國立交通大學
生物科技系
博士論文
亞硫酸基轉移酶之定位與功能分析
Localization and Functional Analysis of Sulfotransferase
研 究 生 : 劉慈安
指導教授 : 楊裕雄 博士
亞硫酸基轉移酶之定位與功能分析
Localization and Functional Analysis of Sulfotransferase
研究生: 劉慈安 Student: Tzu-An Liu
指導教授: 楊裕雄 博士 Advisor: Dr. Yuh-Shyong Yang
國立交通大學
生物科技研究所
博士論文
A Thesis
Submitted to Department of Biological Science and Technology
National Chiao Tung University
In partial fulfillment of the Requirements for the Degree of PhD
In
Biological Science and Technology
Hsinchu, Taiwan, Republic of China
August, 2008
中華民國九十七年八月
亞硫酸基轉移酶之定位與功能分析
學生: 劉慈安 指導教授: 楊裕雄 博士
國立交通大學科技系博士班
中文摘要
亞硫酸基轉移酶參與許多重要生理反應,如藥物代謝、類固醇與荷爾蒙調控、調節 神經傳導物質功能和癌化過程。在亞硫酸基化反應中,磷酸腺苷酸磷酰硫酸鹽 (PAPS) 為亞硫酸基的來源,其受質可廣泛的包含醣類ヽ蛋白質ヽ多種內生性小分子及環境荷爾 蒙等。 為了進一步探討亞硫酸基轉移酶在生物體中可能的生理與功能意義,本研究利 用兩種重要的動物模型,果蠅及斑馬魚,作為亞硫酸基轉移酶之定位與功能分析之平 台。首先,研究結果顯示在果蠅全腦中,類脫氫表雄甾酮亞硫酸基轉移酶之蛋白質 (DHEA ST-like protein) 選擇性地表現在果蠅腦中特殊的神經細胞及神經纖維,推測可能 和記憶與學習迴路相關。在斑馬魚中,有兩種新的亞硫酸基轉移酶被確認。此兩種酶針 對環境荷爾蒙,特別是羥化多氯化聯苯 (hydroxylated PCB) 有最強的亞硫酸基化能力, 在斑馬魚的胚胎形成與發育過程中,此兩種酶大量表現在幼魚時期,推測在斑馬魚器官 發育之初期,魚體已具有代謝環境荷爾蒙的能力。另外,本研究發展出一個針對酪胺酸 亞硫酸基化之偵測方法,並發現三個恆常性血液蛋白有酪胺酸亞硫酸基化的現象。結果 顯示,以果蠅及斑馬魚為平台,本研究詳細地探討亞硫酸基轉移酶之定位與功能分析, 期能為日後之研究提供可用之相關資訊。Localization and Functional Analysis of Sulfotransferase
Student: Tzu-An Liu Advisor: Dr. Yuh-Shyong Yang
Department of Biological Science and Technology
National Chiao Tung University
ABSTRACT
Sulfotransferases (SULTs) constitute a superfamily of related enzymes that play important roles in the regulation of detoxification, steroid hormone action, neurotransmitter function, drug metabolism, and carcinogenesis. Sulfation, catalyzed by SULTs, is the critical biotransformation process for transferring a sulfuryl group from the cofactor 3’-phosphoadenosine-5’-phophosulfate (PAPS) to a wide range of structurally diverse endogenous and xenobiotic compounds. It is believed that understanding the expression pattern and cellular distribution of SULTs, along with the functional and structural information, will assist in determining the physical and functional significance of SULTs. Here we utilized two ideal animal models, fruit fly (Drosophila melanogaster) and zebrafish (Danio rerio), as the alternative testing systems for investigating the localization and functional significance of SULTs. Firstly, a systematical analysis demonstrated that dehydroepiandrosterone sulfotransferase-like protein was expressed in fruit fly, which so far has no evidence indicating the presence of cytosolic SULTs, and was abundant in the specific neural bodies as well as in several bundles of synapses in fruit fly neuronal circuits. Secondly, two novel cytosolic SULTs, SULT1 ST7 and SULT1 ST8, were identified and characterized in zebrafish. They both exhibited strong sulfating activities toward environmental estrogens, particularly hydroxylated polychlorinated biphenyls (PCBs), among various endogenous and xenobiotic compounds tested as substrates. Developmental expression experiments also revealed distinct expression patterns of SULT1 ST7 and ST8 during embryonic development and throughout the larval stage onto maturity. Lastly, to better understand the functional regulation of SULTs, a target-specific approach for the identification of tyrosine sulfation had been established. Three new tyrosine-sulfated hemostatic proteins were identified. Such a target-specific approach will allow for investigation of sulfated-tyrosine proteins of other biochemical/physiological pathways/processes and contribute to a better understating of post-translational modification by tyrosine sulfation.
ACKNOWLEDGEMENT
Science is definitely not done by individuals. No one can pass through this baptism of fire without the help and support from others. At this point in my life, I would like to express my deepest appreciation to those who had have provided me with great help along this long way, and the people who have shaped the person I am today.
In the first place, I would like to thank my advisor Professor Yuh-Shyong Yang. He has opened a new door for me to explore the world through. His enthusiasm and interest for science has invigorated me with a desire to investigate new and interesting topics. In particular, he gave me the opportunity and freedom to experience the research at the forefront of science, and to grow by teaching myself instead of telling me what to do at every juncture. It was with him that I first realized that I am capable of handling endless problems as well as taking on the varied responsibilities.
My deepest appreciation is extended to Professor Ming-Cheh Liu for his willingness to take me into his lab at University of Texas, Health Center at Tyler. I have learned not only scientific techniques from him, but also more importantly, the attitude and characteristics for being a good scientist. I am truly grateful for his guidance and support throughout my stay in Sates. I also want to express my gratitude to my entire dissertation committee: Dr. Ming-Yih Liu, Dr. Horng-Dar Wang, Dr. Chao-Hsiung Lin, Dr. Ching-Te Chien, and Dr. You-Yin Chen. I appreciate them taking the time to review the dissertation. I am so thankful for their valuable suggestions and criticisms.
I wish to extend the personal greetings and appreciation to Dr. Min-Chi Lu, the previous advisor during my master period, for his warm encouragement and inspiration over these years. He makes me believe that, ultimately, everything will turn out to be great! I consider him a teacher, a friend, and a mentor. I really appreciate what he has done for me.
At last, I would like to dedicate this honor to my family. Certainly, without the love and support that they have shown me, I will not be able to reach this goal.
To my lovely family, especially my parents, and all the loved ones—I love you so much!
with loves,
An
CONTENTS
ABSTRACT (Chinese)………. i ABSTRACT (English)……….. ii ACKONWLEDGMENT………... iii CONTENTS……….. iv ABBREVIATIONS………... vi THESIS OBJECTS CHAPTER 1 Introduction………. 1I. Background and significance……….. 1
1. Sulfotransferase……….. 1
2. Localization of human sulfotransferases……… 9
3. Biological importance of sulfotransferases………... 16
4. Invertebrate versus vertebrate: potential application of non-mammalian models for sulfation analysis………... 21
II. References………. 35
CHAPTER 2 Immunohistochemical analysis of a novel dehydroepiandrosterone sulfotransferase-like protein in Drosophila neural circuits………. 53
I. Introduction……… 55
II. Materials and methods………... 58
III. Results……… 64
IV. Discussion……….. 67
V. References………. 71
CHAPTER 3 Identification and characterization of two novel cytosolic sulfotransferases, SULT1 ST7 and SULT1 ST8, from zebrafish……….. 89
II. Materials and methods………... 94
III. Results……… 101
IV. Discussion……….. 107
V. References……….. 113
CHAPTER 4 Identification of tyrosine-sulfated hemostatic proteins by a target-specific chromatography………... 138
I. Introduction……… 139
II. Materials and methods………... 141
III. Results……… 144
IV. Discussion……….. 147
V. References……….. 149
APPENDIX……….. 159 Structure of sulfated compounds
ABBREVIATIONS
3-OST-3A glucosaminyl 3-O-sulfotransferase-3A 3-OST-3B glucosaminyl 3-O-sulfotransferase-3B 3-OST-1 glucosaminyl 3-O-sulfotransferase-1 3-OST-2 glucosaminyl 3-O-sulfotransferase-2 ARS aryl sulfatase
CNS central nervous system DHEA dehydroepiandrosterone
DHEAS dehydroepiandrosterone sulfate
DHEA ST dehydroepiandrosterone sulfotransferase
E2 17β-estradiol
EST estrogen sulfotransferase FGF fibroblast growth factor FTZ-F1 Fushi Tarazu factor-1 genes GI gastrointestinal tract
GlaNAc N-acetylgalactosamine GlcNAc N-acetylglucosamine HMW high molecular weight MEM minimum essential medium
PAPS 3’-phosphoadenosine 5’-phosphosulfate PAP 3’-phosphoadenosine 5’-phosphate PCB polychlorinated biphenyl
PREG pregnenolone PROG progesterone
PTM post-translational modification SULT/ ST sulfotransferase
Sox9 SRY HMG box related gene 9 TLC thin-layer chromatography
tPA tissue-type plasminogen activator TH ST thyroid hormone sulfotransferase TL PST thermolabile phenol sulfotransferase TS PST thermostable phenol sulfotransferase TPST tyrosylprotein sulfotransferase
CHAPTER 1
INTRODUCTION
I. Background and significance
1. Sulfotransferases1.1 General aspects of sulfotransferases
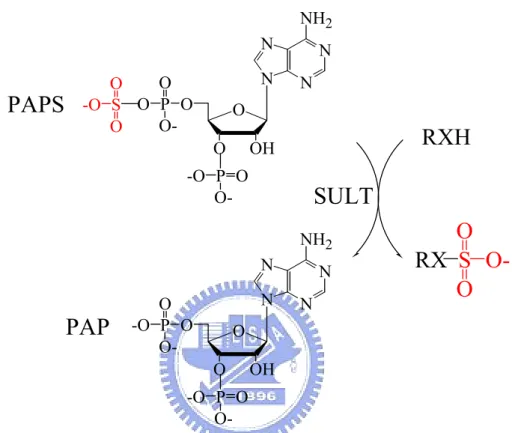
Sulfotransferases (SULTs) constitute a superfamily of related enzymes that catalyze the transfer of a sulfuryl group from the active sulfate donor, 3’-phophoadenosine 5’-phophosulfate (PAPS), to a substrate compound containing either a hydroxyl or an amino group (Figure 1) (Falany, 1997a; Coughtrie et al., 1998; Chapman et al., 2004). The reaction, usually referred to as “sulfation”, occurs in many prokaryotic and vertebrate species and plays an important role in numerous biological processes including detoxification, homeostasis of neurotransmitters, activation and deactivation of hormones and carcinogens, as well as transport and metabolism of steroids in circulation. Sulfation reactions are usually classified by the acceptor group involved in sulfoconjugation, e.g., O-sulfonation (ester) and N-sulfonation (amide) (Strott, 2002; Huxtable, 1986).
O-Sulfonation, a major type of cellular sulfation reaction, involves an alcohol group and can
occur with a numerous of relatively small endogenous and exogenous compounds such as hormones, steroids, catecholamines, drugs, and various xenobiotic agents. N-sulfonation,
although less predominant than O-sulfonation, is nevertheless a critical reaction in the posttranslational modification of macromolecules such as carbohydrates, peptides, and proteins (Strott, 2002). N N N N NH2 O OH O O P O O-O S -O O O O P -O O-N N N N NH2 O OH O O P -O O-O O P -O
O-RXH
RX
S
O
O
O-SULT
PAPS
PAP
Figure 1. Physical sulfation catalyzed by SULT with PAPS as a cosubstrate.
1.2 Classification of sulfotransferases
In vertebrates, SULTs can be divided into two classes: (i) cytosolic SULTs that are responsible for the sulfation of small xenobiotics and endogenous substrates such as hormones, steroids, bile acids, neurotransmitters, as well as multitude of environmental chemicals (Table 1) (Falany, 1997b); (ii) membrane-bound SULTs that are resident
macromolecules such as peptides, lipids, proteins, and glycosaminoglycans, affecting both their structural and functional characteristics, as well as the molecular-recognition events and signaling transduction pathway (Table 2) (Chapman et al., 2004; Gamage et al., 2006; Negishi et al., 2001). There are enormous amount of sulfated molecules in biological system. However, we still understand very little about their biological functions in either metabolic pathway or physiological significance. In particular, how the interplay between various members of the SULTs and arylsulfatase (ARS) families, which are capable of the hydrolysis of sulfate esters, to regulate the availability and biological activity of xenobiotics and endogenous molecules in vivo remains poorly understood.
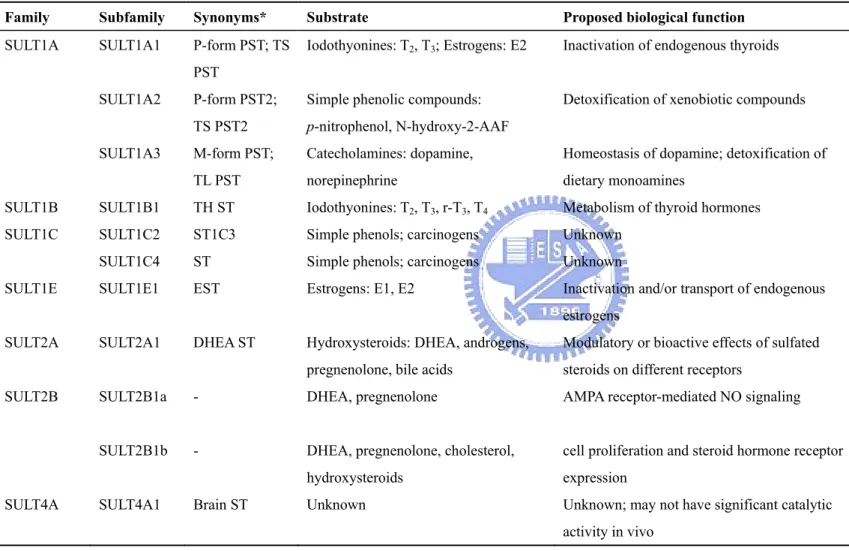
Table 1. Classification of human cytosolic SULTs
*Abbreviations: P-form PST, simple phenol-form phenol sulfotransferase; M-form PST, monoamine-form phenol sulfotransferase; TS PST, thermostable phenol
Family Subfamily Synonyms* Substrate Proposed biological function References
SULT1A1 P-form PST; TS PST
Iodothyonines: T2, T3; Estrogens: E2 Inactivation of endogenous thyroids Wilborn et al., 1994; Falany, 1997a
SULT1A2 P-form PST2;
TS PST2
Simple phenolic compounds:
p-nitrophenol, N-hydroxy-2-AAF
Detoxification of xenobiotic compounds Veronese et al., 1994 SULT1A
SULT1A3 M-form PST;
TL PST
Catecholamines: dopamine, norepinephrine
Homeostasis of dopamine; detoxification of dietary monoamines
Wood et al., 1994; Brix et al., 1999 SULT1B SULT1B1 TH ST Iodothyonines: T2, T3, r-T3, T4 Metabolism of thyroid hormones Wang et al., 1999
SULT1C2 ST1C3 Simple phenols; carcinogens Unknown Her et al., 1997
SULT1C
SULT1C4 ST Simple phenols; carcinogens Unknown Sakakibara et al., 1998a
SULT1E SULT1E1 EST Estrogens: E1, E2 Inactivation and/or transport of endogenous
estrogens
Aksoy et al., 1994; Falany et al., 1995 SULT2A SULT2A1 DHEA ST Hydroxysteroids: DHEA, androgens,
pregnenolone, bile acids
Modulatory or bioactive effects of sulfated steroids on different receptors
Comer et al., 1993; Forbes et al., 1995; Otterness et al., 1992
SULT2B SULT2B1a - DHEA, pregnenolone AMPA receptor-mediated NO signaling Her et al., 1998; Moloche and Falany,
2001; Kohjitani et al., 2008 SULT2B1b - DHEA, pregnenolone, cholesterol,
hydroxysteroids
cell proliferation and steroid hormone receptor expression
Her et al., 1998; He and Falany, 2007; Geese and Raftogianis, 2001
SULT4A SULT4A1 Brain ST Unknown Unknown; may not have significant catalytic
activity in vivo
Falany et al., 2000; Sakakibara et al., 2002; Allali-Hassani et al., 2007
dehydroepiandrosterone sulfotransferase; T2, 3, 3’-diiodothyronine; T3, 3, 3’, 5-triiodothyronine; r-T3, 3, 3’, 5’-reverse triiodothyronine; T4, thyroxine; E1 , estrone; E2,
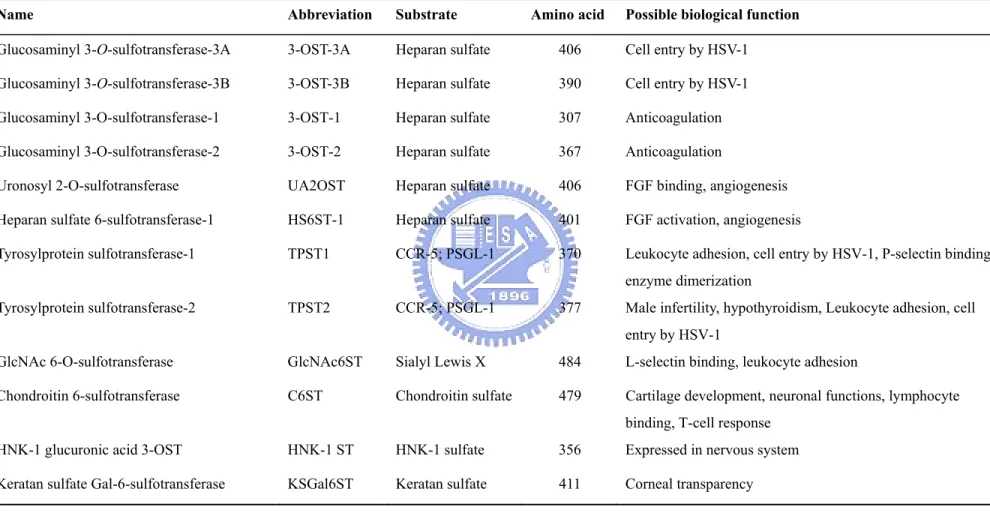
Table 2. Human membrane-bound SULTs
Name Abbreviation Substrate Amino acid Possible biological function
Glucosaminyl 3-O-sulfotransferase-3A 3-OST-3A Heparan sulfate 406 Cell entry by HSV-1 Glucosaminyl 3-O-sulfotransferase-3B 3-OST-3B Heparan sulfate 390 Cell entry by HSV-1 Glucosaminyl 3-O-sulfotransferase-1 3-OST-1 Heparan sulfate 307 Anticoagulation Glucosaminyl 3-O-sulfotransferase-2 3-OST-2 Heparan sulfate 367 Anticoagulation
Uronosyl 2-O-sulfotransferase UA2OST Heparan sulfate 406 FGF binding, angiogenesis
Heparan sulfate 6-sulfotransferase-1 HS6ST-1 Heparan sulfate 401 FGF activation, angiogenesis
Tyrosylprotein sulfotransferase-1 TPST1 CCR-5; PSGL-1 370 Leukocyte adhesion, cell entry by HSV-1, P-selectin binding, enzyme dimerization
Tyrosylprotein sulfotransferase-2 TPST2 CCR-5; PSGL-1 377 Male infertility, hypothyroidism, Leukocyte adhesion, cell entry by HSV-1
GlcNAc 6-O-sulfotransferase GlcNAc6ST Sialyl Lewis X 484 L-selectin binding, leukocyte adhesion
Chondroitin 6-sulfotransferase C6ST Chondroitin sulfate 479 Cartilage development, neuronal functions, lymphocyte binding, T-cell response
HNK-1 glucuronic acid 3-OST HNK-1 ST HNK-1 sulfate 356 Expressed in nervous system
Keratan sulfate Gal-6-sulfotransferase KSGal6ST Keratan sulfate 411 Corneal transparency
1.3 Molecular mechanism of sulfotransferases
To date twenty-seven structures of human SULTs have been solved on twelve different isoforms. Crystal structures for the SULT1A1 (phenol sulfotransferase), SULT1A3 (catecholamine sulfotransferase), SULT1E1 (estrogen sulfotransferase), SULT1B1 (thyronine sulfotransferase), SULT2A1 (dehydroepiandrosterone sulfotransferase), two isozymes of SULT2B1 (pregnenolone sulfotransferase and cholesterol sulfotransferase), SULT4A1 (brain sulfotransferase), three subfamilies of SULT1C, as well as four isozymes of membrane-bound SULTs (Heparan Sulfate Glucosamine 3-O- Sulfotransferase isoform 1-3; Heparan Sulfate N-Deacetylase N-Sulfotransferase) have been characterized (Kakuta et al., 1997 and 1999; Bidwell et al. 1999; Pedersen et al., 2000; Gamage et al., 2003). These structures of SULTs, illustrated highly conserved regions of active sites and PAPS-binding sites, are similar to those of nucleotide kinases (Kakuta et al., 1997). The largest variation among these SULTs is found in the substrate-binding region. The substrate specificity and selectivity of these enzymes have a long and confusing biography due to a great deal of substrate overlap among the known SULTs. Although all SULTs carry out the same function on sulfation, their substrate specificity is significantly distinctive between the enzyme subfamilies, such as phenol and alcohol SULTs. Several recent studies have focused on this topic and implicated some characteristics of cytosolic SULTs toward their substrates, such as stereospecificity (Park et al., 1999; Pai et al., 2002) and enantioselectivity (Sheng and
Duffel, 2003). It is possible that the critical residues interacting with substrate may underscore different substrate specificity of SULTs. In addition to substrate specificity, several imperative amino acid residues in highly conserved region of PAPS-binding site were also found (Chapman et al., 2004). It was demonstrated that Ser134 is the key residue that enables SULT1A to discriminate PAP from AMP (Hsiao and Yang, 2002) and that SULT1A utilizes other nucleotides, besides PAPS, with much less catalytic efficiency (Lin and Yang, 2000). These studies barely focused on the type of nucleotides with adenosine, other nucleotides functioning with SULT1A are poorly understood. Another two structural features of cytosolic SULTs were found recently. One is a flexible loop that was proposed to control the in and out of PAP or PAPS during SULT1A catalysis (Su and Yang, 2003). Previous studies demonstrated that oxidation of SULT1A alters the enzyme’s catalytic activity, pH optima and substrate activity (Marshall et al., 1997). The redox-responsive signal cascade is one of the post-translational modifications of protein, but the functions of this redox-sensing loop on SULT1A catalysis are still not clear. Cytosolic SULTs appeared to be capable of forming either homodimers or heterodimers in solution (Kiehlbauch et al., 1995). Recent articles indicated that there exists a common dimerization motif in cytosolic SULTs (Petrotchenko et al., 2001). However, the functional significance of the dimerization process is not known and the protein-protein interaction of SULTs has not been well studied.
2. Localization of human sulfotransferase
It is believed that knowing the expression pattern and cellular distribution of SULTs, along with the functional and structural information, will assist with determining the physical and functional significance of SULTs. To better understand the role of individual SULT isoforms in the regulation of various biological processes, a comprehensive study of cellular distribution of these enzymes in vivo is needed. Here an inclusive information has been studied in an attempt to illustrate the organ-specific distribution of individual SULT isoforms at mRNA and protein level (Table 3), and moreover, it may provide a useful information for further investigation of thephysiological function of these enzymes.
2.1 SULT1A subfamily
SULT1A1 exhibits the highest expression level among all SULT1 isoforms in the liver. It has also been identified in brain, intestine, breast, endometrium, adrenal gland, placenta, jejunum, platelets, as well as kidney and lung. SULT1A3 is highly expressed in the jejunum and placenta with the exception of adult human liver. For the developmental perspective, a significant level of expression of both SULT1A1 and SULT1A3 appeared in the fetal liver, intriguingly, SULT1A3 nearly disappears in adult kidney and liver. The cellular localization and physiological significance of SULT1A2 is the least understood among the SULT1A members. Dooley et al. (2000) had shown the SULT1A2 encoding mRNA is present in the
liver, brain, kidney, ovary, lung, and gastrointestinal tract (GI) at the lower level. However, it appears that SULT1A2 mRNA expression does not translate into the formation of protein (Gamage et al., 2006).
2.2 SULT1B subfamily
The predominant physical function of SULT1B1 appears to be in the regulation and biotransformation of thyroid hormones. Intriguingly, a significant expression of SULT1B1 mRNA and protein has been detected in the small intestine, colon, and is also found in liver and blood leukocytes at lower levels (Teubner et al., 1998; Wang et al., 1998).
2.3 SULT1C subfamily
The biological function of SULT1C is barely understood. Previous studies have indicated the presence of SULT1C2 in adult human kidney, stomach, thyroid, as well as fetal kidney and liver. It was also demonstrated that SULT1C2 protein was clearly detectable in stomach, ileum, jejunum, rectum, and cecum (Coughtrie, 2002). At the RNA level, the abundant expression of SULT1C4 was observed in fetal kidney and lung, as well as in the adult spinal cord, kidney, and ovary. However, the protein expression of SULT1C4 has not yet been demonstrated in any adult or fetal organisms or tissues (Sakakibara et al. 1998b).
2.4 SULT1E subfamily
SULT1E1 protein was detected in liver (Forbes-Bamforth and Coughtrie, 1994; Falany et al. 1995), endometrium (Falany et al. 1998), jejunum (Her et al. 1996), and mammary epithelial cells in primary culture (Falany and Falany, 1996). The significant level of mRNA encoding SULT1E1 on the immunoblots has been demonstrated in both human liver and jejunum cytosol, and mammary epithelial cells in primary culture (Forbes-Bamforth and Coughtrie, 1994; Falany et al., 1995). The enzymatic activity of SULT1E1 was also assessed by using β-estrogen as a substrate in the different human organs, such as fetal liver, kidney, lung (Adjei et al., 2008).
.
2.5 SULT2A subfamily
Northern analysis has shown SULT2A1 is extensively present in many human tissues, such as liver, adrenal, small intestine, ovary, prostate, lover, stomach, small intestine, colon, as well as brain and bone marrow (Otterness et al., 1992; Tashiro et al., 2000). The immunohistochemical study has revealed that SULT2A1 is selectively expressed in the embryonic human hepatocytes and around the central vein (Baker et al., 1994). Paker et al. (1994) also reported that SULT2A1 expression was detected in the fetal and neocortical zones of the adrenal cortex, liver, testis, and intestine. Further, kidney SULT2A1 immunostaining was present in the proximal and distal tubules, loops of Henle, collecting
ducts, and their progenitors (Baker et al., 1994). Despite the localization of SULT2A1 were inclusively investigated, the comprehensively physiological implication of this isozyme remains ambiguous in vivo, suggesting a simple and convenient genetic model might be needed for the further investigation.
2.6 SULT2B subfamily
Both SULT2B isoforms mRNAs have been detected in many tissues including adrenal gland, placenta, ovary, prostate, lung, kidney, colon and skin (Her et al., 1998; Dooley et al., 2000; Meloche and Falany, 2001; Javitt et al., 2001; Geese and Raftogianis, 2001). Further, the results demonstrated that SULT1Bb was more extensively expressed than SULT1Ba in a variety of hormone-responsive tissues, such as stomach, small intestine, spleen, thymus, thyroid, and liver (Geese and Raftogianis, 2001). Additionally, the protein expression of SULT1Bb is present in prostate and lung, whereas SULT1Ba is present in prostate and placenta (Geese and Raftogianis, 2001).
2.7 SULT4A subfamily
SULT4A1 is also termed “brain sulfotransferase”, which sharing an extremely high degree of sequence homology (97% amino acid identity) with the orthologous rat and mouse SULTs (Blanchard et al., 2004). Immunohistochemical staining of human brain
sections showed that SULT4A1 is located extensively, but exclusively, in a variety of brain regions including cerebral cortex (motor, cingulate, arontal), globus pallidus, medial temporal lobe (island of Calleja), amygdala pituitary, thalamus, cerebellum (dentate nucleus folia: vermis, granular layer), midbrain (oculomotor, red nucleus, magnocellular), and brainstem (Liyou et al., 2003). At present, the physiological substrate of SULT4A1 remains unknown.
2.8 TPST subfamily
TPSTs reside in the Golgi compartment and therefore have access to generate the posttranslational modification of secretory and membrane proteins transported through the trans-Golgi network (Kehoe and Bertozzi, 2000; Monigatti et al., 2006). Northern analysis
has demonstrated that both human TPST1 and TPST2 are broadly expressed in many tissues including cerebellum, fetal brain, trachea, testis, spinal cord, thyroid gland, uterus, lung, kidney, salivary gland, prostate, skeletal muscle and uterus (Ouyang et al., 1998; Mishiro et al., 2006). However, due to the lack of the isoenzyme-specific antibodies or suitable analytic reagents and probes, the cellular distribution and related abundance of TPST isoforms at the protein level have not yet been studied.
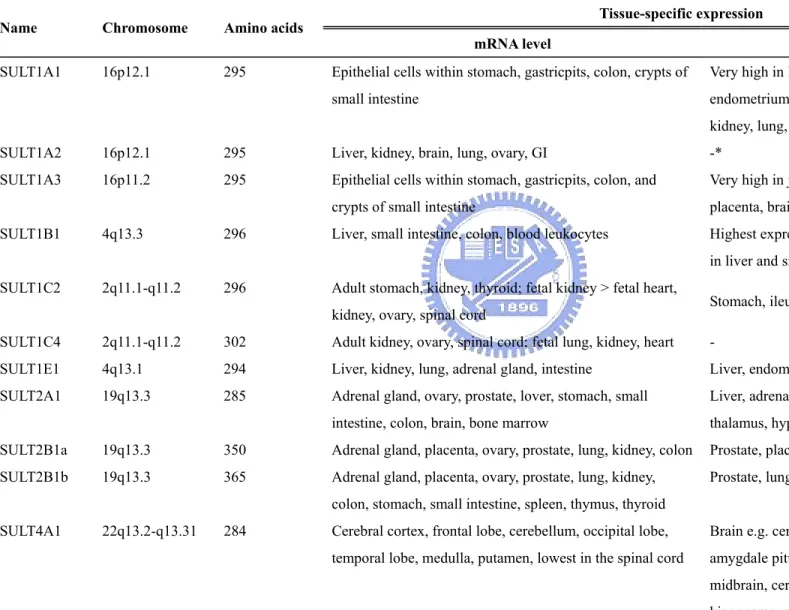
Table 3. Tissue-specific distribution of human cytosolic SULTs and TPST isoforms
Tissue-specific expression Name Chromosome Amino acids
mRNA level Protein expression
SULT1A1 16p12.1 295 Epithelial cells within stomach, gastricpits, colon, crypts of small intestine
Very high in liver; brain, breast, intestine, endometrium, adrenal gland, platelets, placenta, kidney, lung, jejunum
SULT1A2 16p12.1 295 Liver, kidney, brain, lung, ovary, GI -*
SULT1A3 16p11.2 295 Epithelial cells within stomach, gastricpits, colon, and crypts of small intestine
Very high in jejunum and colon; intestine, platelets, placenta, brain
SULT1B1 4q13.3 296 Liver, small intestine, colon, blood leukocytes Highest expression observed in colon; also detected in liver and small intestine
SULT1C2 2q11.1-q11.2 296 Adult stomach, kidney, thyroid; fetal kidney > fetal heart,
kidney, ovary, spinal cord Stomach, ileum, jejunum, rectum, cecum
SULT1C4 2q11.1-q11.2 302 Adult kidney, ovary, spinal cord; fetal lung, kidney, heart -
SULT1E1 4q13.1 294 Liver, kidney, lung, adrenal gland, intestine Liver, endometrium, GI, trachea, pancreas SULT2A1 19q13.3 285 Adrenal gland, ovary, prostate, lover, stomach, small
intestine, colon, brain, bone marrow
Liver, adrenal, duodenum, central vein, brain e.g. thalamus, hypothalamus
SULT2B1a 19q13.3 350 Adrenal gland, placenta, ovary, prostate, lung, kidney, colon Prostate, placenta SULT2B1b 19q13.3 365 Adrenal gland, placenta, ovary, prostate, lung, kidney,
colon, stomach, small intestine, spleen, thymus, thyroid
Prostate, lung SULT4A1 22q13.2-q13.31 284 Cerebral cortex, frontal lobe, cerebellum, occipital lobe,
temporal lobe, medulla, putamen, lowest in the spinal cord
Brain e.g. cerebral cortex, medial temporal lobe, amygdale pituitary, thalamus, hypothalamus, midbrain, cerebellum, lentiform nucleus,
(Continued)
*Related study is currently not available.
TPST1 7q11.21 370 Cerebellum, fetal brain, trachea, testis, spinal cord, thyroid gland, uterus, lung, kidney, salivary gland, prostate, skeletal muscle, uterus
- TPST2 22q12.1 377 Cerebellum, fetal brain, trachea, testis, spinal cord, thyroid
gland, uterus, lung, kidney, salivary gland, prostate, skeletal muscle, uterus
3. Biological importance of sulfotransferases 3.1 Sulfation in biology
In vertebrates, two classes of SULTs can be distinguished. One of which, membrane associated SULTs, sulfonates endogenous macromolecular molecules and comprises mainly membrane-bound forms localized in Golgi apparatus. No xenobiotic-metabolizing properties have been reported insofar. The other class of enzyme is cytosolic SULTs which were responsible for sulfation of various small endo- and exogenous compounds, such as hormones, bioamines, drugs, and various xenobiotic agents (Falany, 1997a). Although the range of compounds which were sulfated is enormous, yet we still understand relatively little of the functions either of the metabolic pathway or physiological significance. Furthermore, large numbers of studies revealed the possible associations between the polymorphisms of SULTs and physiological disorders, for example, cancers, hypertension, neoplasias and neurodegenerative diseases, which were seriously epidemiological risks for human beings (Glatt and Meinl, 2004; Sachse et al., 2002; Wu et al., 2003; Peng et al., 2003; Steiner et al, 2000). Whereas sulfation is vital for various physiological regulations, hydrolysis of sulfate esters, catalyzed by ARS, also linked to many important cellular functions including bioactivation of endogenous compounds, cellular degradation and modulation of signaling pathways. Particularly, how the association between the members of SULT families and diseases regulates the availability and biological activity of
xenobiotics and endogenous chemicals remains to be clarified.
3.2 Hepatic detoxication and biotransformation
Sulfation is a major way in vivo for the biotransformation of a wide range of structurally diverse endo- and exogenous substrates including phenolic (17β-estradiol; E2), alicyclic hydroxysteroids (DHEA), and iodothyronines, phenols (2-naphthol), primary (ethanol) and secondary alcohols (2-butanol) (Gamage et al., 2006; Goldstein and Faletto, 1993; Chapman et al., 2004). The responsible enzymes, cytosolic SULTs, are traditionally known as a group of Phase II detoxifying enzymes widely found among various invertebrate and vertebrate species (Nowell and Falany, 2006). Conjugation with sulfate is generally considered a detoxication or inactivation mechanism (Goldstein and Faletto, 1993). The sulfated compounds may become more soluble in aqueous solution, less ability of the penetration of cell membrane, and therefore, can be more easily excreted from the body via the kidney or bile (Glatt et al., 2001). For example, acetaminophen, an analgesic, is chiefly excreted as sulfated conjugates in the urine (Glatt and Meinl, 2004). From this perspective, SULTs can be considered as a part of the chemical defense mechanism in vivo (Allali-Hassani et al., 2007; Gamage et al., 2006).
compounds including as aminoazo dyes, benzidines, heterocyclic amines, hydroxymethyl polycyclic aromatic hydrocarbons, terpenes, β-aminoethyl alcohols, and 2-nitropropane (Michejda and Kroeger-Koepke, 1994; Gamage et al., 2006). For the N-hydroxy arylamines like 2-acetylaminofluorene (2-AAF), previous studies has demonstrated that the N-hydroxylated 2-AAF mediated by cytochrome 1A2 is a potent substrate of SULT1A2 and SULT1A1 (Glatt, 2000; Meinl et al., 2002). The sulfated metabolites were found to be more carcinogenic than the parent amide, suggesting the importance of SULTs as one of the metabolic pathways involved in the activation of 2-AAF (Meinl et al., 2002).
3.3 Regulation of sulfotransferase and its metabolites in nervous system
In 1913, Harvey Cushing,a giant of American medicine and brain surgery, presented his concept that “psychic conditions profoundly influence the discharges from the glands of internal secretion”. He postulated that each glandular disorder would induce a typical psychopathology. The steroidal environment of the brain varies as a function of the life cycle or apart of intercurrent events such stress and illness means this can have profound consequences both directly on the integrity of the brain and on its resistance to toxic or noxious agents. The term “neurosteroids”, apart from being synthesized by classical steriodogenic organs, such as progesterone (PROG), pregnenolone (PREG) and DHEA, were produced by de novo in central nervous system (CNS) and peripheral nervous system
(PNS) (Vallée et al, 2001). They were found in high levels and regulated the activity of genes and protein synthesis, cellular development, neuroendocrine system and behavioral pattern (De Nicola et al, 1993). SULT2A1 catalyzes the sulfonation of DHEA in its transition into dehydroepiandrosterone sulfate (DHEAS) (Adams et al., 1985; Longcope et al., 1982; Longcope, 1996). Both DHEA and DHEAS are critical precursors for several types of androgens and estrogens. They also involve in numerous neurophysical processes, such as increasing neuronal excitability, and enhancing neural plasticity and neuroprotective properties. Previous studies have reported that the concentrations of DHEA(S) in blood decrease markedly with ageing in humans (Orentreich et al, 1992; Vallée et al, 2000), and have been proposed to be the neuromodulators involved in age-related cognitive decline (Näsman et al, 1991; Sunderland et al, 1989). These findings have led to the hypothesis that elevated concentrations of steroid and its sulfate product may influence both physical and cognitive aging (Vallée et al, 2001).
3.4 Pathopharmacological properties of sulfotransferases
An increasing volume of studies have revealed the possible relationship between the expression pattern of SULTs and physiological disorders such as cancers, obesity, hypertension, neoplasias and certain neurodegenerative diseases. The altered levels of SULTs in individuals may greatly vary the cellular responsiveness to pathological
mechanisms. For example, it is believed that the administration of the progestin-derived anti-breast cancer drugs, medrogestone and tibolone, significantly induces the transcriptional regulation of SULT1E1 in estrogen-responsive breast cancer, which normally expresses lower levels of SULT1E1 than health adults (Chetrite and Pasqualini, 2001; Pasqualini and Chetrite, 2007). SULT1E1 catalyzes the estrogen into the biologically inactive estrogen sulfates that inhibit their action to responsive receptors, indicating the SULT1E1 plays a critical role in the estrogen homeostasis that may be disrupted in breast cancer (Gamage et al., 2006; Chetrite and Pasqualini, 2001; Pasqualini and Chetrite, 2007).
Statistically significant associations were also observed between the SULT1A1 genotype and age, obesity and certain neoplasias including mammary, pulmonary, esophageal and urothelial cancer (Glatt and Meinl, 2004; Sachse et al., 2002; Wu et al., 2003; Peng et al., 2003; Steiner et al, 2000). The elevated SULT activity phenotype was significantly more frequent in the cancer patient than in the controls (Glatt et al., 2001). Furthermore, there continues a broad interest in exploring the physiopathological and neuroregulatory role of SULT2A in the induction and maintenance of the endocrine-dependent cancers, including breast cancer and carcinoma of prostate gland (Qian et al., 1998; Purohit et al., 1999; Billich et al., 2000; Baulieu, 1998; Puia and Belelli, 2001). Theoretically, the sulfate metabolites of steroids, such as estrone, and testerone,
could be critical mechanism in modulating the availability of unconjugated parent molecules interact with corresponding receptors. Because only the unconjugated molecule has the growth-promoting abilities, abnormal regulation or metabolism of SULT2A, therefore, it may have the pathological implications (Strott, 2002). Nevertheless, the association with diseases (e.g. neoplasias, hypertension) and health-related parameters appears to be multifarious and varies between subgroups. It remains an important issue for us to investigate the physiological and pathopharmacological roles of SULTs in more detail.
4. Invertebrate versus vertebrate: potential application of non-mammalian models for sulfation analysis
Human SULT comprises of a multi-gene family of proteins containing at least thirteen distinct members that differ remarkably in their enzymatic properties, localization, regulatory and metabolic mechanism (Lindsay et al., 2008). Due to the multicellular complexity of higher species, the lack of suitable genetic models and increasingly moral concerns, however, studies in sulfation have difficulty circumstantiating the physical functions and regulatory mechanisms of those enzymes in mammals. A simple animal model for elucidating the genetic and molecular dissection of complex mechanism is therefore needed.
Over the past decades, non-mammalian models were thought to be far from the higher organisms for the study of biomedical science because of the phylogenic and functional diversities. To date, it has become abundantly clear that some of the non-mammals are not only convenient materials but are also shared with the physiological and pharmacological properties common to humans (Peterson et al., 2008). Here we utilized two premiere model organisms, Drosophila melanogaster and zebrafish, as the alternative testing systems for the study of the localization and functional analysis of SULTs.
4.1 Drosophil. melanogaster
4.1.1 Drosophila: systematical biomaterial for biomedical study
Cytogenetic studies in the complete mapping of the Drosophila polytene chromosomes and genomic sequence have altered our estimate of the evolutionary and physiological relationship between vertebrate and invertebrate (Reiter et al., 2001; Gilbert, 2008). Among the 929 distinct human disease genes, 714 are associated with 548 unique Drosophila genes, of which 153 are associated with known mutant alleles and 56 more are
tagged by P-element insertions in or near the gene (Reiter et al., 2001; Gilbert, 2008). The rapid and high-throughput genetic screen of P-element insertion or deletion in Drosophila for the study of the mutagenesis and the genetic modifications has been widely utilized for many years (Igarashi, 2005; Gilbert, 2008; Castro and Carareto, 2004). This unique feature
revolutionizes the study of the transposition and regulation of a transposon in a eukaryote by using this tiny insect as a premiere model organism (Castro and Carareto, 2004). In addiction, Drosophila share most of the basic metabolic homeostasis found in vertebrates (Baker and Thummel, 2007). This tiny insect mimics the intricate carbohydrate, sterol, lipid metabolism, the timing and process of developmental events, as well as the pharmacological, neurobiological and pathological mechanisms in humans. Therefore, Drosophila melanogaster, with the functional similarities of genetic, developmental and metabolic
characterizations, provides an important insight for the studies of the higher vertebrate systems (Baker and Thummel, 2007).
4.1.2 SULTs in Drosophila
In Drosophila, several types of membrane-associated SULTs have been identified and characterized, and the physical functions and biological regulations modulated by the sulfate conjugates have been extensively investigated (Table 4) (Kushe-Gullberg and Kjellén, 2003; Xu et al., 2007; Kamimura et al., 2004 and 2006; Sergeev et al., 2001; Zhu et al., 2005; Sen et al., 1998). The sulfation of hexuronate and glucosamine units, usually N-acetylgalactosamine (GlaNAc) or N-acetylglucosamine (GlcNAc), has been shown to be
essential for the fibroblast growth factor (FGF)-mediated tracheal formation, the stability or intracellular trafficking of Notch protein, the developmental processes and embryogenesis,
as well as differentiation and neuronal functions (Lander et al., 1996; Xu et al., 2007; Kamimura et al., 2006; Sen et al., 1998; Sergeev et al., 2001; Zhu et al., 2005). The findings have demonstrated that sulfation is a critical regulator for developmental and neuroregulatory functions in Drosophila, however, there has been no evidence clearly indicating that cytosolic ST(s) is(are) present or expressed in this important animal model. Recently, Hattori et al. have reported that four cytosolic SULT homologs, designed as dmST1-4, were identified by a research of the Drosophila melanogaster genome database
(http://flybase.bio.indiana.edu/). Each of the four isozymes have classified into separate and novel gene family, as determined by the subsequent amino acid sequence alignment and molecular phylogeny analysis (Hattori et al., 2008). Despite the low amino acid sequence homology (less than 40%) between those isozymes, and also toward their vertebrate homologs, a degree of amino acid sequence similarity does exist between dmSTs and the human SULT family members suggests that, at least in part, the insect cytosolic SULTs share an common ancestral gene or conserved residues with other mammal SULTs (Hattori et al., 2008).
Table 4. Membrane-associated SULTs and SULT-related proteins found in Drosophila*
Drosophila Functional similarity Proposed functions References
Hs2st human Hs2st FGF-mediated tracheal formation Xu et al., 2007; Kamimura et al., 2006
Hs3st human Hs3st Notch signal Kamimura et al., 2004
Hs6st human Hs6st Stability or intracellular trafficking of Notch protein Kamimura et al., 2001 and 2006
PIPE-st1 human Hs2st Probably nonfunctional protein Sergeev et al., 2001; Zhu et al., 2005
PIPE-st2 human Hs2st Unknown Sergeev et al., 2001; Zhu et al., 2005
PIPE-st3 human Hs2st Unknown Sergeev et al., 2001; Zhu et al., 2005
PIPE-st4 human Hs2st Unknown Sergeev et al., 2001; Zhu et al., 2005
PIPE-st5 human Hs2st Unknown Sergeev et al., 2001; Zhu et al., 2005
PIPE-st6 human Hs2st Unknown Sergeev et al., 2001; Zhu et al., 2005
PIPE-st7 human Hs2st Unknown Sergeev et al., 2001; Zhu et al., 2005
PIPE-st8 human Hs2st Unknown Sergeev et al., 2001; Zhu et al., 2005
PIPE-st9 human Hs2st Formation of embryonic dorsal-ventral polarity Sen et al., 1998; Sergeev et al., 2001; Zhu et al., 2005
PIPE-st10 human Hs2st Unknown Sergeev et al., 2001; Zhu et al., 2005
Retinol dehydratase SULT-related protein Cytosolic SULT activity Pakhomova et al., 2005
4.2 Danio rerio (zebrafish)
4.2.1 An idea vertebrate model for the genomic and embryonic analysis
Zebrafish has in recent years emerged as a popular animal model for a wide range of studies (Beis and Stainier, 2006; Aleström et al., 2006; Strähle and Blader, 1994). The completion of the genome sequence and expression profiles of zebrafish has made it an ideal vertebrate model of choice for a wide spectrum of biological studies. Numerous scientists take advantages of this photogenic creature as an emerging model for functional genomics, pharmacology, developmental biology, and even human biomedical researches (Strähle and Blader, 1994). As a tropical teleost, zebrafish is remarkable for its many benefits: small size and ease of breeding, transparent embryo, rapidly developmental process, short generating intervals and sexually reproductive ability. The increasing popularity of zebrafish is also due to its amenability to well-characterized gene set of encoded proteins and to an eventual understanding of how they work to mimic the intricate genetic and physiological regulation in higher vertebrate animals. Exploiting the full potential of zebrafish is obligatory. In fact, zebrafish serves as a superior model than other invertebrate system, particularly in studies of large-scale mutagenesis and altered phenotype screening (Currie, 1996). By successful application of forward and reverse genetics, more than 2000 mutations are identified for perturbing the normal development of zebrafish.
4.2.2 Zebrafish cytosolic SULTs
The special characteristics make the zebrafish a premier model for a systematic investigation of the ontogeny, cell type/tissue/organ-specific expression, and physiological involvement of individual SULTs. A prerequisite for using zebrafish in these studies is the identification of the various cytosolic SULTs and their biochemical characterization. By searching the expressed sequence tag database, ten zebrafish cDNAs encoding putative cytosolic SULTs were systematically cloned, expressed, and characterized (Table 5) (Sugahara et al., 2003a, b, c, d; Liu et al., 2005; Yasuda et al., 2005a,b; Yasuda et al., 2006).
Of the ten zebrafish SULTs, six fall within the SULT1 gene gamily (Sugahara et al., 2003a,b; Liu et al., 2005; Yasuda et al., 2005a,b); three belong to the SULT2 gene family (Sugahara et al., 2003c; Yasuda et al., 2006); and one (designated SULT X) appears to be independent from all known SULT gene families (Sugahara et al., 2003d). The analysis of amino acid sequence via BLAST search revealed that these zebrafish SULT isozymes display sequence homology to mammalian SULTs.
The sequence analysis also revealed that the recombinant zebrafish SULTs contains two sequences resembling the so-called “signature sequences" (YPKSGTxW in the N-terminal region and RKGxxGDWKNxFT in the C-terminal region) which are similar to mammalian SULTs. Of these two sequences, YPKSGTxW has been demonstrated by X-ray
Table 5. Features of the zebrafish SULT family
Group Name Gene ID Locus GenBank accession No. Amino aicd Reference
SULT1 ST1 323424 chromosome 8 AY181064 299 Sugahara et al., 2003a SULT1 ST2 368269 chromosome 8 AY181065 301 Sugahara et al., 2003a SULT1 ST3 368270 chromosome 8 AY196985 301 Sugahara et al., 2003b SULT1 ST4 402915 chromosome 20 AY196986 304 Liu et al., 2005 SULT1 ST5 619193 unknown AY879099 293 Yasuda et al., 2005a SULT1
SULT1 ST6 436872 chromosome 12 AY937249 308 Yasuda et al., 2005b SULT2 ST1 338214 unknown AY181063 287 Sugahara et al., 2003c SULT2 ST2 777793 unknown DQ640387 287 Yasuda et al., 2006 SULT2
crystallography to be responsible for binding to the 5'-phosphosulfate group of PAPS, a co-substrate for SULT-catalyzed sulfation reactions, and thus designated the “5’-phosphosulfate binding (5’-PSB) motif” (Chapman et al., 2004). The recombinant zebrafish SULTs also contain the “3’-phosphate binding (3’-PB) motif” responsible for the binding to the 3’-phosphate group of PAPS.
4.2.3 Enzymatic characteristics of the zebrafish cytosolic SULTs
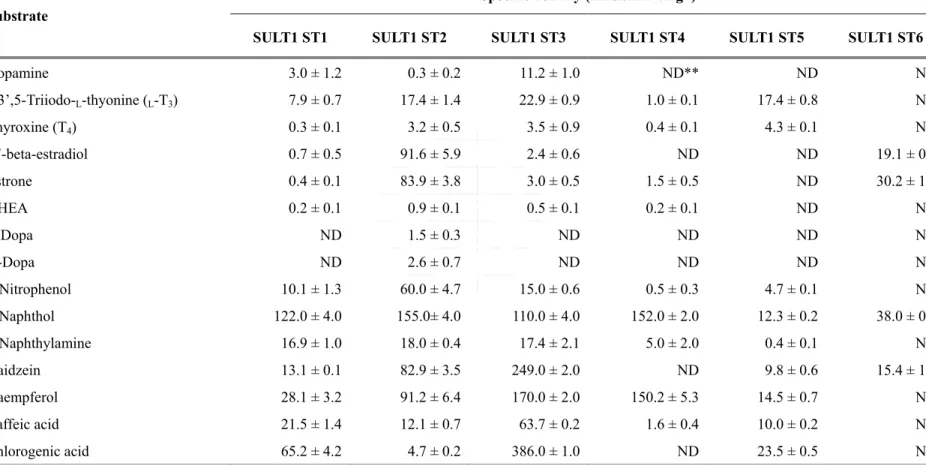
Zebrafish cytosolic SULTs had been shown to involve regulation of endogenous hormones and detoxication of xenobiotic chemicals in past five decades (Nowell and Falany, 2006; Gamage et al., 2006; Armstrong, 1987). A variety of endogenous and xenobiotic compounds were tested as substrates for the zebrafish SULTs in many studies, and the activity data obtained are given in Table 6. In view of substrate specificity and selectivity (Table7), zebrafish SULT1 group was found to prefer catalyzing phenolic compounds, such as dopamine, thyroid hormones, and estrogen. Among zebrafish SULT1 group, all except SULT1 ST6 were able to catalyze thyroid hormones. Intriguingly, zebrafish SULT1 ST5 showed sulfating activities toward only thyroid hormones and their metabolites, including
L-T3, 3,3’,5-triiodo-D-thyronine (D-T3), 3,3’,5’-triiodo-L-thyronine (L-rT3), L-thyroxine (L-T4),
Table 6. Substrate specificity of zebrafish cytosolic SULTsa
aReferences are listed in the Chapter I, 4.2.2.
bIt cannot be classified into any of the existing subfamilies within the SULT1 family. SULT
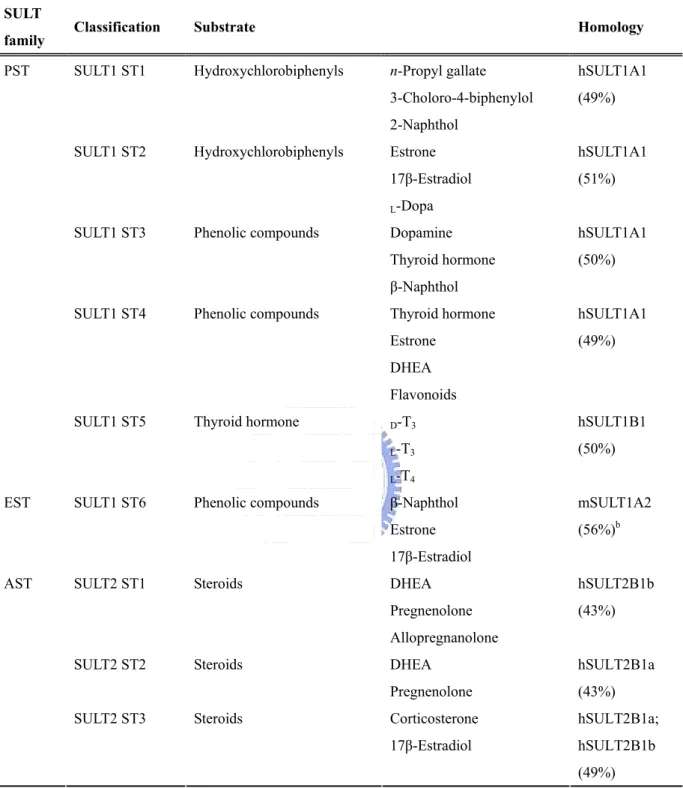
family Classification Substrate Homology
SULT1 ST1 Hydroxychlorobiphenyls n-Propyl gallate hSULT1A1
3-Choloro-4-biphenylol (49%)
2-Naphthol SULT1 ST2 Hydroxychlorobiphenyls Estrone hSULT1A1
17β-Estradiol (51%)
L-Dopa
SULT1 ST3 Phenolic compounds Dopamine hSULT1A1
Thyroid hormone (50%)
β-Naphthol SULT1 ST4 Phenolic compounds Thyroid hormone hSULT1A1
Estrone (49%)
DHEA Flavonoids
SULT1 ST5 Thyroid hormone D-T3 hSULT1B1
L-T3 (50%)
PST
L-T4
SULT1 ST6 Phenolic compounds β-Naphthol mSULT1A2
Estrone (56%)b
EST
17β-Estradiol
SULT2 ST1 Steroids DHEA hSULT2B1b
Pregnenolone (43%)
Allopregnanolone
SULT2 ST2 Steroids DHEA hSULT2B1a
Pregnenolone (43%)
SULT2 ST3 Steroids Corticosterone hSULT2B1a; 17β-Estradiol hSULT2B1b AST
Previous studies had shown that dopamine was catalyzed by human SULT1A3 (Chapman et al., 2004). Among zebrafish SULT1 group, we found ST1, 2, and 3 can utilize dopamine as substrate, with the highest activity by SULT1 ST3 (Sugahara et al., 2003b). It is remarkable that only SULT1 ST2 was determined for the sulfating capacity of the precursor of dopamine, Dopa. On the other hand, estrone and 17β-estradiol, the key steroid hormones for sex determination, were also catalyzed by SULT1 group (except ST4 and 5). It should also be pointed out that, of the six zebrafish SULT1 isoforms (Sugahara et al., 2003a, b; Liu et al., 2005; Yasuda et al., 2005a, 2005b), the SULT1 ST2 also exhibited strong activities toward estrone and 17β-estradiol (Yanagisawa et al., 1998; Sugahara et al., 2003c). In addition, the SULT1 ST 6 appears to be the only zebrafish SULT isoform known to date that displays substrate specificity exclusively for endogenous estrogens. These findings implied that the SULT1 ST6 plays a critical role in the metabolism and homeostasis of endogenous estrogens in vivo. Except endogenous estrogens, xenobiotic plant-derived phytoestrogens and environmental estrogens were also catalyzed by SULT1 group.
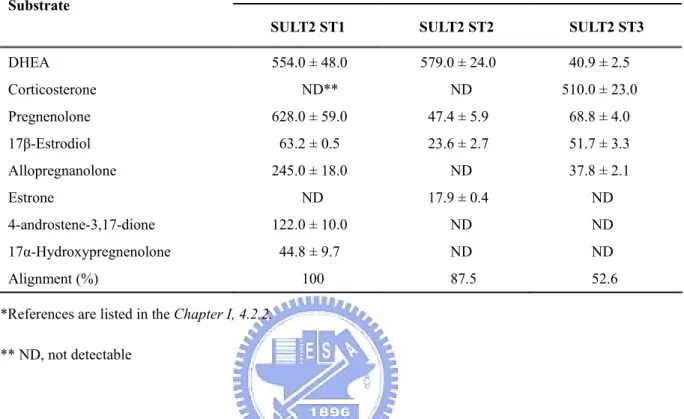
Zebrafish SULT2 was found to prefer catalyzing hydroxysteroids, such as DHEA, PREG, and corticosterone. To date, three zebrafish SULT2 isoforms had been cloned and characterized (as shown in Table 8). Similar to human SULT2A1, the zebrafish SULT2 ST1
Table 7. Specific activities of the zebrafish SULT1 subfamily toward endogenous and xenobiotic compounds*
*References are listed in the Chapter I, 4.2.2.
** ND, not detectable.
Specific activity (nmol.min-1.mg-1)
Substrate
SULT1 ST1 SULT1 ST2 SULT1 ST3 SULT1 ST4 SULT1 ST5 SULT1 ST6
Dopamine 3.0 ± 1.2 0.3 ± 0.2 11.2 ± 1.0 ND** ND ND 3,3’,5-Triiodo-L-thyonine (L-T3) 7.9 ± 0.7 17.4 ± 1.4 22.9 ± 0.9 1.0 ± 0.1 17.4 ± 0.8 ND Thyroxine (T4) 0.3 ± 0.1 3.2 ± 0.5 3.5 ± 0.9 0.4 ± 0.1 4.3 ± 0.1 ND 17-beta-estradiol 0.7 ± 0.5 91.6 ± 5.9 2.4 ± 0.6 ND ND 19.1 ± 0.6 Estrone 0.4 ± 0.1 83.9 ± 3.8 3.0 ± 0.5 1.5 ± 0.5 ND 30.2 ± 1.1 DHEA 0.2 ± 0.1 0.9 ± 0.1 0.5 ± 0.1 0.2 ± 0.1 ND ND L-Dopa ND 1.5 ± 0.3 ND ND ND ND D-Dopa ND 2.6 ± 0.7 ND ND ND ND p-Nitrophenol 10.1 ± 1.3 60.0 ± 4.7 15.0 ± 0.6 0.5 ± 0.3 4.7 ± 0.1 ND 2-Naphthol 122.0 ± 4.0 155.0± 4.0 110.0 ± 4.0 152.0 ± 2.0 12.3 ± 0.2 38.0 ± 0.5 2-Naphthylamine 16.9 ± 1.0 18.0 ± 0.4 17.4 ± 2.1 5.0 ± 2.0 0.4 ± 0.1 ND Daidzein 13.1 ± 0.1 82.9 ± 3.5 249.0 ± 2.0 ND 9.8 ± 0.6 15.4 ± 1.0 Kaempferol 28.1 ± 3.2 91.2 ± 6.4 170.0 ± 2.0 150.2 ± 5.3 14.5 ± 0.7 ND Caffeic acid 21.5 ± 1.4 12.1 ± 0.7 63.7 ± 0.2 1.6 ± 0.4 10.0 ± 0.2 ND Chlorogenic acid 65.2 ± 4.2 4.7 ± 0.2 386.0 ± 1.0 ND 23.5 ± 0.5 ND
Table 8. Specific activities of the zebrafish SULT2 subfamily with endogenous compounds as substrates*
*References are listed in the Chapter I, 4.2.2. ** ND, not detectable
Specific activity (pmol.min-1.mg-1) Substrate
SULT2 ST1 SULT2 ST2 SULT2 ST3
DHEA 554.0 ± 48.0 579.0 ± 24.0 40.9 ± 2.5 Corticosterone ND** ND 510.0 ± 23.0 Pregnenolone 628.0 ± 59.0 47.4 ± 5.9 68.8 ± 4.0 17β-Estrodiol 63.2 ± 0.5 23.6 ± 2.7 51.7 ± 3.3 Allopregnanolone 245.0 ± 18.0 ND 37.8 ± 2.1 Estrone ND 17.9 ± 0.4 ND 4-androstene-3,17-dione 122.0 ± 10.0 ND ND 17α-Hydroxypregnenolone 44.8 ± 9.7 ND ND Alignment (%) 100 87.5 52.6
can catalyze a wide range of neurosteroids, such as DHEA, PREG, allopregnanolone, 17α-hydroxypregnenolone, and 4-androstene-3,17-dione. It displayed strongest sulfating activity toward DHEA, but virtually no activities toward the rest of the endogenous and xenobiotic compounds tested, such as dopamine, T4, Dopa, estrone, β-naphthol, daidzein,
gallic acid, and n-propyl gallate (Sugahara et al., 2003c). It is notable that the SULT2 ST2 shares a high degree of sequence homology (87.5% amino acid identity) with the SULT2 ST1 and displayed strongest sulfating activity toward DHEA, however, the distinct substrate-catalyzing profile was found. Much lower sulfating activity toward PREG, and no activity toward allopregnanolone, 17α-hydroxypregnenolone, and 4-androstene-3,17-dione. The relatively low activity of SULT2 ST3 in catalyzing DHEA, PREG, allopregnanolone, 17α-hydroxypregnenolone, and 4-androstene-3,17-dione was found. Nevertheless, this isozyme appears to play an important role in the metabolism of corticosterone.
II. REFERENCES
Adams, J., Garcia, M., Rochefort, H. 1985. Estrogenic effects of physiological concentrations of 5-androstene-3 beta, 17 beta-diol and its metabolism in MCF7 human breast cancer cells. Cancer Res. 41, 4720-4726.
Adjei, A.A., Gaedigk, A., Simon, S.D., Weinshilboum, R.M., Leeder, J.S. 2008. Interindividual variability in acetaminophen sulfation by human fetal liver: implications for pharmacogenetic investigations of drug-induced birth defects. Birth. Defects Res. A Clin. Mol. Teratol. 82, 155-165.
Aleström, P., Holter, J.L., Nourizadeh-Lillabadi, R. 2006. Zebrafish in functional genomics and aquatic biomedicine. Trends Biotechnol. 24, 15-21.
Allali-Hassani, A., Pan, P.W., Dombrovski, L., Najmanovich, R., Tempel, W., Dong, A., Loppnau, P., Martin, F., Thornton, J., Edwards, A.M., Bochkarev, A., Plotnikov, A.N., Vedadi, M., Arrowsmith, C.H. 2007. Structural and chemical profiling of the human cytosolic sulfotransferases. PLoS Biol.5, e97
Armstrong, R.N. 1987. Enzyme-catalyzed detoxication reactions: mechanisms and stereochemistry. CRC Crit. Rev. Biochem. 22, 39-88.
Baker, E.V., Hume, R., Hallas, A., Coughtrie, W.H. 1994. Dehydroepiandrosterone sulfotransferase in the developing human fetus: quantitative biochemical and
immunological characterization of the hepatic, renal, and adrenal enzymes. Endocrinology 134, 982-989.
Baker, K.D., Thummel, C.S. 2007. Diabetic larvae and obese flies-emerging studies of metabolism in Drosophila. Cell Metab. 6, 257-266.
Baulieu, E.E. 1998. Neurosteroids: a novel function of the brain. Psychoneuroendocrinology 23, 963-987.
Beis, D., Stainier, D.Y. 2006. In vivo cell biology: following the zebrafish trend. Trends Cell Biol. 16, 105-112.
Bidwell, L.M., McManus, M.E., Gaedigk, A., Kakuta, Y., Negishi, M., Pedersen, L., Martin, J.L. 1999. Crystal structure of human catecholamine sulfotransferase. J. Mol. Biol. 293, 521-530.
Billich, A., Nussbaumer. P., Lehr, P. 2000. Stimulation of MCF-7 breast cancer cell proliferation by estrone sulfate and dehydroepiandrosterone sulfate: inhibition by novel non-steroidal steroid sulfatase inhibitors. J. Steroid Biochem. Mol. Biol. 73, 225-235.
Blanchard, R. L., Freimuth, R. R., Buck, J.,Weinshilboum,R. M., and Coughtrie, M. W. H. 2004. A proposed nomenclature system for the cytosolic sulfotransferase (SULT) superfamily. Pharmacogenetics 14, 199-211.
Brix, L.A., Duggleby, R.G., Gaedigk, A., McManus, M.E. 1999b. Structural characterization of human aryl sulphotransferases. Biochem. J.337, 337-343.
Castro, J.P., Carareto, C.M. 2004. Drosophila melanogaster P transposable elements: mechanisms of transposition and regulation. Genetica 121, 107-118.
Chapman, E., Best, M.D., Hanson, S.R., Wong, C.H. 2004. Sulfotransferases: structure, mechanism, biological activity, inhibition and synthetic utility. Angew. Chem. Int. Ed. 43, 3526-3548.
Chetrite, G.S., Pasqualini, J.R. 2001. The selective estrogen enzyme modulator (SEEM) in breast cancer. J. Steroid. Biochem. Mol. Biol. 76, 95-104.
Comer, K.A., Falany, J.L., Falany, C.N. 1993. Cloning and expression of human liver dehydroepiandrosterone sulfotransferase. Biochem. J. 289, 233-240.
Coughtrie, M.W.H., Sharp, S., Maxwell K., Innes N.P. 1998. Biology and function of the reversible sulfation pathway catalysed by human sulfotransferases and sulfatases. Chem Biol Interact. 109, 3-27.
Coughtrie, M.W.H. 2002. Sulfation through the looking glass-recent advances in sulfotransferase research for the curious. Pharmacogenomics J. 2, 297-308.
Currie, P.D. 1996. Zebrafish genetic: mutant cornucopia. Curr. Biol. 6, 1548-1552.
De Nicola, A.F. 1993. Steroid hormones and neuronal regeneration. Adv. Neurol. 59, 1199-206.
Dooley, T.P., Haldeman-Cahill, R., Joiner, J., Wilborn, T.W. 2000. Expression profiling of human sulfotransferase and sulfatase gene superfamilies in epithelial tissues and cultured
cells. Biochem. Biophys. Res. Commun. 277, 236-245.
Falany, C.N., Krasnykh, V., Falany, J.L. 1995. Bacterial expression and characterization of a cDNA for human liver estrogen sulfotransferase. J. Steroid Biochem. Molec. Biol. 52, 529-539.
Falany, C.N. 1997a. Enzymology of human cytosolic SULTs. FASEB J. 11, 206-216.
Falany, C.N. 1997b. Introducing: changing view of the sulfation and the sulfotransferases. FASEB J. 11, 1-2.
Falany, C.N., Xie, X., Wang, J., Ferrer, J., Falany, J.L. 2000. Molecular cloning and expression of novel sulphotransferase-like cDNAs from human and rat brain. Biochem. J. 346, 857-864.
Falany, J.L., Falany, C.N. 1996. Expression of cytosolic sulfotransferases in normal mammary epithelial cells and breast cancer cell lines. Cancer Res. 56, 1551-1555.
Falany, J.L., Azziz, R., Falany, C.N. 1998. Identification and characterization of cytosolic sulfotransferases in normal human endometrium. Chem. Biol. Interact. 109, 329-339.
Forbes, K.J., Hagen, M., Glatt, H., Hume, R., Coughtrie, M.W.H. 1995. Human fetal adrenal hydroxysteroid sulphotransferase: cDNA cloning, stable expression in V79 cells and functional characterisation of the expressed enzyme. Mol. Cell. Endocrinol. 112, 53-60. Forbes-Bamforth, K.J., Coughtrie, M.W.H. 1994. Identification of a new adult human liver sulfotransferase with specificity for endogenous and xenobiotic estrogens. Biochem.
Biophys. Res. Commun. 198, 707-711.
Gamage, N.U., Duggleby, R.G., Barnett, A.C., Tresillian, M., Latham, C.F., Liyou. N.E., McManus, M.E., Martin, J.L. 2003. Structure of a human carcinogen-converting enzyme, SULT1A1. Structural and kinetic implications of substrate inhibition. J. Biol. Chem. 278, 7655-7662.
Gamage, N., Barnett, A., Hempel, N., Duggleby, R.G., Windmill, K.F., Martin, J.L., McManus, M.E. 2006. Human sulfotransferases and their role in chemical metabolism. Toxicol. Sci. 90, 5-22.
Geese, W.J., Raftogianis, R.B. 2001. Biochemical characterization and tissue distribution of human SULT2B1. Biochem. Biophys. Res. Commun. 288, 280-289.
Gilbert, L.I. 2008. Drosophila is an inclusive model for human diseases, growth and development. Mol Cell Endocrinol. (Article in press)
Glatt, H. 2000. Sulfotransferases in the bioactivation of xenobiotics. Chem. Biol. Interact. 129, 141-170.
Glatt, H., Boeing, H., Engelke, C.E., Ma, L., Kuhlow, A., Pabel, U., Pomplun, D., Teubner, W., Meinl, W. 2001. Human cytosolic sulphotransferases: genetics, characteristics, toxicological aspects. Mutat. Res. 482, 27-40.
Glatt, H., Meinl, W. 2004. Pharmacogenetics of soluble sulfotransferases (SULTs). Naunyn Schmiedebergs Arch. Pharmacol. 369, 55-68.
Goldstein, J.A., Faletto, M.B. 1993. Advances in mechanisms of activation and deactivation of environmental chemicals. Environ. Health Perspect. 100, 169-176.
Hattori, K., Motohashi, N., Kobayashi, I., Tohya, T., Oikawa, M., Tamura, H.O. 2008. Cloning, expression, and characterization of cytosolic sulfotransferase isozymes from Drosophila melanogaster. Biosci. Biotechnol. Biochem. 72, 540-547.
He, D., Falany, C.N. 2007. Inhibition of SULT2B1b expression alters effects of 3beta-hydroxysteroids on cell proliferation and steroid hormone receptor expression in human LNCaP prostate cancer cells. Prostate 67, 1318-1329.
Her, C., Szumlanski, C., Aksoy, I.A., Weinshilboum, R.M. 1996. Human jejunal estrogen sulfotransferase and dehydroepiandrosterone sulfotransferase: immunochemical characterization of individual variation. Drug Metab. Dispos. 24, 1328-1335.
Her, C., Kaur, G.P., Athwal, R.S., Weinshilboum, R.M. 1997. Human sulfotransferase SULT1C1: cDNA cloning, tissue-specific expression, and chromosomal localization. Genomics 41, 467-470.
Her, C., Wood, T.C., Eichler, E.E., Mohrenweiser, H.W., Ramagli, L.S, Siciliano, M.J., Weinshilboum, R.M. 1998. Human hydroxysteroid sulfotransferase SULT2B1: two enzymes encoded by a single chromosome 19 gene. Genomics 53, 284-295.
Hsiao, Y.S., Yang, Y.S. 2002. A single mutation converts the nucleotide specificity of phenol sulfotransferase from PAP to AMP. Biochemistry 41, 12959-12966.
Huxtable, R.J. 1986. Biochemistry of sulfur. New York: Plenum Publishing Corp.
Igarashi, P. 2005. Overview: nonmammalian organisms for studies of kidney development and disease. J. Am. Soc. Nephrol.16, 296-298.
Javitt, N.B., Lee, Y.C., Shimizu, C., Fuda, H., Strott, C.A. 2001. Cholesterol and hydroxycholesterol sulfotransferases: identification, distinction from dehydroepiandrosterone sulfotransferase, and differential tissue expression. Endocrinology 142, 2978-2984.
Kakuta, Y., Pedersen, L.G., Carter, C.W., Negishi, M., Pedersen, L.C. 1997. Crystal structure of estrogen sulphotransferase. Nat. Struct. Biol. 4, 904-908.
Kakuta, Y., Sueyoshi, T., Negishi, M., Pedersen, L.C. 1999. Crystal structure of the sulfotransferase domain of human heparan sulfate N-deacetylase/ N-sulfotransferase 1. J. Biom. Chem. 274, 10673-10676.
Kamimura, K., Rhodes, J.M., Ueda, R., McNeely, M., Shukla, D., Kimata, K., Spear, P.G., Shworak, N.W., Nakato, H. 2004. Regulation of Notch signaling by Drosophila heparan sulfate 3-O sulfotransferase. J. Cell Biol. 166, 1069-1079.
Kamimura, K., Koyama, T., Habuchi, H., Ueda, R., Masu, M., Kimata, K., Nakato, H. 2006. Specific and flexible roles of heparan sulfate modifications in Drosophila FGF signaling. J. Cell Biol. 174, 773-778.
protein-protein interactions. Chem. Biol. 7, R57-R61.
Kiehlbauch, C.C., Lam, Y.F., Ringer, D.P. 1995. Homodimeric and heterodimeric aryl sulfotransferases catalyze the sulfuric acid esterification of N-hydroxy-2-acetylaminofluorene. J. Biol. Chem. 270, 18941-18947.
Kohjitani, A., Fuda, H., Hanyu, O., Strott, C.A. 2008. Regulation of SULT2B1a (pregnenolone sulfotransferase) expression in rat C6 glioma cells: relevance of AMPA receptor-mediated NO signaling. Neurosci. Lett. 430, 75-80.
Kushe-Gullberg, M., Kjellén, L. 2003. Sulfotransferases in glycosaminoglycan biosynthesis. Curr. Opin. Struct. Biol. 13, 605-611.
Lander, A.D., Stipp, C.S., Ivins, J.K. 1996. The glypican family of heparan sulfate proteoglycans: major cell-surface proteoglycans of the developing nervous system. Perspect. Dev. Neurobiol. 3, 347-358.
Lin, E.S., Yang, Y.S. 2000. Nucleotide binding and sulfation catalyzed by phenol sulfotransferase. Biochem. Biophys. Res. Commun. 271, 818-822.
Lindsay, J., Wang, L.L., Li, Y., Zhou, S.F. 2008. Structure, function and polymorphism of human cytosolic sulfotransferases. Curr. Drug Metab. 9, 99-105.
Liu, M.Y., Yang, Y.S., Sugahara, T., Yasuda, S., Liu, M.C. 2005. Identification of a
novel zebrafish SULT1 cytosolic sulfotransferase: cloning, expression, characterization, and developmental expression study. Arch. Biochem. Biophys. 437,
10-19.
Liyou, N.E., Buller, K.M., Tresillian, M.J., Elvin, C.M., Scott, H.L., Dodd, P.R., Tannenberg, A.E., McManus, M.E. 2003. Localization of a brain sulfotransferase, SULT4A1, in the human and rat brain: an immunohistochemical study. J. Histochem. Cytochem. 51, 1655-1664.
Longcope, C., Bourgct, C., Flood, C. 1982. The production and aromatization of DHEA in post-menopausal women. Matyritas. 4, 325-332.
Longcope, C. 1996. Dehydroepinandrosterone metabolism. J. Endocrinol. 150, S125-S127. Marshall, A.D., Darbyshire, J.F., Hunter, A.P., McPhie, P., Jakoby, W.B. 1997. Control of activity through oxidative modification at the conserved residue Cys66 of aryl sulfotransferase IV. J. Biol. Chem. 272, 9153-9160.
Meinl, W., Meerman, J. H., Glatt, H. 2002. Differential activation of promutagens by alloenzymes of human sulfotransferase 1A2 expressed Salmonella typhimurium. Pharmacogenetics 12, 677-689.
Meloche, C.A., Falany, C.N. 2001. Expression and characterization of the human 3 beta-hydroxysteroid sulfotransferases (SULT2B1a and SULT2B1b). J. Steroid Biochem. Mol. Biol. 77, 261-269.
Michejda, C.J., Kroeger-Koepke M.B. 1994. Carcinogen activation by sulfate conjugate formation. Adv. Pharmacol. 27, 331-363.
Mishiro, E., Sakakibara, Y., Liu, M.C. 2006. Suiko M. Differential enzymatic characteristics and tissue-specific expression of human TPST-1 and TPST-2. J. Biochem. 140, 731-737.
Monigatti, F., Hekking, B., Steen, H. 2006. Protein sulfation analysis-A primer. Biochim. Biophys. Acta. 1764, 1904-1913.
Näsman, B., Olssen, T., Bäckström, T., Eriksson, S., Grankvist, K., Viitanen, M,, Bucht, G. 1991. Serum dehydroepiandrosterone sulfate in Alzheimer’s disease and in multi-infarct dememtia. Bio. Psychiatry. 30, 684-690.
Negishi, M., Pedersen, L.G., Petrotchenko, E., Shevtsov, S., Gorokhov, A., Kakuta, Y., Pedersen, L.C. 2001. Structure and function of sulfotransferases. Arch. Biochem. Biophys. 390, 149-157.
Nowell, S., Falany, C.N. 2006. Pharmacogenetics of human cytosolic sulfotransferases. Oncogene 25, 1673-1678.
Orentreich, N., Brind, L.J., Rizer, R.L., Vogelman, J.H., Andres, R., Baldwin, H. 1992. Long-term longirudinal measurement of plasma dehydroepiandrosterone sulfate in normal men. J. Clin. Endocrinol. Metab. 75, 1002-1004.
Otterness, D.M., Wieben, E.D., Wood, T.C., Watson, W.G., Madden, B.J., McCormick, D.J., Weinshilboum, R.M. 1999. Human liver dehydroepiandrosterone sulfotransferase: Molecular cloning and expression of cDNA. Mol. Pharmacol. 41, 865-872.
Ouyang, Y., Lane, W.S., Moore, K.L. 1998. Tyrosylprotein sulfotransferase: purification and molecular cloning of an enzyme that catalyzes tyrosine O-sulfation, a common posttranslational modification of eukaryotic proteins. Proc. Natl. Acad. Sci. USA. 95, 2896-2901.
Pai, T.G., Ohkimoto, K., Sakakibara, Y., Suiko, M., Sugahara, T., Liu, M.C. 2002. Manganese stimulation and stereospecificity of the Dopa (3,4-dihydroxyphenylalanine)/tyrosine-sulfating activity of human monoamine-form phenol sulfotransferase. Kinetic studies of the mechanism using wild-type and mutant enzymes. J. Biol. Chem. 277, 43813-43820.
Park, B.C., Lee, Y.C., Strott, C.A. 1999. Identification by chimera formation and site-selected mutagenesis of a key amino acid residue involved in determining stereospecificity of guinea pig 3-hydroxysteroid sulfotransferase isoforms. J. Biol. Chem. 274, 21562-21568.
Parker, C.R., Falany, C.N., Stockard, C.R., Stankovic, A.K., Grizzle, W.E. 1994. Immunohistochemical localization of dehydroepiandrosterone sulfotransferase in human fetal tissues. J. Clin. Endocrinol. Metab. 78, 234-236.
Pasqualini, J.R., Chetrite, G.S. 2007. Correlation of estrogen sulfotransferase activity and proliferation in normal and carcinomatous human breast. A hypothesis. Anticancer Res. 27, 3219-3225.