubulin, anti-Rta and anti-GFP antibodies, respectively (Fig. 5b). SUMO-1 conjugated Rta was detected by immuno-precipitation with anti-Flag antibody in the presence of 0.5μM MG132. Over-expression of GFP-RanBPM-209 and GFP-SPRY resulted in the enhancement of Rta sumoylation (Fig. 5b, lanes 2 and 4); whereas over-expressing GFP-RanBPM-334 did not cause increased SUMO-1 conjugation to Rta (Fig. 5b, lane 5), further confirming that the SPRY domain of RanBPM is necessary to promote Rta sumoylation. Interaction between RanBPM and Ubc9
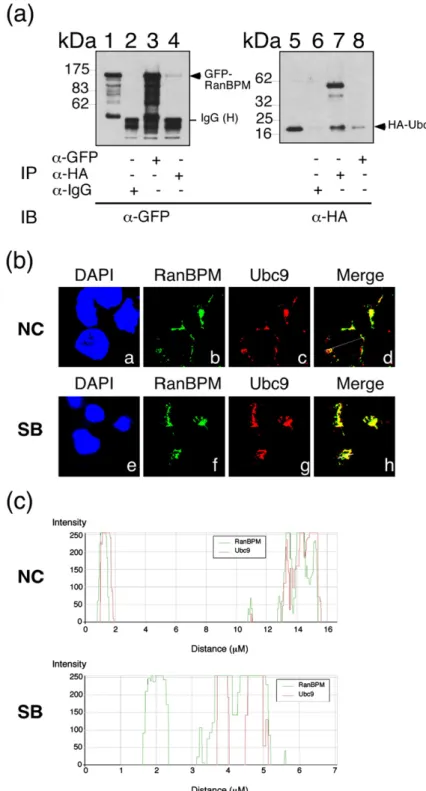
We further investigated whether RanBPM inter-acts with Ubc9 to elucidate further the involvement of RanBPM in protein sumoylation. A lysate from 293T cells that were cotransfected with pEGFP-RanBPM and pcDNA-Ubc9 was prepared 24 h after transfection (Fig. 6a). The presence of GFP-RanBPM in the lysate was confirmed by immunoblot analysis with anti-GFP antibody (Fig. 6a, lane 1). GFP-RanBPM was immunoprecipitated by anti-GFP antibody (Fig. 6a, lane 3) and coimmunoprecipitated with HA-Ubc9 by anti-HA antibody (Fig. 6a, lane 4). GFP-RanBPM was undetected when IgG
anti-body was added to the immunoprecipitation reac-tion (Fig. 6a, lane 2). HA-Ubc9 was also immuno-precipitated by the anti-HA antibody (Fig. 6a, lane 7) and coimmunoprecipitated with GFP-RanBPM by anti-GFP antibody (Fig. 6a, lane 8). However, HA-Ubc9 was not immunoprecipitated by anti-IgG antibody (Fig. 6a, lane 6). In addition, P3HR1 cells untreated (Fig. 6b, a–d) or treated with sodium butyrate (Fig. 6b, e–h) to induce the EBV lytic cycle were used to examine the subcellular location of RanBPM and Ubc9. A confocal microscopy study showed that RanBPM colocalizes with Ubc9 in the nucleus both in the absence (Fig. 6b–d) and in the presence of Rta (Fig. 6b–h). Quantitative analysis of the merged confocal images with Zeiss LSM Image Browser software confirmed the colocalization of RanBPM and Ubc9 (Fig. 6c).
Enhancement of transactivation activity of Rta by RanBPM
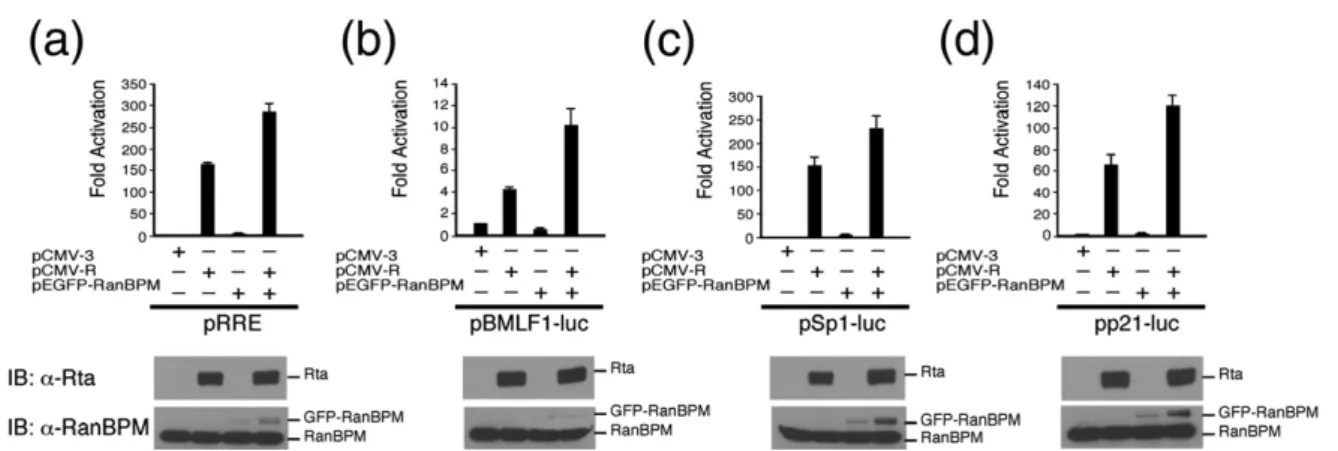
Rta is known to bind to RRE in a promoter to activate transcription or interact with MCAF1 to enhance Sp1-mediated transcription.12 This study performed transient transfection to determine whether the interaction between Rta and RanBPM affects these two types of transcription. As expected, in EBV-negative Akata cells, transfection of pCMV-R activated luc transcription from pRRE and pBMLF1-luc, two reporter plasmids that contain a promoter with an RRE sequence, up to 158-fold and 4.3-fold,
Fig. 5. Enhancement of Rta sumoylation by RanBPM. P3HR1 cells were transfected with pCR-SUMO-1, pCMV-R, pEF-PIASxβ or pcDNA-Ubc9 and treated with 0.5 μM MG132. Proteins that were conjugated by Flag-tagged SUMO-1 were immunoprecipitated (IP) with anti-Flag antibody and detected by immunoblotting (IB) using anti-Rta antibody. The input control was loaded with 2.5% of the cell lysates and proteins were detected by immunoblotting with anti-α-tubulin, anti-Rta, anti-RanBPM, and anti-HA antibodies (a). In addition, P3HR1 cells were transfected with pCR-SUMO-1, pCMV-R, pEGFP-RanBPM-209, pEGFP-RanBPM-334 or pEGFP-SPRYand treated with 0.5μM of MG132. Similarly, proteins that were conjugated by Flag-tagged SUMO-1 were immunoprecipitated (IP) with anti-Flag antibody and detected by immunoblotting (IB) using anti-Rta antibody. The input control was loaded with 2.5% of the cell lysates and proteins were detected by immunoblotting with anti-α-tubulin, anti-Rta and anti-GFP antibodies (b). PSR, polysumoylated Rta.
respectively (Fig. 7a and b). The promoter activity increased by 288-fold and 10.4-fold, respectively (Fig. 7a and b) when the cells were cotransfected with pEGFP-RanBPM and pCMV-R. In the absence of pCMV-R, pEGFP-RanBPM did not activate the transcription (Fig. 7a and b), demonstrating that RanBPM promotes the transactivation activity of Rta. To determine whether RanBPM affected Sp1-mediated transcription, similar experiments were performed using two reporter plasmids that contain a promoter with Sp1 binding sites, pSp1-luc and pp21-luc. As expected, transfecting pCMV-R acti-vated pSp1-luc and pp21-luc by 150-fold and 65-fold,
respectively (Fig. 7c and d); the luciferase activity increased by 234-fold and 121-fold, respectively (Fig. 7c and d), when the cells were cotransfected with pEGFP-RanBPM. Additionally, plasmid pLG5, a reporter plasmid that contains a promoter with five copies of Gal4-binding sites, was utilized to deter-mine how Ubc9, SUMO-1 and RanBPM influence the capacity of Rta to transactivate the promoter. In EBV-negative Akata cells, cotransfecting pLG5 and pM-Rta, which expressed Gal4-pM-Rta, activated the Gal4 promoter (Fig. 8). In the presence of Ubc9, SUMO-1 and RanBPM, the capacity of Gal4-Rta to activate the promoter increased 3.1-fold, 2.5-fold and 3.3-fold,
Fig. 6. Interaction between Ran– BPM and SUMO E2 (Ubc9). (a) 293T cells were cotransfected with pEGFP-RanBPM and pcDNA-Ubc9. Coimmunoprecipitation was per-formed with anti-GFP antibody (lanes 3 and 8) and HA anti-body (lanes 4 and 7). Immunopre-cipitation (IP) was also performed using anti-IgG antibody (lanes 2 and 6). Immunoblotting (IB) was per-formed with anti-GFP (lanes 1–4) and anti-HA antibodies (lanes 5–8). Lanes 1 and 5 were loaded with 5% of the cell lysate. IgG (H), the heavy chain of IgG. (b) In addition, P3HR1 cells were treated with sodium butyrate (SB) to induce EBV lytic cycle. Cells were incubated with monoclonal anti-RanBPM antibody (b and f) and polyclonal anti-Ubc9 antibody (c and g), then observed under a confocal laser scanning microscope. Staining with 4′-6-dia-midino-2-phenylindole revealed the positions of the nucleus. d and h are merged images. Yellow color in the merged images was analyzed by the generation of a white line scan to quantify the fluorescence intensity according to ZEISS LSM Image Browser software (c). NC, P3HR1 cells untreated with SB.
respectively (Fig. 8). The over-expression of two of these three proteins increased the activity of Gal4-Rta by factors of 4.8–6.3 (Fig. 8). The Gal4-Rta activity increased by a factor of 7.5 when Ubc9, SUMO-1 and RanBPM were over-expressed in the cells (Fig. 8), indicating that RanBPM promotes the transactivation activity of Rta by sumoylation.
Activation of the EBV lytic cycle
It is generally known that Rta is usually not ex-pressed during EBV latency in P3HR1 cells. Trans-fecting up to 20μg of pEGFP-RanBPM into 5×106of P3HR1 cells had little effect on the expression of Rta; immunoblot analysis with anti-Rta antibody did not detect the expression of Rta at 24 h after transfection (Fig. 9, lane 3). Transfecting the cells with 0.5μg of pCMV-R, which is about 10% of the amount that is
typically used in EBV lytic induction, resulted in the synthesis of a small amount of Rta (Fig. 9, lane 4). However, the amount of Rta increased markedly when 5 μg or 10 μg of pEGFP-RanBPM was also cotransfected (Fig. 9, lanes 5 and 6), indicating that RanBPM promoted the expression of Rta, presum-ably by autoregulating the BRLF1 promoter. Addi-tionally, the transfection also led to the expression of both phosphorylated and unphosphorylated EA-D (Fig. 9), an early protein of EBV, suggesting that RanBPM affects the lytic development.
Discussion
Rta and Zta are expressed by EBV during the immediate-early stage of the lytic cycle.36,37 These two proteins appear to activate EBV genes in diffe-rent ways. Transcription of many EBV lytic genes, including those expressed during the immediate-early and immediate-early stage of the lytic cycle, seems to depend strongly on Zta. Without Zta, none of these genes is expressed,38–41 indicating that Rta alone
Fig. 7. RanBPM enhances the transactivation activity of Rta in EBV-negative Akata cells. Four reporter plasmids — pRRE (a), pBMLF1-luc (b), pSp1-luc (c) and pp21-luc (d)— were cotransfected with pCMV-3, pCMV-R or pEGFP-RanBPM into EBV-negative Akata cells. Luciferase activity that was exhibited by the cells was examined at 24 h after transfection. Each transfection experiment was performed three times and each sample in each experiment was prepared in duplicate. The level of Rta and RanBPM expression were detected by immunoblotting (IB) with anti-Rta and anti-RanBPM antibodies.
Fig. 8. Enhancement of Rta's transactivation activity by sumoylation enzymes and RanBPM. EBV-negative Akata cells were cotransfected with pcDNA-Ubc9, pCR-SUMO-1, pEGFP-RanBPM, or combinations of these plasmids, to elucidate the effect of sumoylation on the capacity of Gal4-Rta to transactivate the pLG5 promoter. Luciferase activity that was exhibited by the cells was examined at 24 h after transfection. Each transfection experiment was performed at least three times, and each sample in each experiment was prepared in duplicate. RLU: relative light units.
Fig. 9. Effects of RanBPM on EBV lytic activation. P3HR1 cells were treated with 3 mM sodium butyrate to activate the expression of Rta (lane 1). Cells untreated with sodium bu-tyrate (lane 2) were transfected with 20 μg of pEGFP-RanBPM (lane 3). Cotransfection was also performed by transfecting 0.5 μg of pCMV-R and 5–20 μg of pEGFP-RanBPM (lanes 4–7). Lysates were prepared at 24 h after transfection. Immunoblot analysis was conducted using anti-Rta, add 'anti-EA-D, anti-GFP and anti-α-tubulin antibodies.
nuclear-cytoplasmic transport protein, also interacts with Rta. The interaction is verified in vitro by GST-pull-down analysis (Fig. 1) and in vivo by coimmu-noprecipitation (Fig. 2). This interaction apparently involves a segment of the transactivation domain of Rta, spanning amino acid residues 416–476, and the SPRY domain in RanBPM (Fig. 3). The interaction with RanBPM appears to promote the transactiva-tion activity of Rta in transient transfectransactiva-tion analysis (Fig. 7). In fact, RanBPM is known to interact, via its SPRY domain, with many proteins, including TrkA receptor, hDectin-1E, L1,Axl/Sky and hormone re-ceptors,17–20and to affect their biological functions. Since many RanBPM-interacting proteins, including androgen receptor, glucocorticoid receptor, p73 and HIPK2, are conjugated by SUMO, this work investigates whether RanBPM participates in Rta sumoylation. Our previous study demonstrated that Rta is conjugated by SUMO-1 to promote the transactivation activities of Rta.8 The work also found that sumoylated Rta was unstable and sen-sitive to SUMO proteases, causing the detection of sumoylated Rta by immunoblotting difficult.8 Therefore, treating cells with 2.5 μM MG132 is necessary to stabilize and detect sumoylated Rta by immunoblotting. However, under such experimen-tal conditions, the amount of sumoylated Rta becomes abundant and further enhancement of Rta sumoylation by SUMO E3 ligases was difficult to demonstrate. Therefore, this study reduces the con-centration of MG132 to 0.5 μM to decrease the amount of detectable sumoylated Rta so that an increase of Rta sumoylation can be detected after over-expressing SUMO E3 ligase. Under the condi-tions, sumoylated Rta in P3HR1 cells was undetected after cotransfecting pCR-SUMO-1 and pCMV-R (Fig. 5a, lane 2 and b, lane 3). However, over-expression of GFP-RanBPM increases Rta sumoylation (Fig. 5a, lane 3). Previous studies showed that sumoylation efficiency by SUMO E3 ligases is enhanced in the presence of SUMO E2 (Ubc9).42–44In fact, this study find that RanBPM interacts with Ubc9 in vivo (Fig. 6) and over-expression of GFP-RanBPM and Ubc9 profoundly increases the number of sumoylated Rta bands and their intensity in a gel (Fig. 5, lane 6), indicating that RanBPM functions as a SUMO E3 ligase in enhancing the sumoylation of Rta. However, we were unable to demonstrate the same enhance-ment effect in vitro (data not shown). The lack of enhancement may be attributable to the fact that RanBPM expressed by E. coli is not functional and is
ligases, including PIAS1, PIASxα and PIASxβ, inhibiting RanBPM expression alone may not be sufficient to inhibit Rta sumoylation. Since sumoyla-tion is known to enhance the transactivasumoyla-tion activity of Rta,8,27 the fact that RanBPM over-expression promotes Rta sumoylation could explain why the expression of GFP-RanBPM also increases the trans-activation activity of Rta (Fig. 7). Additionally, the fact that RanBPM promotes the capacity of Gal4-Rta to transactivate the Gal4 promoter in pLG5 (Fig. 8) further indicates that RanBPM influences Rta's transactivation activity rather than its DNA-binding capacity. The enhanced Rta transactivation activity is less likely due to stabilization of Rta by sumoylation, since Rta levels were not increased by RanBPM over-expression (Fig. 7), although further studies will be necessary to completely rule out that endogeneous Rta becomes more stable in the presence of RanBPM. Most importantly, over-expression of RanBPM and the resulting enhanced Rta activity seem to influence the activation of the EBV lytic cycle by Rta. This study demonstrates that transfection of 0.5 μg of pCMV-R, an amount tenfold lower than that typically used for EBV lytic activation, induces production of only a small amount of Rta and EA-D. However, cotransfection of pEGFP-RanBPM substantially increases the expression of these two proteins in a dose-dependent manner in P3HR1 cells (Fig. 9). These results clearly indicate the participa-tion of RanBPM in promoting the effect of Rta on EBV lytic activation by Rta.
Materials and Methods
Cell lines and EBV lytic induction
P3HR1 and EBV-negative Akata cells were cultured in RPMI 1640 medium containing 10% (v/v) fetal calf serum. 293T cells were cultured in Dulbecco's modified Eagle's medium (DMEM) that contained 10% fetal calf serum. To activate the EBV lytic cycle, P3HR1 cells were treated with TPA and sodium butyrate, according to a method des-cribed elsewhere.47,48
Plasmids
Plasmid pCMV-R contains BRLF1 that was trans-cribed from the cytomegalovirus (CMV) immediate-early promoter. Plasmid pCMV-3 is a cloning vector that was used to construct pCMV-R. Plasmid pGEX-4T1, which ex-presses GST, was purchased from Amersham Biosciences.
5′-CCGCTCGAGTAATGTAGGTAGTCTTCC
into the HindIII and SalI sites in pEGFP-C1. Plasmid pEGFP-RanBPM-334 was constructed using the same strategy with primers RanBPM-1055:
5′-CCCAAGCTTGTCCTTTCGTGTTTGATATAG and RanBPM-3. Plasmid pEGFP-SPRY was constructed in the same way with the DNA fragment that was amplified with primers RanBPM-684 and GST-BPM:
5′-CCGCTCGAGCACGAAAGGATGTTGCCC Plasmid pFlag-Rta that expresses full-length Rta with a Flag tag at the amino terminus (Flag-Rta) was constructed by inserting a PCR-amplified BRLF1 fragment that was amplified with primers Flag-5R:
5′-CCGGAATTCCAGGCCTAAAAAGGATGGC and GST-3R:
5′-ACGCGTCGACGTCTAAAATAAGCTGGTGTCAA into pCMV-Tag2A at the EcoRI and XhoI sites. Plasmids that expressed deletion mutants of Flag-Rta, including R-DN190, R-DN290, R-DN360, R-DN415 and R-DC416, which lacked Rta regions from amino acids 1–190, 1–290, 1–360, 1–415 and 416–605, respectively, were constructed by inserting BRLF1 fragments that had been amplified by PCR, into the EcoRI-XhoI sites in pCMV-Tag2A. Plasmids that expressed deletion mutants of HA-Rta, including R-DC416 and R-DN415, which lack Rta regions from amino acids 416–605 and 1–415, respectively, were constructed by inserting a PCR-amplified BRLF1 fragment into the EcoRI-XhoI sites in pcDNA3-HA. Plasmids pRRE, pBMLF1-luc, pSp1-luc and pp21-luc were reporter plas-mids that were used to analyze the transactivation activation activity of Rta.27Plasmid pLG5 has five
Gal4-binding sites and a TATA sequence upstream of a luciferase gene. Plasmid pM is an empty vector that contains the GAL4 gene (Clontech). A DNA fragment that encodes an Rta region from amino acids 255–605 was amplified by PCR and inserted into the EcoRI and SalI sites in pM to generate pM-R. Plasmids pCR-SUMO-1 and pcDNA-Ubc9 expressed Flag-tagged SUMO-1 and HA-tagged Ubc9, respectively.8
Yeast two-hybrid screen
Yeast two-hybrid screening of the binding partners of Rta was performed with a bait plasmid, pR476 as described.8,27
were induced with TPA and sodium butyrate for 24 h and cell lysate was mixed with Rta or RanBPM anti-body.24Proteins that had been immunoprecipitated by the antibodies were captured with protein A/G-agarose beads (Oncogene, Boston, MA). Immunoblot analysis was performed to detect immunoprecipitated proteins. Sumoylated Rta was detected as described.51
Immunoblot analysis
Proteins that were separated by SDS-PAGE were electro-transferred to Hybond C membrane (Amersham Bio-sciences, New Jersey) at 90 V for 1 h, and probed using appropriate antibodies. Supersignal West Pico chemilumi-nescent substrate (Pierce, Rockford, IL) was used to visua-lize the proteins on the membrane. The antibodies used included anti-Rta, anti-HA, anti-GFP, anti-α-tubulin (Sigma, St Louis, MO) and anti-Flag antibodies (Sigma). Anti-RanBPM monoclonal antibody is described elsewhere.24
Transient transfection assay
P3HR1, EBV-negative Akata and 293T cells (5 × 106) were transfected with 5μg of plasmids by electroporation using a BTX ECM630 electroporator (BTX Instrument, Holliston, MA) and luciferase assay was performed using a luminometer (Orion II; Berthod, Bad Wildbad, Ger-many) as described.50 Each transfection experiment was performed three times and each sample in each experi-ment was prepared in duplicate.
Immunofluorescence analysis
P3HR1 cells were cultured for 24 h after transfecting pEGFP, pEGFP-Rta, pCMV-3, DC416, or pHA-R-DN415. Cells were harvested by centrifugation; plated on poly-l-lysine (Sigma)-coated coverslips, and fixed with 4% (v/v) paraformaldehyde in phosphate-buffered saline (PBS) for 30 min. The cells were then incubated for 1 h with anti-RanBPM monoclonal antibody, rabbit anti-HA polyclonal antibody (Y-11, Santa Cruz Biotechnology Inc., Santa Cruz, CA), or goat anti-Ubc9 polyclonal antibody (N-15, Santa Cruz Biotechnology Inc.). Following incuba-tion, cells were washed three times in PBS and then treated with fluorescein Alexa 488-conjugated goat anti-mouse IgG antibodies (Molecular Probes, Inc. Eugene, OR), Alexa 598-conjugated goat anti-rabbit IgG antibodies (Molecular Probes), or rhodamine-conjugated donkey anti-goat IgG antibodies. Nuclei were visualized by staining with 5μg/ ml of 4′-6-diamidino-2-phenylindole. Finally, cells were washed in PBS, mounted in CITIFLOUR (Agar Inc. Essex, England), and observed with a Zeiss confocal laser scanning microscope (META, model LSM 510).
References
1. Kieff, E. (1996). Epstein-Barr virus and its replication. In Fields' Virology (Fields, B. N., Knipe, D. M. & Howley, P. M., eds), 3rd edit. Lippincott-Raven, Philadelphia, PA.
2. Speck, S. H., Chatila, T. & Flemington, E. (1997). Reac-tivation of Epstein-Barr virus: regulation and function of the BZLF1 gene. Trends Microbiol.5, 399–405. 3. Liu, P. & Speck, S. H. (2003). Synergistic autoactivation
of the Epstein-Barr virus immediate-early BRLF1 promoter by Rta and Zta. Virology,310, 199–206. 4. Holley-Guthrie, E. A., Quinlivan, E. B., Mar, E. C. &
Kenney, S. (1990). The Epstein-Barr virus (EBV) BMRF1 promoter for early antigen (EA-D) is regulated by the EBV transactivators, BRLF1 and BZLF1, in a cell-specific manner. J. Virol.64, 3753–3759.
5. Gruffat, H., Duran, N., Buisson, M., Wild, F., Buckland, R. & Sergeant, A. (1992). Characterization of an R-binding site mediating the R-induced activation of the Epstein-Barr virus BMLF1 promoter. J. Virol.66, 46–52. 6. Feederle, R., Kost, M., Baumann, M., Janz, A., Drouet, E., Hammerschmidt, W. & Delecluse, H. J. (2000). The Epstein-Barr virus lytic program is controlled by the co-operative functions of two transactivators. EMBO J. 19, 3080–3089.
7. Gruffat, H., Manet, E., Rigolet, A. & Sergeant, A. (1990). The enhancer factor R of Epstein-Barr virus (EBV) is a sequence-specific DNA binding protein. Nucleic Acids Res.18, 6835–6843.
8. Chang, L. K., Lee, Y. H., Cheng, T. S., Hong, Y. R., Lu, P. J., Wang, J. J. et al. (2004). Post-translational modi-fication of Rta of Epstein-Barr virus by SUMO-1. J. Biol. Chem.279, 38803–38812.
9. Swenson, J. J., Holley-Guthrie, E. & Kenney, S. C. (2001). Epstein-Barr virus immediate-early protein BRLF1 interacts with CBP, promoting enhanced BRLF1 trans-activation. J. Virol.75, 6228–6234.
10. Zacny, V. L., Wilson, J. & Pagano, J. S. (1998). The Epstein-Barr virus immediate-early gene product, BRLF1, interacts with the retinoblastoma protein during the viral lytic cycle. J. Virol.72, 8043–8051. 11. Swenson, J. J., Mauser, A. E., Kaufmann, W. K. &
Kenney, S. C. (1999). The Epstein-Barr virus protein BRLF1 activates S phase entry through E2F1 induc-tion. J. Virol.73, 6540–6550.
12. Chang, L. K., Chung, J. Y., Hong, Y. R., Ichimura, T., Nakao, M. & Liu, S. T. (2005). Activation of Sp1-mediated transcription by Rta of Epstein-Barr virus via an interaction with MCAF1. Nucleic Acids Res.33, 6528–6539.
13. Adamson, A. L., Darr, D., Holley-Guthrie, E., Johnson, R. A., Mauser, A., Swenson, J. & Kenney, S. (2000). Epstein-Barr virus immediate-early proteins BZLF1 and BRLF1 activate the ATF2 transcription factor by
cDNA encodes a protein possessing a long stretch of proline and glutamine within the N-terminal region, comprising a large protein complex. Gene,272, 25–33. 17. Yuan, Y., Fu, C., Chen, H., Wang, X., Deng, W. & Huang, B. R. (2006). The Ran binding protein RanBPM interacts with TrkA receptor. Neurosci. Lett.407, 26–31. 18. Xie, J., Sun, M., Guo, L., Liu, W., Jiang, J., Chen, X. et al. (2006). Human Dectin-1 isoform E is a cytoplasmic protein and interacts with RanBPM. Biochem. Biophys. Res. Commun.347, 1067–1073.
19. Cheng, L., Lemmon, S. & Lemmon, V. (2005). RanBPM is an interacting protein that regulates L1-mediated mitogen-activated protein kinase activation. J. Neurochem.94, 1102–1110.
20. Hafizi, S., Gustafsson, A., Stenhoff, J. & Dahlback, B. (2005). The Ran binding protein RanBPM interacts with Axl and Sky receptor tyrosine kinases. Int. J. Biochem. Cell. Biol.37, 2344–2356.
21. Rao, M. A., Cheng, H., Quayle, A. N., Nishitani, H., Nelson, C. C. & Rennie, P. S. (2002). RanBPM, a nuclear protein that interacts with and regulates transcriptional activity of androgen receptor and glucocorticoid receptor. J. Biol. Chem.277, 48020–48027.
22. Poirier, M. B., Laflamme, L. & Langlois, M. F. (2006). Identification and characterization of RanBPM, a no-vel coactivator of thyroid hormone receptors. J. Mol. Endocrinol.36, 313–325.
23. Wang, D., Li, Z., Messing, E. M. & Wu, G. (2002). Activ-ation of Ras/Erk pathway by a novel MET-interacting protein RanBPM. J. Biol. Chem.277, 36216–36222. 24. Denti, S., Sirri, A., Cheli, A., Rogge, L., Innamorati, G.,
Putignano, S. et al. (2004). RanBPM is a phosphopro-tein that associates with the plasma membrane and interacts with the integrin LFA-1. J. Biol. Chem.279, 13027–13034.
25. Bai, D., Chen, H. & Huang, B. R. (2003). RanBPM is a novel binding protein for p75NTR. Biochem. Biophys. Res. Commun.309, 552–557.
26. Kramer, S., Ozaki, T., Miyazaki, K., Kato, C., Hanamoto, T. & Nakagawara, A. (2005). Protein stability and function of p73 are modulated by a physical interaction with RanBPM in mammalian cultured cells. Oncogene,24, 938–944.
27. Liu, S. T., Wang, W. H., Hong, Y. R., Chuang, J. Y., Lu, P. J. & Chang, L. K. (2006). Sumoylation of Rta of Epstein-Barr virus is preferentially enhanced by PIASxbeta. Virus Res.119, 163–170.
28. Ponting, C., Schultz, J. & Bork, P. (1997). SPRY domains in ryanodine receptors (Ca(2+)-release channels). Trends Biochem. Sci.22, 193–194.
29. Poukka, H., Karvonen, U., Janne, O. A. & Palvimo, J. J. (2000). Covalent modification of the androgen recep-tor by small ubiquitin-like modifier 1 (SUMO-1). Proc. Natl Acad. Sci. USA,97, 14145–14150.
30. Le Drean, Y., Mincheneau, N., Le Goff, P. & Michel, D. (2002). Potentiation of glucocorticoid receptor
trans-tion motif. J. Biol. Chem.275, 36316–36323.
34. Bailey, D. & O'Hare, P. (2005). Comparison of the SUMO1 and ubiquitin conjugation pathways during the inhibition of proteasome activity with evidence of SUMO1 recycling. Biochem. J.392, 271–281.
35. Roukens, M. G., Alloul-Ramdhani, M., Vertegaal, A. C., Anvarian, Z., Balog, C. I., Deelder, A. M. et al. (2008). Identification of a new site of sumoylation on Tel (ETV6) uncovers a PIAS-dependent mode of regulating Tel function. Mol. Cell. Biol.28, 2342–2357.
36. Biggin, M., Bodescot, M., Perricaudet, M. & Farrell, P. (1987). Epstein-Barr virus gene expression in P3HR1-superinfected Raji cells. J. Virol.61, 3120–3132. 37. Takada, K. & Ono, Y. (1989). Synchronous and
sequential activation of latently infected Epstein-Barr virus genomes. J. Virol.63, 445–449.
38. Buisson, M., Manet, E., Trescol-Biemont, M. C., Gruffat, H., Durand, B. & Sergeant, A. (1989). The Epstein-Barr virus (EBV) early protein EB2 is a posttranscriptional activator expressed under the control of EBV transcrip-tion factors EB1 and R. J. Virol.63, 5276–5284.
39. Cox, M. A., Leahy, J. & Hardwick, J. M. (1990). An enhancer within the divergent promoter of Epstein-Barr virus responds synergistically to the R and Z transactivators. J. Virol.64, 313–321.
40. Kenney, S., Holley-Guthrie, E., Mar, E. C. & Smith, M. (1989). The Epstein-Barr virus BMLF1 promoter contains an enhancer element that is responsive to the BZLF1 and BRLF1 transactivators. J. Virol.63, 3878–3883.
Mdm2 binding and SUMO-1 conjugation by the E2 SUMO-1 conjugating enzyme Ubc9. J. Biol. Chem.276, 40389–40395.
45. Woods, Y. L., Xirodimas, D. P., Prescott, A. R., Sparks, A., Lane, D. P. & Saville, M. K. (2004). p14 Arf promotes small ubiquitin-like modifier conjugation of Werners helicase. J. Biol. Chem.279, 50157–50166. 46. Chen, L. & Chen, J. (2003). MDM2-ARF complex
regulates p53 sumoylation. Oncogene,22, 5348–5357. 47. Luka, J., Kallin, B. & Klein, G. (1979). Induction of the
Epstein-Barr virus (EBV) cycle in latently infected cells by n-butyrate. Virology,94, 228–231.
48. Davies, A. H., Grand, R. J., Evans, F. J. & Rickinson, A. B. (1991). Induction of Epstein-Barr virus lytic cycle by tumor-promoting and non-tumor-promoting phor-bol esters requires active protein kinase C. J. Virol.65, 6838–6844.
49. Hsu, T. Y., Chang, Y., Wang, P. W., Liu, M. Y., Chen, M. R., Chen, J. Y. & Tsai, C. H. (2005). Reactivation of Epstein-Barr virus can be triggered by an Rta protein mutated at the nuclear localization signal. J. Gen. Virol. 86, 317–322.
50. Chang, P. J., Chang, Y. S. & Liu, S. T. (1998). Role of Rta in the translation of bicistronic BZLF1 of Epstein-Barr virus. J. Virol.72, 5128–5136.
51. Sapetschnig, A., Rischitor, G., Braun, H., Doll, A., Schergaut, M., Melchior, F. & Suske, G. (2002). Trans-cription factor Sp3 is silenced through SUMO mod-ification by PIAS1. EMBO J.21, 5206–5215.