太魯閣國家公園關原地區鳥類利用巢箱之繁殖與棲息
蕭明堂
*莊美真 王穎
國立臺灣師範大學生命科學系 (收稿日期:2007.12.18,接受日期:2008.3.21) 摘 要 為瞭解以次級洞穴為巢的鳥類對巢箱在繁殖或棲息上的利用,吾人於太魯閣國家公園之關原 地區設置兩種規格之巢箱(洞口與底面積:舊巢箱 3 cm 和 169 cm2;新巢箱 4.5 cm 和 182.25 cm2 ), 於 2004 年 2 月至 2005 年 8 月間,監測巢箱中鳥類築巢巢材與棲息所留下排遺的出現情形,分別代 表巢位與棲所之利用,以檢測(1)有鳥排遺出現的巢箱占總數之比例(鳥排遺占用率)與氣候的關係; (2)鳥類對巢位與棲所之利用在新、舊巢箱間之差異;(3)巢位與棲所之出現,彼此間有無關連,以 及(4)同一巢箱其利用型態間在各年間的變化。結果顯示,在平均溫度較低時,鳥排遺占用率較高 (p=0.003),另當降水日數較多時,降水量對鳥排遺占用率之影響亦較高(p=0.003)。在巢位選擇上, 根據苔蘚巢材之占用率(p<0.001)、已完成巢之比例(p<0.001)皆為舊巢箱高於新巢箱以及實際繁殖巢 之資料顯示,青背山雀(Parus monticolus)與煤山雀(P. ater)傾向選擇使用洞口與底面積較小之舊巢箱 繁殖;而普通鳾(Sitta europaea)所利用之樹皮巢材則無顯著差異,惟普通鳾於 2005 年中皆用新巢箱 繁殖,似有傾向選擇使用洞口與底面積較大之新巢箱。在棲所方面,鳥排遺占用率在新、舊巢箱間 無顯著差異(p>0.1)。以所有的巢位與棲所占用巢箱的比例來看,並未有明顯分別使用新、舊巢箱之 情形,然其在巢箱的使用上仍有所分隔,在人為收取舊巢材的 2004 與 2005 年繁殖季中,鳥類傾向 選擇不同巢箱分別進行築巢與棲息(2004 年:p=0.003;2005 年:p<0.001);且於繁殖季中實際有繁 殖的巢箱,其後於非繁殖季中出現鳥排遺之機率有低於其他巢箱之趨勢(p=0.08)。另外,於人為清 除舊巢材後,在前一年具有苔蘚、樹皮或鳥排遺之巢箱,隔年仍有較高的機率被鳥類作為相同之利 用,顯示巢箱周圍的微棲地環境有可能影響鳥類在巢位與棲所上的選擇。 關鍵詞:巢箱、巢位、棲所、青背山雀、煤山雀、普通鳾緒 言
許 多 以 次 級 洞 穴 為 巢 的 鳥 種 (secondary cavity-nester),包括山雀科與鳾科鳥類,除利用 腐 敗 樹 洞 、 裂 縫 或 為 其 他 掘 穴 鳥 種 (primary cavity-nester)使用過的巢洞進行築巢與繁殖外, 在氣候條件較嚴苛的環境中,亦會利用洞穴進行 棲息(Kempenaers and Dhondt, 1991; Báldi and Csörgõ, 1994, 1997; Matthysen, 1998)。良好的棲 所可增加鳥類對抗嚴苛的氣候或捕食者,提高鳥 類的存活率(Kendeigh, 1961)。然而,鳥類在棲息 的地方多會留下排遺,MerilÄ and Allander(1995) 在大山雀(Parus major)的研究中發現,在有個體 棲息的巢箱中,有 92%皆有排遺的累積。由於排 遺的氣味可能吸引捕食者的注意(Pitt, 1976),且 累積的排遺亦會污染洞穴,而洞穴的清潔與外寄 生蟲的多寡則會影響鳥類的繁殖表現,例如:外 寄生蟲的存在會延遲大山雀的產卵時間、降低卵 的孵化並增加親鳥棄巢的比例(Oppliger, 1994)。 因此,鳥類對於洞穴在繁殖與棲息的使用上可能 會有所衝突,Perrins(1979)認為鳥類傾向避免在 較佳的洞穴中棲息而保持那些洞穴的乾淨,以利 之後的繁殖使用。 在繁殖與棲息之不同利用目的下,鳥類對於 洞穴的選擇性可能有所差異,大山雀(Perrins, 1979)與卡羅山雀(P. carolinensis)(Pitt, 1976)通常 多選擇較小或剛好僅能容身的洞穴進行棲息,以 減少輻射熱能的散失。然而在繁殖上,使用較大 的洞穴則有利於親鳥投入更多的巢材,使孵卵期 中卵的保溫效果提高(Löhrl, 1973),在炎熱的天氣 中,亦有利雛鳥散熱,避免過熱(hyperthermia)以 減 少 雛 鳥 在 溫 度 調 節 上 的 能 量 消 耗 (Mertens, 1969; van Balen and Cave, 1970),因而雌鳥可能會 選擇在較大的洞穴中繁殖,並產下較大的窩卵數(Karlsson and Nilsson, 1977; van Balen, 1984; Rendell and Robertson, 1989)。此外,洞穴的其他 形質特徵亦會影響鳥類在繁殖上的使用,例如:
洞 口 的 直 徑 與 使 用 鳥 種 的 體 型 有 關(Nilsson,
1984),且會影響鳥巢被捕食者捕食或被其他鳥種 干擾之程度(Wesołowski, 2002; Wesołowski and Rowiński, 2004)。巢箱的新舊則可能與外寄生蟲 的多寡有關,Perrins(1979)發現當每年研究的樣 區中,有部分巢箱損壞並重新設置時,鳥類多選 擇在新設置的巢箱中進行繁殖。 本研究擬探討鳥類對於繁殖巢位與棲息棲 所之選擇。由於天然洞穴之形質差異較大,於 Wang et al.(2003)天然巢位的觀察,大山雀天然巢 之洞口橫徑為5.4±6.2 cm;沼澤山雀(P. palustris) 為3.9±2.0 cm;普通鳾(Sitta europaea)則為 4.7±1.0 cm;然於 Nilsson(1984)研究中,大山雀的洞口橫 徑則介於 2.5-2.8 cm;藍山雀(P. caeruleus)介於 1.8-2.8 cm;沼澤山雀介於 1.5-1.6 cm;普通鳾介 於 2.8-3.0 cm。藉由人工巢箱的設置能固定其他 因子,方便探討單純因子對於鳥類巢位選擇的影 響,故本研究沿用(Hwang, 1996; Lin, 1999)於太 魯閣國家公園所設置之巢箱,並在既有舊巢箱之 下方或旁邊重新釘置新巢箱,以提供兩種規格之 巢箱予鳥類利用,舊巢箱洞口為3 cm,底面積為 169 cm2,而新巢箱洞口為4.5cm,底面積為 182.25 cm2。於繁殖季與非繁殖季中,記錄鳥類築巢之 巢材及棲息時所遺留之排遺於巢箱中累積之情 形,做為鳥類利用巢箱進行繁殖或棲息之判定, 並探討下列問題: 1.在較嚴苛的氣候環境下,鳥類利用巢箱進行棲 息之比例(具鳥排遺之巢箱數)是否跟著增加? 2.鳥類是否傾向在較乾淨且底面積較大的新巢箱 中進行繁殖,而在洞口與底面積較小的舊巢箱中 棲息,因而在舊巢箱中會有較多的鳥排遺出現? 3.繁殖巢位與棲所間的關連: (1)於同一年中,鳥類對巢箱在繁殖或棲息上的利 用,彼此間有無關連?在繁殖季與非繁殖季間有 無差異?相較於人為清除舊巢材之情況,在自然 環境下又為如何? (2)在不同年間,鳥類是否選擇某些特定巢箱進行 繁殖或棲息?前一年被鳥類利用之巢箱,隔年被 鳥類用來進行相同利用的機率是否較高?
材料與方法
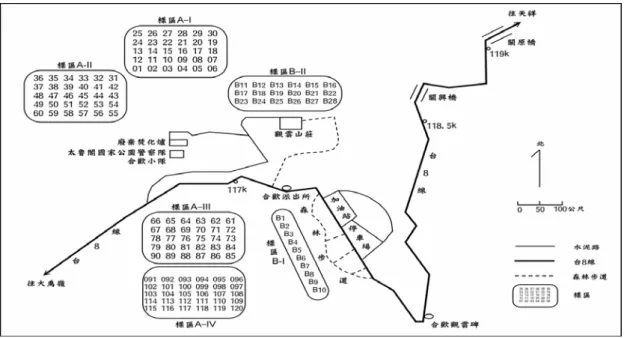
研究地點與巢箱設置 研究地點位於太魯閣國家公園關原地區(圖 一),本研究沿用 Hwang(1996)與 Lin(1999)於中 橫公路 116-117 k 上下的二葉松林中所設置的樣 區(樣區 A),並於 2004 年 1 月初,在既有舊巢箱 之下方或旁邊重新釘置120 個新巢箱,新巢箱之 洞口與底面積皆大於舊巢箱,新巢箱高度與舊巢 箱相近,皆離地高約3 公尺,方位亦同樣朝南。 另外,在原本無舊巢箱之觀雲山莊旁以及關原加 油站對面的步道上,亦增設 20 個新巢箱(樣區 B),以供對照比較鳥類對新、舊巢箱之選擇。於 2005 年 2 月在觀雲山莊旁又再增設 8 個新巢箱。 由於部分巢箱會隨著時間而逐漸腐朽,或遭齧齒 目動物啃咬而無法使用,總計A 樣區於 2004 年 有效之舊巢箱數為 80 個,新巢箱數為 119 個; 於2005 年有效之舊巢箱數為 62 個,新巢箱數為 117 個。B 樣區於 2004 與 2005 年之有效新巢箱 數分別為20 與 27 個。 巢箱監測 吾人分別於2004 年 2 月與 2005 年 1 月將前 年鳥類築巢使用的舊巢材與鳥排遺自巢箱中清 除乾淨。自 2004 年 2 月 25 日起,至 2005 年 8 月9 日止,於繁殖季時(2-8 月),每隔 5-9 天巡視 樣區中巢箱 1 次,在非繁殖季時(9 月至隔年 1 月),每月巡視巢箱 1 次,巡巢時記錄巢箱中有無 巢材、巢材的多寡以及是否有鳥排遺的出現,並 用數位相機拍照記錄。由於山雀科之青背山雀(P. monticolus)與煤山雀(P. ater)皆使用苔蘚植物進 行築巢,在雌鳥孵卵前,無法直接由巢材來判斷 使用鳥種,故所記錄的資料僅分成為山雀科鳥類 使用之苔蘚巢材、普通鳾使用之樹皮巢材以及鳥 排遺等三類。若巢箱中同時出現兩種以上的巢 材,則在兩種巢材的項目中都給予記錄,並記錄 其是否有鳥類產卵繁殖,以及窩卵數大小。另 外,於2001-2003 年間未進行巡巢與舊巢材之收 取,而在 2004 年 2 月收取舊巢材時,亦記錄巢 箱中的舊巢材與鳥排遺,代表在自然環境下之累 積情形。 資料分析與統計 鳥類棲息行為與氣象因子之關連 為同時比較繁殖季與非繁殖季之資料,故在 各月份的資料中,皆選擇1 次與上月巡巢間隔相 近 30 天左右的巡巢記錄(各月平均巡巢間隔 29.6 ±5.04 天),藉由比對照片來判斷在該期間中,巢圖一、太魯閣國家公園關原地區巢箱配置分佈。
Figure 1. Spatial arrangement of nest boxes at Guan-Yuan in the Taroko National Park.
箱有無鳥排遺的增加,並將有鳥排遺增加的巢箱 數 除 以 有 效 巢 箱 數 , 以 鳥 排 遺 占 用 率 (feces occupancy rate)表示。在氣象資料部分,採用中央 氣 象 局 距 關 原 地 區 最 近 之 大 禹 嶺 自 動 測 站 (COT790, 121°18' 29〞E, 24°11' 16〞N)所提供之 逐日降水量與氣溫資料,依據各月巡巢日期,將 巡巢間隔中的逐日氣象資料予以平均,以代表該 段時間之氣候狀況。以逐步迴歸分析(stepwise regression)檢定各月份鳥排遺占用率與氣象因子 (月均溫、降水日數、平均降水量與平均風速)之 關連。由於鳥排遺占用率為百分比資料,故先以 正弦反函數轉換後,再進行分析。 新、舊巢箱占用率與使用程度 將所設置的巢箱分成 3 組,分別為樣區 A 舊 巢箱、樣區 A 新巢箱與樣區 B 新巢箱;將研究期 間分成 3 個時期,分別為 2004 年繁殖季、2004 年非繁殖季與 2005 年繁殖季。計算在不同時期 之各樣區以及新、舊巢箱中,有巢材或鳥排遺出 現之巢箱數,並針對被使用而具巢材之巢箱,依 據巢材量之多寡,區分其使用程度,分為(1)未完 成巢(partial nest):鳥類僅嘗試性築巢,巢材量未 將巢箱底部面積全部鋪滿,(2)已完成巢(complete nest):巢材量已將巢箱底部面積全部鋪滿,具多 層 結 構 ; 另 計 算 實 際 進 行 產 卵 繁 殖 之 巢 數 (breeding nest)占已完成巢之比例。 以卡方分析(chi-square tests)檢定巢材(或鳥排 遺)占用率與使用程度,在不同時期之各樣區以及 新、舊巢箱間有無差異,其中在巢材(或鳥排遺) 占用率項目,是以各樣區中「被使用巢箱個數」 相對於「未被使用巢箱個數」來進行分析;在巢 材之使用程度項目,則以各樣區「未完成巢」與 「已完成巢」之巢箱數進行分析。若卡方檢定結 果達顯著水準,則計算每一格之校正後標準化殘
差(adjusted standardized residual,du)進行事後檢
定,標準化殘差∣du∣≧1.96 表示觀察值與期望 值間具顯著差異。在實際繁殖巢相對於已完成巢 之比例部分,由於樣本數較少,僅以百分比表 示,未進行統計分析。另以 Mann-Whitney U test 檢定於各鳥種中,利用新巢箱與舊巢箱之繁殖巢 其窩卵數之差異。 繁殖巢位與棲所之關連
以費氏精確檢定(Fisher Exact Test)檢測(1) 繁殖季(2004 年、2005 年)、非繁殖季(2004 年)以 及繁殖季與非繁殖季之累計(2001-2003 年、2004 年),巢箱被鳥類繁殖利用與否,與其是否被鳥類 棲息而留下排遺間有無關連?其中,繁殖利用是 以巢箱中是否具有築巢之巢材為判定標準,巢材 則包含苔蘚巢材、樹皮巢材以及極少數棕面鶯 (Abroscopus albogularis)使用之芒草巢材;(2)於 2001-2003 年與 2004 年中,巢箱具苔蘚、樹皮巢 材或鳥排遺與否,與其在次年(2004 年與 2005 年) 是否被鳥類進行相同之利用間有無關連。由於在
不同年與不同繁殖季中,有部分巢箱損毀,因而 有效巢箱數不同,故各檢測以最大巢箱數進行分 析。
結 果
鳥類棲息行為與氣象因子之關連 於 2004 年 9 月中旬至 2005 年 3 月中旬間, 鳥排遺占用率高於其他月份(圖二),特別在 2005 年 2 月中旬至 3 月中旬期間,發生 2 次寒流導致 樣區降雪,該時期鳥排遺占用巢箱之比例亦達到 最高。將各月份氣象因子與鳥排遺占用率進行迴 歸分析(n=18),結果顯示:溫度越低時,鳥排遺 占用巢箱率顯著越高;且在降水日數越多時,降 水量對鳥排遺占用率的影響亦越高。鳥排遺占用 率(arcsine transform)= -0.69(average temperature, t=-3.70, p=0.003) -0.06(precipitation, t=-0.49,p=0.631) +0.10(Number of raining days, t=0.78, p=0.447) +0.06(precipitation*Number of raining days, t=3.64, p=0.003) +14.97 , 決 定 係 數 (R2)=0.767;在考量上述因子下,平均風速對鳥排 遺占用率的影響不顯著(p=0.873)。 新、舊巢箱占用率與使用程度 苔蘚巢材 各樣區巢箱之苔蘚巢材占用率在兩年繁殖 季間無顯著差異(G=4.97, df=5, 0.25<p<0.5)。將兩 年資料合併(表一),苔蘚巢材占用率在不同樣區 之 新 、 舊 巢 箱 間 具 顯 著 差 異 (G=25.72, df=2, p<0.001),樣區 A 舊巢箱之巢材占用率顯著較高 (du=4.32, p<0.001) , 樣 區 A 新 巢 箱 顯 著 較 低 (du=-5.04, p<0.001),樣區 B 新巢箱則無顯著差異 (du=1.49, p=0.14)。 O b s e r v a tio n p e r io d 2 /25 -3 /1 9 3 /20 -4 /1 9 4 /20 -5 /1 8 5 /19 -6 /1 3 6 /14 -7 /1 5 7 /16 -8 /1 9 8 /20 -9 /1 6 9 /17 -1 0 /15 1 0/1 6 -1 1/1 9 1 1/2 0 -1 2/1 3 1 2/1 4 -1 /2 4 1 /25 -2 /1 7 2 /18 -3 /1 9 3 /20 -4 /1 6 4 /17 -5 /1 8 5 /19 -6 /1 2 6 /13 -7 /5 7 /6-8 / 9 0 2 4 6 8 1 0 1 2 1 4 1 6 0 5 1 0 1 5 2 0 2 5 3 0 0 2 4 6 8 1 0 1 2 1 4 1 6 F e ce s oc cup anc y rat e (%) Ave ra g e pr ec ip it at io n (m m ) A v erage t e mp erat ure (℃ ) 2 0 0 4 2 0 0 5 圖二、2004 年 2 月至 2005 年 8 月關原地區各月巡巢期間相對應之平均溫度(實線實心圓)、降水量(虛線空心三 角形)與巢箱之鳥排遺占用率(柱狀)。
Figure 2. Average temperatures (solid circles with solid line), average precipitations (open triangles with dashed line)
and feces occupancy rates of the birds roosting in nest boxes (solid columns) at Guan-Yuan from 25 February 2004 to 9 August 2005.
表一、關原地區鳥類利用巢箱之巢材占用率與各樣區新、舊巢箱之關連(2004 年與 2005 年合計)。
Table 1. Occupancy rates of nest materials of the birds breeding in nest boxes in relation to localities and nest boxes
types at Guan-Yuan (combined data for 2004-2005).
Number of nest boxes occupied (%)a Locality Nest boxes type Number of total nest boxes
Moss Bark flake Empty
Site A Old 142 54 (38.0) 23 (16.2) 68 (47.9)
New 236 37 (15.7) 42 (17.8) 147 (62.3)
Site B New 47 16 (34.0) 25 (53.2) 12 (25.5)
Total 425 107 (25.2) 90 (21.2) 227 (53.4)
表二、關原地區鳥類利用巢箱之使用程度與各樣區新、舊巢箱之關連(2004 年與 2005 年合計)。
Table 2. The level of utilization of the birds breeding in nest boxes in relation to localities and nest boxes types at
Guan-Yuan (combined data for 2004-2005).
Number of complete nest (%)a Number of breeding nest (%)b Locality Nest boxes type
Moss Bark flake Moss Bark flake
Site A Old 38 (70.4) 8 (34.8) 29 (76.3) 4 (50.0)
New 9 (24.3) 8 (19.1) 3 (33.3) 6 (75.0)
Site B New 11 (68.8) 4 (16.0) 7 (63.6) 3 (75.0)
Total 58 (54.2) 20 (22.2) 39 (67.2) 13 (65.0)
a. The percentage of complete nest compared to total occupied nest b. The percentage of breeding nest compared to total complete nest
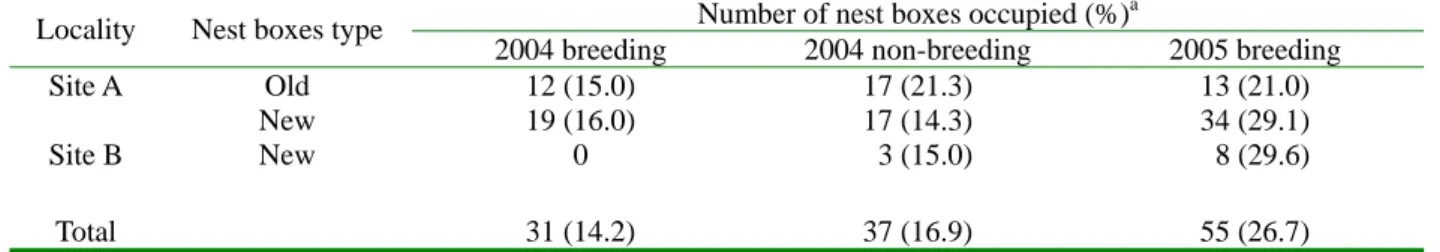
表三、2004 及 2005 年關原地區鳥類利用巢箱之鳥排遺占用率與各樣區新、舊巢箱之關連。
Table 3. Feces occupancy rates of the birds roosting in nest boxes in relation to localities and nest boxes types at
Guan-Yuan, 2004 and 2005.
Number of nest boxes occupied (%)a Locality Nest boxes type
2004 breeding 2004 non-breeding 2005 breeding
Site A Old 12 (15.0) 17 (21.3) 13 (21.0)
New 19 (16.0) 17 (14.3) 34 (29.1)
Site B New 0 3 (15.0) 8 (29.6)
Total 31 (14.2) 37 (16.9) 55 (26.7)
a. The percentage of occupied nest compared to total available nest boxes
在巢箱被使用程度上,不同年間無顯著差異 (G=6.56, df=5, 0.25<p<0.5)。將兩年資料合併(表 二),於被使用而有巢材之巢箱,其使用程度在不 同樣區之新、舊巢箱間具顯著差異(G=21.02, df=2, p<0.001),在樣區 A 被使用之舊巢箱中,有顯著 較高的比例為「已完成巢」(du=3.39, p<0.001), 樣區 A 新巢箱顯著有較高的比例為「未完成巢」 (du=-4.51, p<0.001),樣區 B 新巢箱則無顯著差異 (du=1.27, p=0.205)。在實際繁殖巢的部分(表二), 在樣區 A 舊巢箱與樣區 B 新巢箱中,已完成巢其 後實際有鳥進行繁殖之比例較高,但在樣區 A 新 巢箱中,該比例則相對較低。 比較在新、舊巢箱繁殖的巢之窩卵數大小, 於青背山雀中,兩者間無顯著差異(old nest boxes: 5.67±0.69, n=18; new nest boxes: 5.57±0.53, n=7, p>0.2);於煤山雀中,僅有 2 巢使用新巢箱繁殖, 窩卵數分別為 4 和 6 枚,其和在舊巢箱繁殖巢之 窩卵數(5.38±0.52, n=8),兩者間亦無顯著差異 (p>0.2)。 樹皮巢材 於兩年繁殖季間,各樣區巢箱之巢材占用率 無顯著差異(G=3.31, df=5, 0.5<p<0.75)。將兩年資 料合併(表一),樹皮巢材占用率在不同樣區之 新 、 舊 巢 箱 間 具 顯 著 差 異 (G=27.05, df=2, p<0.001),樣區 B 新巢箱之巢材占用率顯著較高 (du=5.50, p<0.001),樣區 A 的舊巢箱與新巢箱之
巢材占用率則無顯著差異(old nest boxes: du=-1.8,
p=0.718; new nest boxes: du=-1.91, p=0.057)。於使
用程度方面(表二),「樣區」、「年」與「使用程度」 3 個因子 間則 無顯著 交互 作用 (G=8.31 , df=5, 0.25<p<0.5),合併不同樣區與不同年之資料,在 被使用的巢箱中,「已完成巢」占 22.2 %。在實 際繁殖巢的部分(表二),樣區 A 新巢箱與樣區 B 新巢箱中,已完成巢其後有鳥實際進行繁殖之比 例相對較高,且於 2005 年繁殖季,所有實際繁 殖巢亦皆是在新巢箱中。在窩卵數方面,普通鳾 於新巢箱繁殖的巢之窩卵數(4.67±0.5, n=9),與其 在舊巢箱繁殖的巢之窩卵數(4.5±0.58, n=4),兩者 間無顯著差異(p>0.2)。 鳥排遺 於不同樣區之新、舊巢箱間,鳥排遺占用率 無顯著差異(G=13.59, df=10, 0.1<p<0.25, 表三)。 將不同樣區的資料合併,於不同時期間,鳥排遺 占用率具顯著差異(G=11.5, df=5, 0.005<p<0.01), 2004 年繁殖季巢箱之鳥排遺占用率顯著較低 (du=-2.29, p=0.022),2004 年非繁殖季無顯著差異
(du=-1.02, p=0.307),2005 年繁殖季則顯著較高 (du=3.37, p<0.001),且於 2005 年 2/18-3/19 期間, 出現鳥排遺之巢箱數占該年繁殖季出現鳥排遺 之巢箱數之 54.6%。若刪除該月份資料,則不同 時 期 間 , 鳥 排 遺 占 用 巢 箱 率 不 具 顯 著 差 異 (G=15.01, df=5, 0.1<p<0.25)。 在 2005 年 2/18-3/19 期間,出現鳥排遺之巢 箱數相對於該樣區有效巢箱數之比例,樣區 A 舊 巢箱:樣區 A 新巢箱:樣區 B 新巢箱為 9.7:18.8: 7.4,但未達到顯著(G=4.21, df=2, 0.1<p<0.25)。 繁殖巢位與棲所之關連 檢視鳥類繁殖的巢位(具巢材之巢箱)與棲息 的棲所(具鳥排遺之巢箱)之關連(表四),於 2004 與 2005 年繁殖季,具鳥排遺的巢箱中,其出現 巢材之機率皆顯著低於不具鳥排遺的巢箱(2004: p=0.003; 2005: p<0.001)。於 2004 年非繁殖季, 巢箱中出現鳥排遺之有無,則與該巢箱是否具繁 殖季之巢材無關(p=0.590);然而於 2004 年繁殖 季中實際有繁殖的巢箱,在其後非繁殖季出現鳥 排遺之機率(2/34)有較其他未被實際繁殖之巢箱 出現鳥排遺之機率(35/185)為低的趨勢,但在統 計上未達顯著(p=0.08)。將繁殖季與非繁殖季累 計後分析,於 2001-2003 年,巢箱內巢材之有無 與鳥排遺之有無,二者間無顯著關連(p=0.604); 然於 2004 全年中,具鳥排遺之巢箱其出現巢材 之機率則顯著較低(p=0.011)。 同一巢箱連續兩年之利用類型間有所關連 (表五),無論是在山雀科鳥類或普通鳾中,前一 年有被鳥類使用嘗試築巢或實際繁殖之巢箱,其 隔年再出現同種鳥類築巢之機率皆高於前一年 非相同使用之巢箱;而前一年具鳥排遺之巢箱, 次年再出現鳥排遺之機率亦高於前一年不具鳥 排遺之巢箱,然皆僅有 2005 年資料到達統計上 之顯著水準。
討 論
鳥類棲息行為與氣象因子之關連 鳥類會因應氣候環境而調整其行為,例如: 在溫度較低、風速較高的環境,美洲鳳頭山雀(P. bicolor) 和卡羅山雀等數種小型鳥類會增加它們 覓 食 時 停 留 的 時 間 與 次 數 , 減 少 飛 行 的 距 離 (Grubb, 1978);黑頂山雀在極度寒冷的天氣中, 表四、關原地區各時期巢箱有無鳥排遺與其被鳥類繁殖利用之關連。Table 4. Nest boxes quality (with or without feces) and use by cavity nesting birds during the different periods at
Guan-Yuan.
Number of nest boxes which has nest material
Number of nest boxes which has no nest material Periods
With feces Without feces With feces Without feces
p value* 2004 breeding 7 98 24 90 0.003 2005 breeding 9 77 40 80 <0.001 2004 non-breeding 16 89 21 93 0.590 2001-2003 whole year 41 22 12 9 0.604 2004 whole year 22 83 42 72 0.011
* Fisher’s Exact Test
表五、2004 及 2005 年關原地區巢箱被鳥類利用和其前一年利用類型之關連。
Table 5. Nest boxes utilization in relation to the types of use in last year at Guan-Yuan, 2004 and 2005.
Number of nest boxes which has being use
Number of nest boxes which has not being use
Use type Year
The same use as last year
The use different from last year
The same use as last year
The use different from last year
p value* Moss 2004 18 10 23 26 0.162 2005 21 31 25 121 0.001 Bark flake 2004 5 7 11 54 0.113 2005 14 26 30 128 0.035 Feces 2004 19 8 25 25 0.097 2005 26 22 31 119 <0.001
會顯著地減少它們的覓食半徑(Kessel, 1976);在 氣候環境較差時,許多小型鳥類(包括大部分的山 雀科鳥類)會選擇棲息於矮灌叢、洞穴或巢箱來減
少 能 量 的 消 耗(Perrins, 1979; Kempenaers and
Dhondt, 1991)。於本研究中,鳥類利用巢箱棲息 所留下鳥排遺之多寡隨著各月份氣候狀況而有 所變化,在較惡劣的氣候環境下(溫度越低、降水 日數與降水量越高),鳥排遺占用率顯著較高,特 別在2005 年 2 月中旬至 3 月中旬寒流與降雪期 間,鳥排遺占用率更是顯著高於其他月份,表示 鳥類會反應氣候環境狀況而增加棲息行為。另 外,於 2005 年寒流與降雪期間,亦發現部分巢 箱有大量鳥排遺累積之情形。雖然在大多數山雀 科鳥類中,冬季的棲息多半都是單獨棲息(Odum, 1942; Perrins, 1979),但有少數個案發現(Loery and Nichols, 1985),在極度寒冷的環境中,黑頂 山雀會群聚在一起棲息(communal roosting),且這 樣的行為亦發現於相近山雀科的其他鳥種中,包 括:銀喉長尾山雀(Aegithalos caudatus)(Mcgowan et al., 2006) 與 短 嘴 長 尾 山 雀 (Psaltriparus minimus)(Smith, 1972) , 以 及 鳾 科 之 小 鳾 (S. pygmaea)(Hay, 1983),群聚棲息有助於維持體 溫,減少熱量散失(Brenner, 1965),特別是在小型 雀型目鳥類,因此推測大量鳥排遺出現於同一巢 箱中,除可能是鳥類為躲避寒冷天氣而長時間棲 息外,亦無法排除為多數個體群聚棲息所造成。 新、舊巢箱之選擇 在繁殖巢位方面,本研究之山雀科鳥類與普 通鳾對於巢位的選擇並不相同。在同時具有新巢 箱與舊巢箱的環境中,山雀科鳥類傾向在舊巢箱 中繁殖,故舊巢箱之巢材占用率、已完成巢之比 例以及實際繁殖巢之比例都高於新巢箱,然而在 僅有新巢箱的環境中,則對新巢箱仍有相當高的 使用率,表示在有所選擇的情形下,山雀科鳥類 傾向使用舊巢箱進行繁殖。然普通鳾則在新、舊 巢箱之巢材占用率上無顯著差異,而由 2005 年 所有實際繁殖巢皆位於新巢箱來看,普通鳾似有 傾向選擇使用新巢箱進行繁殖之趨勢。兩種鳥類 於新、舊巢箱的巢位選擇之差異,與研究預期「鳥 類傾向在新巢箱中繁殖」未完全符合,推測可能 與新、舊巢箱規格,以及鳥類繁殖生態習性之差 異有關。雖然本研究所提供之兩種洞口直徑規 格,皆為天然環境中山雀科鳥類與普通鳾可能利 用之大小(Edington and Edington, 1972; van Balen, 1982; Nilsson, 1984; Wang et al., 2003),但新巢箱
較大的洞口可能增加鳥類被捕食者捕食之風險 (Wesołowski, 2002; Wesołowski and Rowiński, 2004),以及共域鳥種對巢位之競爭與干擾(van Balen, 1982),或許這些因子造成繁殖失敗風險的 增加,大於使用舊巢箱所導致之外寄生蟲感染或 雛鳥過熱等問題,且於本研究中,舊巢箱之底面 積並未限制繁殖之窩卵數大小,使用新、舊巢箱 繁殖的巢其窩卵數並無顯著差異,因而山雀科鳥 類偏好選擇舊巢箱進行繁殖。相較於Perrins(1979) 觀察英國山雀科鳥類傾向選擇新設置巢箱之結 果,可能係因其所設置之新、舊巢箱皆為同一規 格。推測在山雀科鳥類之巢位選擇上,洞口大小 為較優先考量的因子。另一方面,由於普通鳾能 藉由糊泥土在洞口來縮減洞口直徑,較不受洞口 大小的限制,且新巢箱的底面積較大,可堆疊較 多的樹皮巢材,有助於洞穴的乾燥,而受普通鳾
所 偏 好 (van Balen, 1982; Wesołowski and
Rowiński, 2004),故本研究之普通鳾相對地較傾 向選擇洞口較大的新巢箱中繁殖。 在棲所方面,鳥排遺占用率在不同樣區之 新、舊巢箱間無顯著差異,亦與研究預期「洞口 及底面積較小之舊巢箱鳥排遺出現比例較高」不 符,推測可能的原因有下列幾點:由於山雀科與 鳾科鳥類全年皆具領域行為,其防衛的洞穴可能 涵蓋超過1 個以上,使得部分洞穴無法被其他個
體所利用(Dhondt and Eyckerman, 1980; Enoksson, 1990; Kempenaers and Dhondt, 1991; Báldi and Csörgõ, 1997),因而領域擁有者在棲所的選擇上 能有較多的彈性,較不受限於巢箱的新舊;另一 方面,於本研究主要利用巢箱鳥種之山雀科與普 通鳾中,兩種鳥類繁殖巢位之選擇性不同,山雀 科鳥類傾向於舊巢箱中築巢與繁殖,普通鳾則傾 向於新巢箱中繁殖,或許兩種鳥類對於棲所之選 擇亦有所差異;另外,亦可能有其他非利用巢箱 繁殖之鳥種,於巢箱中棲息並留下排遺,而混淆 了新、舊巢箱中鳥排遺占用率之結果,對於鳥類 棲息行為之觀察與利用鳥種辨識為後續研究之 方向。 繁殖巢位與棲所之關連 雖然以所有的巢位與棲所占用巢箱的比例 來看,並未有明顯分別使用新、舊巢箱之情形, 然其在巢箱的使用上仍有所區隔。在人為收取舊 巢材的2004 與 2005 年繁殖季,具鳥排遺之巢箱 其 出 現 巢 材 之 機 率 較 不 具 鳥 排 遺 之 巢 箱 來 得 低;然於2004 年非繁殖季,兩者則無顯著關連。
表示鳥類在繁殖季時,巢位與棲所之選擇並非互 相獨立,支持 Perrins(1979)鳥類避免在較佳的洞 穴中棲息而保持那些洞穴的乾淨,以利其後繁殖 使用之預期。另外,在繁殖季中實際有繁殖的 巢,於其後非繁殖季中出現鳥排遺之機率有較低 之趨勢,推測此可能與寄生蟲有關。在大山雀以 及其他洞穴巢鳥種中,雞蚤(Ceratophyllus sp.)為 主要的外寄生蟲,雞蚤通常在每巢中產生 2 個世 代(Harper et al., 1992),第二個世代在幼鳥離巢後 會發生,數以千計的雞蚤會以囊鞘的形式黏附在 巢材中,等待有機械性刺激時才由囊鞘中出來 (Humphries, 1968),因此鳥類會避免在已繁殖過 的 巢 箱 中 棲 息 , 減 少 被 外 寄 生 蟲 感 染 之 風 險 (Christe et al., 1994; MerilÄ and Allander, 1995)。 舊巢材的累積除了影響鳥類在該年非繁殖季中 對棲所的選擇,亦會影響隔年鳥類繁殖巢位之選 擇,Lin(1996)發現本區的山雀科鳥類會避免在具 舊巢材的巢箱中繁殖。因而推測在自然環境下, 無人為移除舊巢材與鳥排遺,巢材逐年的累積將 使得鳥類築巢巢位與棲所之選擇跟著減少,故以 較長的時間尺度(2001-2003)進行檢視時,巢材與 鳥排遺間相關性減低而相互獨立。 此外,本研究以人為將舊巢材移除後,不同 年間鳥類傾向在前一年同一巢箱進行相同之利 用,雖於 2004 年的資料中,統計上未達顯著, 但仍有相同之趨勢,推測除巢箱的規格外,微棲 地 環 境 亦 可 能 影 響 巢 位 的 選 擇 (Willner et al., 1982; Munro and Rounds, 1985)。吾人於研究期間 觀察發現,在巢箱洞口前若具有一小段枝幹或闊 葉樹枝條,將有利於親鳥進巢前的停棲;選擇設 置巢箱的台灣二葉松若生長在坡度較陡、急遽升 降的坡面上,該巢箱亦較受鳥類青睞而用來築巢 繁殖;另外,針對使用巢箱繁殖之青背山雀個體 進行繫放,亦發現於 2004 年與 2005 年繁殖季 中,使用同一巢箱進行繁殖之個體皆非同一對個 體(n=3),皆為支持微棲地環境對巢位選擇影響的 間接證據。 本研究設置兩種規格的巢箱,發現不同鳥種 對於新、舊巢箱在繁殖的選擇上有所差異,而鳥 類 對 於 巢 箱 在 繁 殖 與 棲 息 的 利 用 上 亦 有 所 區 隔,且在不同年間傾向對同一巢箱有相同利用型 態。此一研究有助於提供往後規劃設置鳥巢箱進 行監測時,巢箱規格影響所吸引鳥種之參考。未 來可進一步調查巢箱周圍的微棲地因子,以釐清 鳥類在繁殖與棲息之不同利用目的下,對於洞穴 選擇之主要考量因子為何?另外,研究結果顯示 鳥類利用巢箱進行棲息之比例受到嚴苛氣候的 影響,故持續進行巢箱監測,以瞭解氣候因子對 於鳥類利用巢箱之頻率及其存活之影響,亦可為 往後研究之方向。
誌 謝
本研究感謝太魯閣國家公園管理處在經費 上的補助,師大野生動物實驗室在野外工作上的 協助與行政上的支持,謹此一併誌謝。參考文獻
Báldi A and Csörgõ T. 1994. Roosting site fidelity of Great Tits (Parus major) during winter. Acta Zool. Hung. 40: 359-367.
Báldi A. and Csörgõ T. 1997. Spatial arrangement of roosting Great Tits (Parus major) in a Hungarian forest. Acta Zool. Hung. 43: 295-301.
Brenner FJ. 1965. Metabolism and survival time of grouped Starlings at various temperatures. Wilson Bull. 77: 388-395.
Christe P, Oppliger A and Richner H. 1994. Ectoparasite affects choice and use of roost sites in the great tit, Parus major. Anim. Behav. 47: 895-898.
Dhondt AA and Eyckerman R. 1980. Competition between the Great Tit and the Blue Tit outside the breeding season in field experiments. Ecology 61: 1291-1296.
Edington JM and Edington MA. 1972. Spatial patterns and habitat partition in the breeding birds of an upland wood. J. Anim. Ecol. 41: 331-357.
Enoksson B. 1990.Autumn territories and population regulation in the nuthatch Sitta europaea: an experimental study. J. Anim. Ecol. 59: 1047-1062.
Grubb TC, Jr. 1978. Weather-dependent foraging rates of wintering woodland birds. Auk 95: 370-376.
Harper GH, Marchant A and Boddington DG. 1992. The ecology of the hen flea Ceratophyllus gallinae and the moorhen flea Dasypsyllus gallinulae in nestboxes. J. Anim. Ecol. 61: 317-327.
Hay DB. 1983. Physiological and behavioral ecology of communally roosting Pygmy Nuthatches (Sitta pygmaea). Ph.D. thesis.
Northern Arizona University.
Humphries DA. 1968. The host-finding behaviour of the hen flea, Ceratophyllus gallinae (Schrank)(Siphonaptera). Parasitology 58: 403-414.
Hwang, CL 1996. Breeding biology of birds using nest boxes in Taroko National Park. Master’s thesis. National Taiwan Normal University. Taipei, Taiwan. (in Chinese).
Karlsson J and Nilsson SG. 1977. The influence of nest-box area on clutch size in some hole-nesting passerines. Ibis 119:207-211. Kempenaers B. and Dhondt AA. 1991. Competition
between Blue and Great Tit for roosting sites in winter: an aviary experiment. Ornis Scand. 22:73-75.
Kendeigh S. 1961. Energy of birds conserved by roosting in cavities. Wilson Bull. 73: 140-147.
Kessel B. 1976. Winter activity patterns of Black-capped Chickadees in interior Alaska. Wilson Bull. 88: 36-61.
Lin KC. 1999. Nest box site selection and reproductive performance of three sympatric birds. Master’s thesis. National Taiwan Normal University. Taiwan. (in Chinese). Loery G. and Nichols JD. 1985. Dynamics of a
Black-capped Chickadee population, 1958-1983. Ecology 66: 1195-1203.
Löhrl H. 1973. Einfluss der Brutaumfläche auf die Gelegegrösse der Kohlmeise (Parus major). J. Orn. 114: 339-347. (in Germany with English summary)
Matthysen E. 1998. The nuthatches. T and AD Poyser, London.
McGowan A, Sharp SP, Simeoni M and Hatchwell BJ. 2006. Competing for position in the communal roosts of long-tailed tits. Anim. Behav. 72:1035-1043.
MerilÄ J and Allander K. 1995. Do great tits (Parus major) prefer ectoparasite-free roost sites? an experiment. Ethology 99: 53-60.
Mertens JAL. 1969. The influence of brood size on the energy metabolism and water loss of nestling Great Tits Parus major major. Ibis 111: 11-16.
Munro HL and Rounds RC. 1985. Selection of artificial nest sites by five sympatric passerines. J. Wildl. Manage. 49: 264-276. Nilsson SG. 1984. The evolution of nest-site
selection among hole-nesting birds: the
importance of nest predation and competition. Ornis Scand. 15: 167-175.
Odum EP. 1942. Annual cycle of the Black-capped Chickadee-3. Auk 59: 499-531.
Oppliger A, Richner H and Christe P. 1994. Effect of an ectoparasite on lay date, nest-site choice, desertion, and hatching success in the great tit (Parus major). Behav. Ecol. 5: 130-134. Perrins CM. 1979. British Tits. William Collins
Sons & Co, London.
Pitts TD. 1976. Fall and winter roosting habits of Carolina Chickadees. Wilson Bull. 88: 603-610.
Rendell WB and Robertson RJ. 1989. Nest-site characteristics, reproductive success and cavity availability for Tree Swallows breeding in natural cavities. Condor 91: 875-885.
Smith SM. 1972. Roosting aggregations of Bushtits in response to cold temperatures. Condor 74: 478-479.
van Balen JH. 1984. The relationship between nest-box size, occupation and breeding parameters of the Great Tit Parus major and some other hole-nesting species. Ardea 72: 163-175.
van Balen JH and Cavé AJ. 1970. Survival and weight loss of nestling Great Tits, Parus major, in relation to brood-size and air temperature. Neth. J. Zool. 20: 464-474. van Balen JH, Booy CJH, van Franeker JA and
Osieck ER. 1982. Studies on hole-nesting birds in natural nest sites. I. Availability and occupation of natural nest sites. Ardea 70: 1-24.
Wang HT, Gao W, Wan DM, Liu D and Deng WH. 2003. Nest-site characteristics and reproductive success of five species of birds breeding in natural cavity. Acta Ecologica Sinica 23(7): 1377-1385.
Wesołowski T. 2002. Anti-predator adaptations in nesting Marsh Tits Parus palustris: the role of nest-site security. Ibis 144: 593-601. Wesołowski T and Rowiński P. 2004. Breeding
behaviour of Nuthatch Sitta europaea in relation to natural hole attributes in a primeval forest. Bird Study 51: 143-155. Willner GR, Gates JE and Devlin WJ. 1983. Nest
box use by cavity-nesting birds. Am. Midl. Nat. 109: 194-201.
Breeding and Roosting of Birds in Nest Boxes at Guan-Yuan in the
Taroko National Park
Ming-Tang Shiao
*, Mei-Chen Chuang, Ying Wang
Department of Life Science, National Taiwan Normal University Taipei, Taiwan
(Received: 18 December 2007, accepted: 21 March 2008)
ABSTRACT
From Feb 2004 to Aug 2005, we surveyed nest materials and feces, representing breeding sites and roosting sites of the birds respectively, between 2 different types of nest boxes (entrance and bottom: 3 cm and 169 cm2 of old box; 4.5 cm and 182.25 cm2 of new box) at Guan-Yuan in the Taroko National Park, and studied (1) the effect of weather on the presence of feces in nest boxes, (2) the difference between the new and old nest boxes used by the birds on breeding and roosting, (3) the association between breeding and roosting sites and (4) temporal changes of use type in the same nest boxes. The results showed that feces had higher appearance in nest boxes when average temperature was lower (p=0.003). So did it when the precipitation was higher in a longer period of rainy days (p=0.003). In terms of breeding site selection, based upon the moss occupancy rates (p<0.001), complete nests ratio (p<0.001) which are all higher in old boxes than in new ones and the data of breeding nests, green backed tit (Parus monticolus) and coal tit (P. ater) tended to choose old nest boxes to breed. As for Eurasian nuthatch (Sitta europaea), there were no significant differences between old and new boxes of bark flake occupancy rates and complete nest ratio. However, that all breeding nests were found only in new nest boxes in 2005 seemed to indicate that nuthatch tended to choose new nest boxes. In terms of roosting site selection, the percentage of each type of nest boxes with feces showed no significant difference. The utilization of nest boxes was either for breeding or roosting, but rarely for both purposes. When we cleaned the old nest materials before breeding season, the birds tended to use different nest boxes for breeding and roosting respectively in breeding season (2004: p=0.003; 2005: p<0.001). And in non-breeding season, feces had lower appearance in nest boxes that had been used for actual breeding in previous breeding season than in other nest boxes (p=0.08). In addition, the nest boxes with nest materials or feces in previous year have more probability to be used again in the same way in following year. This indicated that the microhabitats around nest boxes could affect the selection of breeding and roosting sites.
Key words: nest boxes, breeding site, roosting site, green-backed tit (Parus monticolus), coal tit (Parus ater), Eurasian nuthatch (Sitta europaea)