40
Bufalin induces G
0/G
1phase arrest through
inhibiting the levels of cyclin D, cyclin E,
CDK2 and CDK4, and triggers
apoptosis
via
mitochondrial signaling
pathway in T24 human bladder cancer
cells
Wen-Wen Huanga , 1, Jai-Sing Yangb , 1, Shu-Jen Paia , Ping-Ping Wuc , Shu-Jen Changc , Fu-Shin Chuehd , Ming-Jen Fane , Shang-Ming Chiouf , g, Hsiu-Maan Kuoh , Chin-Chung Yehi , Po-Yuan Chena , Minoru Tsuzuki j, k, Jing-Gung Chunga , e, , a Department of Biological Science and Technology, China Medical University, Taichung 404, Taiwan b Department of Pharmacology, China Medical University, Taichung 404, Taiwan
c School of Pharmacy, China Medical University, Taichung 404, Taiwan
d Department of Health and Nutrition Biotechnology, Asia University, Taichung 413, Taiwan e Department of Biotechnology, Asia University, Taichung 413, Taiwan
f Department of Functional Neurosurgery & Gamma Knife Center, China Medical University Hospital, Taichung 404, Taiwan
g School of Medicine, China Medical University, Taichung 404, Taiwan h Department of Parasitology, China Medical University, Taichung 404, Taiwan i Department of Urology, China Medical University Hospital, Taichung 404, Taiwan j Department of Biochemistry, Nihon Pharmaceutical University, Saitama 362-0806, Japan k Tsuzuki Institute for Traditional Medicine, China Medical University, Taichung 404, Taiwan
Abstract
Most of the chemotherapy treatments for bladder cancer aim to kill the cancer cells, but a high recurrence rate after medical treatments is still occurred. Bufalin from the skin and parotid venom glands of toad has been shown to induce apoptotic cell death in many types of cancer cell lines. However, there is no report addressing that bufalin induced cell death in human bladder cancer cells. The purpose of this study was investigated the mechanisms of bufalin-induced apoptosis in a human bladder cancer cell line (T24). We demonstrated the effects of bufalin on the cell growth and apoptosis in T24 cells by using DAPI/TUNEL double staining, a PI exclusion and flow cytometric analysis. The effects of bufalin on the production of reactive oxygen species (ROS), the level of mitochondrial
membrane potential (ΔΨm), and DNA content including sub-G1 (apoptosis) in T24 cells were also
determined by flow cytometry. Western blot analysis was used to examine the expression of
G0/G1 phase-regulated and apoptosis-associated protein levels in bufalin-treated T24 cells. The results
indicated that bufalin significantly decreased the percentage of viability, induced the G0/G1 phase
arrest and triggered apoptosis in T24 cells. The down-regulation of the protein levels for cyclin D, CDK4, cyclin E, CDK2, phospho-Rb, phospho-AKT and Bcl-2 with the simultaneous up-regulation of the cytochrome c, Apaf-1, AIF, caspase-3, -7 and -9 and Bax protein expressions and caspase activities were observed in T24 cells after bufalin treatment. Based on our results, bufalin induces apoptotic cell death in T24 cells through suppressing AKT activity and anti-apoptotic Bcl-2 protein as well as inducing pro-apoptotic Bax protein. The levels of caspase-3, -7 and -9 are also mediated apoptosis in bufalin-treated T24 cells. Therefore, bufalin might be used as a therapeutic agent for the treatment of human bladder cancer in the future.
Highlights
► Suppression of AKT activity and anti-apoptotic Bcl-2 protein level in bufalin-treated T24 cells. ► Stimulations of Bax signal and caspases- and mitochondria-dependent pathway in T24 cells after
bufalin exposure. ► Inductions of G0/G1 phase arrest and apoptotic death of T24 cells.
Abbreviations
AIF, apoptosis-inducing factor;
CDK, cyclin-dependent kinase;
CsA, cyclosporine A;
DCFH-DA, 2′-7′-dichlorfluorescein-diacetate;
DiOC6, 3,3′-dihexyloxacarbocyanine iodide;
DMSO, dimethyl sulfoxide;
ECL,enzyme chemiluminescence;
HRP, horseradish peroxidase;
NAC, N-acetyl-cysteine;
PBS, phosphate-buffered saline;
PI, propidium iodide;
SDS, sodium dodecyl sulfate;
TUNEL,terminal deoxynucleotidyl transferase (TdT)-mediated d-UTP nick end-labeling;
z-VAD-fmk, z-Val-Ala-Asp-fluoromethyl ketone (pan-caspase inhibitor);
z-LEHD-fmk, z-Leu-Glu-His-Asp-fluoromethyl ketone (caspase-9 inhibitor)
Keywords
Bufalin;
T24 human bladder cancer cells;
G0/G1 phase arrest;
Apoptosis;
Mitochondrial signaling pathway
1. Introduction
Bladder cancer is the fifth most common cancer among men and women in the USA in 2008 [1]. In
Taiwan, 3.3 persons per 100 thousand die annually from bladder cancer based on the report in 2009 from the Department of Health, R.O.C. (Taiwan). The major treatments for bladder cancer patients are surgery, radiotherapy and chemotherapy, or combine with radiotherapy and chemotherapy, but the efficiency of cure rates are not satisfactory. Therefore, searching for chemoprevention or chemical controls for bladder cancer has become a crucial concern.
Bufalin (Fig. 1) is the major digoxin-like immunoreactive component of Chan-Su extracts from the
venom ofBufo bufo gargarizan[2]. Chan-Su, a traditional Chinese medicine, was obtained from the skin
and parotid venom glands of the toad [3] and its extracts have been applied in the treatment of
various cancers in clinical trials in China [4]. Bufalin has been demonstrated to induce cell cycle arrest
and apoptosis in many human cancer cells including leukemia [5], [6], [7], [8] and [9], prostatic
cancer [2] and [10], endometrial and ovarian cancer [11] and osteosarcoma [12]. Besides, bufalin
induced autophagy in human colon cancer cells through promoted the reactive oxygen species (ROS)
generation and the c-Jun NH2-terminal kinase (JNK) signaling[13]. Bufalin has been shown to inhibit
cytochrome P450 3A4 (CYP3A4) in in vitro and in vivo effects and interacted the CYP3A4-metabolized
agent [14]. However, the effects of bufalin on bladder cancer cells have not yet been thoroughly
reported and knowledge of the molecular mechanisms of bufalin-induced apoptosis in bladder cancer cells was rudimentary and remained to be delineated. Therefore, the purpose of this study was designed to define the biological and therapeutic effects of bufalin-treated human bladder cancer cells
for the first time. We investigated whether or not bufalin was able to mediate growth inhibition of bladder cells, cell cycle arrest and induction of apoptosis in bladder cancer cells.
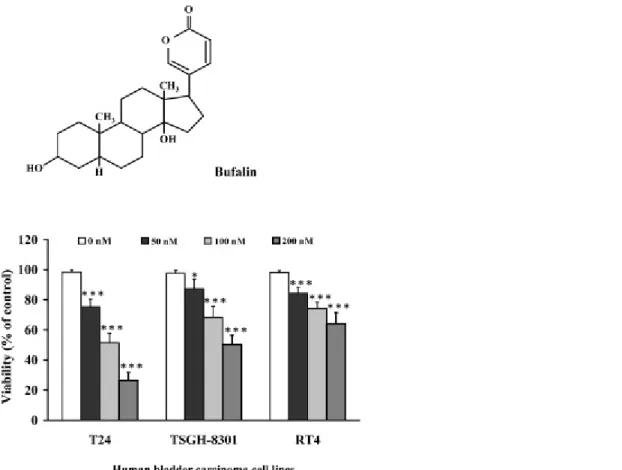
Fig. 1. Effects of bufalin on cell viability in human bladder cancer cells. (A) The chemical structure of bufalin. (B) Bladder cancer cells were treated with 0, 50, 100 and 200 nM of bufalin for 24 h then cell viability was determined by a PI exclusion method. Data were expressed mean ± SEM of three independent experiments. ***P < 0.001, significantly different compared with the control (0 nM) group.
Apoptosis, also known as programmed cell death type I, involves a cascade of molecular changes including loss of organelle trans-membrane potential, swelling of the matrix, and rupture of the outer membrane, DNA fragmentation, chromatin condensation, apoptotic body, and culminates with the
discharge of apoptotic proteins, most notably cytochrome c in the cytosol [15], [16] and [17]. The
anti-apoptotic Bcl-2 proteins regulating outer membrane integrity [18] of mitochondrial permeability
transition [19] have been intensely pursued for novel molecular therapeutics of some human cancer.
In this study, we demonstrated that bufalin inhibited the growth of T24 human bladder cancer cells
through G0/G1 phase arrest and the inhibitions of cyclin D and E, CDK2 and CDK4, and it induced
apoptosis through a mitochondria-dependent pathway. Our results indicated that a decrease of PKB (protein kinase B)/AKT activity and an increase in the pro-apoptotic Bax through dissociation from anti-apoptotic Bcl-2, leading to mitochondrial dysfunction, cytochrome c release, activation of caspase cascades and consequently apoptotic cell death in bufalin-treated T24 cells.
2. Materials and methods
2.1. Chemicals and reagents
Bufalin, dimethyl sulfoxide (DMSO), propidium iodide (PI), RNase A, Triton X-100, proteinase K, cyclosporine A (CsA: a mitochondrial membrane permeability transition inhibitor) and N-acetyl-cysteine (NAC: a ROS scavenger) were purchased from Sigma–Aldrich Corp. (St. Louis, MO, USA).
FCS, L-glutamine, penicillin-streptomycin and trypsin-EDTA were obtained from Invitrogen Life
Technologies (Carlsbad, CA, USA). The z-LEHD-fmk (caspase-9 inhibitor), z-VAD-fmk (a pan-caspase inhibitor) were purchased from R&D Systems (Minneapolis, MN, USA). Sources of antibodies used in this study were as follows: monoclonal antibodies specific for β-actin, cyclin D, CDK4, cyclin E, CDK2, Rb, phospho-Rb, cytochrome c, Apaf-1, AIF, AKT, Bax, Bcl-2 and all peroxidase-conjugated secondary antibodies were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Polyclonal antibodies specific for phospho-AKT, caspase-9, caspase-7 and caspase-3 were obtained from Cell Signaling Technology Inc. (Danvers, MA, USA). Enhanced chemiluminescence (ECL), a Western blot detection reagent, was purchased from Pierce Chemical (Rockford, IL, USA).
2.2. Cell culture
The human bladder cancer cell lines (T24, TSGH-8301 and RT4) were purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan). The cells were grown in McCoy's 5a medium
supplemented with 10% FCS, 2 mM L-glutamine, and 100 units/ml penicillin/100 μg/ml streptomycin at
37 °C under 5% CO2 in air.
2.3. Cell viability assay
Bladder cancer cell lines (2 × 105 cells/well) were individually maintained in 24-well plates with
McCoy's 5a medium for 24 h, and then treated with 0, 50, 100 and 200 nM of bufalin for 24 h at 37 °C,
5% CO2 and 95% air. For incubation with the specific inhibitors, cells seeded into 24-well plates were
pretreated with NAC (10 mM), CsA (5 μM), a pan-caspase inhibitor (z-VAD-fmk; 10 μM) and a caspase-9 inhibitor (z-LEHD-fmk) for 1 h, followed by treatment with or without 100 nM bufalin. Cells were harvested from individual well by centrifugation. For viability determination, cells from each treatment were stained with PI (5 μg/ml) and analyzed by flow cytometry (Becton-Dickinson, FACSCalibur, San
Jose, CA, USA) as previously described[20] and [21].
2.4. Cell morphology were examined by DAPI and TUNEL staining
Aliquots of T24 cells (2 × 105 cells/well) were placed into 24-well plates with McCoy's 5a medium and
then were exposed to 0, 50, 100 and 200 nM of bufalin for 24 h. Cells were examined and photographed under a phase-contrast microscope. Apoptosis was detected using the DAPI/TUNEL double staining method in situapoptosis detection kit (in situ cell death detection kit; Roche Diagnostics, Mannheim, Germany). T24 cells were treated with or without 100 nM bufalin. Cells were
fixed in 4% buffered formaldehyde then were mounted on glass slides. After being washed with PBS,
endogenous peroxidase was blocked by the addition of 3% H2O2. Cells were then treated with FITC-label
terminal deoxynucleotidyl transferase and biotinylated dUTP. After stopping the reaction, the samples were stained with 4′-6-diamidino-2-phenylindole (DAPI, Invitrogen). All cells were stained by DAPI and TUNEL then were examined and photographed by a fluorescence microscope as described
previously [22] and [23].
2.5. DNA content analysis for cell cycle distribution and sub-G1 group
Approximately 2 × 105 cells/well of T24 cells in 24-well plates were treated with 100 nM bufalin for 0, 6,
12 and 24 h. Cells were harvested and washed twice with cold PBS. Cells were fixed by using 70% ethanol at −20 ̊C overnight and washed twice with cold PBS, and then cells were re-suspended in PBS containing 40 μg/ml PI and 0.1 mg/ml RNase and 0.1% triton X-100 in dark room for 30 min at 37 °C. All samples were analyzed by flow cytometry and the cell cycle and sub-G1 (apoptosis) phase were
determined and analyzed as described previously [24] and [25].
2.6. Determination of reactive oxygen species (ROS) and mitochondrial membrane potential (ΔΨm)
Cells (2 × 105 cells/well) in 24-well plates were exposed to 100 nM bufalin and incubated for 0, 2, 4, 6
and 12 h. At the end of incubation, cells from each treatment were harvested by centrifugation and were washed twice by PBS, then were re-suspended in 2,7-dichlorodihydrofluorescein diacetate
(DCFH-DA; 10 μM, Invitrogen) for ROS determination and were re-suspended in DiOC6 (1 μM, Invitrogen) for
measurement of ΔΨm. Then cells were incubated at 37 °C under dark room for 30 min and were
analyzed immediately by flow cytometry as described previously [26] and [27].
2.7. Assays of caspase-3, caspase-7 and caspase-9 activity
Approximately 2 × 105 cells/well of T24 cells in 10 cm culture dish were treated with 100 nM bufalin
and incubated for 0 and 24 h, and then the activities of caspase-3, -7 and -9 were assessed according to manufacturer's instruction of Caspase colorimetric kit (R&D system Inc.). Cells were harvested and lysed in 50 μl lysis buffer which containing 2 mM DTT for 10 min. After centrifugation, the supernatant containing 200 μg protein were incubated with caspase-3, caspase-7 and caspase-9 substrate in reaction buffer. Then all samples were incubated in 96-well flat bottom microplate at 37 °C for 1 h. Levels of released pNA were measured with ELISA reader (Anthos Reader 2001, Anthos Labtec) at
405 nm wavelength [23] and [28].
2.8. Determinations of G0/G1 phase arrest and apoptosis-associated protein levels by Western blotting
T24 cells at a density of 1 × 106 cells in 75 T flasks were exposed to 100 nM bufalin and then incubated
for 0, 1, 2, 6, 12, 18 and 24 h for examining the protein levels correlated with cell cycle arrest and apoptosis. Cell were harvested from each treatment, washed with cold PBS, and lysed in the PRO-PREP™ protein extraction solution (iNtRON Biotechnology, Seongnam, Gyeonggi-Do, Korea). The total
proteins were collected before the levels of cyclin D and E, CDK4, CDK2, Rb, p-Rb, p-AKT, AKT, Bax and Bcl-2 were detected using immunoblotting. The total protein was collected before the cytochrome c, Apaf-1, AIF, caspase-9, caspase-3 and caspase-7 was detected by Western blotting. In brief, about 30 μg protein from each sample was resolved over 10% sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membrane. The blot was soaked in blocking buffer (5% non-fat dry mik/0.05% Tween 20 in 20 mM Tris buffered saline (TBS) at pH 7.6) at room temperature for 1 h then incubated with individual monoclonal antibodies in blocking buffer at 4 °C for overnight. Then followed by secondary antibody-conjugated horseradish peroxidase (HRP) and detected by chemiluminescence kit (Millipore, Bedford, MA, USA) and autoradiography using X-ray film
as described elsewhere [20],[22] and [29]. To ensure equal protein loading, each membrane was
stripped and reprobed with anti-β-actin antibody.
2.9. In vitro AKT kinase assay
This assay was followed as the protocol of the manufacturer's instructions from an AKT kinase assay kit
(Cell Signaling Technology, Beverly, MA, USA). Briefly, about 2 × 106 cells/well of T24 cells in 10 cm
dish were treated with 100 nM bufalin for 0, 1, 2 and 6 h. At the end of incubation, cells were harvested and lyzed using the AKT kinase assay kit and then 200 mg of protein from each time point treatment was immunoprecipitated with 2 mg of anti-AKT antibody overnight. Then all samples were extensive washing, the immunoprecipitates were incubated with 1 mg of glycogen synthase kinase-3 α/β (GSK-3 α/β) fusion protein substrate in 50 ml of kinase buffer for 30 min at 30 °C. Reactions were stop by SDS loading buffer. The samples were separated on 12% SDS-PAGE, and the phospho-GSK-3
α/β (Ser219) was detected by immunoblotting [30], [31] and [32].
2.10. Statistical analyses
Data are presented as the mean ± SEM for the indicated number of separate experiment. Statistical analyses of data were done by Student's t-test, and *P < 0.05, ***P < 0.001 were considered significant.
3. Results
3.1. Bufalin decreased the viability of human bladder cancer cells
We determined the growth inhibition effects of bufalin on the cell viability by using a PI exclusion and
flow cytometric assay, and the results are shown in Fig. 1B. Increasing the dose of bufalin led to the
decrease of the percentage of viability in examined 3 different bladder cancer cell lines (Fig. 1B).
Bufalin at 100 nM decreased by almost 45, 26 and 22% the viable cells of T24, TSGH-8301 and RT4, respectively, at 24 h treatment. The highest sensitive cell is T24 cells; therefore, we selected T24 cells for further experiments.
3.2. Bufalin induced morphological changes and apoptosis in T24 cells
T24 cells were treated with 0, 50, 100 and 200 nM bufalin for 24 h. As shown in Fig. 2A and B, bufalin
induced cell morphological changes and decreased the cells number, and cells became smaller, round
and blunt in size when compared with control in T24 cells. These effects are dose-dependent (Fig. 2A;
arrow). The assay demonstrated that bufalin induced DNA condensation and apoptosis which were
examined by DAPI/TUNEL double staining (Fig. 2B). Taken together, we concluded that 100 nM bufalin
decreased the percentage of viable T24 cells through the apoptotic cell death.
3.3. Bufalin induced cell cycle arrest and affected associated protein levels in T24 cells
Based on the results from growth inhibition, further studies were conducted to investigate the possible mechanisms which are involved in bufalin-induced cell cycle arrest and associated protein levels in T24 cellsin vitro. The results from flow cytometric assay revealed that 100 nM bufalin induced
accumulation of G0/G1phase in T24 cells and this effect is a time-dependent manner (Fig. 3A). Western
blotting also showed that bufalin down-regulated the expressions of cyclin D and E, CDK2 and CDK4
and p-Rb and up-regulated the expressions of Rb, leading to G0/G1 phase arrest in T24 cells and this
effect is a time-dependent response (Fig. 3B).
3.4. Bufalin induced reactive oxygen species (ROS) production and pre-treated with N-acetyl-cysteine (NAC) and a pan-caspase inhibitor (z-VAD-fmk) to increase the viability in treated T24 cells
To verify that ROS and caspase cascade are involved in bufalin-induced cell death of T24 cells. Cells were pretreated with 10 mM ROS scavenger (NAC) or 10 μM z-VAD-fmk and then exposed to 100 nM bufalin before being harvested for measuring the levels of ROS and cell viability by flow cytometric
assay. The results in Fig. 4A indicated that bufalin promoted the ROS production and this effect is a
time-dependent response. T24 cells were also measured the percentage of viability in T24 cells and
results are shown in Fig. 4B, which indicated that NAC and z-VAD-fmk can increased the percentage of
viable T24 cells after bufalin exposure. Based on these observations, bufalin-induced cytotoxic effects were mediated through ROS production and increased caspase activity in T24 cells.
3.5. Bufalin decreased the level of mitochondria membrane potential (ΔΨm) and pre-incubated with cyclosporine A (CsA) and caspase-9 inhibitor to protect against the viability in T24 cells after treatment
To investigate whether mitochondria are involved in bufalin-triggered cell death, T24 cells were pretreated with 5 μM CsA (a mitochondrial membrane permeability transition inhibitor) or 10 μM z-LEHD-fmk (a caspase-9 inhibitor). Cells then were treated with 100 nM bufalin. Cells were harvested
for examining of ΔΨmand viability. The results shown in Fig. 5A and B indicated that 100 nM bufalin
decreased the level of ΔΨm. Furthermore, cells were pretreated with CsA or z-LEHD-fmk and then
the bufalin-treated only cells (Fig. 5B). These results indicated that bufalin-induced cytotoxic effects were mediated through mitochondria-dependent apoptotic signaling pathways.
3.6. Bufalin increased the activities of caspase-3, -7 and -9 and affected the apoptosis-associated protein levels in T24 cells
To determine whether apoptosis is mediated via the activation of caspase-3, -7 and -9 in bufalin-treated T24 cells. Cells were harvested after exposure to 100 nM bufalin and then determined the activities of caspase-3, -7 and -9 by colorimetric assays. The changes of apoptosis-associated protein
levels were determined by Western blotting. The results shown in Fig. 6A indicated that bufalin
promoted the activation of caspase-3, -7 and -9 for a 24-h treatment. Results in Fig. 6B indicated
100 nM bufalin increased the levels of cytosolic cytochrome c, AIF, Apaf-1 and active form of caspase-3, -7 and -9. Our results suggest that bufalin-induced apoptosis is done through the mitochondria-dependent signaling pathway in T24 cells.
3.7. Bufalin inhibited the activity of AKT and affected the Bcl-2 family protein levels in T24 cells
To examine whether bufalin-induced apoptosis is through the inhibition of AKT and involved in Bcl-2 family protein levels in T24 cells. Cells were harvested after treatment with 100 nM bufalin, and then determined the AKT activity and Bcl-2 family-related protein levels by Western blotting. Our results in Fig. 7A revealed that bufalin decreased the AKT activity after bufalin for 2 and 6 h-treatment and this effect is time-dependently. Results from Western blotting also showed that bufalin decreased the levels
of p-AKT, AKT and Bcl-2, but it increased the level of Bax (Fig. 7B) in T24 cells. Based on these
observations, it is suggested that bufalin-induced apoptosis in T24 cells is mediated through the changes of ratio of Bax/Bcl-2 and a decrease in the activities of AKT.
4. Discussion
It was reported that bufalin induced cytotoxic effects in many human cancer cell lines through cell
cycle arrest and induction of apoptosis [2], [5], [6], [7], [8], [9], [10], [11] and [12]. In this study, we
first demonstrated that bufalin induced cytotoxic effects through G0/G1 arrest (Fig. 3A) and inducing
apoptosis in T24 cells. This is in agreement with the reports from Nasu et al. indicated that bufalin
inhibited the cell proliferation through induction of apoptosis and the G0/G1 phase arrest of the cell
cycle of endometriotic stromal cells in vitro[33]. We also used DAPI/TUNEL double staining to confirm
that bufalin induced apoptosis in T24 cells (Fig. 2A and B). Western blotting analysis indicated that
bufalin induced the down-regulation of cyclin D and cyclin E, CDK2 and CDK4 and p-Rb, but it increased the level of Rb in T24 cells. These regulations of cell cycle associated proteins indicated
bufalin induced G0/G1 phase arrest in T24 cells.
Our results showed that bufalin promoted the production of ROS in T24 cells and this effect is time
the viable T24 cells when compared to the bufalin-treated only cells (Fig. 4B). This observation indicated that ROS was involved in bufalin-induced cell death. This is in agreement with other report demonstrated that bufalin induced apoptosis via ROS-dependent mitochondrial death pathway in
human lung adenocarcinoma ASTC-α-1 cells [34]. Our results also showed in Fig. 5A indicated that
bufalin decreased the level of ΔΨmfrom T24 cells and this is also agreement with Sun et al. reported
that bufalin decreased the level of ΔΨmand mitochondria play an important role in bufalin-induced
apoptotic death in ASTC-α-1 cells [34].
It is well known that caspases can be activated in two major apoptotic pathways, the death-receptor and mitochondria-mediated signaling pathways. Bufalin promoted caspase-3, -7 and -9 in T24 cells (Fig. 6A). As shown in Fig. 6B, bufalin increased cytosolic protein levels of cytochrome c, Apaf-1, Pro-caspase-9 and AIF in T24 cells. This is also in agreement with other report showed that the activation of caspase-9, an initiator caspase closely coupled to pro-apoptotic signals, was observed after bufalin treatment, suggesting that caspase-9-mediated mitochondria-mediated signaling pathway is involved
in the mechanism of bufalin-induced apoptosis [11]. However, their report did not show that caspase-3
and -7 also involved in bufalin induced apoptosis. This is our novel finding mechanism of bufalin-induced apoptosis, which is involved in activations of caspase-9, -3 and -7 in T24 cells. Our result does not rule out the involvement of the death receptor apoptotic signaling pathway. The levels of Fas, FasL and FADD protein levels and caspase-8 activity have no significant influence on bufalin-treated T24 cells (data not shown). Our results suggest that the mitochondrial signaling pathway is mediated bufalin-induced apoptotic response in T24 cells.
Our results showed that bufalin induced the down-regulation of the expression of Bcl-2 (Fig. 7B), and
the simultaneous up-regulation of the Bax (Fig. 7B) and activated caspase-3, -7 and -9 expressions
(Fig. 6B) in T24 cells and this is in agreement with reports from Sun et al. in ASTC-α-1 cells after bufalin exposure (27). We determined caspase activity assay to confirm that bufalin promoted the activities of
caspase-3, -7 and -9 in T24 cells (Fig. 6A). This is also agreement with other report indicated that
activation of caspase-9 as observed after bufalin treatment, suggesting that caspase-9-mediated
cascade is involved in the mechanism of bufalin-induced apoptosis [35]. However, another report
showed that bufalin did not affect caspase-3 activity in ASTC-α-1 cells [34]. Therefore, it is suggested
that the cell-specific effects of bufalin on cancer cells such as the mechanisms of bufalin-induced apoptosis of human leukemia cells by the activation of AP-1 and the c-Jun N-terminal protein kinase
(JNK) [8], cdc2 kinase and casein kinase II [5] and [6], the induction of Tiam1 expression [9] and Bcl-2
and c-myc expression [7] and the inhibition of protein kinase A and C [5] and [6]. The interesting point
is that other report indicated bufalin induced G2/M phase arrest in leukemia cells [5] and [6]. Therefore,
we suggest that the effects of bufalin may be cell-type specific.
It was reported that phosphorylated Bax on Ser184 by AKT and then inhibition of conformational change
and inability of Bax to translocate to the mitochondrial membrane [36] and [37]. This action, then,
blocks the pore formation and inhibits the release of cytochrome c, Apaf-1, pro-caspase-9 and AIF proteins from mitochondrial into cytosol. It also reported that the phosphorylated Bax heterodimerizes
with Bcl-xl, the binding of Bcl-2 family members may prevent the translocation of Bax to the
mitochondrial membrane, and then inhibit apoptosis [38]. In this study, bufalin inhibited the activity of
AKT in T24 cells. It is well known that AKT is involved in cell survival or death dependent the associated signal pathway. Oka et al. investigated that the high expression of activated AKT was observed in T24
cells, whereas low expression of that was shown in RT4 cells [39]. We also demonstrated that
treatments of T24 cells with 100 nM bufalin reversed the high constitutive activity of AKT in comparison to those from TSGH-8301 and RT4 human bladder cancer cells (data not shown). It is suggested that more sensitivity in the PI3K inhibitors and AKT protein expression exhibited in T24 cells,
a highly malignant grade III human urinary bladder carcinoma [39]. In the present study, the AKT
activity may play an important role in regulating the Bcl-2 family protein levels to the induction of apoptosis in bufalin-treated T24 cells.
Overall, the outline of molecular signaling pathways is summarized in Fig. 8. These results indicated
that bufalin could be used as a novel therapeutic agent for the medical treatment and/or prevention of bladder cancer.
Conflict of interest statement
None.Acknowledgments
This study was supported by research grant CMU99-TC-05 from China Medical University, Taichung,
Taiwan. We also thank the National Science Council of the Republic of China for financial support (NSC
97-2320-B-039-004-MY3).
References
1.
o A.M. Kamat, D.L. Lamm
o Chemoprevention of urological cancer
o J. Urol., 161 (1999), pp. 1748–1760
2.
o C.H. Yu, S.F. Kan, H.F. Pu, E. Jea Chien, P.S. Wang
o Apoptotic signaling in bufalin- and cinobufagin-treated androgen-dependent and -inandrogen-dependent human prostate cancer cells
o Cancer Sci., 99 (2008), pp. 2467–2476
3.
o L. Krenn, B. Kopp
o Bufadienolides from animal and plant sources
o Phytochemistry, 48 (1998), pp. 1–29
o Z. Meng, P. Yang, Y. Shen, W. Bei, Y. Zhang, Y. Ge, R.A. Newman, L. Cohen, L. Liu, B. Thornton, D.Z. Chang, Z. Liao, R. Kurzrock
o Pilot study of huachansu in patients with hepatocellular carcinoma, nonsmall-cell lung cancer, or pancreatic cancer
o Cancer, 115 (2009), pp. 5309–5318
5.
o S. Numazawa, M.A. Shinoki, H. Ito, T. Yoshida, Y. Kuroiwa
o Involvement of Na+,K(+)-ATPase inhibition in K562 cell differentiation induced by bufalin
o J. Cell. Physiol., 160 (1994), pp. 113–120
6.
o Y. Jing, M. Watabe, S. Hashimoto, S. Nakajo, K. Nakaya
o Cell cycle arrest and protein kinase modulating effect of bufalin on human leukemia ML1 cells
o Anticancer Res., 14 (1994), pp. 1193–1198
7.
o Y. Masuda, N. Kawazoe, S. Nakajo, T. Yoshida, Y. Kuroiwa, K. Nakaya
o Bufalin induces apoptosis and influences the expression of apoptosis-related genes in human leukemia cells
o Leuk. Res., 19 (1995), pp. 549–556
8.
o M. Watabe, K. Ito, Y. Masuda, S. Nakajo, K. Nakaya
o Activation of AP-1 is required for bufalin-induced apoptosis in human leukemia U937 cells
o Oncogene, 16 (1998), pp. 779–787
9.
o N. Kawazoe, M. Watabe, Y. Masuda, S. Nakajo, K. Nakaya
o Tiam1 is involved in the regulation of bufalin-induced apoptosis in human leukemia cells
o Oncogene, 18 (1999), pp. 2413–2421
10.
o J.Y. Yeh, W.J. Huang, S.F. Kan, P.S. Wang
o Effects of bufalin and cinobufagin on the proliferation of androgen dependent and independent prostate cancer cells
o Prostate, 54 (2003), pp. 112–124
11.
o Bufalin induces growth inhibition, cell cycle arrest and apoptosis in human endometrial and ovarian cancer cells
o Int. J. Mol. Med., 21 (2008), pp. 637–643
12.
o J.Q. Yin, J.N. Shen, W.W. Su, J. Wang, G. Huang, S. Jin, Q.C. Guo, C.Y. Zou, H.M. Li, F.B.
Li
o Bufalin induces apoptosis in human osteosarcoma U-2OS and U-2OS methotrexate300-resistant cell lines
o Acta Pharmacol. Sin., 28 (2007), pp. 712–720
13.
o C.M. Xie, W.Y. Chan, S. Yu, J. Zhao, C.H. Cheng
o Bufalin induces autophagy-mediated cell death in human colon cancer cells through reactive oxygen species generation and JNK activation
o Free Radic. Biol. Med. (2011)
14.
o H.Y. Li, W. Xu, X. Zhang, W.D. Zhang, L.W. Hu
o Bufalin inhibits CYP3A4 activity in vitro and in vivo
o Acta Pharmacol. Sin., 30 (2009), pp. 646–652
15.
o D.R. Green, G. Kroemer
o The pathophysiology of mitochondrial cell death
o Science, 305 (2004), pp. 626–629
16.
o L.R. Thomas, A. Henson, J.C. Reed, F.R. Salsbury, A. Thorburn
o Direct binding of Fas-associated death domain (FADD) to the tumor necrosis factor-related apoptosis-inducing ligand receptor DR5 is regulated by the death effector domain of FADD
o J. Biol. Chem., 279 (2004), pp. 32780–32785
17.
o J.H. Chiang, J.S. Yang, C.Y. Ma, M.D. Yang, H.Y. Huang, T.C. Hsia, H.M. Kuo, P.P. Wu, T.H.
Lee, J.G. Chung Danthron
o An anthraquinone derivative, induces DNA damage and caspase cascades-mediated apoptosis in SNU-1 human gastric cancer cells through mitochondrial permeability transition pores and bax-triggered pathways
o Chem. Res. Toxicol., 24 (2011), pp. 20–29
18.
o J.E. Chipuk, T. Moldoveanu, F. Llambi, M.J. Parsons, D.R. Green
o Mol. Cell, 37 (2010), pp. 299–310
19.
o S.W. Fesik
o Promoting apoptosis as a strategy for cancer drug discovery
o Nat. Rev. Cancer, 5 (2005), pp. 876–885
20.
o C.C. Lu, J.S. Yang, A.C. Huang, T.C. Hsia, S.T. Chou, C.L. Kuo, H.F. Lu, T.H. Lee, W.G.
Wood, J.G. Chung
o Chrysophanol induces necrosis through the production of ROS and alteration of ATP levels in J5 human liver cancer cells
o Mol. Nutr. Food Res., 54 (2010), pp. 967–976
21.
o Y.Y. Chen, S.Y. Chiang, J.G. Lin, Y.S. Ma, C.L. Liao, S.W. Weng, T.Y. Lai, J.G. Chung
Emodin
o Aloe-emodin and rhein inhibit migration and invasion in human tongue cancer SCC-4 cells through the inhibition of gene expression of matrix metalloproteinase-9
o Int. J. Oncol., 36 (2010), pp. 1113–1120
22.
o S.H. Wu, L.W. Hang, J.S. Yang, H.Y. Chen, H.Y. Lin, J.H. Chiang, C.C. Lu, J.L. Yang, T.Y.
Lai, Y.C. Ko, J.G. Chung
o Curcumin induces apoptosis in human non-small cell lung cancer NCI-H460 cells through ER stress and caspase cascade- and mitochondria-dependent pathways
o Anticancer Res., 30 (2010), pp. 2125–2133
23.
o J.S. Yang, M.J. Hour, W.W. Huang, K.L. Lin, S.C. Kuo, J.G. Chung
o MJ-29 inhibits tubulin polymerization, induces mitotic arrest, and triggers apoptosis via cyclin-dependent kinase 1-mediated Bcl-2 phosphorylation in human leukemia U937 cells
o J. Pharmacol. Exp. Ther., 334 (2010), pp. 477–488
24.
o J.P. Lin, J.S. Yang, N.W. Chang, T.H. Chiu, C.C. Su, K.W. Lu, Y.T. Ho, C.C. Yeh, D. Mei, H.J.
Lin, J.G. Chung
o GADD153 mediates berberine-induced apoptosis in human cervical cancer Ca ski cells
o Anticancer Res., 27 (2007), pp. 3379–3386
o Y.T. Lin, J.S. Yang, H.J. Lin, T.W. Tan, N.Y. Tang, J.H. Chaing, Y.H. Chang, H.F. Lu, J.G. Chung
o Baicalein induces apoptosis in SCC-4 human tongue cancer cells via a Ca2+
-dependent mitochondrial pathway
o In Vivo, 21 (2007), pp. 1053–1058
26.
o B.C. Ji, W.H. Hsu, J.S. Yang, T.C. Hsia, C.C. Lu, J.H. Chiang, J.L. Yang, C.H. Lin, J.J. Lin,
L.J. Suen, W. Gibson Wood, J.G. Chung
o Gallic acid induces apoptosis via caspase-3 and mitochondrion-dependent pathways in vitro and suppresses lung xenograft tumor growth in vivo
o J. Agric. Food Chem., 57 (2009), pp. 7596–7604
27.
o Y.T. Ho, J.S. Yang, T.C. Li, J.J. Lin, J.G. Lin, K.C. Lai, C.Y. Ma, W.G. Wood, J.G. Chung
o Berberine suppresses in vitro migration and invasion of human SCC-4 tongue squamous cancer cells through the inhibitions of FAK, IKK, NF-kappaB, u-PA and MMP-2 and -9
o Cancer Lett., 279 (2009), pp. 155–162
28.
o W.Z. Ying, P.W. Sanders
o Cytochrome c mediates apoptosis in hypertensive nephrosclerosis in Dahl/Rapp rats
o Kidney Int., 59 (2001), pp. 662–672
29.
o C. Lo, T.Y. Lai, J.H. Yang, J.S. Yang, Y.S. Ma, S.W. Weng, Y.Y. Chen, J.G. Lin, J.G. Chung
o Gallic acid induces apoptosis in A375.S2 human melanoma cells through caspase-dependent and -independent pathways
o Int. J. Oncol., 37 (2010), pp. 377–385
30.
o P.F. Jones, T. Jakubowicz, F.J. Pitossi, F. Maurer, B.A. Hemmings
o Molecular cloning and identification of a serine/threonine protein kinase of the second-messenger subfamily
o Proc. Natl. Acad. Sci. U.S.A., 88 (1991), pp. 4171–4175
31.
o A. Enomoto, H. Murakami, N. Asai, N. Morone, T. Watanabe, K. Kawai, Y. Murakumo, J.
Usukura, K. Kaibuchi, M. Takahashi
o Akt/PKB regulates actin organization and cell motility via Girdin/APE
o Dev. Cell, 9 (2005), pp. 389–402
o P.J. Coffer, J.R. Woodgett
o Molecular cloning and characterisation of a novel putative protein-serine kinase related to the cAMP-dependent and protein kinase C families
o Eur. J. Biochem., 201 (1991), pp. 475–481
33.
o K. Nasu, M. Nishida, T. Ueda, N. Takai, S. Bing, H. Narahara, I. Miyakawa
o Bufalin induces apoptosis and the G0/G1 cell cycle arrest of endometriotic stromal cells: a promising agent for the treatment of endometriosis
o Mol. Hum. Reprod., 11 (2005), pp. 817–823
34.
o L. Sun, T. Chen, X. Wang, Y. Chen, X. Wei
o Bufalin induces reactive oxygen species dependent Bax translocation and apoptosis in ASTC-a-1 Cells
o Evid. Based Complement. Alternat. Med. (2009)
35.
o Y. Otsuki
o Apoptosis in human endometrium: apoptotic detection methods and signaling
o Med. Electron Microsc., 34 (2001), pp. 166–173
36.
o S.J. Gardai, D.A. Hildeman, S.K. Frankel, B.B. Whitlock, S.C. Frasch, N. Borregaard, P.
Marrack, D.L. Bratton, P.M. Henson
o Phosphorylation of Bax Ser184 by Akt regulates its activity and apoptosis in neutrophils
o J. Biol. Chem., 279 (2004), pp. 21085–21095
37.
o M. Xin, X. Deng
o Nicotine inactivation of the proapoptotic function of Bax through phosphorylation
o J. Biol. Chem., 280 (2005), pp. 10781–10789
38.
o H. Yamaguchi, H.G. Wang
o The protein kinase PKB/Akt regulates cell survival and apoptosis by inhibiting Bax conformational change
o Oncogene, 20 (2001), pp. 7779–7786
39.
o N. Oka, S. Tanimoto, R. Taue, H. Nakatsuji, T. Kishimoto, H. Izaki, T. Fukumori, M.
o Role of phosphatidylinositol-3 kinase/Akt pathway in bladder cancer cell apoptosis induced by tumor necrosis factor-related apoptosis-inducing ligand