中草藥對LPS活化NF-kB抑制機轉之研究
60
0
0
全文
(2) 中草藥對 LPS 活化 NF-κB 抑制機轉之研究. Study of the Suppressive Effect of Chinese Herbs to PS-induced NF-κB Activation Student:Yu-Shan Wu. 研 究 生:吳昱珊. Advisor:Dr. Chiun-Jye Yuan. 指導教授:袁俊傑 博士. 國 立 交 通 大 學 生 化 工 程 研 究 所 碩 士 論 文. A Thesis Submitted to Institute of Biochemical Engineering National Chiao Tung University in partial Fulfillment of the Requirements for the Degree of Master in Biochemical Engineering July 2004 Hsinchu, Taiwan, Republic of China. 中華民國九十三年七月 2.
(3) 中草藥對 LPS 活化 NF-κB 抑制機轉之研究 學生:吳昱珊. 指導教授:袁俊傑 博士 國立交通大學生化工程研究所碩士班. 中文摘要 轉錄因子 NF-κB 在免疫發炎反應、細胞的增生、分化與癌化等 生理反應中扮演了關鍵性的角色,且也與許多疾病的發生有相關性。 NF-κB 平時會與其調控蛋白 IκB 結合,而形成一個不活化的複合體存 在細胞質中,當受到外來的刺激,IκB 會降解而使得 NF-κB 會進入細 胞核,進而活化下游基因的轉錄(transcription)。 在此研究中,我們利用實驗室中先前完成的,能穩定持有可檢 驗 NF-κB 活化與否的蟲螢光酵素報導基因的小鼠巨噬細胞株 (RAW264.7) ,來篩檢中草藥內可以調控免疫發炎反應的有效成分。 在篩檢一系列的中草藥初萃層後,發現編號 19-3 的中草藥初萃層能 有效的抑制內毒素(LPS)活化 NF-κB 的作用。 為了進一步的了解此現象的分子調控機轉,設計了一系列的實 驗做研究。初步發現,中草藥初萃層 19-3 抑制內毒素活化 NF-κB, 不是經由降解 NF-κB 的調控蛋白 IκB 所造成。進一步實驗發現,中 草藥初萃層 19-3 可能經由直接或間接的去調控 NF-κB 的活化,其會 i.
(4) 影響細胞內 NF-κB 的分部情形。綜合以上,在此研究中,我們篩檢 了一系列的中草藥後僅發現初萃層 19-3 能抑制 LPS 活化 NF-κB,而 在分子機轉方面,還需由進一步的實驗加以探討。. ii.
(5) Study of the Suppressive Effect of Chinese Herbs to LPS-induced NF-κB Activation. Student : Yu-Shan Wu. Advisor : Dr. Chiun-Jye Yuan. Institute of Biochemical Engineering National Chiao Tung University. Abstract NF-κB is an important transcription factor regulating numerous genes involved in immune and inflammatory responses, cell growth, cell survival, apoptosis, and tissue differentiation. It is also suggested to be involved in the pathogenic progression of many diseases. In resting cells, NF-κB is maintained in an inactive form in the cytoplasm through interaction with inhibitory protein, IκBs. Upon receipt of an appropriate signal, NF-κB is release from IκB and translocate to the nucleus, activate target gene transcription. In this project, we utilized a RAW (p4XNFκB-Lucneo) stable clone generated in our laboratory to screen for the bioactive components in Chinese herbs that were documented to exhibit anti-inflammatory activity. We found that the Chinese herb extract 19-3 could significantly reduce LPS-induced NF-κB activation. The molecular mechanism of Chinese herb extract 19-3 was also partially elucidated by Western blot analysis. Preliminary result shows that 19-3 inhibits LPS-induced NF-κB activation is not involved in suppressing IκBα degradation. Further studies shows that 19-3 may mediate the transactivation of NF-κB iii.
(6) directly or indirectly. The subcellular distribution of NF-κB was also affected by the treatment of 19-3. In conclusion, we screened several Chinese herbs and found at least one fraction from Chinese herb 19-3 exhibited a suppressive activity to LPS-induced NF-κB activity. The molecular mechanism of 19-3 needs to be further elucidated.. iv.
(7) Acknowledgment 感謝指導教授 袁俊傑老師在這兩年內對我的指導與教誨,包容 我在實驗上與生活上的成長,也指導我在實驗上正確的觀念與態度, 也感謝口試委員 廖光文老師 吳文勉老師及 徐祖安老師於口試時給 予的寶貴意見,除了讓我的論文更加完善,也讓我在未來的研究方 向、人生觀上有更成熟的思維,感謝大華技術學院 何姣青老師,除 提供我實驗上的材料外,也不斷在實驗與生活上給我支持與鼓勵。 感謝實驗室中所有成員,讓我這兩年的研究生生涯多采多姿, 實驗室的學長姊 柏文、韻舒、舜評、柏翰、俊炫、威震對我在實驗 與生活上無維不至的照顧,共患難的同學 政剛、欣怡、珮玲、宗翰, 學弟妹 元碩、岳縉、叔青、宜芳,因為你們,讓我的研究生生活更 加完整且充滿回憶。 最後,也最重要的,謝謝我的家人 爸爸、媽媽、昱嫺及俊宏, 這兩年對我的支持、包容與鼓勵,謝謝我的朋友,因為你們,讓我有 動力與勇氣走到這裡。兩年的研究生活,回憶起來,心中滿是感謝, 謝謝每一位陪在我身邊的人。. v.
(8) Contents 中文摘要..................................................................................................... i Abstract ..................................................................................................... iii Acknowledgment ........................................................................................v Contents .................................................................................................... vi Figure Contents....................................................................................... viii Appendix Contents ................................................................................... ix Introduction.................................................................................................1 NF-κB and its regulation ..........................................................2 Biomedical importance of NF-κB ............................................4 The physiological functions of NF-κB.....................................5 Activation of NF-κB by bacterial lipopolysaccharide..............6 Chinese medicinal herbs ...........................................................9 Materials and Methods..............................................................................10 Cell lines .................................................................................10 Reagents..................................................................................10 Plasmids and stable clone .......................................................11 Cell culture and drug treatment ..............................................11 Luciferase assay......................................................................12 MTT assay ..............................................................................13 Determine cell apoptosis using flow cytometry .....................14 Western Blotting .....................................................................14 Cell fractionation ....................................................................16 DNA-protein binding affinity assay .......................................16 vi.
(9) Results.......................................................................................................18 The induction of NF-κB by lipopolysaccharide .....................18 Screening for bioactive fractions from Chinese herbs ...........19 Time- and dose-dependent effect of Chinese herb .................20 Cytotoxicity of 19-3 to RAW264.7 ........................................21 The molecular mechanism of suppressive effect of 19-3 .......22 Discussion .................................................................................................25 References.................................................................................................28. vii.
(10) Figure Contents Fig. 1.. Time response of LPS from Escherichia coli strain on NF-κB. activity. ..............................................................................................35 Fig. 2. Dose response of LPS from Escherichia coli strain on NF-κB activity. ..............................................................................................36 Fig. 3. Effect of various fractions from Chinese herbs to LPS-induced NF-κB activation...............................................................................37 Fig. 4. Dose-dependent effect of 19-3 in the LPS-induce NF-κB activation. ..........................................................................................38 Fig. 5.. Time-dependent effect of 19-3 in LPS-induced NF-κB. activation. ..........................................................................................39 Fig. 6.. Time-dependent effect of 19-3-1, 19-3-2, and 19-3-3 in. LPS-induced NF-κB activation.........................................................40 Fig. 7.. The Cytotoxicity of 19-3. ...........................................................41. Fig. 8. Flow cytometric analysis of the cytotoxicity of 19-3 on RAW264.7.........................................................................................42 Fig. 9.. Western blotting of IκBα and NF-κB p65 before and after. treatment............................................................................................43 Fig. 10.. Western blotting of IκBα and NF-κB p65 in different cell. fractions before and after treatment. .................................................44 Fig. 11. DNA-protein binding affinity assay to quantify the p65 bind to the specific binding site before and after treatment. .........................45. viii.
(11) Appendix Contents Appendix 1.. The NF-κB/IκB family of proteins...................................46. Appendix 2.. Genes were known to be regulated by NF-κB..................47. Appendix 3.. Model of TLR mediated signaling pathways....................48. Appendix 4.. A schematic model of NF-κB activation ..........................49. ix.
(12) Introduction NF-κB (nuclear factor of kappa B) was first identified in 1986 as a nuclear factor bound to an enhancer element of the immunoglobulin κ light chain gene and initially thought to be specifically expressed in mature B- and plasma cells but not in pre B-cells (Sen and Baltimore, 1986a). Later, it was found to be existed ubiquitously in almost all cells. It is involved in the control of a variety of cellular processes, such as immune and inflammatory responses, cell survival, cell proliferation, and apoptosis. Several diseases, including cancer, arthritis, chronic inflammation, autoimmunity, asthma, neurodegenerative diseases, atherosclerosis, and Alzheimer’s disease, are also found to be caused by the activation of NF-κB. The activation of NF-κB was implicated to be independent of protein synthesis (Sen and Baltimore, 1986b). In resting cells, NF-κB is maintained in an inactive form in the cytoplasm through interaction with an inhibitory protein, IκB. Upon reception of appropriate signals, include proinflammatory cytokines, viral infection, bacterial lipopolysaccharide, phorbol esters and okadaic acid, NF-κB is released from IκB and translocates to the nucleus. Specific NF-κB binding site in promoters and enhancers of a big number of inducible genes have been identified with the general consensus sequence of GGGRNNYYCC (R is purine, Y is pyrimidine, and N is any base) (Chen et al., 1999).. 1.
(13) NF-κB and its regulation To date, five NF-κB subunits have been identified in mammalian cells, RelA (also known as p65), c-Rel, RelB, NF-κB1 (p50 and its precursor p105), and NF-κB2 (p52 and its precursor p100) (Appendix 1). All of these subunits contain a highly conserved N-terminal 300-amino acid region, called the Rel homology domain (RHD). RHD contains a phosphotylation site for PKA about 25 amino acid N-terminal to the nuclear localization sequence (NLS). The RHD is required for dimmer formation, interaction with the inhibitory IκB proteins, nuclear translocation and DNA binding. NF-κB subunits can be divided into two subclasses according to their structures and functions. RelA (p65), RelB and c-Rel contain a C-terminal transcription activation domain (TD), which is less conserved at the sequence level across species, even though they can activate the transcription of same set of genes in a variety of species. In contrast, NF-κB1 (p50 and its precursor p105) and NF-κB2 (p52 and its precursor p100) lack TD, therefore, acting as regulatory subunits of other subunits with TD. The long C-terminal domain of NF-κB1 and NF-κB2 contain multiple copies of ankyrin repeats, which act to inhibit the activity of RelA, RelB and c-Rel by association. All vertebrate NF-κB proteins can form homo- or heterodimers, except for RelB, which can only form heterodimers. NF-κB commonly refers to the RelA/NF-κB1 heterodimer (p65/p50 complex), which is one of the most forming dimers and is the major NF-κB complex in most 2.
(14) cells. The composition of differ NF-κB dimers and the sequence context of NF-κB binding sites in different promoters contribute to their differential specificities of gene activation. NF-κB1 is the major partner of RelA, which is required for normal embryogenesis. Mice lacking p50 and p105 exhibit multiple defects in the function of the immune system (Sha et al., 1995). NF-κB2 expression is restricted to the epithelium of the stomach and certain areas of hemopoietic organs such as the thymic medulla, the marginal zone and periarterial sheath of the spleen (Attar et al., 1997). NF-κB2 is the major dimmer partner of RelB, another NF-κB family member important for the function and development of dendritic cells. RelB expression is normally confined to dendritic cells and B lymphocytes. Mice lacking RelB exhibit multiple pathological lesions and defects in acquired and innate immunity (Weih et al., 1995). NF-κB exists in the cytoplasm as an inactive form by associating with inhibitory IκB proteins. This class of inhibitory proteins includes IκBα, IκBβ, IκBγ, IκBε, p100, p105, Bcl-3 and the recently described inducible-IκBξ (Amy et al., 2002). These inhibitory proteins exhibit different affinities for individual NF-κB complex, and are expressed in a tissue-specific manner. IκBα, the most extensively characterized IκB protein, the primary regulator of rapid signal induced activation of NF-κB. Each IκB isoform consist of a conserved ankyrin repeat domain, which mediates the protein-protein interaction with NF-κB. The interaction between NF-κB and IκBα not only blocks the ability of NF-κB to bind to DNA, but also prevent NF-κB heterodimer from entering the uncleus due to the existence of a strong nuclear export signal (NES) in IκBα. Thus, in the resting state, the NF-κB/IκBα complex shuttles continuously between 3.
(15) the nucleus and the cytoplasm with the rate of export exceeds the rate of import, resulting in the cytoplasmic residence of the NF-κB/IκBα. The degradation of IκBα by ubiquitin-base proteosome uncovers a NLS in each subunit of the NF-κB dimer and allows the dimer to translocate from cytoplasm to the nucleus. Upon nuclear translocation, NF-κB dimer binds to the specific DNA sequence (called κB site) and activate NF-κB-dependent gene expression.. Biomedical importance of NF-κB NF-κB regulates the transcription of a large number of genes, especially those participate in immune and inflammatory responses (Siebenlist et al., 1994; Barnes et al., 1997). NF-κB activation also functions in the antiviral responses through regulation of interferons expression (Stark et al., 1998). Many viruses, such as human immunodeficiency virus type 1 (HIV-1) and human T-cell leukemia virus type 1 (HTLV-1), utilize NF-κB to activate their own genes and to stimulate the survival and proliferation of the lymphoid cells, in which they replicate without induce the expression of interferons (Roulston et al., 1995; Bex and Gaynor, 1998). Although NF-κB acts in most cells to suppress apoptosis, under certain circumstances and in certain cell types, however, it may act to induce apoptosis (Sonenshein, 1997; Baichwal and Baeuerle, 1997). Inappropriate regulation of NF-κB contributes to a wide range of human disorders, including cancers, arthritis, chronic inflammation, autoimmunity, asthma, neurodegenerative diseases, 4.
(16) atherosclerosis, and Alzheimer’s disease.. The physiological functions of NF-κB NF-κB can be activated by a variety of stimuli, including inflammatory cytokines (TNF-α, IL-1, IL-2, and IL-12), viral products (HIV-1 and HTLV-1), bacterial toxins (LPS and exotoxin B), stress (UV light and gamma-irradiation), growth factors and apoptotic mediators. Genes positively regulated by NF-κB, are proinfammatory cytokines, chemokines, adhesion molecues, proteases and inflammatory enzymes (Amy et al., 2002, also see Appendix 2). B-cells isolated from p50 knock out mice do not differentiate in response to LPS (Sha et al., 1995). It appears that p50 plays an important role in the immune responses to acute inflammation. The c-Rel-/- mice failed to induce an effective humoral and cellular immune responses to various pathogens. Proliferation defects as well as decreased production of cytokines and immunoglobulin were also observed in T- and B-cells isolated from c-Rel-/- mice. Macrophages from the c-Rel-/- mice killed bacteria poorly. The TNF-α production in response to LPS was reduced, while the expression of GM-CSF, G-CSF, and IL-6 were elevated. Furthermore, the c-Rel-/- mice were more susceptible to influenza virus and leishmania than wild type animals (Baeuerle and Baltimore, 1996). The RelA knock out mice die early during embryonic development due to massive apoptosis of hepatocyte. This observation reveals a role of NF-κB in controlling apoptosis. Thus, NF-κB may play an important role 5.
(17) in protecting cells from apoptosis (Beg et al., 1995). NF-κB also responds to oxidative stress, such as reactive oxygen species (ROS). The activation of NF-κB by ROS can be inhibited by reducing agents and ROS scavengers, such as dithiocarbamates and N-acetyl-L-cysteine (Schreck et al., 1992).. Activation of NF-κB by bacterial lipopolysaccharide Lipopolysaccharide (LPS), the major component of the outer surface of Gram-negative bacteria, is a potent activator of the immune and inflammatory responses, including inflammatory cytokine release, and systemic changes known as septic shock (Schletter et al., 1995; Wenzel et al., 1996). Gram-negative sepsis, a condition that results in approximately 200,000 annual deaths in the Untied States, is believed to be initiated by the stimulation of LPS (Pinner et al., 1996). In Taiwan, septicemia is the thirteenth of the main causes of death for the year 2002. The death rate of septicemia is 30-50 %. Identifying the downstream signaling pathways derived from LPS stimulation, hence, extremely important to understand the cellular mechanism of Gram-negative septic shock. Searching for compounds that can suppress LPS-induced septicemia becomes one of the major trends in the pharmaceutical industry. LPS has been shown to initiate multiple intracellular signaling events, especially the activation of NF-κB, which ultimately leads to the synthesis and release of a number of proinflammatory cytokines, 6.
(18) including interleukin-1 (IL-1), interleukin-6 (IL-6), interleukin-8 (IL-8), and tumor nucrosis factor (TNF) (Schletter et al., 1995). Although the regulation of NF-κB has been intensively studied, most works about NF-κB pathways were focus on those originating from TNF-α, LPS and IL-1 receptors. The mammalian IL-1 receptor (IL-1R) is one of the toll family members (Belvin et al., 1996), mediating the innate immune response in vertebrates and insects. The activation of monocytes and macrophages by LPS is mediated by a cell surface receptor, known as membrane CD14. CD14, a glycosylphosphatidylinositol-linked membrane protein, is widely expressed in mononuclear cells and is considered as a high affinity receptor for LPS (Ingalls et al., 1999) . However, due to lacking a cytoplasmic domain, CD14 has been questioned as how to transmit extracellular signals to downstream intracellular signaling pathways. Recent studies suggest that a toll-like receptor (TLR) can be activated by LPS (Medzhitov et al., 1997; Yang et al., 1998). Human embryonic kidney cells, HEK293, a LPS-unresponsive cell line, is enabled to respond to LPS stimulation by overexpressing TLR-2. These findings suggest that TLR-2 can be a candidate for LPS receptor (Frank et al., 1999). Other studies in the LPS-resistant C3H-HeJ mice implicates that TLR4 is a signal transducing component in the LPS receptor complex (Kirschning et al., 1998). In HEK293 cells, LPS-induced TLR4-mediated activation can be blocked by E5531, a LPS antagonist, in a dose dependent manner. These studies demonstrate that TLR4 is involved in the LPS signaling pathway and serves as a cell-surface co-receptor for CD14 (Jesse et al., 1999). A model has been proposed to explain the 7.
(19) signaling of LPS as illustrated in Appendix 3. The binding of LPS to CD14 is enhanced by a plasma LPS-binding protein (MD-2) (Ulevitch et al., 1994). LPS forms a complex with MD-2, which is subsequently recognized by the soluble CD14 in the circulation. The trimeric complex of LPS, MD-2, and soluble CD14 then binds and activates TLR4 (Faure et al., 2000). The cytoplasmic domain of TLR is highly homologous to the signaling domain of the IL-1 receptor (Gay and Keith, 1991; Heguy et al., 1992). This domain is later referred to as the Toll/IL-1R homology (TIR) domain. Previous study shown, that LPS receptor and IL-1 receptor may share similar signaling components to activate NF-κB (Zhang et al., 1999). Both TLR4 and IL-1R recruit and activate an adaptor protein myeloid differentiation factor 88 (MyD88) (Wesche et al., 1997), which associats with the TIR domain and recruits two other distinct putative serine threonine kinases, namely IL-1 receptor-associated kinase 1 (IRAK1) and IRAK2, to the receptor complex (Muzio et al., 1997). Subsequently, IRAK1 and IRAK2 autophosphorylate and dissociate from the receptor complex. The activated IRAKs then activate TNF receptor-associated factor 6 (TRAF6) (Muzio et al., 1998), which in turn activates downstream kinase, NF-κB-inducing protein kinase (NIK) (Song et al., 1997). NIK activates the IκB kinase complex (IKKα and IKKβ) that facilitates the degradation of IκB and liberates the active NF-κB.. 8.
(20) Chinese medicinal herbs The pharmacology and clinical application of traditional Chinese medicinal herbs has been well documented for centuries in China. Some Chinese herbs are effective in promoting good health and vitality and in curing various diseases. For example, Codonopsis pilosula (Dangshen) has an important effect on antiaging, it has been employed for treatment of dyspepsia, poor appetite, fatigue, psychoneurosis and various diseases (Jin et al., 1996). A large number of traditional Chinese medicinal herbs have been documented to be involved in the modulating immune responses. The molecular mechanism of most of these Chinese herbs of immune modulating ability is, however, still unknown.. Although there are several signaling pathway that lead to immune-response modulation and activation, transcription factor NF-κB plays an important role in the expression of a number of cellular genes, especially those involved in immune and inflammatory responses. Hence, NF-κB is often used as a primary target to screen for anti-inflammatory drugs from a wide range of natural or artificial resources. In this project, we utilized a RAW (p4XNFκB-Lucneo) stable clone generated previously in our laboratory (Huang, 2003) to screen for the bioactive components in Chinese herbs exhibiting anti-inflammatory activity. The molecular mechanism of these bioactive components will be further elucidated in this research. 9.
(21) Materials and Methods 1. Materials. Cell lines RAW264.7 cell line was purchased from Food Industrial Research and Development Institute, Hsinchu.. Reagents The crude extracts of various Chinese herbs, were provided by professor Ho, Jiau-Ching of Ta-Haw Institute of Technology. Prestained protein standard marker was purchased from MBI. Bradford’s reagent was purchased from BioRad. Hyperfilm was obtained from Amersham. Polyvinylidine diflutoide (PVDF) and Nylon membranes and ECL reagent were obtained from Milllipore. Antibody specific against NF-κB p65 subunit was purchased from Abcam. Antibody against IκB-α was from Biosource or Santa-Cruz. HRP-conjugated anti-mouse, anti-rabbit and anti-goat IgG secondary antibodies were bought from Chemicon. Fetal bovine serum (FBS) was purchased from Biological. Dulbecco’s modified egale medium (DMEM) was purchased from Hyclone. 100XPenicillin/streptomycin and 0.5 % trypsin-EDTA were obtained from GIBCO. G418 sulfate, lipopolysaccharide (LPS) were from Calbiochem. NP-40 was from PIERCE. Beetle luciferin reagent was from 10.
(22) Promega. All other reagents and chemicals used in the experiments were reagent grade.. Plasmids and stable clone Plasmid and stable clone used in the thesis are list below: (1) p4XNFκB-Lucneo plasmid is constructed previously in our laboratory (Li, 2002). (2) RAW (p4XNFκB-Lucneo) stable clone is constructed previously in our laboratory (Huang, 2003).. 2. Methods. Cell culture and drug treatment RAW264.7 cells were grown in DMEM medium containing 10 % FBS, 100 µg/mL penicillin and 100 U/mL streptomycin at 37 ℃, in a humidified incubator containing 5 % CO2. Cells were sub-cultured for every 2-3 days. Cells were detached from cultural flask with a gently patting. The sub-cultivation ratio is from 1: 3 to 1: 6. The reporter plasmid p4XNFκB-Lucneo stably transfected RAW264.7, RAW (p4XNFκB-Lucneo), in the T-75 flask were harvested and dispensed into a 24-well cultural plate (4×105 cells/well). Cells were 11.
(23) cultured in a 37 ℃ incubator for 16-24 h or until cells reach 80% confluence, followed by treating cells with various concentrations of herb extracts. At the indicated time, cells were harvested and subjected to luciferase assay. Cells without treatment were used the negative control.. Luciferase assay Preparation of cell lysate Following drug treatment, cells were washed once with 1XPBS, pH 7.4. Cell lysate was then prepared by incubating with 50 µL cell lysis buffer (25 mM Tris-HCl, pH 7.8, 2 mM DTT, 2 mM EDTA, 10 % glycerol, and 1 % Triton X-100) under 4 ℃ for 15 min. Cell lysates can be used directly or stored in a -80 ℃ freezer for long. Luciferase assay The luciferase activity assay of cell lysates was performed by mixing 50 μL luciferin reagent (0.47 mM D-luciferin, 2.67 mM MgSO4, 20 mM Tricine, 0.55 mM ATP, 1.07 mM (MgCO3)4-Mg(OH)2·5H2O, 0.27 mM CoA, 33.3 mM DTT, 0.1 mM EDTA) and 10 μL cell lysate in a test tube (12×75 mm). The activity of luciferase was determined measuring the luminescence on a luminometer (Berthold) with a detection protocol:delay 32 seconds, detect 20 seconds and average the relative light units (RLU) within 20 seconds. The results of luciferase assay (RLU) were normalized by the protein concentration of each cell lysate. 12.
(24) Protein assay Protein concentration of cell lysate was determined using Bradford reagent. The assay was performed in a constant reaction protocol by mixing 160 μL ddH2O, 40 μL Bradford reagent, and 1 μL cell lysate. The mixture was vigorous by vortexed and incubated under room temperature for 10 minutes. Determine the absorbance of mixture at wavelength 595 nm on the plate reader (Fluostar, BMG). Calculate the protein concentration of cell lysate through intrapolation of a standard curve plotted on a same machine.. MTT assay The cytotoxicity of drugs and herb extract was determined by MTT assay. RAW264.7 cells were seeded in 24-well culture plates with 5×105 cells/well. After culturing at 37 ℃ for 14-16 h or until cells reach 80% confluence, cells were then treated with various concentrations of herb extracts at 37 ℃. Following incubation, used medium was replaced with 500 µL thiazole blue solution (0.5 mg/mL in DMEM + 10% FBS) into each well and incubation at 37 ℃ for 2.5-3 h. After reaction, stained cells were washed once with 1XPBS, pH 7.4 and deposited solid formazan was dissolved in 1 mL acidic isopropanol. Measure the absorbance of dissolved formazan of each well at wavelength 570 nm. The spectrophotometer was calibrated with acidic isoproparnal at O.D. 570 nm. 13.
(25) Determine cell apoptosis using flow cytometry The cell apoptosis induced by herb extract was determined by flow cytometry. The apoptotic cells were labeled using Annexin-V apoptosis kit (Gene Research Lab.). RAW264.7 cells were grown in a T-75 cultural flask to about 2×107 cells (14-16 h are usually needed). Following treatment with herb extract (at 37 ℃ for 2 h), cells were suspend using 1 mL 0.025% trypsin/EDTA, followed by gentle pipetting. Remove trypsin/EDTA by centrifugation and wash cell pellet twice with 1XPBS, pH 7.4. Cell labeling was carried out by following manufacturer’s instruction. Suspend cells in 1 mL 1XAnnexin-V binding buffer (10 mM HEPES/NaOH, pH7.4, 140 mM NaCl and 2.5 mM CaCl2). Mix well and aliquot cells suspension in 100 µL/tube. To each tube, 5 µL Annexin-V FITC solution and 10 µL propidium iodide reagent were added, mixed and incubated at room temperature for 15 min in the dark. After incubation, add 1×Annexin-V binding buffer to each tube to make a final volume of 1 mL. Analyze the labeled cells on flow cytometry (Peakflow) within 1 h.. Western Blotting Grow RAW264.7 cells (5×107 cells) in 100 mm dishes in a humidified incubator for 14-16 h or until cells reach 80% confluence. Following treatment, cells were harvested and incubated with 50 µL RIPA buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1 mM EGTA, 1 mM EDTA, 1% Triton X-100, 1% sodium deoxycholate, 0.1 % SDS, 1 mM PMSF, 10 µg/ml aprotinin, 10 µg/mL leupeptin) on ice for 15 min. The 14.
(26) soluble cell lysate was then separated from undissolved particulates by centrifugation at 13,200 rpm for 10 min. Cell lysates (60-80 µg each) were separated on a 10% SDS polyacrylamide electrophoresis gel (SDS-PAGE). The electrophoresis was performed at 100 Volt for 20 min first, followed at 150 Volt for another 1.5 h. The separated protein bands were transferred to a polyvinylidine difluroide (PVDF) membrane on a Bio-Rad Mini Trans-Blot Electrophoretic Transfer Cell soaked in a transfer buffer (48 mM Tris-HCl, pH 9.2, 39 mM glycine, and 10% methanol). After electro-bloting, PVDF membrane was blocked with 5% skin milk in PBST (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, and 0.05% Tween 20) at room temperature for 1-2 h. The membrane was then incubated with first antibody specifically against NF-κB p65 (1: 1000 dilution), IκB-α (1: 1000 dilution), or β-actin (1: 5000 dilution) at 4 ℃ for overnight or at room temperature for 2 h. Subsequently, the membrane was washed three times with PBST, followed by incubating with horseradish peroxidase (HRP)-conjugated secondary antibody against rabbit IgG (1: 5000 dilution) or against goat IgG (1: 5000 dilution) or against mouse IgG (1: 5000 dilution) at room temperature for another one hour. After incubating and washing three times with PBST, 1 mL ECL was spreaded evenly on the membrane for chemiluminescent reaction. The specific protein bands were visualized by exposing on an X-ray film (Hyperfilm). Protein concentration was determined by Bradford’s reagent (Bio-Rad).. 15.
(27) Cell fractionation RAW264.7 cells (5×108 cells) were grown in a 150 mm cultural dish at 37 ℃ for 14-16 h or until they reached 90% confluence. Following treatment with herb extracts, cells were washed twice with cold 1XPBS, pH 7.4 and harvested by scrapping with plastic scraper. Residue PBS in the tube was collected and removed by brief centrifugation. Resuspend cell pellet in 400 µL hypotonic buffer A (10 mM Tris-HCl, pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM DTT, 1 mM PMSF and 1% protease inhibitor cocktail (Sigma)) and incubated on ice for 15 min. After incubation, add 25 µL 10 % Nonidet P-40 into cell suspension and vigorously vortex for 10 sec. To separate nuclei from soluble cytoplasmic component, the resulting cell lysate was centrifugated at 4 ℃ and 13,200 rpm for 5 min. The supernatant or cytoplasmic fraction was collected and kept at -80 ℃. The nuclei were resuspended in 100 µL extraction buffer B (20 mM Tris-HCl, pH 7.9, 400 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM DTT, 1 mM PMSF and 1% protease inhibitor cocktail). The protein concentrations of nuclear and cytosolic fractions were determined using Bradford reagent (Bio-Rad). Normally, 500-700 µg nuclear fraction protein can be isolated from 150 mm cultural dish with 90% cell confluence.. DNA-protein binding affinity assay To identify and quantify the transcription factors that may bind to the specific responding enhancer element. The DNA-protein binding 16.
(28) affinity assay was perform (Deng et al., 2003). This method is carried out by using streptavidin-conjugated agarose beads to pull down biotinated DNA probe, which can interact with certain nuclear proteins or transcription factors. For each assay, 500 µg nuclear extract proteins were mixed with binding buffer (1XPBS, pH 7.4, containing 1 mM EDTA, 1 mM EGTA, 1 mM DTT, 1 mM PMSF, 1 µg/mL leupeptin, 5 µg/mL aprotinin, 10 mM β-glycerophosphate, and 0.1 mM Na3VO4) and 5 µg double-stranded biotinylated oligonucleotides with NF-κB consensus sequence (5’-biotin-GCCTGGGAAAGTCCCCTCAACT-3’ and 5’-biotin-AGTTGAGGGGACTTTCCCAGGC-3’) in 60 µL 4 % streptavidin-conjugated agarose beads (Sigma) at 4 ℃ with gentle tumbling for 2.5 h. Following incubation, beads were washed three times with ice cold 1XPBS, pH 7.4 and resuspended in 50 µL SDS-PAGE loading buffer. Nuclear proteins bound to the beads were denatured by heating at 95 ℃ for 7 min and loaded onto 10% SDS-PAGE. The NF-κB corresponding element associating proteins were identified through Western Blotting using p65 or IκB-α specific antibody.. 17.
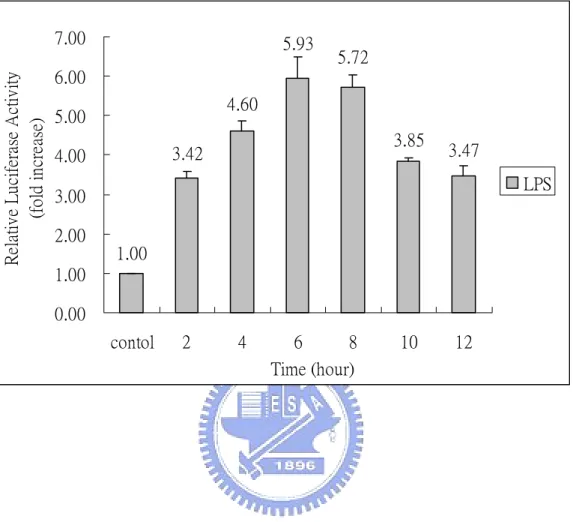
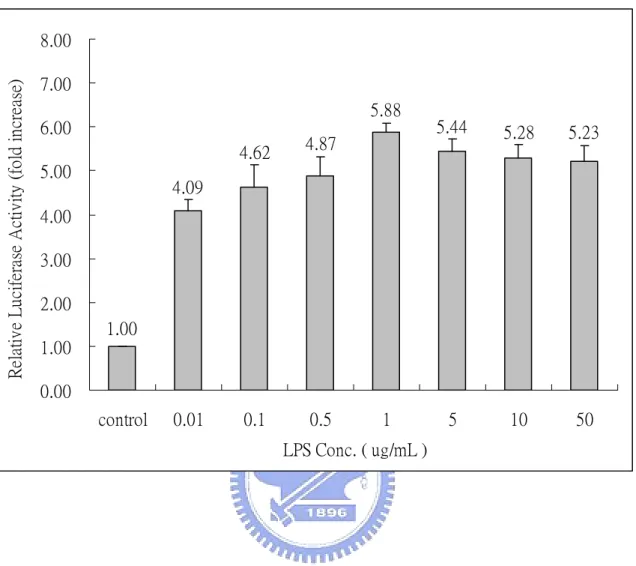
(29) Results The induction of NF-κB by lipopolysaccharide A reporter plasmid p4XNFκB-Lucneo-containing RAW264.7 stable clone, RAW (p4XNFκB-Lucneo), was generated previously in our laboratory (Huang, 2003). It is known that lipopolysaccharide can activate NF-κB in RAW264.7 cells via the Toll-like receptor 4 (TLR4) (Jefferies et al., 2003). We use LPS from E. coli as a stimulant to confirm the effectiveness of this stable clone. Accordingly, RAW (p4XNFκB-Lucneo) cells were treated with 1 µg/mL LPS at 37 ℃ for 2-12 h. As indicated in Fig.1, LPS exhibits a time-dependent stimulatory effect on NF-κB with a maximum activity occurred at 6 h. The activity declined slowly afterward (Fig. 1). RAW (p4XNFκB-Lucneo) cells were further studied by treating with various concentrations of LPS (0.01-50 µg/mL) at 37 ℃ for 6 h. We found that the luciferase activity of the RAW stable clone was 4-6 folds higher than that of the control cells (Fig. 2). Maximal luciferase activity occurred with LPS at concentration of 1 µg/mL. These results confirmed that the generated RAW (p4XNFκB-Lucneo) clone is effective and can be used for drug screening. Various crude extracts from Chinese herbs were screened for their suppressive effect to the LPS-induce NF-κB activation, using this RAW stable clone.. 18.
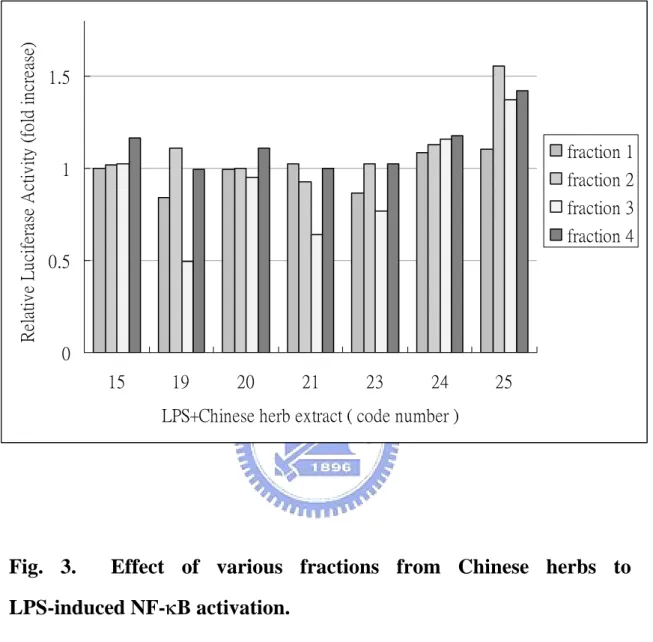
(30) Screening for bioactive fractions from Chinese herbs One of specific goals of this work is to find out bioactive components from Chinese herbs that may suppress LPS-induced NF-κB activation. Hence, about 25 Chinese medicinal herbs, that have been documented to have anti-inflammatory activity including Mimosa pudica L. (Sensitiveplant;含羞草), Rehmanniae radix (Shudihuang;熟地黃), Hairyvein Agrimonia Herba (Xianhecao;仙鶴草), Ligustri Fructus (Nuzhenzi;女貞子), Ecliptae Herba (Mohalian;旱蓮草), Codonopsis radix (Dangshen;黨參), Eriocauli Herba (Gujingcao;穀精草), Glycyrrhizae radix (Gancao;甘草), Rubiae radix (Qiancao;茜草) and Phellodenri cortex (Huangbai;黃柏), were used in this study. The crude extracts of Chinese herbs were fractionated into four fractions based on their polarity. These fractions of various Chinese herbs were obtained from professor Ho, Jiau-Ching of the Ta-Haw Institute of Technology. RAW (p4XNFκB-Lucneo) cells were treated with LPS (1 µg/mL) and various fractions of Chinese herbs (100 µg/mL) at 37 ℃ for 6 h. Fig. 3 showed the effect of various fractions from some Chinese herbs on the LPS-induced NF-κB activation. We found that herb fractions 19-3 and 21-3 could suppress LPS-induced NF-κB activation by 60% and 40%, respectively. Fraction 19-3 is then chosen for the studies of its molecular mechanism in the suppression of LPS-induced NF-κB activation.. 19.
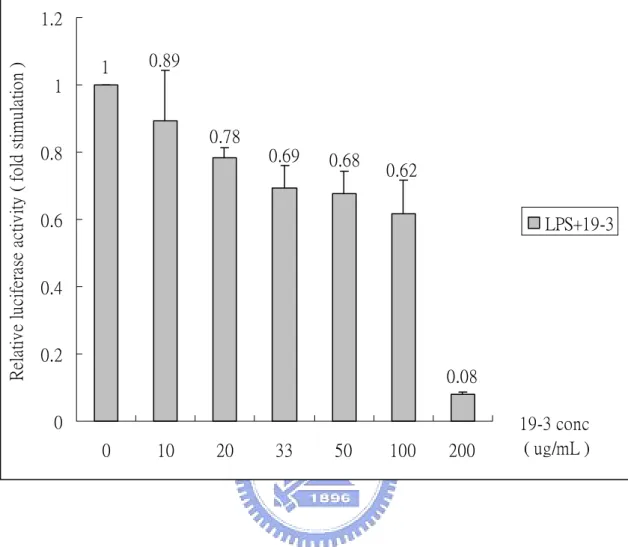
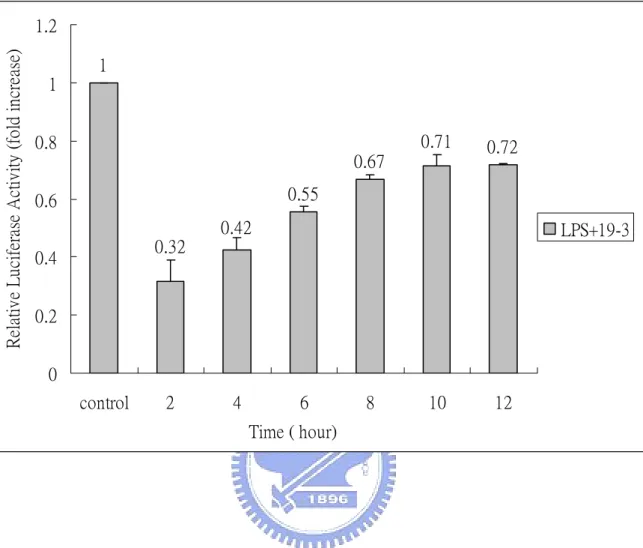
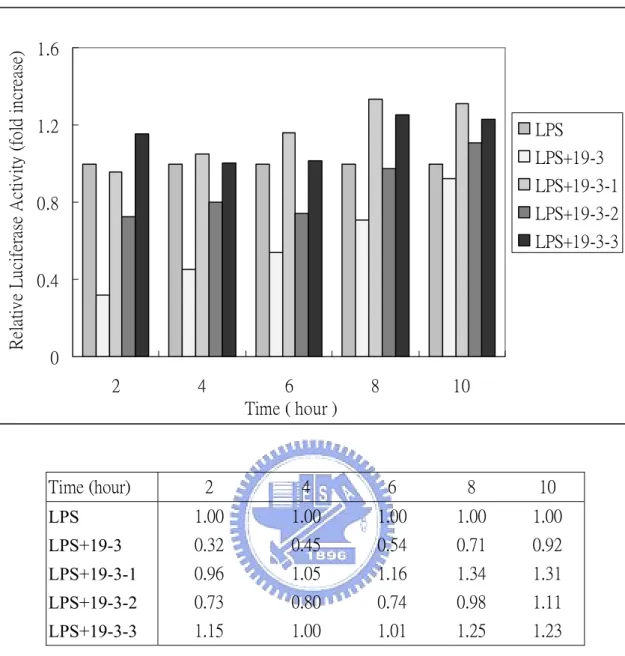
(31) Time- and dose-dependent effect of Chinese herb We first determine the dose- and time- dependent effect of fraction 19-3. Fig. 4 shows dose-dependent effect of fraction 19-3 in the suppression of LPS-induce NF-κB activation. RAW (p4XNFκB-Lucneo) cells were treated with fix concentration of LPS (1 µg/mL) in the presence of various concentrations of fraction 19-3 (10-200 µg/mL) at 37 ℃ for 6 h. We found that the relative luciferase activity of RAW (p4XNFκB-Lucneo) was suppressed with increasing concentration of 19-3. With 19-3 at concentration of 100 µg/mL, the LPS-induced NF-κB activity was suppressed about 40% compared with that of the control cells (treated with 1 µg/mL LPS only). Notably, when increase concentration of 19-3 to 200 µg/mL, the LPS-induced, NF-κB-dependent luciferase activity of RAW (p4XNFκB-Lucneo) cells was almost completely suppressed (8% relative to the control) However, cells were not quite normal under this condition. Time-dependent effect of 19-3 was further studied by treating RAW (p4XNFκB-Lucneo) with LPS (1 µg/mL) and 19-3 (100 µg/mL) at 37 ℃ for 2-12 h. As shown in Fig. 5, 19-3 can suppress LPS-induced NF-κB-dependent luciferase activity at time point as early as 2 h (about 70% suppression). The inhibitory effect of 19-3 decline slowly with the increasing of incubation time. Fraction 19-3 was further fractionated into three subfractions designated as 19-3-1, 19-3-2, and 19-3-3. Fig. 6 shows the time-dependent effect of fractions 19-3-1, 19-3-2, and 19-3-3 in 20.
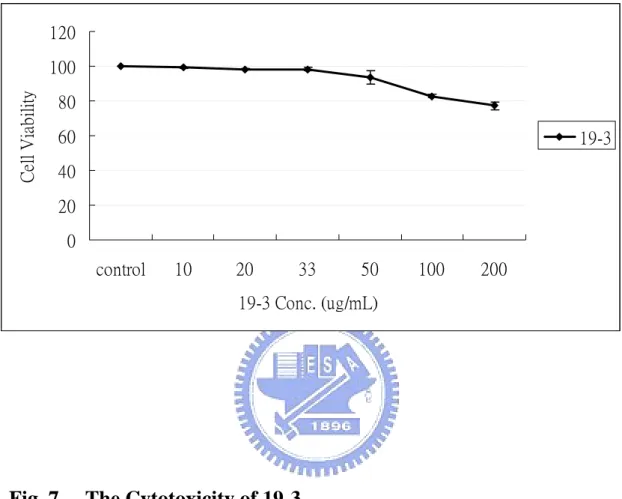
(32) LPS-induced NF-κB activation. Unfortunately, fractions 19-3-1, 19-3-2, and 19-3-3 at 100 µg/mL did not exhibit similar suppression activity as that of 19-3, although 19-3-2 had better suppressive effect than that of 19-3-1 and 19-3-3.. Cytotoxicity of 19-3 to RAW264.7 The cytotoxicity of 19-3 was tested using MTT assay and Annexin V apoptosis assay kit. RAW264.7 cells were seeded in a 24-well culture plate and treated with various concentrations of 19-3 (10-200 µg/mL) at 37 ℃ for 6 h. About 83% cells were still viable in the presence of 100 µg/mL 19-3. The viable cells reduced to about 75% compared with that of control in the presence of 200 µg/mL 19-3. The MTT assay of cells treated with 19-3 also demonstrated that 19-3 may have a low level of cytotoxicity to the RAW264.7 (Fig. 7). The Annexin V biding assay was also performed to investigate whether 19-3 induce apoptosis of RAW264.7 cells. Annexin V is a 35-36 kDa calcium-dependent phospholipids binding protein. It is widely used to detect the loss of phospholipids membrane asymmetry and exposure of phosphatidylserine (PS) at the cell surface, which is an early event in the process that leads to apoptotic cell death. Cells treated with 5 µM staurosporine at 37 ℃ for 4 h was used as a positive control, while cells without treatment were taken as negative control. As shown in Fig. 8, about 95.95% cells in negative control sample are located in Annexin V-negative/PI-negative region, suggesting as the normal cells. In the 21.
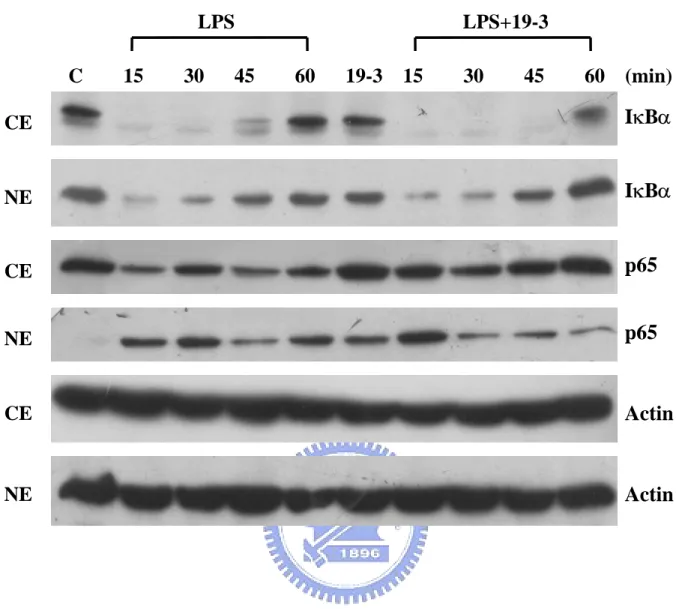
(33) positive control sample (e.g., staurosporine treated RAW264.7 cells), about 14.15% cells are located in Annexin V-positive/PI-negative region (apoptotic cells), while 7.57% cells are present in the PI-positive region (suggesting the necrotic cells). The negative control and positive control were used to find out appropriate condition for further study. As shown in Fig. 8, 19-3 did not induce the apoptosis or necrosis of RAW264.7 cells at the concentration of 100 µg/mL.. The molecular mechanism of suppressive effect of 19-3 Results from previous studies suggest that 19-3 exhibits a suppressive effect to the LPS-induced NF-κB activation. Furthermore, its suppressive effect was not due to its cytotoxicity. Appendix 4 shows a proposed model to explain the signaling of LPS-induced NF-κB activation. It is well known that the activation of NF-κB is initiated by the phosphorylation-dependent degradation of IκB, an inhibitor of NF-κB. In this work, we first want to know whether the inhibitory effect of 19-3 on LPS-induced NF-κB activation is due to the interference of the degradation of IκB. However, we found that 19-3 did not inhibit the LPS-induced IκBα degradation (Fig. 9). Upon the treatment of LPS, IκBα can be degraded within 15 min. The intracellular level of IκBα recovered gradually in cells after treating LPS for 45 min or longer. Chinese herb extract 19-3 alone did not induce IκBα degradation. The NF-B family protein p65 increased slightly 60 min after treatment with LPS or LPS plus 19-3. 22.
(34) Actin was used as an internal control. These findings suggest that 19-3 inhibited the LPS-stimulated NF-κB-activation was not through suppressing phosphorylation dependent proteolysis of IκBα in the cells. NF-κB was initially described as the prototypical p65/p50 heterodimer (Ghosh et al., 1990). Upon activation and IκB-α degradation, p65/p50 complex can translocate to the nucleus and upregulate the corresponding genes. We want to find out whether p65 nuclear translocation is blocked by 19-3. RAW264.7 cells were fractionated into two fractions, cytoplasmic and nuclear fractions, as described in Materials and Methods. Without any treatment, IκBα was distributed in both cytoplasm and nuclear, whereas, NF-κB p65 was only seen in the cytoplasm (Fig. 10). This result is consistent with the finding of other laboratory (Hongyun et al., 2002). The IκBα in both cytoplasm and nucleus were rapidly degraded in response to the treatment of LPS (1 µg/mL). In contrast, NF-κB p65 abruptly appeared in the nucleus in as early as 15 min and steadily present in the nucleus for at least 60 min. Interestingly, we found that NF-κB p65 also abruptly appeared in the nucleus in the presence of both LPS (1 µg/mL) and 19-3 (100 µg/mL), but gradually faded away along with the time. IκBα, on the other hand, exhibited similar responsive patterns in LPS- and LPS/19-3-treated cells, except that after 60 min of treatment the IκBα level is higher in LPS/19-3 treated cells. This result indicates that 19-3 may suppress LPS-induced NF-κB activation through facilitating the export of NF-κB p65 from nucleus. Previous report has demonstrated that newly synthesized IκB can localize to the nucleus and directly regulate with NF-κB-dependent gene 23.
(35) expression by facilitating the dissociation of NF-κB-DNA complexes (Michele et al., 1999). DNA-protein binding affinity assay was then used to investigate whether IκB-α interfere with the interaction of NRE and NF-κB. Since the conventional electrophoretic mobility shift assay (EMSA) is difficult to simultaneously identify and quantify protein complex formed by multiple factor bound to a sequence-specific DNA probe. The DNA-protein binding affinity assay allows to identify and further quantify the transcription factors binding to the specific responding element (Deng et al., 2003). RAW264.7 cells were co-treated with 1 µg/mL LPS and 100 µg/mL 19-3 at 37 ℃ for 15-60 min. At the indicated time intervals, nuclear fractions were prepared and incubated with the biotinated NF-κB consensus sequence and streptavidin-agarose beads for 2.5 h at 4 ℃. The beads were washed and collected by centrifugation, followed by separating on a 10% SDS-PAGE. The NF-κB corresponding element associated protein was identified through Western Blotting using p65 or IκB-α specific antibody. As shown in Fig. 11, only small amount of NF-κB p65 could be seen in the control cells as well as in 19-3 treated cells. In the presence of 1 µg/mL LPS, binding of NF-κB p65 to NRE significant increased. In the presence of 19-3, the LPS (1 µg/mL) treated cells still exhibited similar amount of NF-κB binding on NRE. The reason for this result is not clear so far. Western blotting using IκB-α specific antibody to investigate IκB-α expression, no signal was found (data not shown). These results indicate that IκB-α did not interfere with the binding of NRE and NF-κB. 24.
(36) Discussion RAW (p4XNFκB-Lucneo) cells were generated previously in our laboratory (Huang, 2003). In previous studies, lipopolysaccharied (LPS) was shown to activate NF-κB in RAW264.7 cells via the Toll-like receptor 4 ( TLR4 ) (Jefferies et al., 2003). We utilized LPS from E. coli as a stimulant to confirm that this RAW stable clone (p4XNFκB-Lucneo) was effective for subsequent studies (Fig. 1 and Fig. 2). The early inflammatory response is regulated predominately by the macrophage and is characterized by an increased production of proinflammatory cytokines, mediated, in part, through NF-κB activation (Baeuerle et al., 1996). Aberrant activation of the NF-κB signaling pathway is likely to contribute to the development and progression of various immune and inflammatory disorders (Toshio et al., 2004). We utilized previously generated TF-based testing system, RAW (p4XNFκB-Lucneo) cells, to screen the extracts of various Chinese herbs, and hoped to discover some bioactive components in Chinese herbs that may act as inhibitors of LPS-induced NF-κB activation. Chinese herbs fraction 19-3 was shown in this work to exhibit a suppressive activity to the LPS-induced NF-κB activation. In addition, the molecular mechanism of the suppressive activity of 19-3 to this activity was also elucidated. Western blotting analysis of IκB-α and NF-κB family protein p65 under the stimulation of LPS before and after treatment of Chinese herb fraction 19-3 reveals that 19-3 inhibits LPS-induced NF-κB activation is not due to the suppression of phosphorylation-dependent IκB-α 25.
(37) degradation in the RAW264.7. Instead, 19-3 may inhibit the LPS-induced NF-κB activation through modulation the nuclear localization of NF-κB p65 protein. However, several aspects related to this mechanism remained to be elucidated. First, can IκB-α compete with NRE for the binding of NF-κB? Second, can IκB-α bring NF-κB out of the nucleus ? In first aspects, we utilized DNA-protein binding affinity assay to study DNA binding activity of NF-κB before and after treatment of 19-3. In this study, binding to a specific site in DNA sequence is necessary for the action of transcription factors, and thus represents and obvious target for therapeutic drugs. As noted above, the inhibition of E2F/DP-1 heterodimer actually achieves its effect by preventing DNA-binding of the factor which requires heterodimer formation. A number of DNA-binding drugs such as distamycin have been developed to inhibit DNA replication as a means of cancer therapy (Cozzi, 2003). In protein-protein interaction aspect, interaction of transcription factor with other proteins is important for the regulation of their activity and the mechanism of their action. For example, the newly synthesized IκB can enter the nucleus, directly interfere with gene expression by dissociating protein-DNA complexes, and export NF-κB back to its resting state in the cytoplasm (Thomas, 1999). Experiments will be used to explain whether IκB-α compete with NRE to allow NF-κB dissociate from NRE. In this study, we will utilize Immunoprecipitation (IP) to find out the interaction between NF-κB and IκB-α under various circumstances. On the other hand, numerous traditional Chinese medicinal herbs 26.
(38) have reported to have immunomodulatory properties (Borcher et al., 2000; Borches et al., 1997). Identification of active components from folk medicines is an attractive approach to develop novel therapeutics. Utilizing RAW (p4XNFκB-Lucneo) as a drug testing system, we have screened over 100 different fractions from 30 different Chinese herbs. At least one fraction 19-3 has been shown to have potential to be an anti-inflammatory lead for the further drug development.. 27.
(39) References Amy, K. R., James, F. C., and Simon, M. B. (2002) Small-molecule inhibitors of NF-κB for the treatment of inflammatory joint disease. Current Opinion in Pharmacology 2, 316-32. Attar, R. M., Caamano, J., Carrasco, D., Iotsova, V., Ishikawa, H., Ryseck, R. P., Weih, F. and Bravo, R. (1997) Genetic approaches to study Rel/NF-kappa B/I kappa B function in mice. Semin. Cancer Biol. 8, 93-101. Baeuerle, P. A. and Baltimore, D. (1996) NF-κB: ten year after. Cell 87, 13-20. Baichwal, V. R. and Baeuerle, P. A. (1997) Activate NF-kappa B or die? Curr. Biol. 7, R94-R96. Barnes, P. J. and Baichwal, V. R. (1997) Nuclear factor-κB: A pivotal transcription factor in chronic inflammatory diseases. New Engl. J. Med. 336, 1066-1071. Beg, A. A., Sha, W. C., Bronson, R. T. and Baltimore, D. (1995) Constitutive NF-kappa B activation, enhanced granulopoiesis, and neonatal lethality in I kappa B alpha-deficient mice. Genes Dev. 9, 2736-2746. Beg, A., Sha, W., Bronson, R., Ghosh, S. and Baltimore, D. (1995) Embryonic lethality and liver degeneration in mice lacking the RelA component of NF-κB. Nature 376, 167-170. Belvin, M. P. and Anderson, K. V. (1996) A conserved signaling pathway:the Drosophila toll-dorsal pathway. Annu. Rev. Cell. Dev. Biol. 12, 393-416. 28.
(40) Bex, F. and Gaynor, R. B. (1998) Regulation of gene expression by HTLV-1 Tax proteins. Methods 16, 83-94. Borchers, A. T., Sakai, S., Henderson, G. L., Harkey, M. R., Keen, C. L., Stern, J. S., Terasawa, K. and Gershwin, M. E. (2000) Shosaikoto and other Kampo (Japanese herbal) medicines: a review of their immunomodulatory activities. J. Ethnopharmacol 73, 1-13. Borches, A. T., Hackman, R. M., Keen, C. L., Stern, J. S. and Gershwin, M.. E.. (1997). Complememtary. medicine:. a. review. of. immunomodulatory effects of Chinese herbal medicines. Am. J. Clin. Nutr. 66, 1303-1312. Chen, F. E. and Ghosh, G. (1999) Regulation of DNA binding by Rel/NF-κB transcription factors: structural views. Oncogene 18, 6845-6852. Cozzi, P. (2003) The discovery of a new potential anticancer drug: a case history. Farmaco. 8, 213-220. Deng, W. G., Zhu, Y., Montero, A. and Kenneth, K. W. (2003) Quantitative analysis of binding of transcription factor complex to biotinylated DNA probe by a streptavidin-agarose pulldown assay. Analytical Biochemistry 323, 12-18. Faure, E., Equils, O., Sieling, P. A., Thomas, L., Zhang, F. X., Kirschning, C. J., Polentarutti, N., Muzio, M. and Arditi, M. (2000) Bacterial lipopolysaccharide activates NF-kappaB through toll-like receptor 4 (TLR-4) in cultured human dermal endothelial cells. Differential expression of TLR-4 and TLR-2 in endothelial cells. J. Biol. Chem. 275, 11058-11063. Frank, X. Z., Carsten, J. K., Roberta, M., Xiao, P. X., Yiping, J., 29.
(41) Emmanuelle, F., Alberto, M., Mike, R., Marta, M. and Moshe, A. (1999) Bacterial lipopolysaccharide activates nuclear factor-κB through interleukin-1 signaling mediators in cultured human dermal endothelial cells and mononuclear phagocytes. J. Biol. Chem. 274, 7611-7614. Gay, N. J. and Keith, F. J. (1991) Drosophila Toll and IL-1 receptor. Nature 351, 355-356. Ghosh, S. and Baltimore, D. (1990) Activation in vitro of NF-kappa B by phosphorylation of its inhibitor I kappa B. Nature 344, 678-682.. Heguy, A., Baldari, C. T., Macchia, G., Telford, J. L. and Melli, M. (1992) Amino acids conserved in interleukin-1 receptors (IL-1Rs) and the Drosophila toll protein are essential for IL-1R signal transduction. J. Biol. Chem. 267, 2605-2609. Hongyun, S., Shigang, X., Min, L., Ebrahim, Z., Cecilia, G. and Hidekazu, T. (2002) Iron activates NF-κB in Kupffer cells. Am J Physiol Gastrointest Liver Physiol 283, G719-726. Huang, P. W. (2003) Construction of a transcription factor based drug screening system and the molecular mechanism of PGE1-mediaged NF-IL6 activation. Department of Biological Science and Technology National Chiao Tung University. Ingalls, R. R., Heine, H., Lien, E., Yoshimura, A. and Golenbock, D. (1999). Lipopolysaccharide. recognition,. CD14,. and. lipopolysaccharide receptors. Infect. Dis. Clin. North. Am. 13, 341-353. Jefferies, C. A., Doyle, S., Brunner, C., Dunne, A., Brint, E., Wietek, C., Walch, E., Wirth, T. and O’Neill, L. A. (2003) Bruton’s tyrosine 30.
(42) kinase is a Toll/Interleukin-1 receptor domain-binding protein that participates in Nuclear Factor κB activation by Toll-like receptor 4. J. Biol. Chem. 278, 26258-26264. Jesse, C. C., Donna, W. Y., Douglas, T. G., William, J. C. and Fabian, G. (1999) Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J. Biol. Chem. 274, 10689-10692. Jin, Z. L., Wen, J. and Li, S. F. (1996) Antiaging effect of shenqi mixture tonic. Chin Trad Herb Drugs 27, 90-92. Kirschning, C. J., Wesche, H., Ayres, T. M. and Rothe, M. (1998) Human Toll-like. Receptor. 2 Confers. Responsiveness. to. Bacterial. Lipopolysaccharide. J. Exp. Med. 188, 2091-2097. Li, J. J. (2002) The Development of NF-IL6 and NF-κB based drug screening system and its application on the study of Chinese traditional. herbs.. Department. of. Biological. Science. and. Technology National Chiao Tung University. Medzhitov, R., Preston-Hurlburt, P. and Janeway, C. A. (1997) A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 388, 394-397. Michele, A., Hakju, K., Pierre, G. and John, H. (1999) Identification by in vivo genomic footprinting of a transcriptional switch containing NF-κB and Sp1 that regulates the IκB-α promoter. Molecular and cellular biology 19, 6140-6153 Muzio, M., Natoli, G., Saccani, S., Levrero, M. and Mantovani, A. (1998) The Human Toll Signaling Pathway: Divergence of Nuclear Factor κB and JNK/SAPK Activation Upstream of Tumor Necrosis Factor Receptor-associated Factor 6 (TRAF6). J. Exp. Med. 187, 31.
(43) 2097-2101. Muzio, M., Ni, J., Feng, P. and Dixit, V. M. (1997) IRAK (Pelle) Family Member IRAK-2 and MyD88 as Proximal Mediators of IL-1 Signaling. Science 278, 1612-1615. Pinner, R. W., Teutsch, S. M., Simonsen, L., Klug, L. A., Graber, J. M., Clarke, M. J. and Gerkelman, R. L. (1996) Trends in infectious diseases mortality in the United States J. Am. Med. Assoc. 275, 189-193. Roulston, A., Lin, R., Beauparlant, P., Wainberg, M. A. and Hiscott, J. (1995) Regulation of human immunodeficiency virus type 1 and cytokine gene expression in myeloid cells by NF-kappa B/Rel transcription factors, Microbiol. Rev. 59, 481-505. Schletter, J., Heine, H., Ulmer, A. J. and Rietschel, E. T. (1995) Molecular mechanisms of endotoxin activity. Arch. Microbiol. 164, 383-389. Schreck, R., Albermann, K. and Baeuerle, P. A. (1992) Nuclear factor kappa B: an oxidative stress-responsive transcription factor of eukaryotic cells. Free Radic. Res. Commun. 17, 221-237. Sen, R., and Baltimore, D. (1986a) Inducibility of κ immunoglobulin enhancer-binding protein NF-κB by a post-translational mechanism. Cell 47, 921-928. Sen, R., and Baltimore, D. (1986b) Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell 46, 705-716. Sha, W. C., Liou, H. C., Tuomanen, E. I. and Baltimore, D. (1995) Targeted disruption of the p50 subunit of NF-κB leads to multifocal defects in immune rewsponses. Cell 80, 321-330. Siebenlist, U., Franzoso, G. and Brown, K. (1994) Structure, regulation 32.
(44) and function of NF-κB. Annu. Rev. Cell. Biol. 10, 405-455. Sonenshein, G. E. (1997) Rel/NF-kappa B transcription factors and the control of apoptosis. Semin. Cancer Biol. 8, 113-119. Song, H. Y., Regnier, C. H., Kirschning, C., Goeddel, D. V., Cao, Z. and Rothe, M. (1997) Tumor necrosis factor (TNF)-mediated kinase cascades : Bifurcation of Nuclear Factor-κB and c-jun N-terminal kinase (JNK/SAPK) pathways at TNF receptor-associated factor 2. Proc. Natl. Acad. Sci. U S A. 94, 9792-9796. Stark, G. R., Karin, I. M., Williams, B. R., Silverman, R. H. and Schreiber, R. D. (1998) How cells respond to interferons. Annu. Rev. Biochem. 67, 227-264. Thomas, D. G. (1999) The Rel/NF-κB signal transduction pathway: introduction. Oncogene 18, 6842-6844. Toshio, H., Yoshihiro, W. and Iwao, W. (2004) Protein disulfide isomerase suppresses the transcriptional activity of NF-κB. Biochemical and Biophysical Research Communications. 318, 46-52. Ulevitch, R. J. and Tobias, P. S. (1994) Recognition of endotoxin by cells leading to transmembrane signaling. Curr. Opin. Immunol. 6, 125-130. Weih, F., Carrasco, D., Durham, S. K., Barton, D. S., Rizzo, C. A., Ryseck, R. P., Lira, S. A. and Bravo, R. (1995) Multiorgan inflammation and hematopoietic abnormalities in mice with a targeted disruption of RelB, a member of the NF-kappa B/Rel family. Cell 80, 331-340. Wenzel, R. P., Pinsky, M. R., Ulevitch, R. J. and Young, L. (1996) Current 33.
(45) understanding of sepsis. Clin. Infect. Dis. 22, 407-413. Wesche, H., Henzel, W. J., Shillinglaw, W., Li, W. and Cao, Z. (1997) MyD88: an adapter that recruits IRAK to the IL-1 receptor complex. Immunity 7, 837-847. Yang, R. B., Mark, M. R., Gray, A., Huang, A., Xie, M. H., Zhang, M., Goddard, A., Wood, W. I., Gurney, A. L. and Godowski, P. J. (1998) Toll-like receptor-2 mediates lipopolysaccharide-induced cellular signalling. Nature 395, 284-288. Zhang, F. X., Kirschning, C. J., Mancinelli, R., Xiao-Ping, X., Yiping, J., Emmanuelle, F., Mantovani, A., Rothe, M., Muzio, M. and Arditi, M. (1999) Bacterial Lipopolysaccharide Activates Nuclear Factor-κB through Interleukin-1 Signaling Mediators in Cultured Human Dermal Endothelial Cells and Mononuclear Phagocytes. J. Biol. Chem. 274, 7611-7614.. 34.
(46) Relative Luciferase Activity (fold increase). 7.00. 5.93. 5.72. 6.00 4.60. 5.00. 3.85. 3.42. 4.00. 3.47 LPS. 3.00 2.00. 1.00. 1.00 0.00 contol. Fig. 1.. 2. 4. 6 8 Time (hour). 10. 12. Time response of LPS from Escherichia coli strain on NF-κB. activity. RAW (p4XNFκB-Lucneo) cells were treated with LPS (1 µg/mL) at 37 ℃ for 2-12 h. At the indicated time points, cells were harvested and subjected to the luciferase activity assay. The result of luciferase assay was normalized with total protein used in the assay. Relative activity was determined by comparing with that of control (treated with medium only). Results were the means±S.D. from three separate experiments, each carried out in triplicate.. 35.
(47) Relative Luciferase Activity (fold increase). 8.00 7.00 5.88. 6.00 5.00. 4.62. 4.87. 0.1. 0.5. 5.44. 5.28. 5.23. 5. 10. 50. 4.09. 4.00 3.00 2.00 1.00 1.00 0.00 control. 0.01. 1. LPS Conc. ( ug/mL ). Fig. 2.. Dose response of LPS from Escherichia coli strain on NF-κB. activity. RAW (p4XNFκB-Lucneo) cells were treated with various concentrations of LPS (0.01-50 µg/mL) at 37 ℃ for 6 h. Following treatment, cells were harvested and subjected to the luciferase activity assay. The results of luciferase assay were normalized with total protein used in the assay. Relative activity was determined by comparing with that of control (treated with medium only). Results were the means±S.D. from three separate experiments, each carried out in triplicate.. 36.
(48) Relative Luciferase Activity (fold increase). 1.5. fraction 1 1. fraction 2 fraction 3 fraction 4. 0.5. 0 15. 19. 20. 21. 23. 24. 25. LPS+Chinese herb extract ( code number ). Fig. 3.. Effect of various fractions from Chinese herbs to. LPS-induced NF-κB activation. RAW (p4XNFκB-Lucneo) cells were co-treated with LPS (1 µg/mL) and various Chinese herbs extracts (100 µg/mL) at 37 ℃ for 6 h. Following treatment, cells were harvested and subjected to the luciferase activity assay. Cells incubated with LPS and 1% DMSO were used as control. The results of luciferase assay were normalized with total protein used in the assay and represented relative to the activity of the control cells.. 37.
(49) Relative luciferase activity ( fold stimulation ). 1.2 1. 0.89. 1 0.78. 0.8. 0.69. 0.68. 0.62. 0.6. LPS+19-3. 0.4. 0.2 0.08 0 0. Fig. 4.. 10. 20. 33. 50. 100. 200. 19-3 conc ( ug/mL ). Dose-dependent effect of 19-3 in the LPS-induce NF-κB. activation. RAW (p4XNFκB-Lucneo) cells were treated with LPS (1 µg/mL) and various concentrations of 19-3 (10-200 µg/mL) at 37 ℃ for 6 h. Following treatment, cells were harvested and subjected to the luciferase activity assay. The results of luciferase activity assay were normalized with total protein used in the assay. The results were presented as relative activity compared with that of control (without 19-3). Results were the means±S.D. from three separate experiments, each carried out in triplicate. 38.
(50) Relative Luciferase Activity (fold increase). 1.2 1 1 0.71. 0.8 0.67. 0.72. 0.55. 0.6 0.42. LPS+19-3. 0.32. 0.4 0.2 0 control. 2. 4. 6. 8. 10. 12. Time ( hour). Fig. 5. Time-dependent effect of 19-3 in LPS-induced NF-κB activation. RAW (p4XNFκB-Lucneo) cells were treated with LPS (1 µg/mL) and 19-3 (100 µg/mL) at 37 ℃ for 2-12 h. At the indicated time points, cells were harvested and subjected to the luciferase activity assay. The luciferase activity was normalized with total protein used in each assay. Relative luciferase activity was calculated using the activity of control (without 19-3) as 1. Results were the means±S.D. from three separate experiments, each carried out in triplicate.. 39.
(51) Relative Luciferase Activity (fold increase). 1.6. 1.2. LPS LPS+19-3 LPS+19-3-1. 0.8. LPS+19-3-2 LPS+19-3-3. 0.4. 0 2. Time (hour). 4. 6 Time ( hour ). 8. 10. 2. 4. 6. 8. 10. LPS. 1.00. 1.00. 1.00. 1.00. 1.00. LPS+19-3. 0.32. 0.45. 0.54. 0.71. 0.92. LPS+19-3-1. 0.96. 1.05. 1.16. 1.34. 1.31. LPS+19-3-2. 0.73. 0.80. 0.74. 0.98. 1.11. LPS+19-3-3. 1.15. 1.00. 1.01. 1.25. 1.23. Fig. 6. Time-dependent effect of 19-3-1, 19-3-2, and 19-3-3 in LPS-induced NF-κB activation. RAW (p4XNFκB-Lucneo) cells were co-treated with LPS (1 µg/mL) and 19-3-1, 19-3-2, or 19-3-3 (100 µg/mL) at 37 ℃ for 2-10 h. At the indicated time points, cells were harvested and subjected to the luciferase activity assay. The luciferase activity was normalized with total protein used in each assay. Relative luciferase activity was calculated using the activity of control (treated with LPS) as 1. 40.
(52) 120. Cell Viability. 100 80 60. 19-3. 40 20 0 control. 10. 20. 33. 50. 100. 200. 19-3 Conc. (ug/mL). Fig. 7. The Cytotoxicity of 19-3. RAW264.7 cells were treated with various concentration of 19-3 (10-200 µg/mL) at 37 ℃ for 6 h. Following incubation, the cells were harvested and subjected the MTT assay. Cell viability presented was relative to that of control. Results were the means±S.D. from three separate experiments, each carried out in triplicate.. 41.
(53) control. positive control. 19-3 ( 100 µg/mL ). Fig. 8.. Flow cytometric analysis of the cytotoxicity of 19-3 on. RAW264.7. Cell treated with 5µM staurosporine at 37 ℃ for 4 h was used as a positive control, while cells without treatment were taken as negative control. FL1 was Annexin V and FL3 was PI.. 42.
(54) LPS C. 15. 30. LPS+19-3 45. 60. 19-3. 15. 30. 45. 60 (min) IκBα p65. Actin. Fig. 9. Western blotting of IκBα and NF-κB p65 before and after treatment. Western blot analysis of whole cell lysate from RAW264.7 cells before (C) and after treatment of LPS (1 µg/mL), 19-3 (100 µg/mL), and LPS (1 µg/mL) plus 19-3 (100 µg/mL) was performed as described in Materials and Methods. At the indicated time points, cells were harvested and subjected to cell lysate preparation. Cell lysate (65 µg each) was separated on a 10% SDS-PAGE, followed by Western blot analysis using anti-NF-κB p65, anti-IκBα or anti-β-actin antibody.. 43.
(55) LPS C. 15. 30. 45. LPS+19-3 60. 19-3 15. 30. 45. 60 (min). CE. IκBα. NE. IκBα. CE. p65. NE. p65. CE. Actin. NE. Actin. Fig. 10. Western blotting of IκBα and NF-κB p65 in different cell fractions before and after treatment. RAW264.7 cells were co-treated with LPS (1 µg/mL) and 19-3 (100 µg/mL) at 37 ℃ for 15-60 min. At the indicated time points, cells were harvested and subjected to cell fractionation as described in Material and Methods. The samples (65 µg each) were separated on a 10% SDS-PAGE, followed by Western blotting using anti-NF-κB p65, anti-IκBα or anti-β-actin antibody. The result presented is the representative of three separated experiments. 44.
(56) LPS C. 15. 30. LPS+19-3 45. 60. 25. 22.8 20.7 18.3. 20 fold. 19-3. 15. 30. 45. 60 (min). 22.3 21.4 20.7 20.0 19.9 LPS. 15. LPS+19-3. 10 3.1. 5 1.0 0 control. 19-3. 15. 30. 45. 60. Time ( min ). Fig. 11. DNA-protein binding affinity assay to quantify the p65 bind to the specific binding site before and after treatment. RAW264.7 cells were co-treated with LPS (1 µg/mL) and 19-3 (100 µg/mL) at 37 ℃ for 15-60 min. At the indicated time points, nuclear fraction were prepared as described in Material and Methods. Nuclear fractions were incubated with the biotinylated NF-κB double-stranded oligo and streptavidin-agarose beads at 4 ℃ for 2.5 h. The agarose-bead-bound proteins were determined by Western blotting using anti-NF-κB p65 or anti-IκBα antibody. Each band was analyzed by densitometry (bottom). This result is the representative of two separate experiments. 45.
(57) (Annu. Rev. Immunol. 16:225-260, 1998). Appendix 1. The NF-κB/IκB family of proteins. Membranes of the NF-κB/IκB families are shown. The number of amino acids in each protein is shown on the right. The arrows point to the endoproteolytic cleavage sites of p100/p52 and p105/p50. IκB-ε has been proposed to be translated from either the first methionine or from an internal methionine at position 140. Abbrevation used: RHD, rel homology domain; TD, transactivation domain; LZ, leucine zipper domain of RelB; GRR, glycine-rich region.. 46.
(58) (Current Opinion in Pharmacology 2:316-321, 2002). Appendix 2. Genes were known to be regulated by NF-κB.. 47.
(59) (The journal of Clinical Investigation 107:13-19, 2001). Appendix 3. Model of TLR mediated signaling pathways. TLR4 is activated by Gram-negative bacteria lipopolysaccharied (LPS) or Gram-positive bacterial lipoteichoic acid (LTA). A secreted small molecule MD-2 is essential for TLR4 signaling. Whether bacterial products directly bind to TLRs, or whether they bind through CD14, remains to be fully established. 48.
(60) LPS. TLR4 IL-1RAcP 26S proteosome. MyD88. IκB degradation. IRAK and IRAK2 TRAF6 NIK. ubiquitination. NF-κB activation. P P. IKKα and IKKβ. gene expression. IκB. nucleus. NF-κB cytosol. Appendix 4. A schematic model of NF-κB activation Various stimuli, such as LPS, can activate IKK through the action of adaptors and kinases. Once activated, IKK phosphorylates IκB, leading to polyubiquitinylation of IκB. This modification targets IκB for rapid degradation by the 26S proteosome. IκB degradation express the nuclear localization sequence on NF-κB, resulting in the translocation of NF-κB to the nucleus. In the nucleus, NF-κB regulates transcription of target genes. Some of the NF-κB target genes code for inflammatory mediators, such as TNF-α and IL-1, which can lead to recruitment of additional cells to the inflammatory response. 49.
(61)
數據
+7


相關文件
( D )The main function of fuel injection control system is to _________.(A) increase torque (B) increase horsepower (C) increase fuel efficiency (D) make 3-way catalytic
People of lesser capacities had to learn Hinayana teachings first in order to increase their intellectual power before they turned to Mahayana; the result was the gradual doctrine.
{ As the number of dimensions d increases, the number of points n required to achieve a fair esti- mate of integral would increase dramatically, i.e., proportional to n d.. { Even
In a nonparametric setting, we discuss identifiability of the conditional and un- conditional survival and hazard functions when the survival times are subject to dependent
[r]
Reading Task 6: Genre Structure and Language Features. • Now let’s look at how language features (e.g. sentence patterns) are connected to the structure
(c) Draw the graph of as a function of and draw the secant lines whose slopes are the average velocities in part (a) and the tangent line whose slope is the instantaneous velocity
When the relative phases of the state of a quantum system are known, the system can be represented as a coherent superposition (as in (1.2)), called a pure state; when the sys-
