行政院國家科學委員會專題研究計畫 成果報告
以基因剔除技術研究 cullin-4B 的功能 III
計畫類別: 個別型計畫 計畫編號: NSC93-3112-B-002-039- 執行期間: 93 年 05 月 01 日至 94 年 04 月 30 日 執行單位: 國立臺灣大學醫學院醫事技術學系暨研究所 計畫主持人: 林淑華 報告類型: 完整報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 94 年 7 月 22 日
I 中文摘要 Cullin 蛋白藉由組合成不同的泛素連接酶,以調節細胞內各種重要的蛋白質,而此 一調節是經由泛素-蛋白降解體路徑來完成。迄今 cullin 家族已有七個基因被發現,而我 們的主要研究對象- Cul4B 是否參與調控與發育相關的蛋白仍待釐清。Cul4B 在許多種 組織都有表現,包括腦部、睪丸、卵巢、攝護腺、結腸、脾臟、肝臟及周邊血球細胞甚 至整個胚胎都可偵測到。為了解 Cul4B 的功能,我們同時藉由建造動物模型及細胞培養 的方式來回答此一問題。首先希望建立組織特異性 Cul4B 基因剔除小鼠而能在不同的器 官研究此蛋白質在活體內的功能,我們已建構好 Cul4B 條件式基因剔除載體,並用以轉 染 107小鼠胚胎幹細胞。由抗生素篩選得到的 264 個細胞株中,經由南方墨點法的分析, 共得到 8 個標的成功的細胞株。取出囊胚期卵子,注射幹細胞後,植入代孕母鼠,共產 出 5 隻嵌合鼠。嵌合鼠與黑色 B6 母鼠互配後,得到了栗色毛的子代,證明其遺傳來源 為胚胎幹細胞,且為 Cul4B 基因標定的異型合子。由此一異型合子交配而得到同型合子 後,再與特定在肝或生殖細胞表現 Cre 的轉殖鼠互配,便可得到組織特異性 Cul4B 基因 剔除鼠,以作為進一步研究 Cul4B 功能的材料。 同時,我們也應用生化學的方法來研究 Cul4B 的功能。首先,建立會表現 myc 標 記的 Cul4B 蛋白之細胞株,並以共同免疫沈澱的方式將細胞內與 Cul4B 有交互作用的 蛋白沈澱下來;經由電泳分離後,將特定位置的蛋白切下,以質譜分析鑑別。經由此一 程序,發現 TIP120A 及 DDB1 與 Cul4B 有交互作用;其中,TIP120A 已知會以負調控 的方式調節多種泛素連接酶,而先前的研究則發現 DDB1 是 Cul4A 的轉接蛋白,並且 參與細胞週期的調節。這暗示 Cul4B 在正常生理狀況下可能會以相似於 Cul4A 的作用 方式進行蛋白質降解之調節。 本實驗室未來的目標是分析 Cul4B 基因剔除鼠的表現特徵,以及繼續用共同免疫沈 澱法找尋與 Cul4B 有交互作用的蛋白,以建立由 Cul4B 組成的泛素連接酶調控體內蛋 白表現的模式。
II 英文摘要
Cullin proteins seem to couple to different E3 ubiquitin ligases and regulate the level of key proteins in many cellular processes by degrading proteins through the
ubiquitin-proteasome pathway. Whether CUL-4B, one of the 7 cullin genes identified to date, is involved in degradation of developmentally important proteins remains to be determined. It is highly expressed in various tissues including whole embryo. We have been investigating the function of CUL-4B by tissue-specific gene targeting in mouse embryonic stem cells. We have constructed a targeting vector with CUL-4B gene flanked by three lox P sites, and
transformed107 ES cells with the construct. Of 264 clones isolated with antibiotics selection and southern blot analysis, 8 candidate clones were obtained. We have injected the selected clones into blastocysts and implanted them into foster mice. Five chimeric mice are generated. Germ line offsprings are detected by the presence of agouti litters. We are now breeding the loxP-floxed CUL-4B mice. By mating the loxP-floxed CUL-4B mice with mouse strains that express Cre in liver or germ cells, we will generate mouse strain with deletion of CUL-4B in these two organs. The mice will allow us to investigate the importance of CUL-4B.
We have also taken biochemical approaches to dissecting CUL-4B function. Cell lines stably expressing Myc-tagged CUL-4B were generated. Analyzing the co-immunoprecipitated CUL-4B interacting molecules by MALDI-TOF MS/MS showed that CUL-4B interacts with TIP120A and UV-damaged DNA –binding protein , DDB1. TIP120A functions as a negative regulator of different E3 ubiquitin ligases and DDB1 was reported to be an adaptor protein to mediate substrate binding to CUL-4A associated E3 ubiquitin ligases in regulating cell cycle. It may recruit substrates for the CUL-4B dependent ligases in a similar fashion.
Our future goal is to study CUL-4B function by analyzing the phenotypes of the CUL-4B knockout mice. And we will analyze more CUL-4B interacting molecules pulled down by co-immunoprecipitation and identified by 2D- proteomics to find out the targeting molecules of CUL-4B and the components of SCF-like complex formed by CUL-4B.
1 前言
Targeted protein degradation by the ubiquitin-proteasome pathway plays a vital role in monitoring the abundance of many short-lived regulatory proteins. Through the action of a multienzyme system consisting of the E1 ubiquitin activating enzyme, the E2 ubiquitin conjugating enzyme, and the E3 ubiquitin ligase, multiple ubiquitin moieties are delivered to the target protein to form a polyubiquitin chain and then the target protein was recognized and degraded by the 26S proteasome. The SCF subclass of E3s is a group of multimeric protein complexes that are assembled around a core module composed of Skp1, a cullin family member, the Rbx1 (Roc1/Hrt1) protein and a F-box protein. Each cullin protein seems to form different E3 complex and plays a role in regulating the level of key protein in many cellular processes. Human Cul-4A and Cul-4B are orthologs of nematode Cul-4. Cul-1, Cul-3, Cul-4A, and Cul-7 knockout (KO) mice are embryonic lethal. Cul 4B is a novel member of this family and its function is unknown. Whether KO of Cul-4B is embryonic lethal awaits determination so as its involvement in cell cycles. A conditional tissue-specific knockout is an appropriate strategy to study the function of Cul 4B. We have designed in-vitro and in-vivo experiments to dissect the
physiological functions of Cul4B.
研究目的 The specific aims of this proposal are as follows:
(1) Addressing developmental role of Cul-4B by generating conditional and tissue-specific Cul 4B knockout mice using Cre-LoxP system.
(2) Dissecting whether Cul 4B is essential for ES cell survival by gene targeting in mouse embryonic stem cell.
(3) To identify Cul-4B interacting molecules by Co-immunoprecipitation, 2D gel electrophoresis coupled with Q-TOF using Cul-4B over-expression strategy.
文獻探討 Cullin 4B
Cullin 4B (Cul 4B) is a member of the cullin gene family as defined by sequence homology (1). The full-length cDNA of human Cul 4B have been isolated, and the sequence analysis indicated that the Cul 4B is expressed as a 716-amino acid protein (2). Human cullin 4A(Cul 4A) and Cul 4B are orthologs of nematode Cul 4. The partial C-terminal amino acid sequences of Cul 4A and Cul 4B share 88% identity (1). Cul 4B is expressed in a wide variety of tissues including whole embryo. Recently, the gene coding for human Cul 4B was identified to locate on X chromosome Xq23 by sequence analysis in GenBank and shown that it spans over 36 Kb and contains 20 exons (3).
2
homology in a region of approximately 200 amino acid residues, designated the cullin homology domain (CH) (4, 5). Cullins interact with the RING-H2 domain protein
Rbx1/Roc1/Hrt1 through their CH domains to form core ubiquitin ligase (E3) modules as SCF complex. Among members of the cullin family, Cul 1, Cul 2 and Cul 3 have been demonstrated to mediate the selective degradation of regulators of cell cycle and signaling pathways (see section “cullins”). Recently, the associated proteins and the proteolytic substrates of Cul 4A and Cul 5 were demonstrated. However, little is known about the Cul 4B.
Cullins
Cul1 was originally identified in nematodes in a screen for genes inhibiting hyperplasia (1) and is homologous to Cdc53 (also called CulA) in yeast. A family of four additional
Caenorhabditis elegans and six human genes were identified in databases by searching with
the Cul1 sequence. Cul1 is the best characterized member of the family and is the only one that interacts with Skp1 (6). The name cullin comes from the verb `to cul' in the sense of `to sort'. Cul1 has three domains that mediate its association with other components of the SCF
(reviewed in ref 7). The least conserved domain among cullin members is the N-terminal region which in Cul1 mediates its binding to Skp1. The second and most highly conserved domain is present in the extreme C-terminus of all cullins and mediates the attachment of a small ubiquitin-like protein, Nedd8 (Rub1 in yeast). Covalent modification of cullins by the Nedd8 pathway appears to enhance the ubiquitin-ligating activity of some SCF ligases (reviewed in ref 8). A third domain downstream to this Skp1-binding region is called the cullin homology (CH) region or Cdc34/RING-H2 subunit-recruiting domain. This domain is
conserved among all cullins and binds to Rbx1/Roc1 (Hrt1 in yeast) (ref 9 and reviewed in ref 10, 11). Other cullins also bind to Rbx1 and form different ligases with different specificity. Cul-2 associates with a complex of elongin C (a protein similar to Skp1), and elongin B (an ubiquitin-related protein), and either the VHL (Von Hippel Lindau) tumor suppressor protein or a Socs (suppressor of cytokine signaling)-box-containing protein. Together, this complex is referred to as the VBC-like ligase, in which VHL and probably Socs proteins carry out the role of the substrate-targeting subunit performed by F-box proteins in SCF complexes (reviewed in ref 10, 11). It is likely that other cullins form similar complexes and that the SCF pathway is a prototype system for a larger class of multi-component cullin-based ubiquitin ligases.
Interestingly, APC/C contains a cullin subunit (Apc2) and RING finger protein (Apc11) that targets a specific set of substrates.
Targeted deletion of the Cul1 gene in C. elegans and in mice has provided useful genetic tools for the analysis of the function of this gene. In C. elegans, the loss of Cul1 results in increased proliferation and hyperplasia in all larval tissues examined (1). This suggests that Cul1 is an essential negative regulator of cell cycle. Additionally, Cul1 is required for
developmentally regulated cell cycle exit (G1 to G0 transition), however the loss of function of Cul1 in quiescent cells does not have an effect on cell cycle re-entry (G0 to G1 transition). Mice carrying a deletion in Cul1 die around embryonic day 6.5 and cyclin E protein levels, but
3
not mRNA are increased in these cells (as determined by immunohistochemistry and in situ hybridization, respectively) (12, 13). The mechanism of developmental arrest in Cul1 -/-embryos is unknown. The accumulation of cyclin E is not likely to be the cause, since cells have a tolerance for high levels of this cyclin (14, 15). It is likely that the emerging role of Cul1 in SCFs is as a core component that supports the targeting of multiple substrates, which probably accumulate in these Cul1-/- cells. Very recently, it has been reported that Cul1 is a direct transcriptional target of Myc and that enforced expression of Cul1 or antisense p27 oligonucleotides is capable of overcoming the slow growth phenotype of Myc-deficient mouse embryonic fibroblasts (16). This indicates that p27 is a major obstacle to cell cycle progression in the absence of Cul1.
In addition to Cul1, Cul2 and Cul3 also have been demonstrated to mediate the selective degradation of regulators of cell cycle and signaling pathways. Cul2 associates with elongin C, elongin B, VHL protein and Rbx1 proteins to form VCBCR complex. The VCBCR complex binds HIF-1alpha, transcription factors critically involved in cellular responses to hypoxia, and targets it for ubiquitin-mediated proteolysis (17). Okuda et al. also showed that a member of the atypical protein kinase C group, PKClambda, is ubiquitinated by the VCBCR complex (18). Cul3 may be a core component of an E3 ubiquitin ligase complex that targets free cyclin E for destruction. Genetic evidence from cul3-/- mice (19) is also consistent with the specific involvement of Cul3 in the ubiquitination of cyclin E. Mice homozygous for the cul3 deletion die at or before day 7.5, with disorganized (or in some cases absent) extraembryonic tissues, abnormal development of the trophectoderm, and a failure to undergo proper gastrulation. Importantly, cyclin E levels in cul3-/- embryos are elevated in the ectoplacental cone and the extraembryonic ectoderm (but not embryonic ectoderm) relative to wild-type mice, and this increase correlated with a dramatic increase in the number of S-phase cells as determined by BrdU incorporation (19). Thus, Cul3 is likely to be directly required for destruction of cyclin E in specific tissues in the developing mouse embryo. However, it seems unlikely that the developmental defects observed are solely due to cyclin E accumulation, as Cul3 complexes will likely regulate the levels of other proteins in addition to cyclin E.
Recently, the association proteins and proteolysis substrates of Cul4A and Cul5 were identified. Cul4A can physically associate with the damaged DNA binding protein (DDB), which is composed of two subunits, p125 and p48. DDB binds specifically to UV-damaged DNA and is believed to play a role in DNA repair. Chen et al. reported that Cul4A stimulates degradation of p48 through the ubiquitin-proteasome pathway, resulting in an overall decrease in UV-damaged DNA binding activity (20). Kamura et al. demonstrated that the Cul5/Rbx1 module associates with the Elongin BC complex and a novel elongin BC-box/Leucine-rich repeat-containing protein MUF1 to form a functional ubiquitin ligase (21).
The versatility of protein ubiquitination as a cellular regulatory mechanism is now well established and appears to be comparable to that of other protein modifications such as phosphorylation. Indeed, protein phosphorylation and ubiquitination go hand in hand in the
4
regulation of many cellular pathways. However, the enzymology of the ubiquitination reaction is more complex than that of protein phosphorylation, and the mechanism of polyubiquitin chain formation is unclear yet. It is hoped that the recent characterization of the APC and SCF complexes, and their reconstitution, will allow further progress on this topic to be made. We can expect in the near future is the identification of an ever-increasing number of substrates of the ubiquitin system and of their specific E2/E3 complexes. It remains to be seen whether all the new E3s will belong to one of the known classes, or whether new types of E3s will be identified. The ubiquitin system is involved in many cellular processes, some of which are linked to human pathologies, such as malignancies, hypertension, the inflammatory response and skeletal muscle breakdown (review in ref 22). Therefore, modulation of the activity of the ubiquitin system may be expected to serve in the future as a target for therapeutic intervention. Angelman’s syndrome, a human genetic disease with features of severe motor and intellectual retardation, is due to mutations in the ubiquitin ligase E6-AP (23, 24). Liddle’s syndrome, in which patients develop a severe form of hypertension, is due to a mutation in the kidney epithelial Na+ channel (ENaC) that does not allow interaction of the channel with its ubiquitin ligase, Nedd4. Stabilization of the channel leads to excess reabsorption of Na+ and H2O (25).
It is likely that, in addition to these two syndromes, other human genetic diseases will be linked to genes encoding ubiquitin system components. One intriguing possibility is the existence of slightly altered-activity variants of ubiquitin system genes. In some cases, small differences in steady-state levels of regulatory proteins can have significant effects on the organism; for example, p27Kip1 was shown to be haploinsufficient for tumor suppression, i.e., even
heterozygous mice displayed an increased incidence of cancer (26). Thus, it is conceivable that even minor alterations in the normal degradation of p27Kip1 or of other regulatory proteins by the ubiquitin system can contribute to a disease phenotype. This type of mutations in ubiquitin system genes leading to specific pathologies may be uncovered with the new mapping methods for quantitative trait loci.
In the light of other cullin members involved in the regulation of key proteins of cellular processes, it is possible that Cul 4B also play a role in regulating cell function. Whether KO of Cul-4B is embryonic lethal awaits determination, we try to create conditional and conditional tissue-specific knockout mice at the same time to study the function of Cul 4B. This study will provide an insight into the physiological roles of Cul 4B in cellular processes including cell growth and embryo development. The results of this study will expand the knowledge on the regulation of cell function through ubiquitin-mediated degradation pathway.
研究方法 Designs
To get the Cul-4B genomic DNA is the very first step. Isogenic Cul-4B genomic DNA was obtained by screening a 129/sv-mouse genomic library with a human Cul-4B cDNA. For Specific aim (1)
5
Mouse with deletion of the Cul 4B gene in specific tissues will be generated as an animal model for dissecting the functional roles of Cul 4B. The Cre/LoxP strategy derived from the
bactriophage P1 will be used to perform conditional and tissue specific knockout. We will create mouse lines with the 1.9 kp LoxP sequences integrated in introns 12 and 14 of Cul 4B gene. When the cre recombinase is introduced into these mice by CMV driven-Cre gene expression or by breeding the LoxP carrying mice with the transgenic mice expressing Cre enzyme, we will be able to delete exons 12-14 of Cul 4B in the mouse. The morphology and physical examination of the generated Cul 4B knockout mice will be performed to evaluate consequences of deletion of Cul 4B.
For specific aim (2)
When obtaining the ES cell by the methods for specific aim(1). Introduce the Myc-tagged CUL-4B constructs mentioned above [also containing IRES-GFP (internal ribosome entry site-green fluorescent protein)] into our ES clones by electroporation and study whether CUL-4B is essential to ES cell growth or not. The design of this approach is explained in detail in figure 2.
For specific aim (3)
Cell lines stably expressing CUL-4B and powerful antibodies for immunoprecipitation should obtained.. We use SK-Hep-1 cell stably expressing Myc-tagged full-length and anti-Myc antibody (home made from ATCC hybridoma clones) for co-immunoprecipitation experiments. We also express the Cul-4B cDNA in the pET15 vector in the E. coli BL21(DE3)/pLysS strain to obtain the full-length protein for immunization.
Methods
A. Construction of LoxP containing targeting vector:
The targeting vector is designed based on selection by neomycin resistance gene and HPRT gene. The selection gene (neor) is inserted into the intron 11 and flanked by 2 loxP sites after a 2.9-kb BamHI fragment containing exons 7-11 , followed by a 3.9-kb fragment containing exons 12-14 of mouse Cul
4B gene . The third loxP site is inserted into the intron 14 in the 7 kb fragment with HPRT selection
gene. The cre recombinase will direct homologous recombination of any pairs of the LoxP sites, and excision of some LoxP combinations will result in deletion of exons 12-14.
B. Culturing ES cells and gene targeting at the Cul 4B locus.
Wild type ES cell lines (from Dr. Babinet, Pasteur Institute, France) and E14 (HPRT-) ES cell lines (from Dr. Hung Li, Academia Sinica) were cultured and propagated as described (27) in DMEM media supplemented with 15% heat inactivated fetal bovine serum (FBS), 0.1 mM 2-mercaptoethanol. To maintain the ES cells in their undifferentiated state, mitomycine C treated primary fibroblasts are used as feeder cells, and the leukemia inhibitory factor (LIF) was added to the media at a final concentration of 1000 U/ml. Transfection of the ES cells with
6
the targeting construct was performed by electroporation at 220 V and 960 µF (Gene Pulser, BioRads) using a 1 ml cuvette filled with 20 µg/ml of DNA and 107
ES cells. The targeting plasmid will be linearized at one end of homology prior to its introduction into the cells. The desired ES clones will be selected with G418 (or HAT) and Gancyclovir and survival colonies were isolated and established as clones for further analysis (27).
C. Generation of conditional Cul 4B knockout mice.
The ES cells harboring the targeting construct of Fig. 1, will be identified by Southern blotting, and injected into blastocysts to generate chimeric mice. Breeding of the chimera will be carried out as described (28, 29) to obtain germline transmission. Crossing the mouse line with Cre transgenic mice will generate mice with tissue-specific KO of the Cul 4B gene. We intend to use CMV (cytomegalovirus)- or albumin-driven Cre transgenic mice provided by Dr. Frank Koentgen (Director and CEO, Ozgene, the Australian Gene Targeting Center) and Dr. Iwakura at the Univeristy of ToKyo. Mice with no Cul 4B in liver and in kidney will be investigated.
D. Using electrofusion to generate knockout mice.
Electrofusion is used to generate tetraploid embryos, which can then be used to make aggregation chimeras with ES cells. Tetraploid embryos normally develop very poorly, and in the chimeras, they give rise only to trophoblast cells in the blastocyst and placenta and to a few extraembryonic endoderm cells. The majority of the cells in the embryo, and all of the cells in the newborn mouse, are therefore derived from the donor ES cells. It means we don’t need to generate chimeric mice by injection ES cells into blastocyst, then mating chimera mice to generate KO mice. We can obtain KO mice by aggregation using electrofused embryo and ES cells. To do so, we need the eletrofusion machine (e.g. Multiporator, Eppendorf, Germany) as list in the budget for equipment. We will follow the methods described in ref 30. Briefly, collect two-cell embryos and fuse these embryos by electrofusion, then make aggregation with ES cells. The formed blastocysts are implanted into foster mice.
E. Determination of Cul 4B expression in tissues from knockout mice.
The expression of Cul 4B in the tissues will be determined by RNA and protein levels by methods established in our laboratory as described previously (29). In-situ hybridization and immunohistochemistry (30) will be performed to assess the Cul 4B expression pattern in tissues first, then quantitated by RT-PCR and Western blot analysis respectively.
F. Morphology and functional study of Cul 4B knockout mice.
The morphology of organs of Cul 4B knockout mice will be analyzed by histochemical methods. We will evaluate functional assays depending on the results of phenotype
manifestations of Cul 4B knockout mice.
G. Production of antibodies.
7
polyclonal and/or monoclonal antibodies by immunize animals with synthetic peptides. Another strategy is to express the human Cul 4B cDNA in the T7 phage system (31) to obtain the full-length protein for immunization. We will express the Cul 4B protein using the pET15 vector in the E. coli BL21(DE3)/pLysS strain. The fusion protein will be purified and used to raise polyclonal as well as monoclonal antibodies. We are experienced in making antibodies (32-34).
H. Introduce the Myc-tagged CUL-4B constructs containing IRES-GFP into ES clones
Because CUL-4B in on the X chromosome. The ES cells we used to disrupt the Cul-4B is XY genotype and single targeting event will generate Cul-4B null ES cells. If we supply an exogenous CUL-4B by electroporation before deleting the endogenous genomic copy, we should be able to rescue the line, which will shine for the presence of GFP. If supplying a defective CUL-4B or a CUL-4B floxed by loxP as the genomic CUL-4B we have engineered for conditional KO, the clone will not survive after Cre excision and therefore, no GFP clones will be observed.
I. Expression of Cul-4B protein in yeast system
The Myc-tagged C-terminal truncated Cul-4B mutant DNA can be generated by PCR-directed mutagenesis. The Myc-tagged Cul-4B DNA is cloned into pGapZαA vector, and the plasmid is transfected to yeast. After antibiotics selection, positive clones can be obtained and screend by dot blot with anti-Myc antibody. After gel electrophoresis, the molecular weight of the protein can be determined. Purify these protein can be used to screen the antibody. To find the specific interaction proteins of Cul-4B, if the coimmunoprecipitation of mammalian system is not work, we can use this system to perform the yeast-two hybrid screen to find the Cul4B interacting proteins.
J. Expression of Cul-4B protein in mammalian system
We use SK-HEP-1 (human, Caucasian, liver, adenocarcinoma) cell to express the
Myc-tagged Cul4B. SK-HEP-1 cells were grown in 10-cm dishes in Dublecco modified Eagle medium supplement with 10% fetal bovine serum in a 5% CO2 –supplemented atmosphere. The Myc-tagged Cul-4B DNA were transfected to cells. After antibiotics selection, positive clones can be obtained and screend by dot blot with anti-Myc antibody. Cell lines stably expressing Myc-tagged CUL-4B were generated.
K. Delineating the Cul-4B interacting molecules by coimmunoprecipitation
Cell lines stably expressing Myc-tagged CUL-4B were harvested. The harvested cells were washed twice with phosphate-buffered saline, and are suspended in lysis buffer that contain 50 mM HEPES (pH7.4), 0.1%Triton X-100, 2 mM PMSF 15 µg/mL aprotinin 2 µg/mL Pepstatin and 5 µg/mL leupeptin. After incubation at 4o
C for 30 min, the lysates are centrifuged at 13,000Xg for 30 min. The supernatants are used for the immunoprecipitation experiments. Cell lysates (0.5 mg) will be incubated with beads containing covalently bound antibodies for 16 hours at 4oC. The beads will be collected by centrifugation. Precipitates are washed three
8
times with 1 ml wash buffer. The bound proteins are subjected to western blot analysis. Specific protein can be identified by peptide analysis by the Core facility on site.
L. 2D-proteomics to differentially expressed in CUL-4B expression cell line
We will perform 2D-proteomics (by core facility) to figure out if there is any proteins differentially expressed in CUL-4B expression cell line.
結果與討論
In an attempt to study the function of Cul 4B, a novel gene with unknown function, by creating Cul 4B null mice, we have spent nearly a year to clone the mouse homolog of human Cul 4B, generating a positive-negative selection based-targeting vector, and used the vector to disrupt Cul 4B in mouse embryonic stem (ES) cells.
(1) Cloning and analysis of mouse Cul 4B genomic DNA.
Isogenic Cul 4B genomic DNA was obtained by screening a 129/sv-mouse genomic library (a gift of Dr. Begue, Pasteur Institute, France) with a human Cul 4B cDNA. A total of 14 positive clones were obtained. One of the clone, Clone Q with an insert size of approximately 12.25 kb was isolated and further characterized (data not shown). By analogy to human Cul 4B, physical mapping showed that it contains 15 (exons 3-17) of the 20 exons of the mouse Cul 4B gene (fig. 2).
(2) Construction of targeting vector for tissue specific KO of the Cul 4B gene
Targeting vectors will be designed based on selection by neomycin resistance gene and HPRT gene. As shown in fig. 3 upper.
(3) Targeting of ES cells by tissue specific pPNT-HPRT-Cul 4B-3 loxP.
The transfection experiments were quite successful and nearly 264 clones were isolated and subjected to Southern blot analysis with appropriate combinations of restriction enzyme digestion and probe hybridization (fig.3). 8 candidate clones were obtained.
(4) ES cell survived after Cul-4B deleted.
Exons 12-14 will be deleted in the presence of Cre recombinase . Cre can be introduced into ES cell clone and after excision the correct clones can be identified by Southern blotting and PCR. (Fig 4) .This result indicated that the Cul4B null ES cell can be survived. Whether KO of Cul-4B is embryonic lethal awaits determination
(5) Chimera mice were generated.
We have injected the selected clones into blastocysts and implanted them into foster mice. Five chimeric mice are generated. Germ line offsprings are detected by the presence of agouti litters (Fig 5).
(6) Expression of Cul-4B in E. coli, yeast and mammalian systems.
9
to Top 10 host. The transformed cells were growth in 18 oC for 2 hours and protein expression was induced by IPTG. After IPTG induction, the specific protein can be expressed and detected by anti-Xpress epitope antibody. Fig 6 shows the construct and results of silver stain and Western blot.
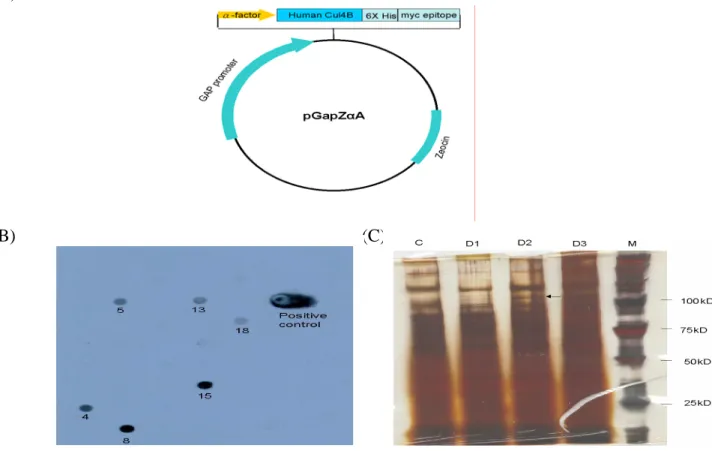
The Myc-tagged Cul-4B DNA was cloned into pGapZαA vector (fig 7A), and the plasmid was transfected to yeast. After antibiotics selection, 20 clones were selected, 6 of them can express Myc-tagged protein when screened by dot blot analysis (fig 7B). After gel
electrophoresis, specific protein was probed by anti-Myc antibody. The molecular weight is about 110 kDa (Fig 7C).
When expressed the Cul-4B in mammalian system, a CMV driven vector was used. SK-Hep-1 cells are transfected with the plasmid as shown in Fig 8 (Left). Transfected SK-Hep-1 cells with GFP expression can be observed under fluorescence microscope (Fig 8 Middle). Cellular protein were extracted and subjected to electrophoresis. Specific protein was probed by anti-Myc antibody. A band of 85 kDa was observed (Fig 8 Right).
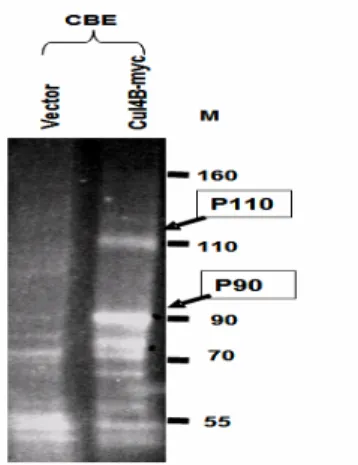
(7) Delineating the Cul-4B interacting molecules by coimmunoprecipitation Cell lines stably expressing Myc-tagged CUL-4B were subjected to
co-immunoprecipitation. The bound proteins are subjected to SDS-PAGE analysis. Sypro Ruby stain gel showing the result of Co-immunoprecipitation. (Fig.9) .Comparing to the cells transfected with vector only, Cul-4B over-expressed cells shows 2 different bands,P110 and P90 bands. The P90 is Cul-4B which is also identified by Myc antibody (data not shown). Analyzing the co-immunoprecipitated CUL-4B interacting molecules by MALDI-TOF MS/MS showed that CUL-4B interacts with TIP120A and UV-damaged DNA –binding protein , DDB1. TIP120A functions as a negative regulator of different E3 ubiquitin ligases and DDB1 was reported to be an adaptor protein to mediate substrate binding to CUL-4A
associated E3 ubiquitin ligases in regulating cell cycle. It may recruit substrates for the CUL-4B dependent ligases in a similar fashion.
(8) 2D-proteomics to differentially expressed in CUL-4B expression cell lin
We have performed 2D-proteomics that showed some interesting spots differentially expressed in CUL-4B expression cell line(fig.10). 2D gel reveals two distinct spots highly expressed in the presence or the absence of excess Cul-4B. The identity of these proteins is under investigation.
參考資料
1. Kipreos ET, Lander LE, Wing JP, He WW, and Hedgecock EM. (1996). Cul-1 is required for cell cycle exit in C. elegans and identifies a novel gene family. Cell 85: 829-839
2. Ishikawa K, Nagase T, Suyama M, Miyajima N, Tanaka A, Kotani H, Nomura N, and Ohara O. (1998). Prediction of the coding sequences of unidentified human genes. X. The
10
complete sequences of 100 new cDNA clones from brain which can code for large proteins in vitro. DNA Res 5: 169-176.
3. UniGene. http://www.ncbi.nlm.nih.gov/UniGene/
4. Yu H, Peters JM, King RW, Page AM, Hieter P, and Kirschner MW. (1998). Identification of a cullin homology region in a subunit of the anaphase-promoting complex. Science 279: 1219-22.
5. Zachariae W, Shevchenko A, Andrews PD, Ciosk R, Galova M, Stark MJ, Mann M, and Nasmyth K. (1998). Mass spectrometric analysis of the anaphase-promoting complex from yeast: identification of a subunit related to cullins. Science 279: 1216-9.
6. Michel JJ, and Xiong Y. (1998). Human CUL-1, but not other cullin family members, selectively interacts with SKP1 to form a complex with SKP2 and cyclin A. Cell Growth Differ 9: 435–449.
7. Deshaies RJ. (1999). Annu Rev Cell Dev Biol 15: 435-467.
8. Yeh ET, Gong, L. and Kamitani T. (2000). Ubiquitin-like proteins: new wines in new bottles. Gene 248: 1-14.
9. Ohta T, Michel JJ, Schottelius AJ, and Xiong Y. (1999). ROC1, a homolog of APC11, represents a family of cullin partners with an associated ubiquitin ligase activity. Mol Cell 3: 535–541
10. Jackson PK, Eldridge, A.G., Freed, E., Furstenthal, L., Hsu, J.Y., Kaiser, B.K. and Reimann, J.D. (2000). The lore of the RINGs: substrate recognition and catalysis by ubiquitin ligases. Trends Cell Biol 10: 429-439.
11. Tyers M and Jorgensen P. (2000). Proteolysis and the cell cycle: with this RING I do thee destroy. Curr Opin Genet Dev 10: 54-64.
12. Wang Y, Penfold S, Tang X, Hattori N, Riley P, Harper JW, Cross JC and Tyers M. (1999). Deletion of the Cul1 gene in mice causes arrest in early embryogenesis and accumulation of cyclin E. Curr Biol 9: 1191-1194.
13. Dealy MJ, Nguyen KV, Lo J, Gstaiger M, Krek W, Elson D, Arbeit J, Kipreos ET and Johnson RS. (1999). Loss of Cul1 results in early embryonic lethality and dysregulation of cyclin E. Nature Genet 23: 245-248.
14. Sherr C. (1994). G1 phase progression: cycling on cue. Cell 79: 551-555.
15. Spruck CH, Won KA and Reed SI. (1999). Deregulated cyclin E induces chromosome instability. Nature 401: 297-300.
16. O'Hagan R, Ohh M, David G, Moreno de Alboran I, Alt F, Kaelin JW and DePinho R. (2000). Myc-enhanced expression of Cul1 promotes ubiquitin-dependent proteolysis and cell cycle progression. Genes Dev 14: 2185-2191.
17. Maxwell PH, Wiesener MS, Chang GW, Clifford SC, Vaux EC, Cockman ME, Wykoff CC, Pugh CW, Maher ER and Ratcliffe PJ. (1999). The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 399: 271–275.
11
(2001). The von Hippel-Lindau Tumor Suppressor Protein Mediates Ubiquitination of Activated Atypical Protein Kinase C. J. Biol Chem 276: 43611-43617
19. Singer JD, Gurian-West M, Clurman B and Roberts JM. (1999). Cullin-3 targets cyclin E for ubiquitination and controls S phase in mammalian cells. Genes Dev 13: 2375– 2387.
20. Chen X, Zhang Y, Douglas L and Zhou P. (2001). UV-damaged DNA binding proteins are targets of Cul4A-mediated ubiquitination and degradation. J Biol Chem 276:
48175-48182.
21. Kamura T, Burian D, Yan Q, Schmidt SL, Lane WS, Querido E, Branton PE, Shilatifard A, Conaway RC, and Conaway JW. (2001). Muf1, a novel Elongin BC-interacting
leucine-rich repeat protein that can assemble with Cul5 and Rbx1 to reconstitute a ubiquitin ligase. J Biol Chem 276: 29748-29753.
22. Kornitzer D and Ciechanover A. (2000). Modes of regulation of ubiquitin-mediated protein degradation. J Cell Physiol 182: 1-11.
23. Kishino T, Lalande M, and Wagstaff J. (1997). UBE3A/E6-AP mutations cause Angelman syndrome. Nat Genet 15: 70–73.
24. Matsuura T, Sutcliffe JS, Fang P, Galjaard RJ, Jiang YH, Benton CS, Rommens JM, and Beaudet AL. (1997). De novo truncating mutations in E6-AP ubiquitin-protein ligase gene (UBE3A) in Angelman syndrome. Nat Genet 15: 74–77.
25. Staub O, Dho S, Henry P, Correa J, Ishikawa T, McGlade J, Rotin D. (1996). WW domains of Nedd4 bind to the proline-rich PY motifs in the epithelial Na1 channel deleted in Liddle’s syndrome. EMBO J 15: 2371–2380.
26. Fero ML, Randel E, Gurley KE, Roberts JM, Kemp CJ. (1998). The murine gene p27Kip1 is haplo-insufficient for tumour suppression. Nature 396:177–180.
27. Colucci-Guyon E, Portier MM, Dunia I, Paulin D, Pournin S, Babinet C. (1994). Mice lacking vimentin develop and reproduce without an obvious phenotype. Cell 79: 679-694.
28. Lin SR, Yu IS, Huang PH, Tsai CW, and Lin SW. (2000). Characterization of chimeric mice with disruption of the gene coding for phosphatidylinositol glycan-A (Pig-a). Brit J Haematology 110: 682-693.
29. Yu IS, Chen HJ, Lee YS, Huang PH, Lin SR and Lin SW. (2000). Mouse deficient of hepsin, a serine protease, exhibits normal embryogenesis and unchanged hepatocyte regeneration ability. Thrombosis and Haemostasis 84: 865-875.
30. Hogan B, Beddington R, Costantini F, and Lacy E. (1994). Manipulating the mouse embryo. 2nd Edition, CSH Laboratory Press.
31. Studier FW, Rosenberg A, Dunn J and Dubendorff J. (1990). Use of T7 RNA polymerase to direct expression of cloned genes. Methods Enzymology 185: 60-88.
32. Lin SW, Lin CN, Hamaguchi N, Smith KJ, and Shen MC. (1994). Characterization of a factor IX variant with a glycine207 to glutamic acid mutation. Blood 84: 1866-1873.
33. Lin SW, Dunn JJ, Studier FW, and Stafford DW. (1987). Expression of human factor IX and its subfragments in Escherichia coli and generation of antibodies to the subfragments.
12 Biochemistry 28: 5267-5274.
34. Huang CC, Li LT, Shen MC, Chen JY, and Lin SW. (2001). Domain specific monoclonal anti-factor VIII antibodies generated by inclusion body-renatured factor VIII peptides. Thrombosis Research 101: 405-415.
13
計畫成果自評
本研究自得到小鼠同源 Cul 4B 基因後即開始進行研究,由反轉錄聚合脢鏈反應 (RT-PCR)證實 Cul 4B 在 ES 細胞已表現,由於 Cul 4B 位於 X 染色體上,被剔除後 ES 細胞無正常的 Cul 4B 功能支持,可能因此導至 ES 細胞不易存活。所以利用組織特異性 (tissue specific)基因剔除鼠技術,進行 Cul 4B 的基因剔除,以得到 Cul 4B 基因剔除鼠。
首先進行標的質體的構築,當將標的質體轉感入小鼠胚胎幹細胞(embryonic stem cell; ES cells)中,目前已完成所有標的 DNA 之建構,至並經兩次轉染實驗,篩選得到 264 個 ES 細胞株經南方墨點法確認,有 8 個在 cul4B 基因序列中具有 loxP 序列的細胞 株,將此 ES 細胞株以顯微注射的方式打入囊胚,嵌合鼠已出生,嵌合鼠與黑色 B6 母 鼠互配後,得到了栗色毛的子代,證明其遺傳來源為胚胎幹細胞,且為 Cul4B 基因標定 的異型合子。由此一異型合子交配而得到同型合子後,相信很快可以進行分析。 篩選得到的 ES 細胞株經南方墨點法確認,具有 loxP 序列的細胞株以帶有 Cre 的 質體轉感後可以得到的 Cul 4B 基因剔除的 ES 細胞株,初步確認 Cul 4B 基因剔除的 ES 細胞株仍可存活。 在細胞培養方面,已成功表現 Cul 4B 蛋白,並以共同免疫沈澱的方式將細胞內與 Cul4B 有交互作用的蛋白沈澱下來;以質譜分析鑑別,發現 TIP120A 及 DDB1 與 Cul4B 有交互作用。
Cullin family 蛋白對細胞的生長及分化相當重要,而 cullin 4B 目前還沒有研究團 隊發表有關該基因的的生理功能或重要性,因此本研究極具發表潛力,就整個計劃進度 而言已達到預期目標。
14 附錄 Figures:
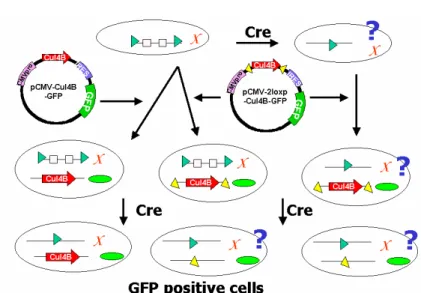
Fig 1 Dissecting whether Cul 4B is essential for ES cell survival by gene targeting in mouse embryonic stem cell. ES cells (oval circle) harboring a loxP (triangle)-floxed genomic Cul-4B (on the X chromosome) were generated by conditional gene-targeting strategy. The ES cells can be subjected to Cre deletion experiments. If Cul-4B is essential for ES cell survival, one would not be able to recover the ES cells (top right). We plan to supply vectors
(pCMV-cul4b-GFP and pCMV-2loxP-cul4b-GFP) carrying Cul-4B cDNA (arrow bars) and GFP marker (small oval circle) into the ES cells with the loxP-floxed Cul-4B (or the ES cells that should survive from Cre-excision). Upon Cre excision of these ES cell clones, the ES cells carrying CUL-4B protein expressed from the Cul-4B cDNA will be isolated and shine with GFP (bottom left). When the ES clones (bottom center and right) transfected with
pCMV-2loxP-cul4B and thus carrying Cul-4B cDNA also flanked by additional loxP will not have CUL-4B proteins after Cre excision because of the loss of both chromosomal and plasmid copies of the Cul-4B genes. Whether one can recover this cell by identifying GFP means that CUL-4B is essential for ES cell survival or not.
15
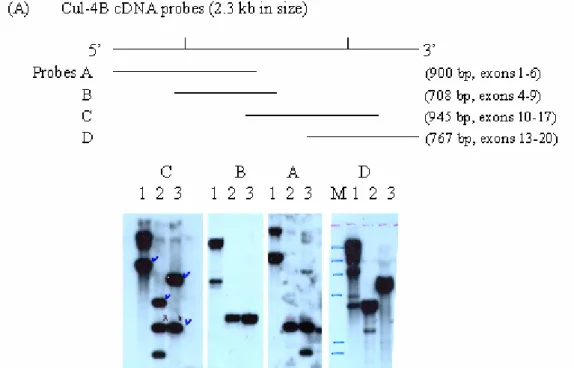
1:phage Q digested with BamHI. 2: digested with BamHI and EcoRI. 3: digested with EcoRI. M: phage lambda DNA digested with HindIII. Note that as references of sizes of the signals, the signals in probe C, lane 3 are 5.5 kb and 2.7 kb, in probe B is 2.7 kb and in probe A are 5.5,2.3 and 1.65 kb, respectively.
Fig. 2. Restriction enzyme mapping of the DNA structure of mouse cullin 4B genome. A human cullin 4B cDNA was used to screen a mouse genomic DNA library constructed by inserting the BamHI treated total genomic DNA from the 129sv strain mice into lambda phages. More than 10 phage clones carrying different and overlapping fragments were identified and isolated. Phage clone Q containing a 12.25 kb insert covering exons 6-17 was identified by sequencing with exon specific primers and by Southern blotting of the EcoRI and BamHI single- or double-digested DNA probed with 3 cDNA subfragments (probes A-C) of cul-4B (panle A). The 12.25 kb insert was subjected to detailed restriction enzyme mapping using more than 15 different enzymes. Only results of the mapping of EcoRI and BamHI were shown below. The 12.25 kb insert can be divided in to 5 EcoRI fragments or 4 BamHI
fragments. The EcoRI fragments were, from 5’ to 3’, 1.65 (fused to the left arm of the lambda vector), 0.65, 2.3, 2.7, and 5.5 kb (fused to the right arm of the vector) in size. The BamHI fragments were, from 5’ to 3’, 1.3, 0.2-0.3, 7.0 and 3.7 kb. The sizes of the introns are not to scale (panel B).
16
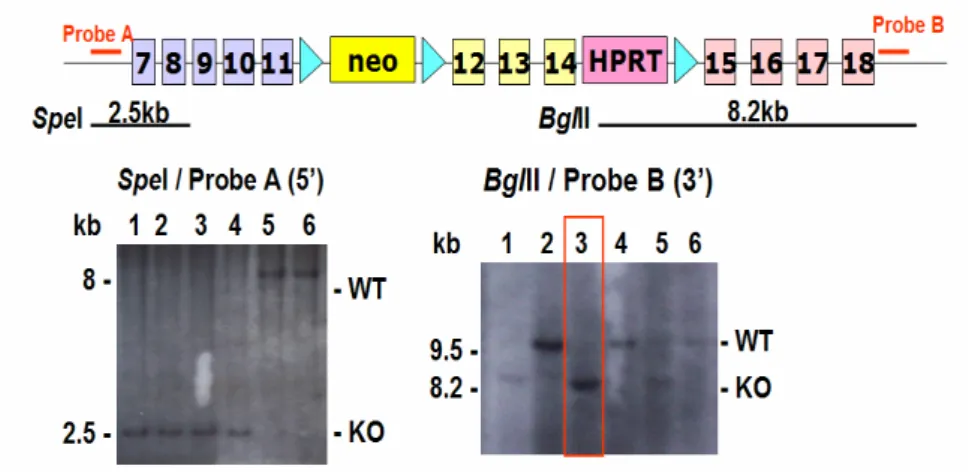
Fig. 3. Conditional loxP-floxed Cul-4B allele. Green triangle: loxP neomycine and HPRT are selection markers. Exons 12-14 will be deleted in the presence of Cre recombinase . Genotyping of the ES cell clones are performed by Southern blotting showing correct configurations with both 5’ and 3’ probes. Lanes 1-4 (left panel) and lane 3 (right panel) are KO lines..
Fig.4. Genomic structure of mouse Cul 4B gene and the targeting construct for whole - body knockout of the Cul 4B gene in ES cell. Green triangle: loxP neomycine and HPRT are selection markers. Exons 12-14 will be deleted in the presence of Cre recombinase . Cre can be introduced into ES cell clone and after excision the correct clones can be identified by Southern blotting and PCR. (left, Southern) lanes 3,4 are KO configuration lanes 1,2,5 are before Cre. (right, PCR) lanes 1,2 are KO and lane 3 is before Cre excision.
17
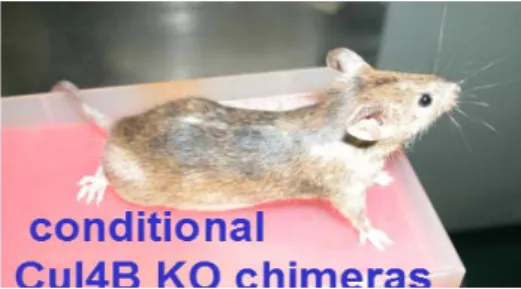
Fig 5. The ES cells of fig.2 are injected into blastocyst to obtain chimera (left) harboring the conditional loxP-floxed ul-4B allele which gave germline transmission of agouti mice.
Fig.6 Expression of full-length Cul-4B in E coli system. Left: construct of bacteria
expression plasmid. Middle: Silver stain of the bacterial protein. Right: western blot analysis probed by antiXpress epitope antibody. S1: Supernatants of Bacteria lysate before IPTG induction. S2: Supernatants of bacteria lysate after IPTG induction. P1: Pellets of Bacteria lysate before IPTG induction. S2: Pellets of bacteria lysates after IPTG induction.
18 (A)
(B) (C)
Fig. 7 Expression of Myc-tagged Cul-4B protein by a yeast system. Left: construct of yeast expression plasmid. Middle: Myc-tagged Cul-4B cDNA was transfected to yeast. After growing in antibiotic containing medium for two days, the yeast was spin down, the
supernatant were collected and subjected to dot blot analysis. 6 positive clones were obtained when probed with anti-Myc antibody. Right: Clone 15 is subjected to gel electrophoresis C: Yeast before plasmid transfection as negative control, D1: supernatant after 1 day culture, D1: supernatant of after 1 day culture, M: marker.
55
Vector Cul4B-myc90
70
110
55
Vector Cul4B-myc90
70
110
Fig 8. Expression of Myc-tagged Cul-4B in mammalian cells. Left: the constructs for the mammalian expression plasmids. Right: Cell extracts were subjected to Western blot analysis and probed by anti-Myc antibody.
19
Fig 9. Co-immunoprecipitation. Sypro Ruby stain gel showing the result of
Co-immunoprecipitation identifying the TIP120A and DDB1 proteins in over-expressed SK-Hep1 cell lines (right lane), showing P110 bands. The P90 is Cul-4B which is also identified by Myc antibody (data not shown).
Fig. 10 2D-proteomics to differentially expressed in CUL-4B expression cell line (left two panels). Two-D gel reveals two distinct spots (circled in red) highly expressed in the presence (lower) or the absence (upper) of excess Cul-4B. The identity of these two spots is in progress. (right panel).