牛磺酸對四氯化碳誘發大鼠肝臟纖維化的改善效果; Taurine reduces hepatic fibrosis induced by carbon tetrachloride in rats
68
0
0
全文
(2) 圖 目 錄 圖 1. Effect of taurine on liver weight (A) and spleen weight (B) in rats with chronic liver injury induced by CCl4 .. ………………………59. 圖 2. Effect of taurine on GOT (A) and GPT (B) in rats with chronic liver injury induced by CCl4. ………………………………………………60 圖 3. Effect of taurine on Liver protein (A) and Hydroxyproline (B) in rats with chronic liver injury induced by CCl4.……………………………61 圖 4. Effect of taurine on LPO (A) and GSH (B) in rats with chronic liver injury induced byCCl4. ………………………………………………62 圖 5. RT-PCR products analysis of taurine effect on liver Tgf-beta1 (A) and and HGF (B) RNA content in rats with chronic liver injury induced by by CCl4. ……………………………………………………………63. 圖 5. RT-PCR products analysis of taurine effect on liver TIMP-1(C) and Pro-collagen-1(D) ……………………………………………………64 圖 5. RT-PCR products analysis of taurine effect on liver MAT1A (E) and MAT2A (F) RNA content in rats with chronic liver injury induced by CCl4 …………………………………………………………………65 圖 6. Western blot products analysis of taurine effect on liver TGF-beta1 protein contents in rats with chronic liver injury induced byCCl4……66 圖 7. The photomicrographs of liver section taken from rats and stained with hematoxylin-enosin.. ………………………………………………67. 2.
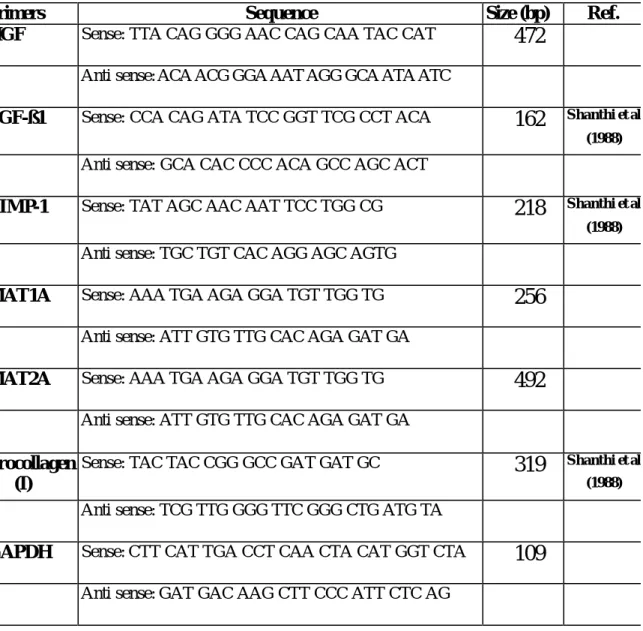
(3) 表 目 錄 表 1. PCR primer sequences and expected size of amplified products …56 表 2. Effect of taurine on CCl4 chronic treated rats liver SOD,Catalase, GSH-Px activities. ……………………………………………………57 表 3. List of liver genes expression in normal/CCl4 ratio ………………58. 3.
(4) 牛磺酸對四氯化碳誘發大鼠肝臟纖維化的改善效果. 研究生 詹明哲. 中國醫藥學院 醫學研究所. 摘要. 本研究目的在於探討牛磺酸 (taurine)對四氯化碳 (CCl4)誘發大鼠肝 臟纖維化的改善作 用。使 用 Wistar 雄性大鼠, 每週兩次經口投予 CCl4,為期八週誘發肝臟纖維化。實驗期間,每日經口投予牛磺酸一次, 使用劑量為 400、800 mg/kg,實驗對照組投予逆滲透水。結果顯示,四 氯化碳誘發大鼠慢性肝炎,使血清 GOT、GPT 活性明顯上升;牛磺酸 的處理第八週能顯著降低 GOT、GPT 活性。四氯化碳誘發大鼠肝纖維 化,脾臟重量明顯增大,肝臟蛋白質含量下降,肝臟脂質過氧化及膠原 蛋白含量明顯增加。800 mg/kg 牛磺酸的處理,脾臟重量有減輕的傾向, 增加蛋白質含量,降低脂質過氧化程度及減少膠原蛋白沉積。肝臟組織 抽取全量 RNA 進行與肝纖維化相關的 RT-PCR 反應,結果顯示,四氯 化碳組明顯增加 transforming growth factor-beta 1 (TGF-beta1)、tissue inhibitor of metalloproteinase-1 (TIMP-1) methionine adenosyltransferase 2A (mat2a)的表現,牛磺酸的處理明顯能抑制這些基因表現。四氯化碳 組減少 hepatocyte growth factor (HGF)、methionine adenosyltransferase 1A (mat1a)的表現,牛磺酸的處理能增加這些基因表現。整體而言,牛 磺酸具有的保肝作用是透過減緩肝纖維化的產生。. 4.
(5) Taurine reduces hepatic fibrosis induced by carbon tetrachloride in rats Ming-Che Chan Institute of Medical Science,China Medical College. The aim of this study was to investigate the improvement of taurine on the fibrosis of rat liver induced by carbon tetrachloride. Liver fibrosis was induced in male Wistar rats by chronic administration of CCl 4 for twice a week for 8 weeks. There are four groups of rats in this study, including normal group and group treated with carbon tetrachloride, and two experimental groups. The rats of two experimental groups were induced liver fibrosis by carbon tetrachloride and received taurine at 400 and 800 mg/kg/day through the whole experimental period. The normal group was treated reverse osmosis water. According to the resultds, rats treated with taurine had obviously reduced levels of the serum activities of GOT and GPT, the degree of LPO, the content of hydroxyproline and the swelling of spleens, besides the increased effects on the level of the protein content of the liver, when compared to the group treated with carbon tetrachloride. The livers were analyzed by reverse transcription-polymerase chain reaction for the transcription of genes involved in liver fibrosis. Taurine had obviously reduced the gene expression in transforming 5.
(6) growth factor beta1 (TGF-beta1), tissue inhibitor of metalloproteinase (TIMP-1), methionine adenosyltransferase 2a (mat2a) and obviously increased level of hepatocyte growth factor (HGF) and methionine adenosyltransferase 1a (mat1a) when compared to the group treated with carbon tetrachloride. In conclusion, the results suggest that taurine has hepatoprotective effect and attenuates the degree of liver fibrosis.. 6.
(7) 前 言. 慢性肝病及肝硬化為 2002 年台灣地區十大死亡原因第六位,肝癌 亦為十大癌症之首。目前台灣地區慢性肝炎患者約有肆佰萬人,顯示台 灣是肝細胞癌,肝硬化及慢性肝炎盛行地區。(參考行政院衛生署衛生 統計資訊網 http://www.doh.gov.tw/statistic/index.htm) 所有慢性肝炎包含病毒性肝炎及酒精性肝炎皆有轉成肝硬化的可 能,目前肝硬化被認為是不可逆的,現代的藥物對於肝硬化尚沒有很好 的治療效果,僅能針對肝硬化產生的併發症處理,如用 β-阻斷劑降門脈 高壓,用利尿劑改善腹水(Gill et al., 1997),因此使用保肝的藥物或食品 來預防慢性肝炎轉變成肝硬化受到重視。 肝纖維化的發生機轉非常複雜,目前所瞭解的是多種造成肝損傷 的因子引起肝細胞損傷、壞死、凋亡及肝組織炎症反應,激活 Kupffer 細胞分泌多種細胞因子;以及肝細胞、血小板及竇內皮細胞等分泌的 細胞因子、脂質過氧化等化學媒介物質共同作用於肝星狀細胞,使之 激活轉變成肌纖維母細胞,經由旁分泌及自分泌的機轉,使肌纖維母 細胞增殖,合成大量的膠原及蛋白多糖等細胞外基質成分(Nebubauer et al., 2001)。 因此具保肝作用或減緩肝纖維化作用的藥物其可能的作用機轉如 下:(1) 保護肝細胞不受到傷害,此又可分為直接的增強肝細胞本身 的保護作用,及間接的消除傷害物質,如清除自由基;(2) 抑制肝臟. 7.
(8) 發炎;(3) 抑制星狀細胞活化;(4) 抑制細胞外基質成分的合成及分 泌;(5) 促使細胞外基質成分降解。上述 5 種作用機轉中,目前市場 上有的以保護肝細胞,減輕肝細胞傷害的藥物為主,其中尤以牛磺酸 (taurine, Tau)使用最多,但其作用的分子機轉尚不十分明確。 已有多數文獻指出牛磺酸可能經由鈣離子的調節(John et al., 1995) 、抗氧化作用(You and Chang, 1998)、polyamine 的合成(Wu, 1999)及經 由 PI3-kinase 的機轉(Marucci, 2002)而具有保肝的效果。最近的研究顯 示牛磺酸能改善肝纖維化,對四氯化碳所誘發的 procollagen I、III 及 tissue inhibition of metalloproteinase-1 (TIMP-1)的基因表現有降低作用( 陳等,1999),唯其作用機轉不明之處仍多。如在肝臟產生纖維化的過程 中佔關鍵性角色的 Transforming growth factor beta1 (TGF-beta1),牛磺 酸對其作用不明。最近的研究指出,肝臟中合成 S-adenosylmethionine (SAM)的酵素 methionine adenosyltransferase (MAT),其活性是肝細胞損 傷的重要指標之一(Shelly et al., 2001),牛磺酸的保肝作用,對 MAT 基 因的表現也未明。 基於上述,本研究主要目的在分析牛磺酸的保肝及抗肝纖維化的作 用機轉中,TGF-beta1 與 MAT 表現的關連性,並利用分子生物學的技 術,包括反轉錄? 鏈合反應(reverse transcriptase polymerase chain reaction, RT-PCR)、 西方墨點法(Western blot)及表現型微陣列分析 (expressed microarray analysis)來闡明其作用之分子機轉。. 8.
(9) 第一章 文獻探討 第一節 牛磺酸. 一、 化 學 及 來 源 牛磺酸是一種含硫的 β 氨基酸,英文名為 taurine, 分子式是 C2H7NO3S (如下圖),它是由牛黃中分離出來的,因此得名牛磺酸,又名 牛膽鹼。其外觀為白色柱狀結晶,無臭、味弱酸。工業合成是以乙醇胺、 硫酸,及硫酸鈉為主要原料,經酯化磺化反應,再用鹽酸和醇萃取分離 製得。 牛磺酸結構式圖:. 牛磺酸以游離的方式廣泛存在動物各種組織細胞內液中,其中以海 螺、烏賊、章魚、文蛤含量較高。之前,人們僅知它的主要功能是與膽 酸結合形成牛磺膽酸。過去幾年來,對牛磺酸的生理功能及營養日漸重 視,因此牛磺酸的應用越來越廣泛。國際上發表許多關於牛磺酸的討論 (Chesney, 1985)。例如,Hayes 氏等在 1975 年提出貓缺乏牛磺酸可導致 失明。近年來人們已注意到牛磺酸在預防醫學中的應用,包括調節細胞 內外鈣平衡、直接參與細胞膜穩定、抗氧化作用及對外源性毒物的結合 等(Waterfield et al., 1993)。. 9.
(10) 二、 生 物 活 性 (一 ) 1.. 中樞神經系統 促進大腦及智力發育的作用 動物實驗證明,牛磺酸具有促進大腦及智力發育的作用,表現為學. 習、記憶能力的提高(韓, 1988)。其作用是調節體內微量元素的代謝,提 高大腦組織中與腦發育有關的必要元素鋅、銅、鐵及游離的氨基酸等含 量,進而促進大腦中的 DNA,RNA 及蛋白質的合成。牛磺酸缺乏的大 鼠,腦及智力發育受到不良的影響,表現與正常組有差異性。牛磺酸缺 乏的幼貓,其大腦重量也顯著低於補充牛磺酸組。牛磺酸在腦神經的增 殖週期中具明顯的促增殖作用(韓, 1989)。牛磺酸對神經細胞的分化成熟 也起著促進作用(韓, 1992),當牛磺酸缺乏,體外培養的人類腦神經的分 化明顯受阻。. 2. 抗 癲 癇 牛磺酸在中樞神經系統的作用除促進腦發育外,也是一種神經抑制 因子,可以抗痙攣作用;也是調節神經及調節滲透及抗氧化的因子 (韓,1991)。牛磺酸能拮抗多種物質誘發的癲癇,其可延長潛伏期,降低 發作次數及發作強度。對於酒精成癮後戒酒發生神經錯亂及局部痙攣, 牛磺酸有可使反應減輕(Thurston, 1987)。. (二 ) 視 網 膜 Hayes 氏等(1975)提出貓缺乏牛磺酸可導致失明。Geggel 氏等(1985) 報導長期進行胃腸道外營養注射(不含牛磺酸)的 23 名成年人及 21 名兒 童的觀察結果,兒童組空腹血中牛磺酸含量下降,視網膜色素出現瀰漫 性顆粒,另外錐體也異常。給與牛磺酸後,視網膜接近正常,而成人組. 10.
(11) 不受影響,證明幼兒在缺乏牛磺酸,會發生視力障礙。. (三 ) 心 血 管 系 統 牛磺酸在心血管組織中含量豐富,具有廣泛的心血管作用,一旦缺 乏,會導致心血管疾病(Huxtable, 1992)。. 1. 對 心 肌 的 保 護 作 用 近年來發現牛磺酸對心肌缺血再灌注損傷具有非常有效的防治作 用(劉, 1991)。其能通過抑制心肌脂質過氧化而保護心肌。此外,還能 抑制缺血在灌流時鈣內流,防止細胞內鈣離子過多。此外,另一研究證 實,用腎上腺素及高濃度的鈣灌流大鼠心臟造成心肌內鈣過多的模式 中,牛磺酸的治療具有抑制心肌組織和粒線體鈣聚積,增加心肌 ATP 含 量,因此牛磺酸有改善細胞內鈣過多的現象(王, 1993)。. 2. 心 衰 竭 牛磺酸可顯著改善瓣膜性和缺血性心衰竭的功能,並且全面改善病 人的血液動力學,它是治療心衰竭的有效藥物且無毒性,增加左心室的 收縮而不引起血壓降低,也不影響洋地黃類的藥物作用,還可降低洋地 黃的副作用(黃,1991)。. 3. 高 血 壓 牛磺酸可通過中樞及周邊降壓作用,僅對高血壓有降低作用,正常 血壓不受影響。在腦室注射血管收縮素 II 和腎上腺素引起的高血壓, 牛磺酸可改善之(Azuma et al., 1987)。此外,牛磺酸和 Mg2+ 可以拮抗 endothelin,防止血管平滑肌細胞(VSMC)過渡增生而造成高血壓疾病(吳. 11.
(12) 等 ,1991)。. 4. 動 脈 粥 狀 硬 化 動脈粥狀硬化有多種因素,其中包括血小板功能異常及膽固醇累 積。有學者指出血小板聚集與牛磺酸含量呈負相關,服用牛磺酸後,血 小板聚集降低(Pronczal et al., 1988)。此外也有人發現牛磺酸在體外實驗 明顯抑制花生四烯酸(arachidonic acid)、ADP 和膠原誘導的血小板聚集, 並且呈與劑量反應的關係,而體內的研究也指出有抑制血小板聚集的作 用(陳等,1991)。所以牛磺酸對動脈粥狀硬化疾病有防治作用。. (四 ) 肝 膽 系 統. 1. 肝 細 胞 保 護 作 用 正常大鼠肝和尿中牛磺酸含量明顯正相關,四氯化碳和半乳糖胺造 成大鼠肝損傷,均可使肝臟中的牛磺酸含量下降,顯示牛磺酸具有一定 的肝細胞保護作用(Waterfield et al., 1993)。. a ) 鈣離子 Nakashima 氏等(1990)發現在含鈣的介質中,牛磺酸可防止因缺氧而 造成的肝胞死亡,而在缺鈣的介質中則沒有這種作用,說明牛磺酸對肝 細胞的保護作用可能透過細胞鈣離子含量而定。. b) 抗氧化 牛磺酸可直接或間接抗氧化所造成的損傷。直接方式是利用分子中的 氨基與氧化劑結合而阻止氧化作用。如牛磺酸與強氧化劑 HClO 作用時. 12.
(13) 生成牛磺氯氨,從而解除了 HClO 對組織的氧化損傷作用。間接方式是 牛磺酸能穩定細胞膜成分,從而抵抗氧化劑的攻擊(Banks et al., 1991)。. c) polyamine 有文獻指出,缺少合成 polyamine 酵素 ornithine decarboxylase 時, 牛磺酸不具肝細胞保護作用,加入 polyamine 後,牛磺酸具有保護肝細 胞的功能,推測 polyamine 的存在與牛磺酸保護肝細胞的功能上有關 (Wu et al., 1999)。. 2. 肝 臟 纖 維 化 最近的研究顯示,牛磺酸對實驗大鼠肝臟纖維化具有改善的效果(陳 等, 1999),可以減少慢性肝損傷時,tissue inhibition of metalloproteinase (TIMP)、procollagen I 及 III 的基因表現,可能具有臨床治療功效的潛力 (Chen et al., 1999)。. 3. 參 與 肝 脂 質 代 謝 體外研究表明,人體肝細胞牛磺酸含量與膽汁酸合成速度,細胞游 離膽固醇濃度及高親合性低密度脂蛋白受體表達有關。對倉鼠、豚鼠和 小鼠的體內研究表明,補充牛磺酸可影響 7α -膽固醇脫氫? 和 HMGCoA 還原? 的活性。牛磺酸不僅與膽汁中的脂質分泌有關,而且與脂質 代謝有關係。用含牛磺酸的飼料餵大鼠可明顯降低血液及肝臟中的膽固 醇,也可以減少膽固醇結石的形成。大鼠飲用牛磺酸水溶液兩週後,肝 臟中的膽固醇、膽固醇脂及三酸甘油脂明顯低於正常組(周,1995)。. (五 ) 免 疫 系 統. 13.
(14) 牛磺酸具有保護淋巴細胞並可促進其增殖、產生抗體的作用,還可 促進巨嗜細胞產生 IL-1,增強噬中性白血球的吞噬殺菌的功能,提高人 和動物的特異性和和非特異性免疫功能。另外牛磺酸可以促使對 B 肝疫 苗無反應的小鼠產生抗 HBsAg 抗體(冉, 1993)。. (六 ) 其 他 牛磺酸還具有防治糖尿病(Maturo et al., 1987)、調節腦垂體分泌(Pow, 1993)、防治缺鐵性貧血及治療凍傷寒偏頭痛作用。. 第二節 肝臟之簡介. 一、 肝 臟 的 型 態 肝臟是最大的器官,也是最大的消化腺。其位於右側橫隔膜下腔, 重量約 1,200 至 1,500 公克,其表面光滑且非常脆弱,受到外力時容易 破裂,引起大出血。肝臟呈楔形,由鍊狀韌帶分左右葉。右葉較大,佔 肝體積的五分之四,左葉較小,佔肝體積的五分之一 ; 其於較小,包括 尾葉及方葉。. 二、 肝 臟 的 顯 微 結 構 肝臟是由大量的肝小葉組成,肝小葉為多邊柱體,它是以中央靜 脈為中心,單排的肝細胞呈現放射狀,成群的肝細胞與肝血竇(sinusoid) 存在著狄氏腔(space of Disse),然而肝血竇的周圍並不是完整的基膜, 因此肝血竇與狄氏腔呈現開放式循環。肝細胞的微絨毛深入狄氏腔,能 與血液接觸。而肝細胞膜通透性高能讓高分子量的物質進出,所以肝細. 14.
(15) 胞可與血液進行有效的物質交換。 三、 肝 臟 的 血 流 系 統 肝臟的血管特點有肝動脈和肝門靜脈的雙重血液供應,肝臟可以 在肝動脈的體循環下接受肺部及其他組織過來的氧氣及代謝物,也可以 由肝門靜脈的血液取得大量由消化道吸收的營養物質。肝臟也有兩條輸 出的通路,除了肝靜脈與體循環相關連外,還可以通過膽道系統與腸道 相連,使得一些肝內代謝產物、助消化物質及有毒物質或解毒物可以隨 膽汁的分泌而排入腸道,再經由糞便排出體外(見下圖)。. 肝血管圖:. 四、 肝 臟 的 功 能. (一) 膽 汁 的 分 泌. 膽汁由肝細胞分泌,它不僅促進脂類消化吸收,同時將體內某些代 謝產物及生物轉化產物如膽固醇、膽紅素及藥物、毒物等產物不斷排入 腸道隨糞便排出體外。所以他即是消化液也是排泄液。正常人肝細胞每. 15.
(16) 天分泌 500~1,000 ml 液體,比重約 1.01,稱為膽汁。肝膽汁進入膽囊後, 由於膽囊壁分泌大量黏液物質以及對液體的重吸收,再濃縮成 300~700 ml,呈現褐色黏稠不透明,比重約為 1.04,稱為膽囊膽汁,儲存於膽囊, 經膽總管排入十二指腸。 無論是膽汁或是膽囊膽汁,均有苦味,其組成除水之外還有固體物 質,以膽汁酸含量最高,佔總固體物質的一半以上,其它還有卵磷脂、 蛋白質、脂肪酸、膽固醇、膽紅素及無機鹽等。. (二 ) 蛋 白 質 的 代 謝. 1. 多 種 蛋 白 質 的 合 成 器 官 身體細胞雖都有合成蛋白質的能力,但以肝細胞為最強。據估計肝 臟合成的蛋白質佔全身蛋白質總量的 40 % 以上。肝臟內蛋白質代謝極 為活躍。肝臟會合成肝細胞本身的結構蛋白,並且會合成大部分血漿蛋 白,包括白蛋白、球蛋白、纖維蛋白原、凝血? 原等。. 2. 氨 基 酸 分 解 及 合 成 肝內含有豐富的與氨基酸代謝有關的? 類,因而氨基酸代謝很活 躍;如氨基酸的轉氨基,脫氨基及各別氨基酸的代謝過程,都能在肝臟 內進行。肝臟可以根據有機體的需要,合成一些非必須氨基酸,也可處 理多餘的氨基酸。例如將其轉變為糖、脂肪酸或氧化成二氧化碳和水以 及合成各種含氮化合物及尿素。當肝臟功能有障礙時,可能會導致血液 中的氨基酸濃度上升。. 3. 合 成 尿 素 肝臟能利用氨合成尿素。氨基酸分解代謝及蛋白質或其消化的中間. 16.
(17) 產物在大腸均會有氨氣產生。氨在經過肝臟鳥氨酸循環時合成尿素以解 毒。當肝障礙時,尿素無法合成,血氨增高,易生肝昏迷。 (三 ) 脂 肪 的 代 謝. 1. 促 進 脂 類 的 消 化 合 吸 收 肝能將膽固醇轉化為膽汁酸,膽汁酸是膽汁主要成分。膽汁酸鹽不 僅是乳化劑能乳化脂質,還能活化腸胰脂? ,從而促進脂類吸收。如果 肝障礙時,易引起腹瀉等症狀。. 2. 膽 固 醇 的 代 謝 器 官 肝臟是合成膽固醇的重要場所之一,血液膽固醇大部分來自肝臟, 一部份來自食物。在血液中游離膽固醇可經代謝接受卵磷脂分子上的脂 肪酸形成膽固醇脂。當肝功能障礙時,血漿中的膽固醇量不一定變化, 但膽固醇酯必然減少。而肝內膽固醇的轉化,約 80 % 以上是形成膽汁 酸鹽後排入膽道,所以膽道阻塞時,血漿膽固醇及酯類將增加。. 3. 磷 脂 、 脂 蛋 白 的 合 成 脂質的運輸和轉化均與肝臟密切相關,一方面無論從乳糜微粒、極 低密度脂蛋白中釋放的脂肪酸均能被血漿白蛋白結合成脂肪酸-白蛋白 複合體被運輸,另一方面,三酸甘油脂,膽固醇依不同比例,與極低密 度脂蛋白和高密度脂蛋白參與脂類的運輸與轉化。一旦合成磷脂的原料 缺乏或是肝功能障礙,三酸甘油脂將無法運出在肝中沈積,形成脂肪 肝。正常肝脂類 4 % ~ 7 %,脂肪肝則大於 10 %。. (四 ) 肝 臟 的 其 他 功 能. 17.
(18) 肝臟分泌的膽汁酸可協助脂溶性維生素的吸收,當膽道阻塞時,脂 類及脂溶性維生素的吸收發生障礙。而維生素 K 參與凝血過程,故維生 素 K 吸收障礙時會有出血情況。肝臟有儲存維生素 A、D、E、K 及 B12 的功能。肝臟儲存的維生素為體內的 95 %。另外,肝臟也可進行生物轉 化,可將外源性物質及內源性營養物進行化學轉變,增加極性使其易隨 膽汁或尿液排出。. 第三節 肝硬化簡介. 肝硬化(liver cirrhosis)是一種常見的慢性疾病。由於各種原因引起肝 細胞瀰漫性變性、壞死,以及伴隨著炎症反應,間質結締組織增生,肝 細胞再生等病變反覆交錯進行。至最後,肝臟正常小葉結構逐漸被增生 的結締組織纖維取代,形成假小葉(pseudolobule),導致肝臟肝臟全面改 造,稱為肝硬化。肝硬化有多種分類,其中以門脈性肝硬化最常見,其 次是膽汁性肝硬化,以下個別介紹。. (一) 門 脈 性 肝 硬 化 : 多數研究認為肝硬化是由不同原因造成的,如病毒性肝炎、慢性 酒精性中毒、藥物性肝炎或是營養缺乏引起的脂肪肝。然而造成肝纖維 化的原因通常是兩種因素造成,例如酒精性肝炎加上營養缺乏,或是營 養缺乏加上藥物因素而導致。. 1. 病 毒 性 肝 炎 為世界最常見的肝硬化原因,其中在中國人是以慢性 B 型肝炎及. 18.
(19) 非 A 非 B 型肝炎發展為較多。很多資料顯示,慢性 B 型肝炎對門脈性 肝硬化有關連,例如在免疫組織染色法檢驗出 HBsAg。. 2. 慢 性 酒 精 中 毒 長期酗酒被認為是肝硬化的一個重要因素,因此又有人稱為酒精 性肝硬化。在歐美較常發生,中國人較少,據統計肝硬化患者慣性酗酒 者僅佔 13.0 % ~ 20.5 %,曾經有人作實驗,以大量乙醇餵食狒狒,並給 予高蛋白、纖維素及膽鹼飼料,但仍導致肝硬化,因此乙醇對肝有直接 的毒性作用。. 3. 營 養 缺 乏 實驗證明,大鼠食物中如果長期缺乏膽鹼、蛋白質、尤其是氮氨 酸和維生素,可導致肝細胞內中性脂肪沈積而引起脂肪肝。經過一年, 肝實質細胞可發生脂肪囊腫(fatty cyst),繼之出現肝纖維化。但也有學者 持相反意見,認為只因營養缺乏所引起的脂肪肝,至少在人不能導致肝 硬化,因為除了營養缺乏外,似乎需要損害肝細胞代謝的機能從而引起 肝細胞壞死。. 4. 化 學 毒 物 及 藥 物 中 毒 很多實驗證明,經不同途徑給大鼠以化學藥物,如四氯化碳、二甲 基氨基偶氮苯 (diaminobenzidine, DAB)、二甲基亞硝胺(dimethylnitrosamine, DMN)及 thioacetamide 均可作為實驗性肝硬化的模式,其他氯 仿、黃磷、砷也會造成肝硬化。雖然由這些毒物引起的肝硬化與人類肝 硬化有些差異,但在病理上仍認為是門脈型肝硬化。所以像四氯化碳或 其他服用藥物均可能引起慢性肝炎並同時肝纖維化,而這些過程均會導. 19.
(20) 致結締組織增生,並且反覆出現肝細胞壞死,肝細胞結節再生,最後自 然是肝硬化。. (二) 膽 汁 性 肝 硬 化 : 任何原因引起的肝內、外膽管疾病,導致膽汁淤積,皆可發展為膽 汁性肝硬化。分原發性膽汁性肝硬化(pimary bliary crrhosis, PBC)和續發 性膽汁性肝硬化(secondary biliary cirrhosis, SBC)。後者由肝外膽管長期 阻塞引起。一般認為 PBC 是一種自身免疫性疾病,淋巴細胞被激活後, 攻擊中、小膽管,導致炎症反應。組織學上,頗似宿主對移植器官的排 斥反應。與肝臟同種移植的排斥反應有許多相似之處。臨床上,病情緩 解與惡化交替出現,常伴有其他自身免疫性疾病,如乾燥綜合症,系統 性紅斑性狼瘡病,類風濕性關結炎及慢性淋巴細胞性甲狀腺炎等。. 第四節 肝纖維化在分子生物學上的研究. 肝纖維化的形成是由於長期的慢性肝損傷,造成庫氏細胞(kupffer cell)、血小板、血管內皮細胞等釋放細胞激素,細胞激素作用於星狀細 胞,使星狀細胞活化成纖維母細胞,產生大量的細胞外基質,例如: 第 I、 III 型膠原蛋白而造成肝纖維化。活化的星狀細胞雖能釋放 matrix metalloproteinase (MMP,即 collagenase) 來降解纖維狀膠原蛋白,但也 會產生 tissue inhibition of metalloproteinase (TIMP)來抑制 MMP 降解纖 維蛋白的作用,造成肝纖維化更加嚴重。. 一、 星狀細胞. 20.
(21) 肝 臟 星 狀 細 胞 是存在狄氏腔(space of Disse),主要功能是儲存 vitamin A,星狀細胞佔肝臟非實質細胞(non-parenchymal cells) 的三分之 一,或肝臟細胞總數的 15 % 。星狀細胞在正常肝臟執行重要功能,包 括: (1) retinoids 的代謝,星狀細胞儲存了約全身 80% 的 vitamin A 的代 謝物 retinoids (Senoo et al., 1988) ; (2) 細胞激素的分泌,例如: 表皮生 長因子 (epidermal growth factor, EGF)、 肝細胞生長因子 (hepatocytes growth factors, HGF)等,這些因子可以維持肝細胞的量與質; (3) 調控血 流,由於星狀細胞接近神經(Ueno, 1997),故星狀細胞可能受到神經刺激 而收縮(Friedman, 1996)。 隨著肝毒物造成的傷害,肝臟中環境的改變,例如: 庫氏細胞、內 皮細胞釋放細胞激素,或活性氧化物、氧化壓力的增加,星狀細胞會因 此活化,由富含 vitamin A 的休止狀態,轉變為活化態的星狀細胞。. 二 、 TGF- beta. 刺 激 肝 臟星 狀 細胞 合 成細胞外基質的最主要細胞激素是 Transforming growth factor beta1 (TGF-beta1) (Czaja et al., 1989)。TGFbeta1 是由兩個相結構相同或相近的、分子量為 12.5 kDa 合併單位借二 硫鍵連接的雙體。TGF-beta 的生物學功能研究主要在炎症、組織修復和 胚胎發育等方面,近年來發現 TGF-beta 對細胞的生長、分化和免疫功能 都有重要的調節作用。TGF-beta 家族中,包括有三種 isoform ( beta1, beta 2, beta 3)。一般來說,TGF-beta1、beta2 和 beta3 功能相似,TGF-beta 對間質起源的細胞起刺激作用,而對上皮或神經外胚層來源的細胞抑制 作用。 當組織受損而產生發炎反應時,發炎細胞或血小板會釋放 TGF-beta1. 21.
(22) ,來調控白血球的補給與活性。接下來 TGF-beta1 釋放的量與活性的表 現,取決於星狀細胞內的結合蛋白的數目,也就是 TGF-beta1 接受器的 多寡(Gong et al., 1998)。當 TGF-beta1 與 TGF-beta1 receptor (type 、 I II) 結 合,則活化的星狀細胞 TGF-beta1 gene 會有增加的現象(Friedman et al., 1996; Kim et al., 1998),使 TGF-beta1 生成增加,進而刺激星狀細胞生成 更多的膠原蛋白。 在肝纖維化的過程中,TGF-beta1 扮演著很重要的角色,肝臟的星 狀細胞受 TGF-beta1 刺激活化成肌纖維母細胞,造成肝纖維化。. 三 、 TIMP 及 MMP. 當持續肝受損時,肝臟星狀細胞就會增殖,而且在型態上會形成 smooth muscle α-actin positive myofibroblast (activated HSC),並且產生多 樣化的膠原蛋白及非膠原蛋白蛋白(Friedman et al., 1989)。在一般慢性受 損的肝中,膠原蛋白型 I 和 IV、 undulin、 elastin 和 laminin 則 會增加 (Schuppan D, 1990)。細胞外基質的膠原蛋白中,膠原蛋白型 I 是增加 最多的,再來是膠原蛋白型 III 和 IV,隨著肝損傷的日趨嚴重,就會產 生一些無法由巨噬細胞分解的結構蛋白,壞死細胞形成結(nodules),造 成肝硬化。 肝纖維化的原因,亦是細胞外基質的合成失衡, 而肝纖維化本身 即是細胞外基質生成及其降解酵素不斷調控的過程。細胞外基質的降解 主 要 是 matrix metalloproteinase (MMP)酵素的催化, 其種類包括有 collagenase、stromelysins、gelatinase 及 membrane type (MT)-MMPs, 而這些酵素主要是受到 tissue inhibition of metalloproteinase (TIMP)抑 制。. 22.
(23) TIMP 的家族至今已被證實包括有四種, TIMP-1、 TIMP-2、 TIMP-3、TIMP-4,而 TIMP-1 及 TIMP-2 已被證實可以抑制所有已知的 MMP 活 性 。 一般認為, 在肝臟病理狀態, TIMP-1 的重要性大於 TIMP-2 。TIMP-1 是一種 29-kD glycoprotein,主要功能是抑制膠原蛋白 水解? MMP 的活性。在人類及動物模式中,TIMP-1 及 TIMP-2 都增加。 TIMP 的主要來源是被活化的肝臟星狀細胞,而在肝纖維化的初期, TIMP 會被表現出來,相對於 TIMP,interstitial collagenase (MMP-1 在 人類,MMP-13 在大鼠)活性未改變(Benyon et al., 1996)。因此當 TIMP : MMP 動態的比值增加時,就會增加胞外基質的沈積。. 四、肝細胞生長因子. 肝細胞生長因子(HGF)最早在 1984 年從肝切除後的剩餘組織中被 發現,因最初被發現可刺激肝細胞合成 DNA 而命名之(Nakamura et al., 1984)。在正常的情況下 HGF 在組織中以非活性的形式存在。非活性的 HGF 必? 經蛋白水解後才能發揮其生物活性。非活性的 HGF 是由 728 個氨基酸構成的單鍵分子,活化態時是一條重鏈(α chain)和一條輕鏈(β chain)由雙硫鍵連接而成的二聚體。 HGF 是很強的促肝細胞增生因子,不僅能刺激肝細胞再生,也能 促進肝功能回復。肝毒素 CCl4 和氨基半乳糖(D-galactosamine)造成大鼠 急性肝損傷中, HGF mRNA 顯著增加(Asam et al., 1991),但 HGF 在慢 性肝損傷表現情況尚不明。. 五 、 S-腺 ? 甲 硫 氨 酸 (S-adenosylmethionine, SAM). 23.
(24) Kinsell 氏等人(1947)首次發現在肝硬化病人的 methionine途徑有損 害。methionine 的第 3、4 兩個碳原子經酵素作用與 ATP 結合形成活化 狀態的 SAM(見下圖)。Methionine adenosyltransferase (MAT)是 methionine 形成 SAM 的重要催化酵素(Cantoni, 1953)。. http://www.indstate.edu/thcme/mwking/sam.gif. SAM 是許多甲基化反應的甲基提供者,對 polyamine 及 glutathion 的合成相當重要,在 folate cycling 也有其重要性(見下圖)。除此之外, MAT 在肝細胞功能的調控,如肝再生、分化及肝損傷也扮演相當重要 的角色(Paneda et al., 2002)。. http://www.humboldt.edu/~rap1/BiochSupp/note_pics/aaMetab/CholineSyn.gif. 24.
(25) http://lpi.oregonstate.edu/infocenter/othernuts/choline/cholinter.html. 在哺乳動物中 MAT 有三種形式,MAT I、MAT II 及 MAT III 由兩 種基因 mat1a 及 mat2a 所調控。mat1a 僅在肝臟中表現,產生 MAT III (dimmer) 及 MAT I (tetramer) 。mat2a 則是產生 MAT II,其廣泛分佈於 肝臟外的組織,雖存在於新生兒的肝臟中,但長大後即被 mat1a 所取代。 在肝硬化病人(Cabrero et al., 1988),大鼠肝損傷實驗如酒精(Lieber et al., 1990)、四氯化碳(Corrales et al., 1992)、paracetamol (Stramentinoli et al., 1979)、buthionine 及 sulfoximine (Corrales et al., 1991)等,MAT I/III 的 活性明顯下降。另一方面,當發生肝癌 (Cai et al., 1996)、肝臟切除手術 (Huang et al., 1998)及 thioacetamide 誘導肝纖維化時 (Huang et al., 1999),mat2a 表現會明顯上升,mat1a 開始下降。mat1a 及 mat2a 之間 的轉換機轉尚不明確。. 六 、 麩 胱 甘 ? (GSH). 麩胱甘? (GSH)在細胞內抗氧化系統中扮演有重要的角色,GSH 是由 三個胺基酸所構成的。GSH 的生理功能很多,主要有:(1)可預防過氧化. 25.
(26) 物的毒性,本身當作受質(substrate)利用 glutathione peroxidase (GSH- Px) 來 移 除 過 氧 化 氫 (hydrogen peroxide) 或 其 他 有 機 過 氧 化 物 (organic hydro-peroxides);(2)參與解毒代謝,可與多種外來毒物(xenobiotics)或內 生性反應物透過 glutathione S-transferase 催化進行結合,增加外來毒物的 水溶解能力,達到解毒的目的(Ishige et al., 2001)。. 第五節 四氯化碳肝纖維化. 本實驗以四氯化碳作為誘發大鼠肝損傷模式。四氯化碳在工業上作 為脫脂劑及有機溶劑。在 1936 年,有人就提出多重四氯化碳會誘導大 鼠肝硬化的形成(Cameron and Karunaratue, 1936)。連續五星期的投予, 會造成肝硬化、肝結節及膽管的增生。Sipes 氏等人研究指出四氯化碳 的造成肝損傷可能與 Kupffer cell 活性有關(Sipes et al., 1989)。Kupffer cell 為內皮網狀系統之一,與肝臟的排除病菌有關。四氯化碳的投予會 誘導 Kupffer cell 活化而產生大量的活性氧(ROS)對生物造成破壞。 此外也有人提出,四氯化碳進入肝細胞後,會經由內質網中的 cytochrome P-450 的氧化? 還原代謝。cytochrome P-450 氧化? 釋放出 一個電子使四氯化碳還原成三氯甲基,其所形成的自由基以共價鍵方式 與脂質結合,造成脂質過氧化,進而破壞細胞膜結構,嚴重時可能造成 細胞死亡(Reynolds and Ree, 1991)。 綜合上述,四氯化碳是一種具選擇性的肝毒物,當其進入肝臟時, 病理上可發現肝小葉中央靜脈週圍會壞死,並有纖維增生。此外,四氯 化碳的誘導途徑有口服、腹腔注射及皮下注射等,所以四氯化碳是理想 的肝纖維化的動物模式。. 26.
(27) 第二章 實驗材料及方法 一、動物 使用 Wistar 雄性大白鼠,體重約 180-220 公克。購自國家實驗動物 繁殖及研究中心。飼養環境維持溫度 22 ± 3°C,相對溼度 55 ± 5 % ,明、 暗各十二小時的環境。餵食標準飼料(福壽公司),飲水經過逆滲透處理。. 二、四氯化碳誘發慢性肝毒性 實驗前將大鼠分四組,每組十隻,一為控制組不投予 CCl4,其餘 三組實驗組每週兩次口服投予 CCl4 (20 %,0.2 ml/rat ),為期八週。其 中兩組每日口服投予牛磺酸(購自 ACROS ORGANICS;400、800 mg/kg) 水溶液,另一組僅投予 RO 水(逆滲透水)。第八週時,犧牲大鼠,解剖 取出肝臟及脾臟稱重。肝最大葉分三部分,第一部分固定於 10 % 中性 福馬林溶液中供病理切片觀察,第二部分稱重後烘乾供膠原蛋白含量測 定,第三部分快速冷凍於液態氮供 mRNA 的抽取。其餘肝臟依解剖相 關位置分成四袋,儲存於 -80°C,供脂質過氧化、抗氧化酵素活性及西 方墨點法等實驗測定。. 三、肝功能之測定 將採取收集的血液靜置,於 4°C 離心十分鐘(3,000 rpm)。取上層 血清以自動生化儀(COBAS MIRA Plus; Roche)檢測麩氨酸草乙酸轉氨? (GOT)、麩氨酸草丙氨基轉氨? (GPT)、總蛋白(total protein)。 使用 Roche 公司的生化測定試劑(test kits) : AST (GOT ; glutamate oxaloacetate transaminase ): Art. 0736414 ALT (GPT ; glutamate pyruvate transaminase ): Art. 0736384. 27.
(28) 四 、 羥 脯 氨 酸 (hydroxyproline)含 量 測 定 Hydroxyproline 的含量測定是依照 Neuman and Logan (1950)的方 法。將肝組織秤取約 600 mg 切成細長條狀於 100°C 烘箱中隔夜乾 燥。秤取乾燥肝臟組織約 200 mg 放於試管中,加入 5 ml 的 6 N HCl, 於 100°C 烘箱中放置 16 小時水解之。 冷卻後取 1 ml 水解物以 10,000 rpm 離心 30 分鐘。取 0.2 ml 標準品 hydroxyproline 或上清液加 至 0.2 ml 的 6 N NaOH 中和水解物,之後加入 0.6 ml 水。加 1 ml 新 鮮配置 0.01 M 的 CuSO 4 至試管內,取 1 ml 2.5 N NaOH、1 ml 6% H2O2 先後加入試管內,震盪混合 5 分鐘後,加熱和振搖可破壞過多的 過氧化物。取出以冰水冷卻,加入 2 ml 之 5 % p-dimethyl aminobenzaldehyde 及 4 ml 之 3 N H2SO4 振盪均勻,於 70°C 水浴 16 分 鐘,以冰水冷卻至室溫,以分光光度計(HITACHI U-2001; Japan) 於 540 nm 測定吸光值。利用純品 hydroxyproline 當作標準,以多重校正曲線 計算 hydroxyproline 的含量。 hydroxyproline 的含量以 µg/g wet liver 表示。. 五 、 肝 臟 脂 質 過 氧 化 (lipid peroxidation) 之 測 定 依據 Ohkawa et al. (1979)的方法測定脂質過氧化。取肝組織 1 g 加 10 ml KCl (1.15 %)溶液,以均質機均質化。取 0.2 ml 均漿順序加入 0.2 ml sodium dodecyl sulfate (8.1 %)、1.5 ml acetic acid (20 %),並以 NaOH 調整酸鹼度(pH 3.5)。加入 1.5 ml 2-thiobarbituric acid (0.8 %),再以去離 子水調整容積為 4 ml,99°C 熱水中加熱一小時。取出,以冰水冷卻, 加入 1 ml 去離子水及 5 ml 的 n-butanol/pyridine 混合溶液(15:1,v/v),劇 烈震盪均勻後,在 25°C 離心 10 分鐘(4,000 rpm),取有機層以分光光 度計(HITACHI U-2001; Japan)於 532 nm 測定吸光值,以多重校正曲線. 28.
(29) 計算濃度。肝組織的脂質過氧化量以 nmol malondialdehyde (MDA)/mg protein 表示。同時依照 Lowry 等人(1951)所述的方法,測定肝組織蛋 白含量,以牛血漿蛋白(Bovine serum albumine, BSA)為標準品。. 六 、 肝 臟 Glutathione (GSH)含 量 測 定 實驗依照 Sedlak and Lindsay (1968)的方法。取肝組織 0.4 g 加 16 ml EDTA (0.02 M)溶液,以均質機質化。取 5 ml 肝均漿順序加入 4 ml 蒸餾水、1 ml trichloroacetic acid (50 %),震盪混合均勻,離心 15 分鐘 (3,000 xg)。取上清液 2 ml 加入 4 ml Tris buffer 與 0.1 ml 5,5’- dithio bis (2-nitro benzoic acid) (0.36 % in methanol),混合均勻後以分光光度計 於 412 nm 測 定 吸 光 值 , 以 多 重 校 正 曲 線 計 算 濃 度。 肝 組 織 的 glutathione 以 µmol/g wet liver 表示。. 七 、肝 臟 超 氧 化 物 歧 化 ? (superoxide dismutase ; SOD)活 性 測 定 實驗參照 Marklund (1974)等人所描述之方法進行,取肝組織 0.5 g,加入 5 ml buffer (0.32 M sucrose、1 mM EDTA、10 nM Tris-HCl,pH 7.4),以均質機均質化,高速離心 30 分鐘(13,600 ×g、4°C)。取上清液 稀釋,稀釋液取 50 µl,再加上 100 µl Tris buffer (pH 8.2,50 mM)、去 離子水 830 µl,最後加入 20 µl pyrogallol (10 mM),混合均勻後,於 420 nm 測量吸光值。每隔 40 秒測量一次,共測量 5 分鐘,計算每分鐘吸 光值變化速率。以多重校正曲線計算濃度。SOD 活性以單位時間內抑 制 pyrogallol 自動氧化速率達 50 % 時的量,為一單位(U)。組織內 SOD 活性以 U/mg protein 表示。. 八 、 肝 臟 過 氧 化 氫 ? (catalase)活 性 之 測 定. 29.
(30) 實驗參照 Aebi (1984)所描述之方法進行。取肝組織 0.5 g 加入 5 ml 磷酸鹽緩衝液(Na2HPO4 0.01 M,NaH2PO4 0.01 M,NaCl 2 M,pH 7.4),以均質機均質化,離心 10 分鐘(700 ×g,4°C),取上清液 0.9 ml 加 入 0.1 ml Triton X-100 (10 %),混合均勻。取混合液以磷酸鹽緩衝液(50 mM,pH 7.0)稀釋,將 4 ml 稀釋液加入 2 ml H2O2 (30 mM),混合均勻 後於溫控 25°C、波長 240 nm 條件下測吸光值,每隔 5 秒測量一次, 時間總共 1 分鐘。計算求得反應速率常數 K (1/min)。組織 catalase 活 性以 K/mg protein 表示。 K = (2.3/Δt) logA1/A2 Δt :時間間隔(分鐘)。 A1 : T1 時間,檢品之吸光值。 A2 : T2 時間,檢品之吸光值。 九、肝臟麩胱? 過氧化? (Glutathione peroxidase ; GSH-Px)活性之測定 實驗參照 Hafeman 等人(1974)所描述之方法進行,取肝組織 0.5 g 加 5 ml buffer (0.32 mol/L sucrose、 1 m mol/L EDTA 、 10 nmol/L Tris-HCl,pH 7.4),以均質機均質化,高速離心 30 分鐘 (13,600 ×g、4 °C)。取上清液稀釋。稀釋液取 0.4 ml 加入 0.4 ml glutathione (1 mM ), 置於 37°C 水浴加熱 3 分鐘。之後再加入 H2O2 0.2 ml,置於 37°C 水 浴加熱 5 分鐘。然後迅速置於冰中冷卻。冷卻後加入 4 ml 偏磷酸鈉溶 液(偏磷酸鈉 0.635 %,NaCl 30 %,EDTA 0.2 % ),混合後以 3,000 rpm 離心 10 分鐘。取上清液 2 ml,加入 2 ml Na2 HPO4 溶液(0.4 M)與 1 ml 5,5'-dithio-bis (2-nitrobenzoic acid) (0.04 %,sodium citrate 1 %)溶液,混 合均勻後於 412 nm 測量吸光值,以多重校正曲線計算濃度。另外測量 非酵素所造成 GSH 的氧化,其步驟與測量酵素反應步驟相同,兩者差. 30.
(31) 別只是將組織稀釋液以蒸餾水取代。GSH-Px 以 Hafeman 單位 (U)表 示,Hafeman 單位定義為單位時間內檢品消耗之 glutathione (GSH)濃度 對數值(log[GSH]E),減去非酵素反應消耗之 glutathione (GSH)濃度對數 值(log[GSH]NE),所得值之千分之一為 GSH-Px 活性單位(U) : U = (log[GSH]E - log[GSH]NE)/1000×Δt 組織內 GSH-Px 活性以 U/mg protein 表示。. 十 、 反 轉 錄 聚 合 ? 鍊 鎖 反 應 (reverse transcriptase polymerase chain reaction ; RT-PCR) (一 ) 全 量 RNA 之 萃 取 : 全量 RNA 之萃取是參考 Chomczynski 等人(1987)所描述之方法進 行,取 0.2 g 肝組織於均質器內,加入 2 ml TRI Reagent® 於低溫下均 質。取適量的均質液於 eppendorf 內,在 4°C 下靜置 5 分鐘。再使用 1 ml 滅菌針筒和 20 G 注射針頭重覆均質之。加入 0.2 ml chloroform (SHOWA),上下強烈振搖約 20 秒,再於 4°C 離心(14,600 ×g) 15 分 鐘,取 80 % 上清液至已滅菌的 eppendorf,加入等體積的 isopropanol (WAKO)震盪混合均勻,於 -20°C 下靜置。隔日,以於 4°C 離心(12,000 ×g)10 分鐘。移除上清液,加入 75 % 酒精清洗 RNA pellet ;於 4°C 離 心(7,500 ×g) 5 分鐘去酒精,於室溫下風乾(約兩分半鐘)。加 70 µl DEPC 處理過的去離子水溶解之, 最後再以分光光度計(HITACHI U-2001; Japan) 於 260 nm 及 280 nm 下測量吸光值,估算 RNA 的純度及濃度。 檢測後將 RNA 調整為適當濃度,置於 -80°C 冷凍備用。. (二 ) Primer (引 子 ) 的 設 計 與 合 成 :. 31.
(32) 進行反轉錄聚合? 反應時所需的引子,是參考有關文獻(Shanthi et al , 1988)及使用電腦軟體設計(Table 1)。引子的合成,則是委託生工有限 公司合成。. (三 ) RT-PCR: 1.取 0.2ml 的微量離心管,將 One-Step RT-PCR 試劑加入如下: NO. 1 2 3 4 5. Components 2× Reaction Mix RT/P LATINUM Taq Mix Sense Primer (10µM) Anti-sense Primer (10µM) Template RNA (1µg/µl). Volume 12.5 µl 0.5 µl 0.5 µl 0.5 µl 10.0 µl. 2.PCR 反應的條件 mRNA HGF TGF-ß1 TIMP-1 Mat Ia Mat IIa Procollagen I GAPDH. Annealing temp (°C ) 52.0 45.0 57.6 47.0 52.0 59.0 52.0. No.of cycles 35 35 30 40 40 30 35. 將上述反應物混合均勻,置入 GeneAmp PCR system 2400 (PERKIN ELMER)並輸入反應條件後,進行 RT-PCR。. (四 )洋 菜 凝 膠 電 泳 (Gel electrophoresis): 以 0.5× TBE buffer 配製 2 % agarose gel,以微波爐加熱直到完全溶 解。微溫時進行造膠,待膠硬化加入 running buffer (0.5× TBE buffer)。. 32.
(33) 將 DNA 及 marker loading 在 well 中以 100 伏特電壓下進行電泳。 完成後,DNA 以 ethidium bromide 染色,再以 UV 激起螢光呈色後以 拍立得相機紀錄之。將相片掃描後以 Kodak Digital Science™ 1D 影像分 析電腦軟體定量其光密度。本實驗是以 GAPDH 當作對照,如欲求特 定 mRNA 表現量,將特定 mRNA 光密度除以 GAPDH 光密度值,其 相對量即為所求。. 試劑: 1.TRI Reagent® (MRC,Molecular Research Center) 2.Super Script™ One-Step RT-PCR with PLATINUM® Taq (Invitrogen) 3.Agarose (J.T Baker) 4.100 bp DNA Laddr (Gene Marker) 5.TBE Buffer (Sigma) 6.Ethidium bromide (Sigma). 十 一 、 西 方 點 墨 法 ( western blot ). (一) 蛋 白 質 萃 取 取 0.1 g 肝臟組織加入 1 ml 的 lysis buffer,在冰上加以剁碎並且 均質。將均質液放入離心管,進行高速離心(50,000 ×g),15 分鐘。 (二) 蛋 白 質 定 量 從離心過的均質液取上清液,加入 coomassie blue G-250 反應, 以可見光 595 nm,測定吸光值。同時以蛋白質標準品(BSA)所測 得的標準曲線,來計算各個樣本的濃度。 (三) SDS-PAGE 調整各組樣本蛋白質濃度為一致, 以一定量的蛋白與四倍的. 33.
(34) loading buffer 以四比一的比例混合均勻,於 100°C 下水浴加熱五 分鐘,使蛋白質四級結構瓦解成一級結構。將加熱完成的樣本 loading 在 SDS-PAGE (15 % running gel 及 4 % stacking gel),以 50 volt 跑 stacking gel,最後再以 100 volt 跑 running gel。將跑完 的膠片取出,以染劑 Commassie blue R-250 染一小時,再以脫色 劑(5 % acetic acid, 50 % methanol)脫色,直到 band 清晰可見,比 較 band 的寬度及顏色。 (四) 西 方 點 墨 法 結束電泳,將膠片小心取出,利用轉漬設備及 PVDF 轉漬膜,於 轉漬緩衝液中將膠內蛋白轉漬附著於 PVDF 膜上,轉漬過程於室 溫下以 100 volt 電泳 90 分鐘。將轉漬膜浸潤於 20 毫升含有百 分之五脫脂奶粉的 PBS-T (blocking solution)以牛奶蛋白覆蓋於轉 漬膜上尚未有蛋白接合的位置,於室溫下慢速搖晃 1 小時。倒掉 blocking solution 後,加入所期望的特定蛋白專一抗體溶液,於 4°C 下反應整夜。隔日,倒掉抗體溶液,加入 PBS-T 溶液於室溫下搖 晃五分鐘,倒掉 PBS-T 清洗液後再重覆加入 PBS-T 溶液清洗轉 漬膜,前後共四次,最後一次倒掉清洗液,加入過氧化氫酵素接 合的二次抗體(peroxidase-conjugated secondary antibody)溶液以結 合於轉漬膜上的初級抗體,在室溫下以中等速度搖晃一小時,反 應完後,倒掉二次抗體溶液,並加入 PBS-T 溶液清洗殘餘的二級 抗體溶液四次。於暗房中,將化學螢光試劑與轉漬膜於室溫下反應 一分半鐘。反應完後,將轉漬膜瀝乾,迅速放入壓片夾內使底片 感光,時間依螢光強弱決定。感光完的底片再進行顯色及固定。. Lysis buffer (pH=7.3)製備. 34.
(35) Tris-HCl. 20 mM. Glycerol. 10 %. Sodium chloride. 137 mM. Triton X-100. 1%. Protease inhibitor Cocktail. 1 mM. 轉漬方法 著 手 套 將 轉 漬 設 備 中 的 海 綿 (sponge)與 裁 成 適 當 大 小 的 濾 紙 (Whatman 3MM paper)浸泡於一倍的 Transfer buffer 中,再與 methonal 處理後的 PVDF 轉漬膜及電泳膠片於 Transfer buffer 蓋下依序排列,由陰極至陽極為海綿、濾紙、電泳膠、轉漬膜、 濾紙,及海綿。放置時確定各層無氣泡並確認所接電源極性方向 正確。此時即可進行轉漬。. Transfer buffer (pH 8.3)(10×) Tris base. 25.0 mM. Glycine. 192.0 mM. SDS. 3.5 mM. Methanol. 20.0 %. Blocking solution PBS (pH=7.4) Tween-20. 0.5 %. Skin milk. 5.0 %. 專一抗體溶液 TGF-beta1 抗體(Santa Cruz)以百分之五脫脂牛奶的 PBS-T 稀釋. 35.
(36) 二次抗體溶液 抗體(Amersham Bioscience)以百分之五脫脂牛奶的 PBS-T 稀釋. 十二、病理檢驗 肝組織在 10 % 的中性福馬林固定一週,再將此組織以脫水滲臘機 (LIPSHOW)先進行脫水,再進行石腊包埋。包埋後以旋轉式切片機切成 約 3-5 µm 的厚度。再進行肝臟膠原纖維特殊染色(Masson stain),於光 學顯微鏡下觀察肝細胞之特異情況及組織病理變化,比較並拍照評估肝 纖維化程度。. 十 三 、 cDNA 基 因 微 陣 列 試 驗. (一 ) 全 量 RNA 之 萃 取 詳見反轉錄聚合? 鍊鎖反應. (二 ) 晶 片 之 操 作 正常組、四氯化碳組、四氯化碳+牛磺酸水溶液高劑量組。每組抽 RNA 委託百恩諾生物科技公司進行兩對晶片操作,即正常組和四 氯化碳組一對、四氯化碳組和四氯化碳+牛磺酸水溶液高劑量組一 對。每對 RNA 分別先以 Cy 3 及 Cy 5 兩種螢光試劑加以標定再 與基因晶片上的 cDNA 進行雜合反應,反應完成後之晶片經過洗 去多餘物質及乾燥後,再以雷射光打出波長 635 nm 及 523 nm 的 光源,以激發晶片中的螢光試劑 Cy 3 及 Cy 5,並且以軟體加以判 讀所激發出的訊號強弱,並計算其訊號強度以作為量化之數據,進. 36.
(37) 而得知每一基因之表現情形,而得到的資料再交由本研究室解析。 十四、統計方法 以單尾變異數分析(One-way analysis of variance; ANOVA)方法分析 ,並進行 Dunnett 測試,以 P 值小於 0.05 為有差異。. 37.
(38) 第三章 研究結果 一、對肝臟和脾臟重量的影響 CCl4 處理的大鼠肝臟,平均重量為 23.1 ± 0.8 公克明顯高於正常 對照組平均肝臟重量 14.9 ± 0.5 公克(P <0.001) 。牛磺酸 400 mg/kg 的 重量為 21.6 ± 0.6 公克,與 CCl4 組比較不具統計差異,牛磺酸 800 mg/kg 組的重量為 20.0 ± 0.8 公克,與 CCl4 組比較有明顯減輕作用(P <0.01),如 Fig.1 (A)。CCl4 處理的大鼠脾臟平均重量為 2.01 ± 0.22 公 克明顯高於正常對照組平均重量 1.00 ± 0.03 公克(P <0.001),牛磺酸 400 mg/kg、800 mg/kg 組的平均重量分別為 1.63 ± 0.13 公克及 1.53 ± 0.11 公克,與 CCl4 組比較,雖不具統計差異,但有減少脾臟腫大的傾 向,Fig.1 (B)。. 二、血清生化值的影響 CCl4 處理的大鼠,第一、三及第六週,其血清 GOT 及 GPT 值明 顯高於正常組,最終血清(第八週) GOT 及 GPT 值為 1,856.9 ± 237.5 U/L 及 1,866.4 ± 247.3 U/L,也明顯高於正常對照組的 90.1 ± 4.2 U/L 及 65.7 ± 4.4 U/L (GOT: P <0.001; GPT: P <0.001),牛磺酸 400 mg/kg、800 mg/kg 組的第一、三及第六週,血清 GOT 及 GPT 值與 CCl4 組比較 沒有差異。牛磺酸 800 mg/kg 組最終血清 GOT 及 GPT 值分別為 1247.0 ± 103.4 U/L 及 506.8 ± 50.7 U/L,與 CCl4 組比較具統計差異(P <0.05),Fig.2 (A)。牛磺酸 CCl4 400 mg/kg 的 GPT 值為 1,101.6 ± 139.8 U/L,與 CCl4 組比較明顯下降(P <0.05),Fig.2 (B)。. 三、 對 肝 臟 蛋 白 質 及 hydroxyproline 含 量 的 影 響. 38.
(39) CCl4 誘發大鼠慢性肝損傷,其肝臟蛋白質平均含量為 160.4 ± 6.3 mg/g tissue,明顯低於正常對照組的 242.9 ± 5.8 mg/g tissue (P <0.001)。 低劑量與高劑量牛磺酸的處理,肝臟蛋白質平均含量分別為 179.7 ± 6.0 mg/g tissue 及 192.7 ± 10.4 mg/g tissue,與 CCl4 組比較,可以增加肝臟 蛋白質含量(P <0.05),Fig.3 (A)。 CCl4 誘發大鼠慢性肝損傷,hydroxyproline 平均含量為 1048 ± 95.1 µg/g tissue,其明顯高於正常對照組的 590.5 ± 38.4 µg/g tissue (P <0.001)。 低劑量與高劑量牛磺酸的處理,hydroxyproline 的含量為 758.3 ± 55.1 µg/g tissue 及 747.6 ± 81.7 µg/g tissue,與 CCl4 組比較,均明顯下降(P <0.01 ), Fig.3 (B)。. 四 、 對 肝 臟 脂 質 過 氧 化 程 度 及 glutathione 含 量 的 影 響 CCl4 誘發大鼠慢性肝損傷,其肝臟脂質過氧化程度為 3.19 ± 0.44 nmol/mg protein,明顯高於正常對照組的 1.67 ± 0.07 nmol/mg protein (P <0.05),低劑量與高劑量牛磺酸的處理,其肝臟脂質過氧化程度分別為 3.42 ± 0.45 nmol/mg protein 及 2.17 ± 0.25 nmol/mg protein,與 CCl4 組比較, 僅高劑量組具差異性(P <0.05),Fig.4 (A)。 CCl4 誘發大鼠慢性肝損傷,其 glutathione 含量為 5.43 ± 0.09 µmol/g tissue 高於正常對照組的 4.88 ± 0.17 µmol/g tissue (P <0.05),低劑量與高劑 量牛磺酸的處理,其 glutathione 含量為 5.33 ± 0.21 µmol/g tissue 及 4.66 ± 0.20 µmol/g tissue,與 CCl4 組比較,低劑量組不具統計差異。高劑量組 明顯低於 CCl4 組(P <0.05),Fig.4 (B)。. 五 、 對 肝 臟 SOD、 catalase、 glutathione peroxidase (GSH-Px)活性影響 CCl4 誘發大鼠慢性肝損傷,其肝臟 SOD 活性為 1.84 ± 0.18 U/mg. 39.
(40) protein,明顯低於正常對照組的 3.15 ± 0.20 U/mg protein (P <0.001), 低劑量與高劑量牛磺酸的處理,其肝臟 SOD 分別為 1.51 ± 0.10 U/mg protein 及 1.42 ± 0.14 U/mg protein,與 CCl4 組比較沒差異。CCl4 誘發 大鼠慢性肝損傷,其肝臟 catalase 活性為 21.9 ± 1.4 U/mg protein,明顯 低於正常對照組的 25.9 ± 1.2 U/mg protein (P <0.001),低劑量與高劑量 牛磺酸的處理,其肝臟 catalase 活性分別為 22.9 ± 0.9 U/mg protein 及 20.2 ± 1.2 U/mg protein,與 CCl4 組比較,均不具差異性。CCl4 誘發大 鼠慢性肝損傷,其肝臟 GSH-Px 活性為 719.0 ± 34.6 U/mg protein,明 顯低於正常對照組的 953.7 ± 32.1 U/mg protein (P <0.001),低劑量與高 劑量牛磺酸的處理,其其肝臟 GSH-Px 分別為 657.7 ± 30.0 U/mg protein 及 599.9 ± 53.8 U/mg protein,與 CCl4 組比較,明顯下降(P <0.05) 如 Table 2。. 六 、 反 轉 錄 聚 合 ? 鍊 鎖 反 應 (RT-PCR) CCl4 誘 發 大 鼠 肝 臟 纖 維 化 , RNA 分 析 結 果 發 現 TGF-beta1/ GAPDH 的平均值為 0.37,而正常組表現微弱,偵測不到,低劑量與高 劑量牛磺酸的處理,其平均值為 0.23 及 0.21,與 CCl4 組比較,有減 少情形,Fig.5 (A)。 CCl4 誘發大鼠肝臟纖維化,RNA 分析結果發現 HGF/GAPDH 的 平均值為 0.43,明顯低於正常對照組的 0.82,低劑量與高劑量牛磺酸 的處理,其平均值為 0.35 及 0.89。高劑量牛磺酸的處理,有增加情形, Fig.5 (B)。 CCl4 誘發大鼠肝臟纖維化,RNA 分析結果發現 TIMP-1/GAPDH 的平均值為 0.96,高於正常對照組的 0.72,低劑量與高劑量牛磺酸的 處理,其平均值為 0.76 及 0.78,與 CCl4 組比較,有減少情形,Fig.5 (C)。. 40.
(41) CCl4 誘發大鼠肝臟纖維化,RNA 分析結果 Procollagen I/GAPDH 平均值為 1.1,高於正常對照組的 0.70,低劑量與高劑量牛磺酸的處 理,其平均值為 0.97 及 0.87,與 CCl4 組比較,有減少情形,Fig.5 (D)。 CCl4 誘發大鼠肝臟纖維化,RNA 分析結果發現 MAT1A/GAPDH 的平均值為 0.74,明顯低於正常對照組的 0.94,低劑量與高劑量牛磺 酸的處理,其平均值為 0.56 及 1.24,與 CCl4 組比較,低劑量牛磺酸 的處理,有減少情形。高劑量牛磺酸的處理,有增加情形,Fig.5 (E)。 CCl4 誘發大鼠肝臟纖維化,RNA 分析結果發現 MAT2A/GAPDH 的平均值為 0.28,而正常組偵測不到,低劑量與高劑量牛磺酸的處理, 其平均值為 0.61 及 0.15,與 CCl4 組比較,低劑量有增加傾向,高劑量 牛磺酸的處理,有減少傾向,Fig.5 (F)。. 七 、 西 方 點 墨 法 ( western blot ) CCl4 誘發大鼠肝臟纖維化,蛋白質分析結果發現 TGF-beta1 的平 均值為 59.2,而正常組表現的平均值為 40.2,低劑量與高劑量牛磺酸 的處理,其平均值為 52.8 及 44.6,與 CCl4 組比較,有減少傾向 (P <0.05),Fig.6 (A)。. 八、病理切片 肝臟病理切片的觀察顯示,投予四氯化碳八週之大鼠,肝臟纖維化 和發炎程度很明顯;出現膠原蛋白纖維素,充滿發炎細胞、腫脹、玻璃 樣變性(Fig.7)。投予牛磺酸後,肝纖維化和發炎的情形有改善。. 九、基因微陣列 本實驗樣本分成四氯化碳對正常組及四氯化碳對四氯化碳+牛磺酸. 41.
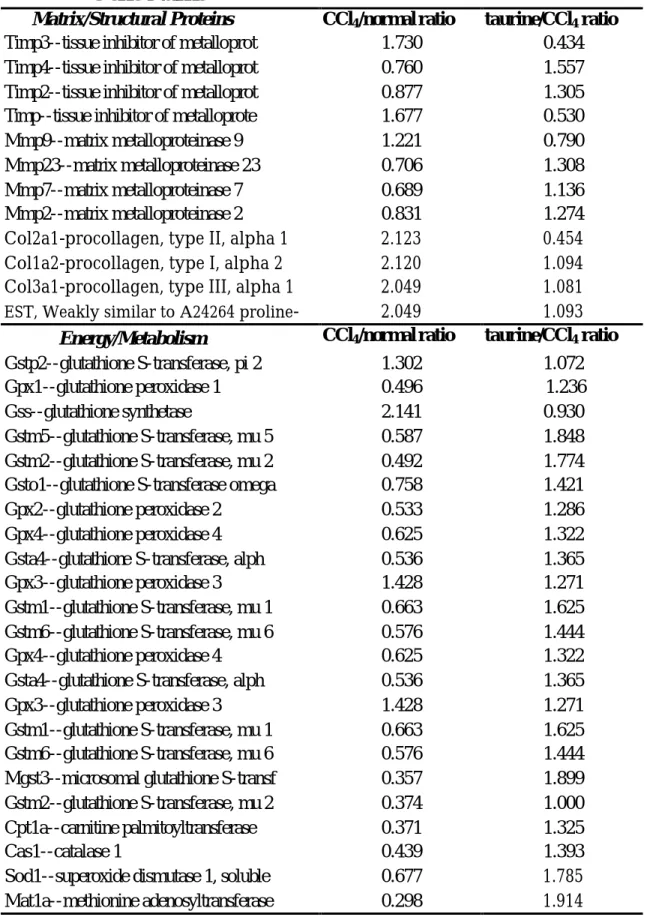
(42) 水溶液組,兩對晶片進行比對,由百恩諾公司資訊部門判讀及進行篩選 後,得到之數據中篩選出特定基因,其訊號大於 3000,即視為此基因 有大量表現,其結果列於表中 (Table 3)。其中與 matrix/structural protein 有關的基因,包括有 TIMP、MMP 及 procollagen,與 energy /metabolism 有關的基因,包括有 glutathione S-transferase、 microsomal glutathione S-transferase、glutathione peroxidase、catalase、superoxide dismutase 及 methionine adenosyltransferase。. 42.
(43) 第四章 討論. 在本次實驗中,大鼠以四氯化碳投予八週並且分別給與高劑量與低 劑量的牛磺酸水溶液,結果顯示高劑量的牛磺酸可以有效的抑制肝臟纖 維化。 GOT、GPT 酵素大量存在肝細胞內,肝臟受到損害時,GOT、GPT 酵素會釋出至血液中,因此,血清中的 GOT、GPT 值最常作為肝損傷 的指標(Sturgill and Lambert, 1997)。其中 GPT 較具專一性,因 GOT 除 了肝臟含有外,心臟、腎臟、骨骼肌、腦部也含有(Sturgill and Lambert, 1997)。大鼠經過八週四氯化碳的處理其血清 GOT、GPT 皆明顯高於正 常組,顯示四氯化碳的確可以誘導慢性肝損傷。牛磺酸的投予在第三及 第六週沒有使血清 GOT、GPT 下降的作用;然在最後第八週出現明顯 的 GOT、GPT 下降作用。此結果有可能是牛磺酸不是保護早期因四氯 化碳代謝活化引起的肝損傷,而是作用於細胞損傷的後期(Waterfield et al., 1993)。 肝硬化時,血流進入肝臟受阻,引起門脈高壓,影響到脾臟的血流, 會使脾臟腫大、門脈高壓及引起腹水(Gill and Kircbain, 1997)。本實驗 中,四氯化碳誘發慢性肝炎,最後引起脾腫大。牛磺酸具改善脾臟腫大 的傾向。 慢性肝炎會引起肝臟纖維化,即結締組織增生。結締組織主要是由 膠原蛋白構成,hydroxyproline 是膠原蛋白特有的成分,測定肝臟中 hydroxyproline 的含量可以反應膠原蛋白的含量,表示肝臟纖維化的程 度(Hanauske-Abel, 1996)。在本實驗,四氯化碳誘發慢性肝炎,其肝臟 hydroxyproline 含量明顯增加,牛磺酸能減少肝臟中 hydroxyproline 含. 43.
(44) 量,表示有減緩肝纖維化的能力。 四氯化碳引起的肝纖維化,主要是自由基造成脂質過氧化而引起的 肝損傷(Camps et al., 1992)。已有文獻指出牛磺酸具抗氧化作用(John et al., 1995)。在本實驗,四氯化碳誘發慢性肝炎,造成肝臟組織的脂質過 氧化明顯增加,高劑量的牛磺酸能降低脂質過氧化程度,顯示與降低自 由基的傷害有關。 基因微陣列的結果顯示,四氯化碳誘導組的 GSH synthetase 活性 明顯下降,而生化的測定結果卻是 GSH 含量增加,結果相反的原因有 待進一步探討。牛磺酸對 GSH 的影響可以反轉四氯化碳的作用,可推 測 GSH 也參與牛磺酸的護肝作用。 在本實驗酵素活性測定及基因微陣列的結果顯示,四氯化碳誘發大 鼠肝纖維化,肝組織中清除自由基的三種酵素 SOD、catalase、GSH-Px 活性明顯下降,牛磺酸對此 SOD 及 catalase 兩種酵素的活性沒有影 響,但會使 GSH-Px 的活性下降,所以牛磺酸抗纖維化是否與 GSH-Px 活性下降有關,有待進一步研究。雖有文獻指出牛磺酸減少 thioacetamide 引起的慢性肝損傷,可能與 GSH-Px 酵素活性的增加有關(Balkan et al., 2002)。但也有報告顯示牛磺酸能減少乙醇及 thioacetamide 造成的慢性 肝損傷,與 SOD、GSH-Px 及 GSH-ST 酵素活性無關(Balkan et al., 2001;Dogru-Abbasoglu et al., 2001)。 先前實驗大鼠的研究顯示牛磺酸有改善肝纖維化的作用,主要可以 明顯抑制四氯化碳所提升的 Procollagen-1 及 TIMP-1 基因表現(Chen, 1999; 陳,1991)。在我們的研究模式中,也獲得相同的結果,證實牛磺酸 明顯抑制四氯化碳所提升的 Procollagen-1 及 TIMP-1 基因表現(Fig.5 (C)(D))。 TGF-beta1 是目前發現的最重要的致纖維化細胞因子(Shanthi et al.,. 44.
(45) 1998)。 TGF-beta1 的表現增加,可促進 TIMP-1 的表現,進而增強 Procollagen-1 的表現(Eng, 1998)。因此抑制 TGF-beta1 的表現,是目前 抗肝纖維化藥物開發的重點。本實驗以 mRNA 及蛋白質的表現確認牛 磺酸可以抑制四氯化碳所提升的 TGF-beta1 表現。可以證實牛磺酸具抗 肝纖維化作用(Fig.5 (A))。 HGF 是一種具有多種生物學作用的蛋白質分子,實驗證明,在肝 切除和各種肝病血液中 HGF 含量會明顯升高, 肝組織中的 HGF mRNA 亦會表現,其目的是在促進肝臟再生及修復 (Lindroos et al., 1991)。雖然如此,Masson 氏等人的四氯化碳肝纖維化實驗顯示 HGF mRNA 表現不受影響(Masson et al., 1999)。本實驗結果發現在八週四氯 化碳的誘導下,HGF mRNA 的表現明顯下降,高劑量的牛磺酸可以消 除 HGF mRNA 的下降表現。然而此四氯化碳組的結果與 Masson 氏等 人(1999) 的結果不同,其 HGF mRN 在四氯化碳誘導下,與正常組無差 異性,我們認為是四氯化碳損傷到肝細胞的再生功能,真正原因有待進 一步研究。 肝損傷時 MAT 的活性降低(Mato et al., 1997),調控 MAT 的基因是 mat1a 的表現也會下降 (Avila et al., 2002)。MAT1A 的表現與 SAM 生成 有 關, SAM 與 polyamine 及 glutathione 的生合成有關 (Chen et al., 2000),此二者對肝細胞有很重要的保護作用(Chen et al., 2000)。本實驗結 果顯示四氯化碳的誘導下,MAT1A 的表現明顯下降,此與其他研究一致 (Avila et al , 2002),高劑量牛磺酸能明顯提升 MAT1A 的表現,顯示牛 磺酸確實具有保肝的效果。 雖然 MAT2A 在一般肝細胞不會表現,但 MAT1A 及 MAT2A 兩者在肝臟的存在是互相調控的,當肝損傷或肝細胞增生時 MAT2A 會 開始表現,取代 MAT1A,雖然此機轉不明,但仍待研究(Shelly et al.,. 45.
(46) 2001)。本實驗也發現正常肝組織 MAT2A 沒有表現,四氯化碳誘發的 肝損傷會造成 MAT2A 表現,而 MAT1A 的表現明顯下降。高劑量的 牛磺酸能抑制 MAT2A 表現,更進一步支持牛磺酸具有保護肝細胞避免 肝損傷的功能。 牛磺酸對四氯化碳造成的肝纖維化有明顯減輕現象,其作用機轉可 能經由抗氧化(如: GSH-ST)和肝細胞保護作用(如: MAT1A),降低肝細 胞損傷 (MAT2A 表現下降),進而減弱 TGF-beta1 的表現,因此減少 TGF-beta1 所誘發肝纖維化的基因表現(如: TIMP-1、Procollagen-1),最 後減少細胞外基質的存積,達到改善肝纖維化的效果。. 46.
(47) 結 論 在 四 氯 化 碳 誘 導 大 鼠 慢 性 肝 損 傷 模 式 下 , 由 RT-PCR 及 expressed microarray analysis 結果得知適量的牛磺酸可以降低 TGFbeta1、TIMP 及 Procollagen-I mRNA 的表現,提升 HGF、MAT1A 及 GSH-ST mRNA 的表現,因此牛磺酸可能是透過 GSH-ST 催化 GSH 與四氯化碳代謝物結合,並且由 MAT 促進 SAM 的生成來穩定細胞膜 脂質及調節細胞膜通透性,使牛磺酸能保護肝細胞避免受到傷害。. 47.
(48) 參考文獻 王紅,等.(1993).牛磺酸對半乳糖胺及硫酸亞鐵-抗壞血酸所致大鼠肝細 胞損傷的保護作用.中國藥理學與毒理學雜誌.7(4):282~284 冉 霞 .(1993)牛磺酸與機體的免疫功能.國外醫學 衛生分冊 6:344-346 李兆萍,等.(1990)牛磺酸對離體大鼠心臟缺血一再灌流損傷的保護作用. 北京醫科大學學報, 22 : 208 吳建民,等.(1991) Mg2+和牛磺酸對內皮素促血管平滑肌細胞增殖的影 響. 北京醫科大學學報, 23(5):351-352 研崇超,等.(1998)大鼠飼料中補充牛磺酸對血、肝膽固醇水平的影響. 衛生研究.17(6):25-28 周健偉 .(1995).牛磺酸與肝臟關係的研究進展 .國外醫學衛生分冊, 22(2):97~100 黃 金 保 .(1991)牛磺酸對心血管的藥理作用.中國藥理學通報. 7(1):8~11 陳長勛,等.(1991)牛磺酸對血小板聚集含實驗性血栓形成的影響.中國 藥學雜誌.26(4):207-209 陳岳祥,等(1999) 牛磺酸對四氯化碳誘導大鼠肝纖維化的抑制作用.中 華消化雜誌.19(3)185-187 劉慶德,等.(1991).牛磺酸對缺血再灌流損傷大鼠心肌鈣超載的影響.北 京醫科大學學報. 23(5):348 韓 曉 濱 .(1988). 牛 磺 酸 與 大 腦 發 育 關 係 的 初 步 探 討 . 衛 生 研 究 . 17(3):22~26 韓曉濱.(1989).牛磺酸與大腦發育關係的研究.營養學報. 11(4):319~324 韓曉濱.(1992).牛磺酸與人腦神經細胞分化影響的研究.生理學科學進 展. 23(4):339~341 劉慶德,等.(1991)牛磺酸的生物學效應.國外醫學生理病理學與臨床分 冊. 11(1):3 劉 錫 光 ,等 .病毒性肝炎實驗診斷學(第二版).人民衛生出版社 韓曉濱.(1991)牛磺酸與中樞神經系統關係的研究新進展.國外醫學 衛 生學分冊.3:143-146. 48.
(49) Aebi H. (1984) Catalase in vitro. Methods Enzymol. 105:121-126. Alvarez L, Corrales F, Martin-Duce A, Mato JM. (1993) Characterization of a full-length cDNA encoding human liver S-adenosylmethionine synthetase: tissue-specific gene expression and mRNA levels in hepatopathies. Biochem J. 293 ( Pt 2):481-486. Andreu H, Pares A, Mato JM, Galli-Kienie M. (1979) S-adenosyl-lmethionine tretment prevents carbon tetrachloride-induced S-adenosyl-lmethionine synthetase inactivation and attenuates liver injury. Hepatolgy 16:1022-1027 Anscher M, Peters W, Reisenbichler H, Petros W, Jirtle R. (1993) Transforming growth factor beta as a predictor of liver and lung fibrosis after autologous bone marrow transplatation for advanced breast cancer. New England of Medicine. 328:1592-90-95 Akerboom TP, Narayanaswami V, Kunst M, Sies H. (1991) ATP-dependent S-(2,4-dinitrophenyl) glutathione transport in canalicular plasma membrane vesicles from rat liver. J Biol Chem.266(20):13147-52 Asami O, Ihara I, Shimidzu N, Shimizu S, Tomita Y, Ichihara A, Nakamura T. (1991) Purification and characterization of hepatocyte growth factor from injured liver of carbon tetrachloride-treated rats. J Biochem (Tokyo). 109(1):8-13. Azuma J, Hamaguchi T, Ohta H, Takihara K, Awata N, Sawamura A, Harada H, Tanaka Y, Kishimoto S. (1987) Calcium overload induced myocardial damage caused by isoproterenol and adramycin : possible role of taurine in its prevention. Adv Exp Med Biol. 217:167-79. Balkan J, Dogru-Abbasoglu S, Kanbagli O, Cevikbas U, Aykac-Toker G, Uysal M. (2001) Taurine has a protective effect againstthioacetamide induced liver cirrhosis by decreasing oxidative stress. Hum Exp Toxicol. 20(5):251-4. Balkan J, Kanbagli O, Aykac-Toker G, Uysal M. (2002) Taurine treatment reduces hepatic lipids and oxidative stress in chronically ethanol-treated rats. Biol Pharm Bull. 25(9):1231-3. Banks MA, Porter DW, Pailes WH, Schwegler-Berry D, Martin WG, Castranova V (1991) Taurine content of isolated rat alveolar type I cells. Comp Biochem Physiol B. 100(4):795-9. Bea F, Hudson FN, Chait A, Kavanagh TJ, Rosenfeld ME. (2003) Induction of glutathione synthesis in macrophages by oxidized low density lipoproteins is mediated by consensus antioxidant response elements. Circ Res. 92(4):386-93. Benyon RC, Arthur MJ. (1998) Mechanisms of hepatic fibrosis. J Pediatr Gastroenterol Nutr.27(1):75-85. Benyon RC, Iredale JP, Goddard S, Winwood PJ, Arthur MJ. (1996). 49.
(50) Expression of tissue inhibitor of metalloproteinases 1 and 2 is increased in fibrotic human liver. Gastroenterology. 110(3):821-31. Biempica L, Zern MA. (1989) In vitro and in vivo association of transforming growth factor-beta 1 with hepatic fibrosis. J Cell Biol. 108(6):2477-82. Cai J, Sun WM, Hwang JJ, Stain SC, Lu SC.(1996) Changes in S-adenosylmethionine synthetase in human liver cancer: molecular characterization and significance. Hepatology. 24(5):1090-7. Cameron GR, Karunaratne WAE. (1936) Carbon tetrachloride cirrhosis in relation to liver regeneration. J.Pathol.Bacterol. 42:1-21 Camps J, Bargallo T, Gimenez A, Alie S, Caballeria J, Pares A, Joven J, Masana L, Rodes J. (1992) Relationship between hepatic lipid peroxidation and fibrogenesis in carbon tetrachloride-treated rats: effect of zinc administration. Clin Sci (Lond).83:695-700 Cantoni, G. L. (1953) J.Biol.Chem 204,403-416 Cabrero C, Suma Duce, AM, Oritz P. Alemany S, Mato JM. (1988) Specfic loss of the high-molecular-weight form of S-adenosyl-l methionine synthetase in human liver cirrhosis.Hepatology 8:1530-1534 Castilla A, Prieto J, Fausto M. (1991) Transforming growth factor 1 and alpha in chronic liver diease. Effets of interferon alpha therpy. New England Journal of Medcine. 324:933-940. Chen W, Kennedy DO, Kojima A, Matsui-Yuasa I. (2000) Polyamines and thiols in the cytoprotective effect of L-cysteine and L-methionine on carbon tetrachloride-induced hepatotoxicity. Amino Acids.18(4):319-27. Chen Y, Li S, Zhang X. (1999) Taurine inhibits deposition of extracellular matrix in experimental liver fibrosis in rats. Zhonghua Gan Zang Bing Za Zhi. 7(3):165-7. Chesney RW. (1985) Taurine: its biological role and clinical implications. Adv Pediatr. 32:1-42. Chomczynski P, Sacchi N. (1987)Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Analytical Biochemistry.162(1):156-159. Corrales F, Gimenez A, Alvarez L, Caballeria J, Pajares MA, Stramentinoli G, Pezzoli C, Galli-Kienle M .(1979) Protective role of S-adenosyl-l-methionine against acetaminophen induced mortality and hepatotoxity in mice. Biochem.Pharmacol. 28:3567-33571 Corrales F, Ochoa P, Rivas C, Martin-Lomas M, Mato JM. Pajares M A. (1991) Inhibition of glutathione synthesis in the liver leads to S-adenosyl-methionine synthetase reduction. Hepatolgy 14:528-533 Czaja MJ, Weiner FR, Flanders KC, Giambrone MA, Wind R, Chen YM, Wu KD, Tsai TJ, Hsieh BS. (1999) Pentoxifylline inhibits. 50.
(51) PDGF-induced proliferation of and TGF-beta-stimulated collagen synthesis by vascular smooth muscle cells. J Mol Cell Cardiol. 31(4):773-83. Dogru-Abbasoglu S, Kanbagli O, Balkan J, Cevikbas U, Aykac-Toker G, Uysal M. (2001) The protective effect of taurine against thioacetamide epatotoxicity of rats. Hum Exp Toxicol. Jan;20(1):23-7. Duffy MC, Blitzer BL, Boyer JL. (1983) Direct determination of the driving forces for taurocholate uptake into rat liver plasma membrane vesicles. J Clin Invest. 72(4):1470-81. Eng Looi. (1998) Salvia miltiorrhiza reduces experimentally induced hepatic fibrosis in rats. Jounal of hepatology 9(5):760-771 Friedman SL. (1996) Hepatic stellate cells. Prog Liver Dis. 14:101-30. Friedman SL, Roll FJ, Boyles J, Arenson DM, Bissell DM. (1989) Maintenance of differentiated phenotype of cultured rat hepatic lipocytes by basement membrane matrix. J Biol Chem . 264(18):10756-62. Gebhardt R. (2002) Prevention of taurolithocholate-induced hepatic bile canalicular distortions by HPLC-characterized extracts of artichoke (Cynara scolymus) leaves. Planta Med. 68(9):776-9. Geggel H S (1985) Nutritional requirement for taurine in patients receiving long-term parenternal nurtrion. N Engl J Med 312:142A Gill MA, Kircbain WR. (1997) Alcoholic liver disease. In:Dipiro JT, Taibert RL,Yee GC,Matzke GR,Wells BG and Posey LM. Editors , Pharmacotherapy:A Pathophysiological Approach. Stamford : Appleton &Lange.pp785-800. Gong H, Wu X, Shu Y. (1998) A novel gene delivery system targeting to epidermal growth factor receptor overexpressing cancer cells Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi. 12(1):43-6. Hafeman DG, Sunde RA, Hoekstra WG.(1974). Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. Journal of Nurtrition. 104(5):580-587. Hanauske-Abel HM. (1996) Fibrosis: representive molecular elements, a basic concept, and emerging targets for suppressive treatment. In: Hepatology: A textbook of liver disease. Hanauske-Abel HM.(1996) Fibrosis: representive molecular elements, a basic concept, and emerging targets for suppressive treatment. In: Hepatology: A textbook of liver disease. Zakim, D. and Boyer, T.D.(Eds) W.B. Saunders Company, Philadelphia,third edition,pp465-506 Hayes KC, Carey RE, Schmidt SY. (1975) Retinal degerenertion associated with taurine deficiency in the cat. Science 188:949. 51.
數據
Outline
相關文件
Normalization by the number of reads in the sample, or by calculating a Z score, should be performed on the reported read counts before comparisons among samples. For genes with
at each point of estimation, form a linear combination of a preliminary esti- mator evaluated at nearby points with the coefficients specified so that the asymptotic bias
A Complete Example with equal sample size The analysis of variance indicates whether pop- ulation means are different by comparing the variability among sample means with
The nanostructure with anisotropic transmission characteristics on ITO films induced by fs laser can be used for the alignment layer , polarizer and conducting layer in LCD cell.
• QCSE and band-bending are induced by polarization field in C-plane InGaN/GaN and create triangular energy barrier in active region, which favors electron overflow. •
Optim. Humes, The symmetric eigenvalue complementarity problem, Math. Rohn, An algorithm for solving the absolute value equation, Eletron. Seeger and Torki, On eigenvalues induced by
Moreover, for the merit functions induced by them for the second-order cone complementarity problem (SOCCP), we provide a condition for each stationary point being a solution of
Moreover, for the merit functions induced by them for the second- order cone complementarity problem (SOCCP), we provide a condition for each sta- tionary point to be a solution of
