The adjuvant effect of high-molecule-weight polysaccharides purified from
Antrodia cinnamomea on dendritic cell function and DNA vaccine
Abstract
The biological activity of an edible basidiomycete Antrodia cinnamomea (AC) has been studied extensively. Many effects, such as anti-cancer, anti-inflammation, and antioxidant, are reported by evaluating either crude extracts or compounds isolated from AC. However, the research for AC function in enhancing immunity is rare. The aim of this study is to investigate the active components and the mechanism involved in the immunostimulatory effect of AC. We found that polysaccharide (PS) played a major role to activate dendritic cell (DC), a critical leukocyte in initiating immune response, in the water extract of AC. According to the molecular size, we further purified and identified that the PS fraction with high molecular weight (more than 100 kDa) exhibited the activating effect. The AC high-molecular-weight PSs (AC
addition, the DC-induced antigen-specific T cell activation and Th1 differentiation were
increased by AC hmwPSs. Studying the molecular mechanism, we confirmed the activation of MAPK and NF-B pathways in DCs after AC hmwPSs treatment. Furthermore, we proved that TLR2 and TLR4 were required for the stimulatory activity of AC hmwPSs on DCs. In a mouse tumor model, we demonstrated that AC hmwPSs enhanced the anti-tumor efficacy of HER-2/neu DNA vaccine by facilitating specific Th1 responses. Thus, we conclude that hmwPSs are the major components of AC to stimulate DCs via TLR2/TLR4 and NF-B/MAPK signaling pathway. The AC hmwPSs have potential to be applied as adjuvant.
Introduction
Edible fungi have nutritional and medicinal values and can be the functional food as dietary supplements . Many edible fungi belong to Aphyllophorales (polypore), a large group of terrestrial fungi of the phylum Basidiomycota (basidiomycetes) . Recently, a precious
basidiomycete Antrodia cinnamomea [syn. A. camphorata] (AC) became a popular health food for reducing cancer and improving liver function in Taiwan . The effects of bioactive
components extracted or purified from AC have been studied extensively; however, the majority of investigations focus on the anti-cancer and anti-inflammation . The immunostimulatory effect of AC remains to be explored.
Polysaccharides (PSs) are the well-known bioactive compounds and broadly exist in living organisms. A great deal of PSs has been isolated from the fruiting bodies, cultured mycelia and culture filtrates of basidiomycetes . Many functions of dietary PSs have been identified, including anti-cancer, anti-inflammatory, anti-oxidant, and immunomodulatory effects . These biological activities of PSs are closely correlated to their physicochemical properties such as molecular size, types and ratios of constituent monosaccharides, and features of glycosidic linkages . Recently, studying the mechanism for immunomodulatory effect of PSs has become a major topic in glycobiology and immunology.
Immune system plays the most essential role to maintain human health against infectious agents and tumor cells. There are many types of immune cell in this system. A specialized leukocyte named dendritic cell (DC) is critical in initiating immune responses . They are in immature state in peripheral tissues and become mature when pathogens are recognized by pattern recognition receptors (PRRs) on DCs . Mature DCs then migrate to secondary lymphoid organs where they present antigens to naive T cells. The activated DCs up-regulate the
expression of MHC and costimulatory molecules for T cell activation and secrete a variety of cytokines and chemokines for T cell differentiation and recruitment . According to the key role in immune system, DCs have being applied as potent vaccines in treating cancer and viral infections . Thus, natural or synthetic substances that can stimulate DCs may potentially be developed into adjuvants and applied to immunotherapy and vaccination.
In this study, we examined the immunostimulatory effect of AC on mouse bone marrow-derived DCs (BMDCs). Our results showed that the PSs of AC possessed the ability to promote DC activation and function. Subsequently, we identified that the activating effect on DCs was mainly contributed by the high molecular weight PSs (hmwPSs) of AC. Furthermore, the AC hmwPSs enhanced the therapeutic effect of a DNA vaccine in mouse tumor model. Thus, we concluded that the AC hmwPSs may be an immunopotentiator and candidate for vaccine adjuvant.
Materials and Methods
Preparation and subfractions of AC PSs
The fruiting bodies of A. cinnamomea (10 g) were treated with distilled water (100 mL) at 100 °C for 1 h. The water extract was centrifuged, filtered through the Toyo No. 1 filter paper (Toyo roshi kaisha, Ltd., Tokyo, Japan), and concentrated with a rotary evaporator (R-210, Buchi Labortechnik AG, Flawil, Switzerland). Next, the extract was mixed with ethanol (80%, v/v), incubated overnight at 4 °C, and centrifuged to obtain the crude PSs. For further
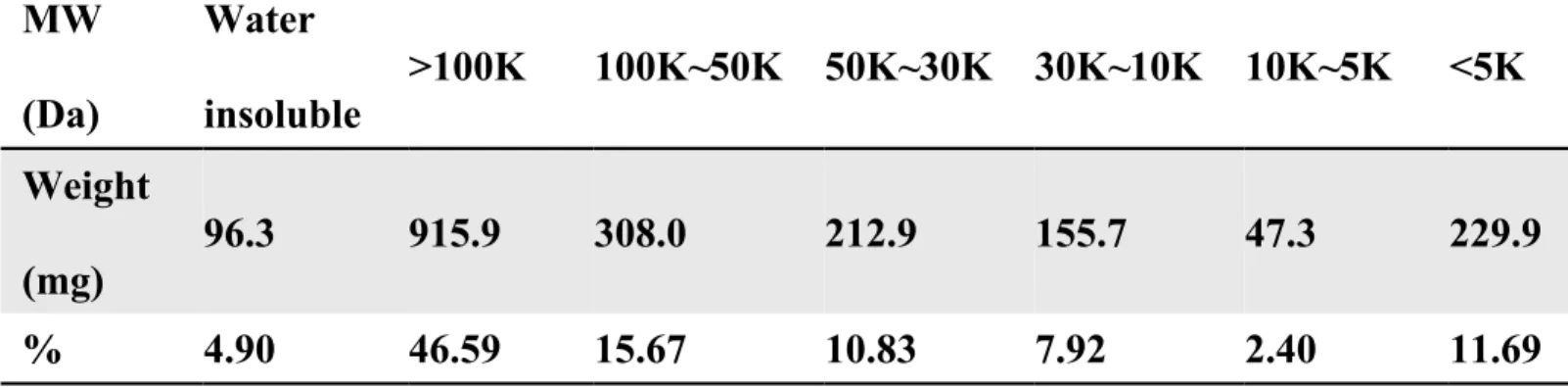
fractionation, the crude PSs were partially separated into 6 fractions by Amicon Ultra-15 5K, 10K, 30K, 50K, 100K centrifugal filter devices (Millipore, County Cork, Ireland). Briefly, crude PSs (2 g) was suspended with distilled water (20 mL)and centrifuged. The supernatant (10 mL) was applied to 100K filter unit of centrifugal filter device. After centrifugation, the 100K filter was transferred to a new tube and PSs (> 100 kDa) were eluted by centrifuged with the filter upside down, and then the PSs were lyophilized. Repeating the same protocol, we obtained other 5 fractions of PSs by using the reduced sizes of filter serially (50K, 30K, 10K, and 5K). The percentage and weight of each fraction were shown in Table 1.
Mice
C57BL/6, C3H/HeN, and TLR4 mutant (C3H/HeJ) mice were purchased from National Laboratory Animal Center (NLAC, Taipei, Taiwan) and National Cheng-Kung University (NCKU, Tainan, Taiwan). OT-I and OT-II TCR transgenic mice were provided by Dr. Clifford Lowell (UCSF, San Francisco, CA). TLR2-deficient mice were provided by Dr. Chih-Peng Chang (NCKU, Tainan, Taiwan). All animals were kept in a specific pathogen free facility at
Taichung Veterans General Hospital (Taichung, Taiwan) and handled according to protocols approved by Institutional Animal Care and Use Committee.
Preparation of mouse DCs
The mouse DCs were derived from bone marrow of mice as described previously . Briefly, bone marrow cells were isolated from the femurs and tibia of mice. After removing red blood cells, the cells were then seeded in 24-well plates with completed RPMI 1640 medium (L-glutamine, nonessential amino acids, sodium pyruvate, HEPES, 2-ME, penicillin/streptomycin) supplemented with 10% FBS and 10 ng/mL recombinant mouse GM-CSF (PeproTech). On day 7, the DCs (> 70%) were collected for analysis.
Detection of cytokine production
DCs were treated with AC PSs (crude extract or various fractions) or LPS (100 ng/mL) for 24 h. The production of cytokines (TNF-, IL-6, and IL-12) was quantified by ELISA
(eBioscience and R&D systems) as described previously .
Measurement of DC maturation
DCs were treated with PBS (control) or AC hmwPSs (5 or 10 g/mL) for 16 h.
Maturation was determined by measuring the expression of MHC and co-stimulatory molecules as described previously . After treatment, cells were collected and immunostained with
fluorescent dye-conjugated mAbs specific for mouse CD11c, I-Ab, H-2b, CD40, CD80, and CD86 (Biolegend), and then analyzed by flow cytometry. The mean fluorescence intensity (MFI) was indicated in each graph.
Assay for OVA-specific T cell activation
The DC-induced OVA-specific T cell activation was determined as described previously . Briefly, DCs were incubated with OVA257-264 (for OT-I) or OVA323-339 (For OT-II) (1 g/mL, synthesized by Echo Chemical Co., Taiwan) in the presence or absence of AC hmwPSs (10 g/mL) for 3 h, and then OT-I or OT-II T cells were added at ratio DC:T cell = 1:5 to the culture. T cell proliferation was measured by [3H]thymidine incorporation after 72 h. To detect IFN-γ production, supernatants were collected from the DC/T cell cultures and the IFN-γ production was measured by ELISA (eBioscience).
Detection of MAPK activation
The MAPK activity was determined by Western blot. Briefly, DCs were treated with AC hmwPSs (10 g/mL) and total proteins were extracted by lysis buffer (50 mM Tris/HCl; 150 mM NaCl; 1% (v/v) Nonidet P-40; 0.25% (w/v) sodiumdeoxycholate; 1 g/mL leupeptin; 1 g/mL pepstatin; 1 g/mL aprotinin; 1 mM PMSF; 1 mM EDTA; 1 mM NaF, and 1 mM Na3VO4). The protein concentration was measured using BCA protein assay kit (Pierce, Rockford, IL, USA). Then, proteins were mixed with sample buffer, boiled, and separated on 10% SDS polyacrylamide gel. All proteins were transferred to nitrocellulose membrane and blocked in TBS with 0.1% (v/v) Tween 20 and 5% (w/v) non-fat dry milk. After blocking, the membranes were incubated with primary antibodies against ERK1/2 (no. 9102), p38 (no. 9212), JNK (SC- 571, Santa Cruz Biotechnology, Danvers, MA, USA), -Actin (no. 4970), and their
phosphorylated forms phospho-ERK1/2 (no. 9101), phospho-p38 (no. 9211), phospho-JNK (no. 4688) (all purchased from Cell Signaling Technology). Subsequently, the blots were washed and
incubated with secondary HRP-conjugated goat anti-mouse or goat anti-rabbit antibody (Jackson Immunoresearch, West Grove, PA, USA). Finally, the blots were washed and developed by enhanced chemiluminescence (GE Healthcare, UK) and analyzed using the LAS3000 system (Fujifilm, Tokyo, Japan). Densitometric analysis was performed with ImageJ software (National Institute of Health, Bethesda, MD, USA).
Analysis of NF-B activity
For detection of NF-B activity, we prepared the nuclear extracts first. DCs were treated with PBS (control), LPS (100 ng/mL), or AC hmwPSs (10 g/mL) and nuclear extracts were prepared using the NE-PER Nuclear and Cytoplasmic Extraction system (Pierce Rockford, IL, USA) according to manufacturer’s instruction. Protein concentrations were determined using a BCA protein assay kit. For each assay, 5 mg/mL of nuclear extracted was used in a TransAM NF-�B p65 ELISA kit (Active Motif, Carlsbad, CA) according to the manufacturer’s instruction.
The HER-2/neu DNA vaccine on a mouse model with p185neu-expressing MBT-2 tumor The mouse model implanted with p185neu-expressing MBT-2 tumor and immunization with HER-2/neu DNA vaccine were described previously . Briefly, p185neu-expressing MBT-2 cells were injected subcutaneously into the flank of C3H/HeN mice. After ten days, the mice were immunized with naked HER-2/neu DNA vaccine using a low pressure-accelerated gene gun (Bioware, Technologies Co. Ltd, Taipei, Taiwan) for three times in a week. In co-administration with AC hmwPSs, the PSs were intraperitoneally injected at the immunization site after vaccination. The anti-tumor effect of vaccination was then determined by monitoring the size of p185neu-expressing MBT-2 tumor every three days. Tumor size was calculated using
the formula for a rational ellipse: (m1 × m2 × m2 × 0.5236), where m1 represents the longer axis and m2 the shorter axis. Mice were sacrificed when tumor size was bigger than 2500 mm3 in mean diameter.
Analysis for Th1 responses in mice immunized with HER-2/neu DNA vaccine
The Th1 responses were determined by intracellular staining for IFN- production by CD8+ T cells. The inguinal lymph node cells were isolated from mice with various treatments and stimulated with recombinant HER-2/neu protein (10 g/mL, R&D System) in complete RPMI 1640 medium overnight. Cells were staining with anti-CD8 and anti-IFN- antibodies and then analyzed by flow cytometry. The percentage of CD8+IFN-+ cells in total CD8+ T cells was shown in each graph. In addition, the amount of IFN- and IL-4 were measured by qPCR. CD4+ T cells were purified using Dynal CD4 Negative Isolation Kit (Dynal AS, Oslo, Norway). Then, total RNA was prepared by using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and cDNA was generated using oligo dT as primer and reverse transcriptase (Promega, Madison,
Wisconsin, USA) according to the manufacturer’s instructions. Primer pairs for IFN- IL-4, and hypoxanthine guanine phosphoribosyl transferase 1 (HPRT) were used as described previously . ABI 7500 Fast Real-Time system with SYBR Green PCR Master Mix (Applied Biosystems) was used for quantitative real-time PCR. The expressions of IFN- and IL-4 were normalized with HPRT expression. Data were shown as fold comparing to control group with vector alone.
Data analysis
Significance of the AC PS treatment in comparison with control PBS treatment or HER-2/neu DNA vaccine immunization in comparison with vector alone vaccination was calculated
using a Student’s t-test with 2-sample equal variance with a 2-tailed distribution. P < 0.05 was considered significant.
Results
Identification of high molecular weight PSs (hmwPSs) as major fraction of AC for DC activation
Various activities of the PSs isolated from AC have been reported; however, the enhancement of immune response has not been examined. Therefore, we prepared a crude extract of AC PS for testing the effect on DCs, which play a critical role in initiating immune responses. The crude AC PSs promoted the production of TNF-, IL-6, and IL-12 by DCs, which is a hallmark for DC activation, in a dose-dependent manner (Fig. 1A). The results suggested that AC PSs may have stimulatory effect on DCs.
To verify the major PS compositions responding to DC activation, we further purified thePSs from crude extract according to molecular weight as shown in Table 1. Six fractions were obtained and their effect on DC activation was analyzed. Obviously, the fraction contained hmwPSs (> 100 kDa) had the strongest induction of TNF- and IL-12 produced by DCs (Fig. 1B). Thus, we identified that hmwPSs were very likely the major components in the total PSs of AC for DC activation.
To confirm that the stimulatory effect of crude AC PSs on DCs was mainly contributed by AC hmwPSs, we determined the ability of AC hmwPSs to induce cytokine production by DCs as did in Fig. 1A. We found that AC hmwPSs also augmented the secretion of TNF-, IL-6, and IL-12 by DCs in a dose-dependent manner (Fig. 1C), and the saturated dose (about 10 g/mL) was less than the dose of crude AC PSs (about 20-40 g/mL). At mean time, we excluded the possible endotoxin contamination because polymyxin B did not significantly reduce the activity of AC hmwPSs compared to the inactivation of LPS (Supplementary Fig. 1). These data clearly indicated that the AC hmwPSs can activate DCs and may be a potent immunostimulator.
AC hmwPSs promoted DC maturation
Maturation is a critical process for DC function to initiate adaptive immunity. Therefore, we evaluated the effect of AC hmwPSs on DC maturation. Upon AC hmwPS treatment, the expression levels of costimulatory molecules (CD40, CD80, and CD86) and MHCs (class I and II) were increased in DCs, indicating that DC became mature (Fig. 2). These results showed that the AC hmwPSs promote DC maturation.
AC hmwPSs facilitated DC-induced T cell activation and Th1 differentiation Activation of naive T cells is initiated by mature DC, so we examined whether AC
hmwPSs affect the ability of DCs to induce T cell activation. DCs were treated with PBS or AC hmwPSs and loaded with OVA257-264 or OVA323-339 peptide, and then co-cultured with OT-I or OT-II T cells. As shown in Fig. 3A, AC hmwPSs-stimulated DCs increased T cell proliferation compared to control DCs. In addition, we also measured the IFN- production by these activated T cells, a hallmark for Th1 differentiation. Significantly, AC hmwPSs-treated DCs induced higher amount of IFN- secreted by T cells than control DCs (Fig. 3B). Our data illustrated that AC hmwPSs augment the ability of DCs to activate Ag-specific T cells and Th1 polarization. This conclusion is in agreement with the enhancement of DC activation and maturation after AC hmwPSs treatment (Figs. 1 and 2).
Activation of MAPKs and NF-B by AC hmwPSs in DCs
We have proved the stimulatory activity of AC hmwPSs in DCs. Next, we explored the molecular mechanism involved in the DC activation by AC hmwPSs. MAPK and NF-B
pathways have been identified to be critical in mediating activation signals in DCs. Therefore, we checked the status of MAPK and NF-B in DCs after treatment of AC hmwPSs. As shown in Fig. 4, AC hmwPSs induced the phosphorylation of MAPKs (ERK, p38, and JNK) (Fig. 4A) and the activation of NF-B p65 (Fig. 4B), indicating that both pathways were activated by AC hmwPSs resulting to DC activation and maturation.
TLR2 and TLR4 were required in DC activation by AC hmwPSs
The MAPK and NF-B pathways can be triggered by many receptors on DC surface. TLRs are the most important receptor to sense microbes in environment. TLR2 and TLR4 have been revealed to participate in the DC activation by several sources of PSs. Thus, we asked whether these two receptors also participate in the stimulation of AC hmwPSs on DCs. We generated BMDCs from TLR2-/- and TLR4 mutant mice and then stimulated with AC hmwPSs. Comparing to DCs derived from wild type mice, TLR4 mutant DCs secreted definitely less IL-6 and IL-12 after AC hmwPS treatment (Fig. 5A), while the reduction of cytokines in TLR2-/- DCs was not as much as that in TLR4 mutant DCs but still significant (Fig. 5B). According to the results, we concluded that AC hmwPSs activate DCs via both TLR2 and TLR4 recognition.
AC hmwPSs strengthened the therapeutic effect of HER-2/neu DNA vaccine in mouse tumor model
We have demonstrated the promoting effect of AC hmwPSs on DC function. In previous studies we have established that the naked HER-2/neu DNA vaccine delivered via gene gun can be used for therapy in p185neu-expressing MBT-2 tumors in C3H/HeN mice . Therefore, we tested the possibility to use AC hmwPSs as adjuvant in this mouse DNA vaccine model. As
shown in Fig. 6A, when we immunized the tumor-implanted mice with the control vector alone or AC hmwPSs alone the tumors continuously grew in mice, and then all mice have to be sacrificed due to the size of their tumors. In contrast, vaccination with the HER-2/neu DNA alone or HER-2/neu DNA plus AC hmwPSs (5 or 10 g) inhibited tumor growth after tumor implantation. Significantly, the HER-2/neu DNA vaccine plus AC hmwPSs reduced more tumor sizes and prolonged the survival of the mice than that with the HER-2/neu DNA vaccine alone (Fig. 6B). These data strongly illustrated that AC hmwPSs enhance the therapeutic efficacy of HER-2/neu DNA vaccine against p185neu-expressing MBT-2 tumors in vivo.
AC hmwPSs polarized the Th1 responses induced by HER-2/neu DNA vaccination
In Fig. 3, we have found that DCs activated by AC hmwPSs increased T cell proliferation and preferentially skewed Th1 differentiation in vitro. Since Th1 response is necessary for tumor clearing, we then examined whether AC hmwPSs also augmented Th1 responses induced by HER-2/neu DNA vaccination in vivo. Total lymph node cells isolated from mice with
various treatments were incubated with recombinant HER-2/neu protein, and then detected IFN- production by CD8+ T cells using intracellular staining. Definitely, more CD8+ T cells from mice immunized with HER-2/neu DNA vaccine plus AC hmwPSs secreted IFN- than cells from immunized with HER-2/neu DNA vaccine alone (Fig. 7A). Furthermore, we determined the expression of IFN- (Th1) and IL-4 (Th2) in CD4+ T cells by quantitative real-time PCR.
Consistent to the results in CD8+ T cells, the vaccination with HER-2/neu DNA vaccine plus AC hmwPSs induced higher generation of IFN- in CD4+ T cells than that with control vector, AC hmwPSs alone, or HER-2/neu DNA vaccine alone (Fig. 7B). In contrast, no IL-4 production could be detected in all tested cells. These data pointed out that AC hmwPSs facilitate
HER-2/neu-specific Th1 responses induced by HER-2/neu DNA vaccination and then enhance the anti-tumor efficacy of HER-2/neu DNA vaccine.
Discussion
In this study, we characterized the activity of AC on the immune function of DCs. We discovered the promoting effect of AC PS on DC activation. Furthermore, we identified that a high molecular weight fraction of AC PS majorly contributed to the induction of DC activation and function. Thus, we report here that AC PS has immunostimulatory activity on DCs and that AC hmwPSs may be a potent adjuvant for the first time.
AC is an endemic mushroom in Taiwan. Its crude extracts and pure compounds, such as polysaccharides, terpenoids, proteins, and phenol derivatives, possess a variety of
health-promoting functions, including anti-hypertension, anti-inflammation, anti-oxidation, anti-cancer, and immunomodulation . According to literature, the majority of AC studies emphasize the inhibitory effect of AC on inflammation and tumor, especially the effect of small compounds isolated from AC . Thus, our study about the immune-enhancing effect of AC PS uniquely promotes the knowledge in AC research. AC PS can now be added as a new immunostimulator for DCs.
The biological activities of AC PS have been revealed, including anti-hepatitis B virus , anti-tumor , anti-angiogenesis , protection from hepatic injury , and anti-inflammation . In addition, AC PS has been shown to modulate the immune system for inhibiting infection of Schistosoma . In these reports the researchers use total AC PSs for their studies. Interestingly, Kuo et al. demonstrated that the immunomodulatory effect of AC mycelia is mainly attributed to the 10-20 kDa polysaccharides , which is much smaller than the active hmwPSs in our study. The difference may be due to different type of immune cells for test - they took the diluted peripheral blood with mainly polymorphonuclear neutrophils (PMN) but we used pure DCs. However, Yang et al. showed that only the AC PSs with high molecular weight ( > 100 kDa)
increase the IL-12 production by mononuclear cells and have anti-angiogenic effect . Actually, the immunostimulatory activity of hmwPSs has also been reported in other natural sources , in agreement with our data here.
About the DC research in AC, we have previously reported that a triterpenoid isolated from AC promotes DC activation and Th2 differentiation . This is the first study to examine the effect of AC on DC. Although we have previously shown that AC can enhance the anti-tumor efficacy of a DNA vaccine in mouse tumor model, we only used crude extract and did not check the effect of AC on pure DCs . In another work, Liu et al. also checked the regulatory role of AC PSs on DCs, but they observed that AC PS treatment on LPS-activated DCs enhanced IL-10 production and suppressed naive CD4 T-cell proliferation in an allogenic mixed lymphocyte reaction . It is possible that our AC hmwPSs may be different from their PSs.
AC hmwPSs activated the NF-κB and MAPK pathways (Fig. 4), that may be the mechanism by which it enhances DC function. A number of reports have shown this unique ability of PS . However, other mechanisms, such as PI3 kinase and PKC, have also been reported to involve in immunomodulation of mushroom polysaccharides . It is interesting to further investigate whether these pathways are also related to the DC activation by AC hmwPSs. To initiate these signaling events, we identified that TLR2 and TLR4 were required for the stimulatory effect of AC hmwPSs, but TLR4 might play a more important role than TLR2 (Fig. 5). Consistent to our finding, several types of PS can also activate immune cells via TLR2 and TLR4 . Thus, AC hmwPSs are likely a new ligand of TLRs.
In vaccine development, DNA vaccines have undergone a number of technological improvements and become attractive and promising in the field, especially for therapeutic
vaccination against intracellular pathogens and cancer . PSs have been used as adjuvant in DNA vaccine against infectious diseases, such as HBV , Trypanosoma cruzi , Chlamydophila abortus , and HIV. Notably, we reported here for the first time that AC hmwPSs significantly enhance the effect of HER-2/neu DNA vaccine against p185neu-expressing MBT-2 tumors in vivo (Fig. 6 and 7), suggesting the application of AC hmwPS in therapeutic vaccine against cancer. In addition, our data also imply the benefit of AC PS in immunotherapy for HER-2-expressing breast cancer . That will be valuable to test the adjuvantisity of AC hmwPSs in other types of vaccine, such as DC vaccines for cancer therapy.
In conclusion, AC hmwPSs have immunostimulatory activity on DCs. Our results emphasize again that AC can serve as an important biofactory because it contains various bioactive components for health promotion. Moreover, we provide the evidence that AC hmwPSs may have potential to be developed into adjuvant in immunology and vaccinology. Recently, polysaccharides have been applied as polymer adjuvants in vaccination because of the biocompatibility, biodegradability, and non-toxic properties, making them attractive candidates for substituting conventional adjuvants . This concept may speed the clinical application of AC PS in human and then enhance the efficacy of vaccine against infectious diseases and cancer.
Acknowledgments
We thank for technical help. This work was supported by (for C.C.L.), (for C.L.C.). The authors declare no financial or commercial conflict of interest.
References
1. El Enshasy HA, Hatti-Kaul R (2013) Mushroom immunomodulators: unique molecules with unlimited applications. Trends Biotechnol 31: 668-677.
2. Mizuno M, Nishitani Y (2013) Immunomodulating compounds in Basidiomycetes. J Clin Biochem Nutr 52: 202-207.
3. Yue PY, Wong YY, Chan TY, Law CK, Tsoi YK, et al. (2012) Review of biological and pharmacological activities of the endemic Taiwanese bitter medicinal mushroom,
Antrodia camphorata (M. Zang et C. H. Su) Sh. H. Wu et al. (higher Basidiomycetes). Int J Med Mushrooms 14: 241-256.
4. Chen YF, Lu WL, Wu MD, Yuan GF (2013) Analysis of Taiwan patents for the medicinal mushroom "Niu-Chang- Chih". Recent Pat Food Nutr Agric 5: 62-69.
5. Ren L, Perera C, Hemar Y (2012) Antitumor activity of mushroom polysaccharides: a review. Food Funct 3: 1118-1130.
6. Ramberg JE, Nelson ED, Sinnott RA (2010) Immunomodulatory dietary polysaccharides: a systematic review of the literature. Nutr J 9: 54.
7. Hu DJ, Cheong KL, Zhao J, Li SP (2013) Chromatography in characterization of polysaccharides from medicinal plants and fungi. J Sep Sci 36: 1-19.
8. Steinman RM (2012) Decisions about dendritic cells: past, present, and future. Annu Rev Immunol 30: 1-22.
9. Merad M, Sathe P, Helft J, Miller J, Mortha A (2013) The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol 31: 563-604.
10. Akira S (2011) Innate immunity and adjuvants. Philos Trans R Soc Lond B Biol Sci 366: 2748-2755.
11. Palucka K, Banchereau J (2013) Dendritic-cell-based therapeutic cancer vaccines. Immunity 39: 38-48.
12. Freitas-Silva R, Brelaz-de-Castro MC, Pereira VR (2014) Dendritic Cell-Based Approaches in the Fight Against Diseases. Front Immunol 5: 78.
13. Huang RY, Yu YL, Cheng WC, OuYang CN, Fu E, et al. (2010) Immunosuppressive effect of quercetin on dendritic cell activation and function. J Immunol 184: 6815-6821. 14. Yu YL, Chen IH, Shen KY, Huang RY, Wang WR, et al. (2009) A triterpenoid methyl
antcinate K isolated from Antrodia cinnamomea promotes dendritic cell activation and Th2 differentiation. Eur J Immunol 39: 2482-2491.
15. Lin MK, Yu YL, Chen KC, Chang WT, Lee MS, et al. (2011) Kaempferol from Semen cuscutae attenuates the immune function of dendritic cells. Immunobiology 216: 1103-1109.
16. Lin CC, Yu YL, Shih CC, Liu KJ, Ou KL, et al. (2011) A novel adjuvant Ling Zhi-8 enhances the efficacy of DNA cancer vaccine by activating dendritic cells. Cancer Immunol Immunother 60: 1019-1027.
17. Chen MH, Li WS, Lue YS, Chu CL, Pan IH, et al. (2013) Clitocybe nuda Activates Dendritic Cells and Acts as a DNA Vaccine Adjuvant. Evid Based Complement Alternat Med 2013: 761454.
18. Chu CL, Chen Dz C, Lin CC (2011) A novel adjuvant Ling Zhi-8 for cancer DNA vaccines. Hum Vaccin 7: 1161-1164.
19. Lu MC, El-Shazly M, Wu TY, Du YC, Chang TT, et al. (2013) Recent research and development of Antrodia cinnamomea. Pharmacol Ther 139: 124-156.
20. Lee IH, Huang RL, Chen CT, Chen HC, Hsu WC, et al. (2002) Antrodia camphorata polysaccharides exhibit anti-hepatitis B virus effects. FEMS Microbiol Lett 209: 63-67. 21. Liu JJ, Huang TS, Hsu ML, Chen CC, Lin WS, et al. (2004) Antitumor effects of the
partially purified polysaccharides from Antrodia camphorata and the mechanism of its action. Toxicol Appl Pharmacol 201: 186-193.
22. Chen SC, Lu MK, Cheng JJ, Wang DL (2005) Antiangiogenic activities of polysaccharides isolated from medicinal fungi. FEMS Microbiol Lett 249: 247-254.
23. Cheng JJ, Huang NK, Chang TT, Wang DL, Lu MK (2005) Study for anti-angiogenic activities of polysaccharides isolated from Antrodia cinnamomea in endothelial cells. Life Sci 76: 3029-3042.
24. Yang CM, Zhou YJ, Wang RJ, Hu ML (2009) Anti-angiogenic effects and mechanisms of polysaccharides from Antrodia cinnamomea with different molecular weights. J Ethnopharmacol 123: 407-412.
25. Han HF, Nakamura N, Zuo F, Hirakawa A, Yokozawa T, et al. (2006) Protective effects of a neutral polysaccharide isolated from the mycelium of Antrodia cinnamomea on
Propionibacterium acnes and lipopolysaccharide induced hepatic injury in mice. Chem Pharm Bull (Tokyo) 54: 496-500.
26. Chen CC, Liu YW, Ker YB, Wu YY, Lai EY, et al. (2007) Chemical characterization and anti-inflammatory effect of polysaccharides fractionated from submerge-cultured Antrodia camphorata mycelia. J Agric Food Chem 55: 5007-5012.
27. Wu YY, Chen CC, Chyau CC, Chung SY, Liu YW (2007) Modulation of inflammation-related genes of polysaccharides fractionated from mycelia of medicinal basidiomycete Antrodia camphorata. Acta Pharmacol Sin 28: 258-267.
28. Meng LM, Pai MH, Liu JJ, Yeh SL (2012) Polysaccharides from extracts of Antrodia camphorata mycelia and fruiting bodies modulate inflammatory mediator expression in mice with polymicrobial sepsis. Nutrition 28: 942-949.
29. Chen YJ, Cheng PC, Lin CN, Liao HF, Chen YY, et al. (2008) Polysaccharides from Antrodia camphorata mycelia extracts possess immunomodulatory activity and inhibits infection of Schistosoma mansoni. Int Immunopharmacol 8: 458-467.
30. Cheng PC, Hsu CY, Chen CC, Lee KM (2008) In vivo immunomodulatory effects of Antrodia camphorata polysaccharides in a T1/T2 doubly transgenic mouse model for inhibiting infection of Schistosoma mansoni. Toxicol Appl Pharmacol 227: 291-298. 31. Kuo MC, Chang CY, Cheng TL, Wu MJ (2008) Immunomodulatory effect of Antrodia
camphorata mycelia and culture filtrate. J Ethnopharmacol 120: 196-203.
32. Lobner M, Walsted A, Larsen R, Bendtzen K, Nielsen CH (2008) Enhancement of human adaptive immune responses by administration of a high-molecular-weight polysaccharide extract from the cyanobacterium Arthrospira platensis. J Med Food 11: 313-322.
33. Ishijima Y, Kawamura T, Kimura A, Kohno A, Okada T, et al. (2011) Toll-like receptor 4-dependent adjuvant activity of Kakkon-to extract exists in the high molecular weight polysaccharide fraction. Int J Immunopathol Pharmacol 24: 43-54.
34. Lemmon HR, Sham J, Chau LA, Madrenas J (2012) High molecular weight polysaccharides are key immunomodulators in North American ginseng extracts: characterization of the ginseng genetic signature in primary human immune cells. J Ethnopharmacol 142: 1-13. 35. Jang JY, Moon SY, Joo HG (2014) Differential effects of fucoidans with low and high
molecular weight on the viability and function of spleen cells. Food Chem Toxicol. 36. Huang CH, Chang CC, Lin CM, Wang ST, Wu MT, et al. (2010) Promoting effect of
Antrodia camphorata as an immunomodulating adjuvant on the antitumor efficacy of HER-2/neu DNA vaccine. Cancer Immunol Immunother 59: 1259-1272.
37. Liu KJ, Leu SJ, Su CH, Chiang BL, Chen YL, et al. (2010) Administration of
polysaccharides from Antrodia camphorata modulates dendritic cell function and alleviates allergen-induced T helper type 2 responses in a mouse model of asthma. Immunology 129: 351-362.
38. Xu Z, Chen X, Zhong Z, Chen L, Wang Y (2011) Ganoderma lucidum polysaccharides: immunomodulation and potential anti-tumor activities. Am J Chin Med 39: 15-27. 39. Lin KI, Kao YY, Kuo HK, Yang WB, Chou A, et al. (2006) Reishi polysaccharides induce
immunoglobulin production through the TLR4/TLR2-mediated induction of transcription factor Blimp-1. J Biol Chem 281: 24111-24123.
40. Figueiredo RT, Bittencourt VC, Lopes LC, Sassaki G, Barreto-Bergter E (2012) Toll-like receptors (TLR2 and TLR4) recognize polysaccharides of Pseudallescheria boydii cell wall. Carbohydr Res 356: 260-264.
41. Zhang X, Ding R, Zhou Y, Zhu R, Liu W, et al. (2013) Toll-like receptor 2 and Toll-like receptor 4-dependent activation of B cells by a polysaccharide from marine fungus Phoma herbarum YS4108. PLoS One 8: e60781.
42. Khan KH (2013) DNA vaccines: roles against diseases. Germs 3: 26-35.
43. Chen L, Shao HJ, Su YB (2004) Coimmunization of Agaricus blazei Murill extract with hepatitis B virus core protein through DNA vaccine enhances cellular and humoral immune responses. Int Immunopharmacol 4: 403-409.
44. Du X, Zhao B, Li J, Cao X, Diao M, et al. (2012) Astragalus polysaccharides enhance immune responses of HBV DNA vaccination via promoting the dendritic cell maturation and suppressing Treg frequency in mice. Int Immunopharmacol 14: 463-470.
45. Mussalem JS, Vasconcelos JR, Squaiella CC, Ananias RZ, Braga EG, et al. (2006) Adjuvant effect of the Propionibacterium acnes and its purified soluble polysaccharide on the immunization with plasmidial DNA containing a Trypanosoma cruzi gene. Microbiol Immunol 50: 253-263.
46. Ling Y, Li S, Yang J, Yuan J, He C (2011) Co-administration of the polysaccharide of Lycium barbarum with DNA vaccine of Chlamydophila abortus augments protection. Immunol Invest 40: 1-13.
47. Sun J, Hou J, Li D, Liu Y, Hu N, et al. (2013) Enhancement of HIV-1 DNA vaccine immunogenicity by BCG-PSN, a novel adjuvant. Vaccine 31: 472-479.
48. Milani A, Sangiolo D, Montemurro F, Aglietta M, Valabrega G (2013) Active immunotherapy in HER2 overexpressing breast cancer: current status and future perspectives. Ann Oncol 24: 1740-1748.
49. Shakya AK, Nandakumar KS (2013) Applications of polymeric adjuvants in studying autoimmune responses and vaccination against infectious diseases. J R Soc Interface 10: 20120536.
Table 1.
MW
(Da)
Water
insoluble
>100K
100K~50K 50K~30K 30K~10K 10K~5K
<5K
Weight
(mg)
96.3
915.9
308.0
212.9
155.7
47.3
229.9
%
4.90
46.59
15.67
10.83
7.92
2.40
11.69
Figure legends
Figure 1. The hmwPSs were major fraction of AC for DC activation. DCs were treated with PBS (control), LPS (100 ng/mL), or various doses of AC PSs as indicated. Supernatants were collected after 24 h (6 h for TNF-α). The amounts of TNF-α, IL-6, and IL-12 were determined by ELISA. (A) The effect of crude AC PSs in the first water extraction. (B) The effect of each fraction of AC PSs with different molecular weights. (C) The effect of hmwPS fraction. Data shown are the mean + SD of three samples. NSp>0.05; *p<0.05; **p<0.01; ***p<0.001 (Student's t-test) are comparisons between LPS- or AC PS-treated and PBS-treated DCs. All results are representative of three to four independent experiments.
Figure 2. AC hmwPSs promoted DC maturation. DCs were treated with PBS (control) or AC hmwPSs (5 or 10 g/mL) for 16 h. The expressions of MHC class I & II, CD40, CD80, and CD86 (gray-filled area) were determined by immunostaining and flow cytometry. All data shown were gated on CD11c+ cells. The black line represents staining with an isotype-matched control antibody. The level of expression is indicated as mean fluorescence intensity (MFI) in each graph. All results are representative of three to four independent experiments.
Figure 3. AC hmwPSs facilitated DC-induced T cell activation and Th1 differentiation.
OT-I or -II T cells were co-cultured with PBS- (white bar) or AC hmwPS-treated (10 g/mL, black bar) DCs pulsed with OVA257-264 (for OT-I) or OVA323-339 (For OT-II) peptides at the indicated ratio of DC:T cell. (A) T cell proliferation was determined by [3H]thymidine
incorporation after 3 days. (B) Supernatants were collected from the cultures after 4 days. IFN- production was measured by ELISA. Data are mean + SD of three samples. NSp>0.05;
***p<0.001 (Student's t-test) are comparisons between AC hmwPS-treated and PBS-treated DCs. All results are representative of three independent experiments.
Figure 4. Activation of MAPKs and NF-B by AC hmwPSs in DCs. DCs were collected and treated with AC hmwPSs (10 �g/mL), and then added lysis buffer at indicated time points to obtain whole cell lysates or nuclear extracts. (A) The activation of MAPKs was assayed by western blot. Whole cell lysates were incubated with antibodies against phosphorylated protein of ERK, JNK, and p38 MAPK, and then detected with specific 2nd antibodies and ECL system. The loading controls were shown by using antibodies against total protein of these MAPKs. Data are representative of three independent experiments. (B) The activation of NF-�B was determined by p65 binding assay. The NF-�B binding activity in nuclear extract was measured using TransAM NF-�B p65 ELISA kit and shown as value read at OD450. Data are mean + SD of three samples. NSp>0.05; ***p<0.001 (Student's t-test) are comparisons between LPS- or AC hmwPS-treated and PBS-treated DCs. All results are representative of three independent experiments.
Figure 5. TLR2 and TLR4 were involved in DC activation by AC hmwPSs. DCs were generated from (A) C3H/HeN and C3H/HeJ (TLR4 mutant), or (B) C57BL/6 and TLR-2 KO mice, and then treated with PBS (control), LPS (100 ng/mL), LTA (1 �g/mL), or AC hmwPSs (10 �g/mL). Supernatants were collected after 24 h. The amounts of IL-6 and IL-12 were determined by ELISA. Data shown are the mean + SD of three samples. NSp>0.05; *p<0.05; ***p<0.001 (Student's t-test) are comparisons between LPS-, LTA- or AC hmwPS-treated and PBS-treated DCs. All results are representative of three independent experiments.
Figure 6. AC hmwPSs enhanced the therapeutic effect of HER-2/neu DNA vaccine in mouse tumor model. The p185neu-expressing MTB-2 cells were inoculated in mice as
described in Materials and Methods section. After ten days, the tumor-bearing mice were treated with AC hmwPSs, HER-2/neu DNA vaccine, or the combination as indicated. Data shown are the mean + SD of six to seven mice per group. (A) Tumor growth curve in mice after
vaccination. The tumor volumes were calculated as described in Materials and Methods section at the indicated day. (B) The Kaplan-Meier survival curve of mice (N = 5) after vaccination. All results are representative of three independent experiments.
Figure 7. AC hmwPSs polarized the Th1 responses induced by HER-2/neu DNA
vaccination. The tumor-bearing mice were treated with AC PSs, HER-2/neu DNA vaccine, or the combination as indicated for 6 days. Then, total inguinal LN cells were isolated from various vaccinated mice and incubated with recombinant HER-2/neu protein (10 g/mL) for 16 h. (A) The production of IFN- by stimulated CD8+ T cells was detected by intracellular staining and flow cytometry. The percentage of CD8+IFN-+ cells in total CD8+ T cells was shown in each graph. (B) CD4+ T cells were purified from the total LN cells. The production of IFN- and IL-4 by stimulated CDIL-4+ T cells were determined by quantitative real-time PCR. The data are normalized to HPRT expression in each sample and shown as the mean + SD of five mice. NSp>0.05; *p<0.05 (Student's t-test) are comparisons of treatment between HER-2/neu DNA vaccine and control. All data are representative of three independent experiments.