Interpretation of geographic variation in size
of American eel Anguilla rostrata
elvers on the
Atlantic coast of North America using their life
history and otolith ageing
Chia-Hui
Wang,
Wann-Nian
Tzeng*
Department of Zoology, College of Science, National Taiwan University. No. 1, Sec. 4, Roosevelt Rd, Taipei, Taiwan 10617, ROC
ABSTRACT: Elvers of the .4merican eel Anguilla rostrata (LeSueur) were collected from 6 estuaries ranging in location from Haiti to the U.S. Atlantic coast and north to Canada during the fishing season in 1995. Time of metamorphosis from leptocephalus to glass eel was determined from otolith daily growth increments, In which the increment width and strontium/calcium ratlos changed drastically. Mean age of the leptocephalus at metamorphosis (T,) was 189 to 214 d , while age upon arrival at the estuaries (T,) was 220 to 284 d . Time from metamorphos~s to arrival at the estuanes (T,-,,) was 32 to 80 d . Faster-growing and earlier-metamorphosing leptocephali migrate to the mid-polnt of the sampling range, but slower-growing and delayed-metamorphosis leptocephali occur in both the southern and northern ranges. Duration of the glass eel phase was longer in the north than in the south, while glass eels grew faster in the south than in the north. Mean total length of elvers at entry to the estuaries increased significantly from 48 mm in the south to 60 mm in the north. Geographic variation in elver length was due to duration of the glass eel phase rather than timing of metamorphosis from lepto- cephalus to glass eel, because elver length was significantly positively correlated with T, , (K = 0.79, p < 0.01) but not significantly correlated with T,, (p > 0.05). Duration of the glass eel phase was influ- enced by the coastal oceanographic conditions.
KEY WORDS: Otolith microchemistry Aging - Early life history. Geographic vanation in size . Elvers American eel
INTRODUCTION
The American eel Anguilla rostrata (LeSueur) is a temperate catadromous fish, widely distributed in the rivers of eastern North and Central America (Schmidt 1909). Developn~ental stages of the American eel are similar to those of Anguilla anguilla and include: lepto- cephalus, glass eel, elver, yellow eel and silver eel (Bertin 1956). The American eel has a panmictic popu- lation (Avise et al. 1986) which spawns in the Sargasso Sea in the southwestern North Atlantic Ocean (Schrnidt 1925, Kleckner e t al. 1983, Kleckner & McCleave 'Addressee for correspondence
E-mail: wnt@ccms.ntu.edu.tw
1988). After a long-distance oceanic migration, the leptocephali metamorphose to glass eels on the continental shelf, and then become elvers in the estu- aries (Bertin 1956, Tesch 1977). Due to their wide dis- tribution in the open ocean and difficulty in collectlon, the biology of leptocephali is poorly known (Kleckner & McCleave 1982, 1985, Schoth & Tesch 1982, Caston- guay & McCleave 1987, Miller & McCleave 1994). The size of elvers at entry to the estuary tends to increase with increasing distance from the spawning area (Vladykov 1966, Smith 1968, Haro & Krueger 1988). This phenomenon was initially explained by a sex/size hypothesis that the female grows faster than the male and that the female predominates more in the north than in the south (Vladykov 1966). Haro & Krueger O Inter-Research 1998
36 Mar Ecol Prog Ser 168: 35-43, 1998
N. Atlantic Current
L
N. Equatorial Current(1988) rejected Vladykov's hypotheses 90°W 75OW 60°W 45"W 30°W and proposed that the geographic trend
in elver length may be due to the variation
in the timing of metamorphosis of lepto- cephali to the glass eel stage. However, until now there have been no studies on
the a g e determination of leptocephalus at metamorphosis to substantiate this speculation.
Daily growth increments in otoliths 30°N
(Pannella 1971) have been widely used to determine the daily age (Campana & Neilson 1985), growth rate (Volk et al. 1984), life history and change of habit of fish (Campana 1984). The age of Japan-
ese eel Anguilla japonica elvers at meta- 10oN morphosis and entry to various estuaries
has been studied (TsukamOtO Fig. 1 Sampling sites of Anguilla rostrata elvers (RI. Haiti; R2: Florida; R3: & Umezawa 1990,1994, Tzeng 1990,1996, North Carolina; R4: Rhode Island; R5: New Brunswick; R6: Nova Scotia)
Otake et al. 1994, Tzeng & Tsai 1994, Cheng & Tzeng 1996). These studies con-
clude that a visible metamorphosis check is deposited EPMA (Electron Probe Microanalysis; EPMA Model in the otolith at metamorphosis. Strontium concentra- 8800) was used to analyze the weight ratio (W %o) tion in the otolith decreases dramatically during meta- of strontium (Sr) to calcium (Ca) in eel otoliths. SrTi03 morphosis. Since growth increments in otoliths of and C a C 0 3 were used as standards for Sr and Ca, American eel elvers a r e deposited daily (Martin 1995), respectively. Sr and Ca were measured along the oto- the a g e of the leptocephalus at metamorphosis and the lith maximum axis through the primordium with a age of the elver at entry to the estuary can be deter- beam condition of 0.01 p A and 15 kV. At intervals of mined from daily growth increments and the strontium approximately 10 pm, the electron beam was focused content of the otolith. on an area approximately 5 pm in diameter. The This study investigates the causes of the geographic energy dispersive strength of Ca and Sr was evaluated variation in size of American eel elvers on the east using four 5 S scanning periods. After ZAF (Z, atomic coasts of North America and Haiti by using the number effect; A, absorption factor; F, fluorescence
microstructure and microchemistry of otoliths. effects) correction (Goldstein et al. 1984), the weight
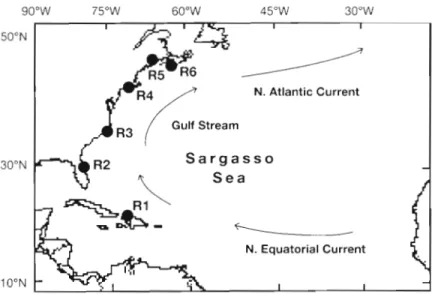
ratio of Sr and Ca was calculated using a constant CO2 of 43.87% (Chen & Tung 1984). The metamorphosis MATERIALS AND METHODS check at which the leptocephalus metamorphosed to the glass eel stage was inferred from the point at which American eel elvers were collected from 6 estuaries the Sr/Ca ratio changed (Tzeng & Tsai 1994, Tzeng on the Atlantic coasts of Haiti, the USA a n d Canada 1996). To link the time series change of Sr/Ca ratio (Table 1, Fig. 1). All samples were collected in 1995, with otolith growth increment, the otolith was washed with 1 additional sample collected in 1997 from Florida
because the corresponding sample size in 1995 was
Table 1. Sampling sites (shown in Fig. 1) and dates and small. The elvers were collected with a dip net during sample sizes of Anguilla rostrata elvers
their upstream migration on a night-time flood tide (Usui 1991) and, were preserved in 95% alcohol. Total lengths to 0.1 mm were measured after alcohol fixa- tion. Pigmentation stages, which indicate the develop- mental stage from glass eel to elver, were assessed according to pigment distribution on the body surface fstrubbera 1913). Saaittal otoliths of the elvers were
Site Date N
R1 Haiti D e c l 7 , 1995 115
R2 USA Feb 28. 1995 4
Jan 22, 1997 50 R3 North Carolina, U.S.A. Mar 22, 1995 50 R4 Annaauatucket River.
4 , 4
extracted from the vestibular apparatus and prepared for microchemistry analysis a n d a g e determination as described by Tzeng (1990, 1996) and Tzeng & Tsai
~ h o ' d e Island, USA Apr 14, 1995 100 R5 musquash River.
New Brunswick, Canada Apr 28, 1995 67 R6 East River, Nova Scotia, Canada May 29, 1995 93
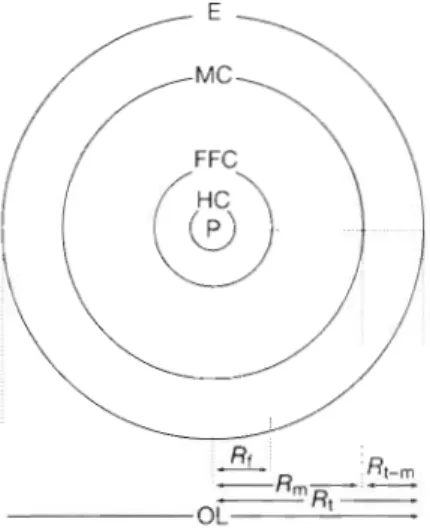
Fig. 2. A schematic diagram of radii measurements in elver otoliths. P: primordium; HC: hatching check; FFC: first feed- ing check; MC: metamorphosis check; E: edge. Radii were measured from the pnrnordium to the first feeding check ( R , ) , to the first feed~ng check (R,,) meta~norphosis check, and to the otolith edge (R,); R,.,: distance from the metamorphosis
check to the otolith edge; OL: otollth length
with acetone to take off the carbon coating after EPMA analysis a n d was then prepared for a g e determination using an SEM (scanning electron microscope, Hitachi S-520) (Tzeng 1996).
Otolith radii from the primordium to the first feeding check (Rf), to the metamorphosis check (R,) a n d to the otolith e d g e (R,) a n d the distance from the metamor- phosis check to the otolith e d g e (R,-,) w e r e measured using a n SEM photograph taken a t 300x magnification along the longest sagittal axis of the otolith (Fig. 2). The growth increments (N,,, N
,_,,,
a n d N,) on the radii of R,, R,_,, and R, were counted on SEM photographs at 1200x magnification. Because increments a r e not deposited in the core of the otolith for the 5 d of the yolk-sac stage (Yamamoto & Yamauchi 1974, Yama- uchi e t al. 1976, Tzeng 1990, Tzeng & Tsai 1994), 5 d ( N o ) were added to the number of growth increments to estimate the daily a g e of elvers at metamorphosis (T,, = N,,+
No) a n d the daily a g e at estuarine arrival (T, = N,+
N o ) . The time between metamorphosis a n d arrival at the estuary (T,-,) equals N l ~ ,. Daily growth increments were counted for 89 otoliths.Three kinds of otolith growth rate estimates were made:
Overall growth rate of otolith, C, = R,/T, (1)
Early growth rate of otolith, G, = Rnl/Tm (2) Estuarine growth rate of otolith, G,_, = R,-,/T,-, (3) Because the counting of daily growth increments was time consuming and the increments near the metamorphosis a r e a on the otolith w e r e often diffusive
a n d obscure, the daily a g e of samples without counting daily growth increment was calculated from otolith growth rate a n d otolith radius:
R,,-& + N o
T", = -
G", (4)
where G,, and C,-, were obtained from Eqs. (2) & ( 3 ) , a n d No is the adjustment factor (5 d ) for yolk-sac stage duration. Otolith growth rate of the elvers in the 6 estuaries was site-specific; t h e rate was calculated a t each sampling site to estimate daily ages.
T h e homogeneity of m e a n total length, various daily a g e s (T,, T,,, a n d T,_,) a n d otolith growth rates (C,, G,, a n d G,.,) among the 6 estuaries was tested with Scheffe's multiple range analysis (Sokal & Rohlf 1969). Correlations between total length a n d T,, T,,, a n d T,-, for the elvers were calculated.
RESULTS
Total length and pigment stage
Total lengths of the elvers collected from the 6 estu- aries on the Atlantic coasts of Haiti a n d North America ranged between 41 a n d 65 mm. Two different length groups w e r e identified: m e a n length of the northern group [Nova Scotia (R6), New Brunswick (R5) a n d Rhode Island (R4)] was significantly greater than that of the southern group [North Carolina (R3), Florida (R2) and Haiti ( R I ) ] . Mean lengths ranged from 47.8
to 49.0 mm for the southern group and from 58.5 to 60.0 mm for the northern group, with a difference in m e a n length of approximately 12 m m (Fig. 3 ) .
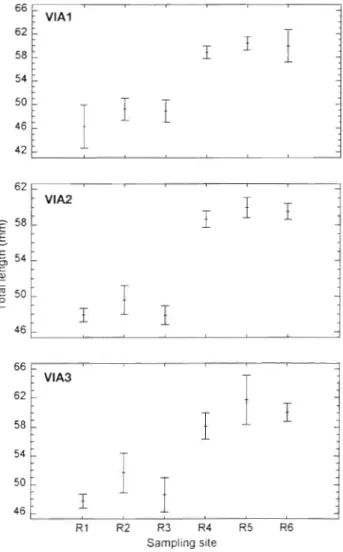
T h e pigment development stage of elvers a t estuar- ine arrival was mostly VB a n d VIAl for R2, but VIAl and VIA2 for the other 5 sites. This may indicate that the immigration of elvers in Florida occurred at a n earlier stage than for the other sites. In addition, the trend of mean total length of elvers from different sampling sites was similar among pigmentation stages, indicating that t h e difference in m e a n length between northern a n d southern groups w a s independent of elver pigmentation stage (Fig. 4 ) .
Otolith microstructure a n d microchemistry The primordium of the elver otoliths w a s amorphous. Between the hatching check (HC) a n d the first feeding
3 8 Mar Ecol Prog Ser 168: 35-43, 1998
Feb 18.1995. N = 4 (Black) Jan 22.1997. N = 50 (Grey) 49.03 + 2.58 mm
M - R 6 May 29- 1995. N =93 leptocephalus stage. Daily growth increments were
Dec 17. 1995. N - 115 47.79 + 2.33 mm 10 o 50 60 70 Total length (mm) Mean G SDI:
-1
59.64 t 2.42 mm wide a n d clear in the beginning of the stage, but-
became indistinct and almost uncountable across the . m m , < . - . . 8 . . , MC check. This indicated that the growth of the lepto-
Fig. 3. Anguilla rostrata. Length frequency distribution of elvers collected from 6 estuaries in Haiti and North Amenca
(Sites R1 to R6 in Fig. 1). Mean + S D of each distribution is
also indicated
check (FFC), no distinct daily growth was discernible. Beyond the FFC, there were distinct daily growth in- crements. The shape of the otolith crystalline arrange- ment changed from a circular to a radial form at approximately
2h
of the total radius distance from the primordi.um. Also, the increment widths at the transl- tion boundary become very narrow. The changes in both crystalline pattern a n d increment width yielded a distinct check, which was interpreted as the metamor- phosis check (MC) (Fig. 5 ) .Changes in crystalline arrangement and increment width in the otolith reflect d~fferent developmental stages of the fish. The nucleus from the primordium to the FFC, which is deposited before the yolk-sac stage, has no discernible daily growth increments. The mean nucleus radius ranged between 9.5 and 15.0 pm. The
zone from the FFC to the MC was deposited during the
cephalus was fast in the early stage and then gradually
slow. From the MC to the otolith edge, the crystalline 2 0 - ~ 5 Apr28.1sss.N=67 arrangement became radial in form and had wide
59.99 3 3.15 mm
10 - increments, indicating that growth became fast after
metamorphosis (Fig. 5). Daily growth increments from the MC to the otolith edge were wider in the southern grap than in the northern group of elvers, i.e. the daily
R1 R 2 R3 R4 R5 R6 Sampling site 2 0 - 10
-
5
Fig. 4. Anglulla rostrata. Mean (* 95".. confidence intervals) total length by pigmentation stage (VIA1, VIA2 and VIA3) of elvers collected from 6 estuaries in Haiti and North America
R 4 growth of glass eel in the period after metamorphosis
npr 14, 1995. N = 100
-
58.52 t 2.83 mm was faster in the southern group than in the northernI
, , , , , , , , group.X 0 . 7 - n * . . .
- ~ - ~ -
o Sr/Ca ratio in otoliths was approximately 5 X 10-3 in
the nucleus and increased with otolith growth, reach-
"'"- R 3 ing a maximum ranging between 1.4 and 2.0 X 10-* a t
LL 10
0
Mar 22,1995, N = 50
48.79 t 2.87 mm the MC. Beyond the check, the Sr/Ca ratio sharply
-
decreased (Fig. 6). The dramatic changes in Sr/Carailos .n rhe otolith has been proposed to correspond to both rhe time oi metamorphosis from leptocephdlus
I
to glass eel and the time of migration from oceanic to coastal waters in Anguilla japonica (Otake et al. 1994,I
Tzeng & Tsai 1994, Arai et al. 1997).I
Geographic trend in otolith growth rateI
The geographic trend in mean growth rate of the elver otoliths at the 6 sampling sites differed in the period before metamorphosis (G,) and in the period from metamorphosis to estuarine arrival (G,.,),
(Table 2 ) . G,, was larger at the mid-polnt (R4) of the 6I
sampling sites than at the southward and northward sites ( R 1 and R6) (Table 2). Conversely, the estuarine growth rate (G,-,) was larger in the southern group (R1 to R3) than in the northern group (R4 to R 6 ) (Table 2). The geographic trend in G, roughly paral- leled that of C,-, because the variance of G,_,, was larger than C, (Table 2).Timing of metamorphosis and estuarine arrival
The mean ages of the elvers at the 6 sampling sites ranged from 220 to 284 d at estuarine arrival (7,) a n d from 189 to 214 d at metamorphosis (T,), with 32 to 80 d between metamorphosis and estuarine arrival
(T,-,). An analysis of variance (ANOVA) indicated that
T,, T, and T,-, differed significantly among sampling sites (Table 3).
T,, was shortest at the mid-point (R3) of the 6 sam- pling sites, and increased toward the south (R1 and R2) and north (R4 to R6) (Table 3). On the other hand, T,-, was larger in the northern group (R4 to R6) than the southern group (R1 to R3) (Table 3), consistent with the geographic trend of elver length (Fig. 3 ) . The geo- graphic trend in T, roughly paralleled that of T,_, be- cause the variance of T,-, is larger than Tm (Table 3).
The geographic trends in Tm and T,_, were inversely related to G, and G,_, (Table 2).
Relationship between total length and T,. T,,, and
T,-,
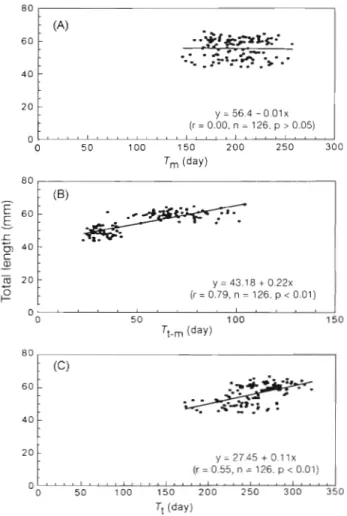
The total length of American eel elvers at estuarine
arrival was not significantly correlated with T,,, (r =
Fig. 5. Anguilla rostrata. Daily growth increment of an elver (49.35 mm) otolith collected from Halt1 [Site R I ) . FFC: first feeding check; HC: hatching check; MC: metamorphosis
40 Mar Ecol Prog Ser 168. 35-43, 1998
Table 2 Angculla rostrata. Homogeneity tests for otolith growth rate (mean k SD, pm d - ' ) in elvers In Haiti and North America measured from the primordium to the otolith edge (G,], to the metan~orphosis check (G,), and between the metamorphosis
check and the otolith edge (G,.,)
u
-200 -150 -100 -50 0 50 100 150 200 Distance from primordium (pm)
Fig. 6. Ang~iilla rostrata. Sr/Ca concentration ratios measured along the maximum axis through the pnmordium of elver otoliths collected from 6 estuanes in Haiti and North Amenca.
Arrows indicate position of the metamorphosis check
0.00, p > 0.05), but was significantly positively corre- lated with T,_, (r = 0.79, p < 0.01) and also with T,
(r = 0.55, p < 0.01) (Fig. 7). Thus, the size of elvers at estuarine arrival increased with the duration from metamorphosis to estuarine arrival, but was indepen-
dent of the age at metamorphosis.
DISCUSSION AND CONCLUSION
The mean total length of elvers at entry to 6 estuaries on the Atlantic coasts of Haiti, USA and Canada dif- fered significantly between northern and southern groups. Factors which have the potential to influence elver length include pigmentation stage of the elver in the estuary (Tzeng 1985), seasonal decline in length of elvers at entry to the estuary (Strubberg 1913, Matsui 1952, Jellyman 1979, Tzeng 1985) and annual vari- ability in elver characteristics at entry to the estuary
Site Mean
*
SD G, R6 R5 R4 R1 R2 R3 Total G, R6 R 1 R2 R3 R5 R4 Total G , _ , R5 R6 R 4 R 1 R3 R2 Total Homogeneous groups A A AB AB AB BTable 3. Anguilla rostrata. Homogeneity test for daily a g e (mean r SD) of elvers from estuaries in Haiti and North America (T,), for daily age at metamorphosis (T,,), and for the time between metamorphosis and arrival at the estuary (T,_,)
Site 7, R3 R 1 R2 R4 R5 R6 Total T, R3 21 R4 26 R5 17 R 1 25 R6 3 2 R2 4 Total 125 R2 4 R4 26 R6 32 R5 17 Total 125 Mean
*
SD Homogeneous groups 220.4 k 33.22 A 241.6*
18.51 B 247.8*
16.24 BC 251.8 r 16.62 BCD 272.3*
15.72 CDE 283.5+
18.18 E 255 3*
30.24Fig 7. A n g u ~ l l a rostrata. Relationships between total length of elvers and ( A ) a g e of leptocephali at metamorphosis (T,),
(B) time between metamorphosis and arrival at the estuaries
(T, .,,,) and (C) age at estuarine arrival (T,)
(Haro & Krueger 1988). Pigmentation stage was found to have no effect on the elver length at estuarine arrival. Annual variability was negligible because the elvers from all estuaries were collected during 1995. Also, the elvers collected from Florida in 1995 and 1997 had similar length distributions. Within an estuary, elver length tends to decline through the run (Tzeng 1985). Run timing is earlier in the southern than in the northern portions of the range and elver length increases from south to north (Vladykov 1966, Haro & Krueger 1988). The observed size difference of 12 mm between the mean lengths of elvers from Haiti and Canada was larger than the seasonal decline, which is usually less than 6 % in American eel (Haro & Krueger 1988). The geographic difference in elver length of American eel was not d u e to differences in sampling date.
Haro & Krueger (1988) proposed that the geographic trend in elver length may be d u e to the variation in the timing of metamorphosis of leptocephali to the glass
eel stage. However, we found that the correlation between the age at metan~orphosis and the length of elvers at estuarine entry was not significant, although the correlation between elver length and the time between metamorphosis and entry to estuaries was significant. This indicated that the geographic differ- ence in elver length along the east coast of North America was correlated with the difference in the duration of the glass eel stage in coastal waters. In coastal waters, the glass eel phase in the northern part of the range lasted approximately 62 to 80 d , which is 2 to 3 times longer than in the southern part of the range (32 to 34 d ) . Thus, higher elver length at north- ern sites in con~parison to southern sites may be due to the longer duration of the glass eel phase in coastal waters. The early life history, described in the follow- ing, of the elver contributes to the further understand- ing of this relationship.
The early life history of the American eel Anguilla rostrata is similar to that of the Japanese eel Anguilla japonica (Tsukamoto 1990, 1992, Tsukamoto & Ume- zawa 1990, 1994), with 2 distinct stages: the lepto- cephalus in the open ocean and the glass eel and elver in coastal waters. American eels spawn in the south- western Sargasso Sea, after which the leptocephalus drifts with the Florida Current and Gulf Stream, then metamorphoses into a glass eel a s it approaches the continental shelf and becomes an elver a t entry to the estuary. The geographic trend in the mean daily ages of the leptocephalus (T,) and the time from metamor- phosis to arrival at the estuary (T,-,) differed among the 6 estuaries sampled. T,, was smallest at the mid- point of the 6 sampling sites (North Carolina) and in- creased towards the southern sites (Haiti and Florida) and northern sites (New Brunswick and Nova Scotia). On the other hand, T,-, was smaller in the south (Haiti to North Carolina) than in the north (Rhode Island to Nova Scotia). This suggests that the migration a n d dispersal patterns differ between the leptocephalus and glass eel stages. In addition, there was an in- verse geographic trend between growth rate a n d leptocephalus a g e at metamorphosis. This indicates that faster-growing a n d earlier-metamorphosing lepto- cephali entered the middle of the sampling range, while slower-growing, delayed-metamorphosis lepto- cephali dispersed southward and northward. This re- sult is consistent with leptocephali entering the Gulf Stream system directly from the Sargasso Sea, as pos- tulated by Kleckner & McCleave (1982), rather than by a more southern route through the Bahamas.
However, when the eel metamorphoses from lepto- cephalus to glass eel and enters coastal waters, geo- graphic trends in a g e (T,-,) and growth rate (G,-,) differed completely from that of the leptocephalus in the open ocean. T,-, was smaller in the southern part
4 2 Mar Ecol Prog Ser
of the sampling range (Haiti to North Carolina) than in the northern part (Rhode Island to Nova Scotia), but G,.,, was the opposite, being smaller in the north than in the south. Thus, glass eels grew faster, but stayed in coastal waters a shorter time, in the south than in the north. The duration of the glass eel phase in coastal waters may be regulated by coastal oceanographic conditions. The coastal waters off the southern sam- pling sites (Haiti to North Carolina) are influenced by the warm Gulf Stream, but the northern waters (Rhode Island to Nova Scotia) a r e influenced by the cold Labrador Current (Lalli & Parsons 1993). Higher tem- peratures in the south will accelerate the development of the glass eel, producing faster glass eel growth, more rapid elver development and earlier immigration to the estuary in the south than in the north. Although growth of the elver in the glass eel phase was faster in the south, their duration in coastal waters was shorter, which may lead to a smaller total length of elvers at entry to the estuary in the south than in the north.
In conclusion, the American eel AnguiUa rostrata elvers are smaller in the southern than in the northern estuaries of the Atlantic coast of North America be- cause the d u r a t ~ o n of the glass eel phase in the coastal waters is shorter in the southern than in the northern part of their range and not because of differences in the timing of metamorphosis from leptocephalus to glass eel.
Acknowledgements. This study was financially supported by the National Science Council, Republic of China (NSC 85231 lB002032, a research project to W N.T.). The authors are grateful to Dr G. Williamson for arranging the sampling, to Dr W. H. Krueger, Mr B. M. Jessop and Mr A. T Peng for providing the fish specimens from the United States, Canada and Haiti, to Dr C. H. Chen for electron probe microanalysis, and to Mr B. M Jessop and Dr S. E . Campana for revlewing the manuscript.
LITERATURE CITED
Arai T, Otake T, Tsukamoto K (1997) Drastic changes in otolith microstructure and microchemistry accompanying the onset of metamorphosis in the Japanese eel Anguilla japonicd. Mar Ecol Prog Ser 161:17-22
Avise JC, Helfman GS, Saunders NC, Hales LS (1986) Mito- chondrial DNA differentiat~on in North Atl.antic eels: pop- ulation, genetic consequences of a n unusual life hlstory pattern. Proc Natl Acad Sci USA 83:4350-4354
Bertin L (1956) Eels-a biological study. Cleaver-Hume Press, London
Campana SE (1984) Microstructural growth patterns in the otoliths of larvdl and juvenile starry flounder, Platichthys s t ~ l l a t u s . Can J Zool 62.1507-1512
Campana SE, Neilson JD (1985) Microstructure of fish oto- liths. Can J Fish Aquat Sci 42:1014-1032
Cdstonguay M, McCleave JD (1987) Vertical distributions, die1 and ontogenetic vertical migrations and net avoid- ance of leptocephali of Anguilla and other common spe-
cies in the Sargasso Sea. J Plankton Res 9:195-214 Chen CH, Tung TC (1984) On-line data reduction for electron
mlcroprobe analysis. Acta Geol Taiwan 22:196-200 Cheng PW, Tzeng WN (1996) Timlng of metamorphosis and
estuanes arrival across the dispersal range of the Japan- ese eel Anguilla japonica. Mar Ecol Prog Ser 131:87-96 Goldstein Jl, Newbury DE. Echlln P, Joy DC, Fion C, Lifshin E
(1984) Scanning elrctron microscopy and x-ray inicro- analysis. Plenum Press, New York
Haro AJ. Krueger WH (1988) Pigmentation, size. and migra- tion of elvers (Anguilla rostrata (Lesueur)) in a coastal Rhode Island stream. Can J Zool 66:2528-2533
Jellyman DJ (1979) Upstream migration of glass-eels (Anguilla spp.) in the Waikato a v e r . Ibid 13(1):13-22 Kleckner RC, McCleave JD (1982) Entry of migrating
American eel leptocephali into the Gulf Stream system. Helgolander Meeresunters 351329-339
Kleckner RC, McCleave JD (1985) Spatial and temporal dis- tnbution on American eel larvae in relatlon to North Atlantic Ocean current systems. Dana 4:67-92
Kleckner RC, McCleave JD (1988) The northern limit of spawning by Atlantic eels (Anguilla spp ) in the Sargasso Sea in relation to thermal fronts and surface water masses J Mar Res 46:647-667
Kleckner RC, McCleave JD, Wippelhauser GS (1983) Spawn- ing of Amencan eel, Anguilla rostrata, relative to thermal fronts in the Sargasso Sea. Environ Biol9:289-293 Lalli CM, Parsons TR (1993) Biological oceanography-an
~ntroduction. Pergamon Press, Oxford
Martln MH (1995) Validation of daily growth Increments In otoliths of Anguifla rostrata (Lesueur) elvers. Can J 2001 73:208-211
Matsui I (1952) Studies on the morphology, ecology and pond- culture of the Japanese eel (Anguilla japonica Temminck & Schlegel). J Shlmonosekl Col1 Fish 2:l-245
Miller MJ, McCleave JD (1994) Species assemblages of lepto- cephali in the Subtropical Convergence Zone of the Sargasso Sea. J Mar Res 52:743-772
Otake T, lshii T, Nakahara M , Nakamura R (1994) Drastic changes in otolith strontium/calcium ratlos In leptocephall and glass eels of Japanese eel Anguilla]aponica. Mar Ecol Prog Ser 112:189-193
Pannella G (1971) Fish otoliths: daily growth layers and peri- odical patterns. Science 173 1124- 1127
Schmldt J (1909) Remark on the metamorphosis and distnbu- tlon of the eel (Anguilla vulgaris TURT.) Meddr Komm Havunders Ser Fisk 3(3): 1-1 7
Schmidt J (1925) The breeding places of the eel. Rep Smith- son Inst 1924-279-316
Schoth M , Tesch FW (1982) Spatlal distribution of 0-group eel larvae (Angujlla sp.) in the Sargasso Sea Helgolander Meeresunters 35:309-320
Smith DG (1968) The occurrence of larvae of the Amencan eel, Anguilla rostrata, in the Straits of Florida and nearby areas. Bull Mar Sci 18:280-293
Sokal RR, Rohtf FJ (1969) Biometry. WH Freeman, San Fran- CISCO
Strubberg AC (1913) The metamorphosis of elvers a s influ- enced by outward conditions. Medd Komm Dan Fisk Havunders 4.1-1 1
Tesch FW (1977) The eel-biology and management of Anguilla eel Ch.apman and Hall Press, London
Tsukamoto K (1990) Recruitment mechanism of the eel, Anguilla japonica, to the Japanese coast. J Fish Biol 36: 359-371
Tsukamoto K (1992) Discovery of spawning area for Japanese eel. Nature 355789-791
Tsukamoto K, Umezawa A (1990) Early life history and oceanic migration of the eel Anguilla japonica. La Mer 28: 188-198
Tsukamoto K , Umezawa A (1994) Metamorphosis: a key fac- tor of larval migration determining geographic distribu- tion and speciation of eels. In: Proc 4th Indo-Pacif~c F ~ s h Conf, Bangkok, Thailand, 28 Nov-4 Dec 1993. Faculty of Fisheries, Kasetart University, Bangkok, p 231-248 Tzeng WN (1985) Immigration timing and activity rhythms of
the eel, Anguilla japonica, elvers in the estuary of north- ern Taiwan with emphasis of environmental influences. Bull Jpn Soc Fish Oceanogr 47/48:11-28
Tzeng WN (1990) Relationship between growth rate and age at recruitment of Anguilla japonica elvers in the Taiwan estuary as inferred from otolith growth increment. Mar Biol 107:75-81
Tzeng WN (1996) Effects of salinlty and ontogenetic move- ment on strontium:calciun~ ratio in otolith of the Japanese eel, Anguilla japonica Temminck & Schlegel. J Exp Mar Biol Ecol 199:lll-122
Editorial responsibjlitjf- Otto Kinne (Editor), Oldendorf/Luhe, Germany
Tzeng WN, Tsai YC (1994) Changes in otolith microchemistry of the Japanese eel, Anguilla japonjca, during its migra- tion from the ocean to the rivers of Taiwan. J Fish Biol 45:671-683
Usul A (1991) Eel culture. Fishing News Books Blackwell Scientific Publications Inc, Oxford
Vladykov VD (1966) Remarks on the American eel (Anguilla rostrata Lesueur). Sizes of elvers entering streams; the relative abundance of adult males and females; and the present economic importance of eels in North America. Verh Int Verein Theor Angew Limnol 16:100?-1017 Volk EC, Wissmar RC, Simenstad C A , Eggers DM (1984) Re-
lationship between otolith microstructure and the growth of juvenile chum salmon (Oncorhynchus keta) under dif- ferent prey rations. Can J Fish Aquat Sci 41:126-133 Yamamoto K, Yamauchi K (1974) Sexual maturation of Japan-
ese eel and production of eel larvae in the aquarium. Nature 251:220-222
Yamauchi K, Nakamura M, Takahashi H, Takano K (1976) Cultivation of larvae of Japanese eel. Nature 263:412 Submitted: September 15, 1997; Accepted: January 22, 1998 Proofs received from author(s). J u n e 15, 1998

![Table 2 Angculla rostrata. Homogeneity tests for otolith growth rate (mean k SD, pm d - ' ) in elvers In Haiti and North America measured from the primordium to the otolith edge (G,], to the metan~orphosis check (G,), and between the met](https://thumb-ap.123doks.com/thumbv2/9libinfo/8846429.240674/6.882.132.398.127.656/angculla-rostrata-homogeneity-otolith-america-measured-primordium-orphosis.webp)