行政院國家科學委員會專題研究計畫 成果報告
自然弒細胞發生學的雙系模式:白血球介質 15 及 21 分別導
引 CD56+Kir±lectis+ 及 CD56±Kirlectin 自然弒細胞
(3/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2314-B-002-005- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學醫學院免疫學研究所 計畫主持人: 許世明 報告類型: 完整報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 94 年 8 月 3 日
90/12/21 修訂
Introduction
We have proposed a moedl for NK-cell development
Mature human NK-cells are heterogeneous, and their developments are incompletely understood. We want to prove that interleukin 15 and interleukin 21 drive respectively the development of CD56brigthtKIRdimLectinbright type I (IFN-γ+) NK-cells and CD56dimKIRbrightLectindim type II (IL-13+) NK-cells. Biologically, this model can be used to further analyze the interrelationship between various NK-cell subsets. Clinically, this model can be used to further sub-classify NK-cell lymphomas or to analyze the role of NK-cells in transplantation pathology. Thus this model will have both biological and clinical significance. As summarized in the abstract and to be expanded in details in the Background section, we will use a developmental approach to integrate previous functional-structural studies. More specifically, we will achieve the following:
(1) Purification of NK-precursors. NK-cells and T-cells arise from a common CD44+, CD25-, CD161+ bipotent T/NK precursor, which subsequently diverges into T-cells under interleukin 7 and the transcription factors Notch and E2A, or into NK-cells under interleukin 15 and the transcription factor Id3. Id3 and E2A are both helix-loop-helix transcription factors, and interleukin 7 and interleukin 15 have a similar structure of 4-alpha-helical bundles using a common γ-chain (γc) as part of their receptors. Based on this observation, we propose to isolate by flow cytometry the common bipotent T/NK cells from bone marrow, thymus, or peripheral blood as the starting point
90/12/21 修訂
for the study of NK-cell development. These rare bipotent T/NK precursors should be CD44+, CD25-, CD161+, γc +. They will be positively selected for interleukin 15 receptor and Id3, and negative selected for interleukin 7 receptor and E2A, and used as the starting materials for NK-cell development.
(2) CD56brigtht vs CD56dim. Human NK-cells uniquely express CD56, and could be divided into a CD56brigtht subset and a CD56dim subset. We want to show that, based in part on recent reports, that NK-cell precursors will develop into either the CD56brigtht subset in the presence of interleukin 15 or the CD56dim subset in the presence of interleukin 21.
(3) Type I vs Type II. NK-cells can be divided into type I NK-cells secreting predominantly IFN-γ and type II NK-cells secreting predominantly interleukin 4 and interleukin 13. Traditionally, it is thought that a common type zero cell will give rise to either type I or type II cells. However, recently data seems to indicate that, in fact, the development follows a type II-type 0-type I sequence, an important issue to be clarified. We want to determine which one of these model, dual lineage vs sequential model, is correct.
(4) C-type Lectin vs KIR. Human NK-cells use both C-type lectins and KIRs as their antigen receptors. C-type lectin might be expressed earlier in the development of NK-cells, and represents a more primitive antigen receptor. But it is still unclear how the expression patterns of C-type lectins and KIRs are related to each other and to their functions, type I vs type II. We want to resolve these problems.
(5) A dual lineage model for the development of mature human NK-cells in which interleukin 15 and interleukin 21 drive respectively the development of CD56brigthtKIRdimLectinbright type I (IFN-γ+) NK-cells and CD56dimKIRbrightLectindim type II (IL-13+) NK-cells. This will be done through integration of the data in step 2,3, & 4.
(6) Clinical application. One obvious application is to continue our previous works in the classification of sinonasal lymphoma. Another field of possible application is in tthe transplantation pathology, ie, whether NK-cells participate in the rejection of graft.
90/12/21 修訂
Results
1) We have used the model to identify NK-prcursors from the thymus, and establised the existence of malignat transformation of these precursors into an NK-lymphoblastic lymphoma. For details, please refer to the attached paper, recently published in Blood.
2) We are currently working on the Type I and Type II mature NK-cells. A manuscript is in progress.
90/12/21 修訂
參考文獻(Reference)
Preliminary Data from our lab
Lien HC, Lin CW, Huang PH, Chang ML, Hsu SM: Expression of cyclin-dependent kinase 6 (cdk6) and frequent loss of CD44 in nasal-nasopharyngeal NK/T cell lymphomas: comparison with CD56-negative peripheral T cell lymphomas. Lab Invest 2000, 80:893-900.
Lin CW, Lee WH, Chang CL, et al. Restricted killer cell Immunoglobulin-like receptor repertoire without T-cell receptor γ rearrangement supports a true NK-cell lineage in a subset of sinonasal lymphomas. Am J Path 2001;159: 1671
Lin CW, Yang JY, et al. KIR+ T-cell lymphoma: Restricted killer immunoglobulin-like receptor repertoire and monoclonal T-cell receptor g rearrangement in a subset of sinonasal lymphoma with dual NK/T cell differentiation. Submitted to Blood.
Models of NK-cell development
Barten R, Torkar M, et al. Divergent and convergent evolution of NK-cell receptors. Trends Immunology, 2001, 22, 52-57.
Loza, MJ, Perussia B. Final steps of natural killer cell maturation: a model for type 1-type 2 differentiation? Nature Immunology, 2001, 10, 917-924.
Parrish-Novak J, Dillion SR, et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature, 2000, 408, 57-63.
Cooper, MA, Fehniger TA, Caligiuri MA. The biology of human natural killer cell subsets (Review). Trends Immunology, 2001, 22, 623-640.
Fehnigewr TA, Caliguiri MA. Interleukin 15: Biology and relevance to human disease (Review). Blood, 2001, 97, 14-32.
NK and T-cell development
Spits H, Lanier LL, Phillips JH: Development of human T and natural killer cells. Blood 1995, 85:2654-2670.
Carlyle JR, Michie AM, Furlonger C, Nakano T, Lenardo MJ, Paige CJ, and Zúñiga-Pflücker JC. Identification of a novel developmental stage marking lineage commitment of progenitor thymocytes. J Exp Med, 1997, 186:173-182.
Mingari MC, Vitale C, Cantoni C, Bellomo R, Ponte M, Schiavetti F, Bertone S, Moretta L: Interleukin-15 induced maturation of human natural killer cells from early thymic precursors: selective expression of CD94/NKG2-A as the only HLA class I-specific inhibitory receptor. Eur J Immunol 1997, 27:1374-1380.
Yu H, Feniger TA, et al. Flts3 ligand promotes the generation of a distinct CD34+ human natural killer cell progenitor that respond to interleukin 15. Blood, 1998, 92, 3647-3657.
Parrish J, Dillion SR, et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature, 2000, 408, 57-63.
Ozaki K, Kikly K, et al. Cloning of as type I cytokine receptor most related to the IL-2 receptor β chain. Proc Natl Aca Sci, 2000, 97, 21, 11439-11444.
Liu CC, Perussia B, Young JDE. The emerging role of IL-15 in NK-cell development. Trends Immunology, 2000, 21, 113-116.
Fehniger TA, Caliguiri MA. Interleukin 15: biology and relevance to human disease. Blood, 2001, 97, 14-32.
90/12/21 修訂
Immunology, 2001, 22, 633-640.
Expression of T-cell markers in fetal NK-cells
Phillips JH, Hori T, Nagler A, Bhat H, Spits H, Lanier LL: Ontogeny of human natural killer cells: Fetal NK cells mediate cytolytic function and express cytoplasmic CD3ε, δ proteins. J Exp Med 1982, 175:1055-1066.
Lanier LL, Chang C, Spits H, Phillips JH: Expression of cytoplasmic CD3ε proteins in activated human adult NK cells and CD3γ,δ,ε complexes in fetal NK cells: Implications for the relationship of NK and T lymphocytes. J Immunol 1992, 14:1876-1880.
Suzumiya J, Takeshita M, Kimura N, Kikuchi M, Uchida T, Hisano S, Eura Y, Kozuru M, Nomura Y, Tomita K, Komiyama S, Okumura M: Expression of adult and fetal natural killer cell markers in sinonasal lymphomas. Blood 1994, 83:2255-2260.
Killer cell antigen receptor
Moretta A, Botino C, Vitale M, Pende D, Biassoni R, Mingari MC, and Moretta L: Receptors for HLA class I molecules in human natural killer cells. Annu Rev Immunol 1996, 14:619-648. Wilson MJ, Torkar M, Trowsdale J. Genomic organization of a human killer cell inhibitory receptor
gene. Tissue Antigens.1997;49:574-579.
Steffens U, Vyas Y, Dupont B, et al. Nucleotide and amino acid sequence alignment for human killer cell inhibitory receptors (KIR), 1998. Tissue Antigens.1998;51:386-413.
Long EO and Rajagopalan S: HLA class I recognition by killer cell Ig-like receptors. Semin Immunol 2000, 12:101-108.
Raulet DH: Development and tolerance of natural killer cells. Curr Opini Immunol 2000, 11:129-134.
Raulet DH, Vance RE, McMahon CW. Regulation of the natural killer cell receptor repertoire. Annu Rev Immunol. 2001;19:291-330.
Special NK- or T-subsets: KIR+ T-cell & NKT-cells
Mingari MC, Schiavetti F, Ponte M, Vitale C, Maggi E, Romagnani S, Demarest J, Pantaleo G, Fauci AS, Moretta L: Human CD8+ T lymphocyte subset that express HLA class I specific inhibitory receptors represent oligoclonally or monoclonally expanded cell populations. Proc Natl Acad Sci USA 1996, 93:2433-12438.
Bertrand H, Karlsson L. KIR expression on self-reactive CD8+ T cells is controlled by T-cell receptor engagement. Nature. 2000;403:321-328.
McMahon CW, Raulet D. Expression and function of NK cell receptors in CD8+ T cells. Current Opinion Immunol. 2001;13: 465-470.
Ugolini S, Arpin C, Anfossi N, et al. Involvement of inhibitory NKRs in the survival of a subset of memory-phenotype CD8+ T cells. Nat Immunol. 2001;2:430-435.
Godfrey, DI, Hammond JL, Poulton LD, Smyth MJ, Baxter AG. NKT cells: fact, functions and fallacies. Immunol Today. 2000;21:573-583.
Sinonasal lymphomas of NK- and T-lineages
Kanavaros P, Lescs MC, Briere J, Divine M, Galateau F, Joab I, Bosq J, Farcet JP, Reyes F, and Gaulard P. Nasal T-cell lymphoma: a clinicopathologic entity associated with peculiar phenotype and with Epstein-Barr virus. Blood 1993, 81:2688-2695.
Nagata H, Konno A, Kimura N, Zhang Y, Kimura M, Demachi A, Sekine T, Yamamoto K, Shimizu N: Characterization of novel natural killer (NK)-cell and gammadelta T-cell lines established from primary lesions of nasal T/NK-cell lymphomas associated with the Epstein-Barr virus. Blood 2001, 97:708-713.
90/12/21 修訂
Introduction
We have proposed a moedl for NK-cell development
Mature human NK-cells are heterogeneous, and their developments are incompletely understood. We want to prove that interleukin 15 and interleukin 21 drive respectively the development of CD56brigthtKIRdimLectinbright type I (IFN-γ+) NK-cells and CD56dimKIRbrightLectindim type II (IL-13+) NK-cells. Biologically, this model can be used to further analyze the interrelationship between various NK-cell subsets. Clinically, this model can be used to further sub-classify NK-cell lymphomas or to analyze the role of NK-cells in transplantation pathology. Thus this model will have both biological and clinical significance. As summarized in the abstract and to be expanded in details in the Background section, we will use a developmental approach to integrate previous functional-structural studies. More specifically, we will achieve the following:
(1) Purification of NK-precursors. NK-cells and T-cells arise from a common CD44+, CD25-, CD161+ bipotent T/NK precursor, which subsequently diverges into T-cells under interleukin 7 and the transcription factors Notch and E2A, or into NK-cells under interleukin 15 and the transcription factor Id3. Id3 and E2A are both helix-loop-helix transcription factors, and interleukin 7 and interleukin 15 have a similar structure of 4-alpha-helical bundles using a common γ-chain (γc) as part of their receptors. Based on this observation, we propose to isolate by flow cytometry the common bipotent T/NK cells from bone marrow, thymus, or peripheral blood as the starting point
90/12/21 修訂
for the study of NK-cell development. These rare bipotent T/NK precursors should be CD44+, CD25-, CD161+, γc +. They will be positively selected for interleukin 15 receptor and Id3, and negative selected for interleukin 7 receptor and E2A, and used as the starting materials for NK-cell development.
(2) CD56brigtht vs CD56dim. Human NK-cells uniquely express CD56, and could be divided into a CD56brigtht subset and a CD56dim subset. We want to show that, based in part on recent reports, that NK-cell precursors will develop into either the CD56brigtht subset in the presence of interleukin 15 or the CD56dim subset in the presence of interleukin 21.
(3) Type I vs Type II. NK-cells can be divided into type I NK-cells secreting predominantly IFN-γ and type II NK-cells secreting predominantly interleukin 4 and interleukin 13. Traditionally, it is thought that a common type zero cell will give rise to either type I or type II cells. However, recently data seems to indicate that, in fact, the development follows a type II-type 0-type I sequence, an important issue to be clarified. We want to determine which one of these model, dual lineage vs sequential model, is correct.
(4) C-type Lectin vs KIR. Human NK-cells use both C-type lectins and KIRs as their antigen receptors. C-type lectin might be expressed earlier in the development of NK-cells, and represents a more primitive antigen receptor. But it is still unclear how the expression patterns of C-type lectins and KIRs are related to each other and to their functions, type I vs type II. We want to resolve these problems.
(5) A dual lineage model for the development of mature human NK-cells in which interleukin 15 and interleukin 21 drive respectively the development of CD56brigthtKIRdimLectinbright type I (IFN-γ+) NK-cells and CD56dimKIRbrightLectindim type II (IL-13+) NK-cells. This will be done through integration of the data in step 2,3, & 4.
(6) Clinical application. One obvious application is to continue our previous works in the classification of sinonasal lymphoma. Another field of possible application is in tthe transplantation pathology, ie, whether NK-cells participate in the rejection of graft.
90/12/21 修訂
Results
1) We have used the model to identify NK-prcursors from the thymus, and establised the existence of malignat transformation of these precursors into an NK-lymphoblastic lymphoma. For details, please refer to the attached paper, recently published in Blood.
2) We are currently working on the Type I and Type II mature NK-cells. A manuscript is in progress.
90/12/21 修訂
參考文獻(Reference)
Preliminary Data from our lab
Lien HC, Lin CW, Huang PH, Chang ML, Hsu SM: Expression of cyclin-dependent kinase 6 (cdk6) and frequent loss of CD44 in nasal-nasopharyngeal NK/T cell lymphomas: comparison with CD56-negative peripheral T cell lymphomas. Lab Invest 2000, 80:893-900.
Lin CW, Lee WH, Chang CL, et al. Restricted killer cell Immunoglobulin-like receptor repertoire without T-cell receptor γ rearrangement supports a true NK-cell lineage in a subset of sinonasal lymphomas. Am J Path 2001;159: 1671
Lin CW, Yang JY, et al. KIR+ T-cell lymphoma: Restricted killer immunoglobulin-like receptor repertoire and monoclonal T-cell receptor g rearrangement in a subset of sinonasal lymphoma with dual NK/T cell differentiation. Submitted to Blood.
Models of NK-cell development
Barten R, Torkar M, et al. Divergent and convergent evolution of NK-cell receptors. Trends Immunology, 2001, 22, 52-57.
Loza, MJ, Perussia B. Final steps of natural killer cell maturation: a model for type 1-type 2 differentiation? Nature Immunology, 2001, 10, 917-924.
Parrish-Novak J, Dillion SR, et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature, 2000, 408, 57-63.
Cooper, MA, Fehniger TA, Caligiuri MA. The biology of human natural killer cell subsets (Review). Trends Immunology, 2001, 22, 623-640.
Fehnigewr TA, Caliguiri MA. Interleukin 15: Biology and relevance to human disease (Review). Blood, 2001, 97, 14-32.
NK and T-cell development
Spits H, Lanier LL, Phillips JH: Development of human T and natural killer cells. Blood 1995, 85:2654-2670.
Carlyle JR, Michie AM, Furlonger C, Nakano T, Lenardo MJ, Paige CJ, and Zúñiga-Pflücker JC. Identification of a novel developmental stage marking lineage commitment of progenitor thymocytes. J Exp Med, 1997, 186:173-182.
Mingari MC, Vitale C, Cantoni C, Bellomo R, Ponte M, Schiavetti F, Bertone S, Moretta L: Interleukin-15 induced maturation of human natural killer cells from early thymic precursors: selective expression of CD94/NKG2-A as the only HLA class I-specific inhibitory receptor. Eur J Immunol 1997, 27:1374-1380.
Yu H, Feniger TA, et al. Flts3 ligand promotes the generation of a distinct CD34+ human natural killer cell progenitor that respond to interleukin 15. Blood, 1998, 92, 3647-3657.
Parrish J, Dillion SR, et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature, 2000, 408, 57-63.
Ozaki K, Kikly K, et al. Cloning of as type I cytokine receptor most related to the IL-2 receptor β chain. Proc Natl Aca Sci, 2000, 97, 21, 11439-11444.
Liu CC, Perussia B, Young JDE. The emerging role of IL-15 in NK-cell development. Trends Immunology, 2000, 21, 113-116.
Fehniger TA, Caliguiri MA. Interleukin 15: biology and relevance to human disease. Blood, 2001, 97, 14-32.
90/12/21 修訂
Immunology, 2001, 22, 633-640.
Expression of T-cell markers in fetal NK-cells
Phillips JH, Hori T, Nagler A, Bhat H, Spits H, Lanier LL: Ontogeny of human natural killer cells: Fetal NK cells mediate cytolytic function and express cytoplasmic CD3ε, δ proteins. J Exp Med 1982, 175:1055-1066.
Lanier LL, Chang C, Spits H, Phillips JH: Expression of cytoplasmic CD3ε proteins in activated human adult NK cells and CD3γ,δ,ε complexes in fetal NK cells: Implications for the relationship of NK and T lymphocytes. J Immunol 1992, 14:1876-1880.
Suzumiya J, Takeshita M, Kimura N, Kikuchi M, Uchida T, Hisano S, Eura Y, Kozuru M, Nomura Y, Tomita K, Komiyama S, Okumura M: Expression of adult and fetal natural killer cell markers in sinonasal lymphomas. Blood 1994, 83:2255-2260.
Killer cell antigen receptor
Moretta A, Botino C, Vitale M, Pende D, Biassoni R, Mingari MC, and Moretta L: Receptors for HLA class I molecules in human natural killer cells. Annu Rev Immunol 1996, 14:619-648. Wilson MJ, Torkar M, Trowsdale J. Genomic organization of a human killer cell inhibitory receptor
gene. Tissue Antigens.1997;49:574-579.
Steffens U, Vyas Y, Dupont B, et al. Nucleotide and amino acid sequence alignment for human killer cell inhibitory receptors (KIR), 1998. Tissue Antigens.1998;51:386-413.
Long EO and Rajagopalan S: HLA class I recognition by killer cell Ig-like receptors. Semin Immunol 2000, 12:101-108.
Raulet DH: Development and tolerance of natural killer cells. Curr Opini Immunol 2000, 11:129-134.
Raulet DH, Vance RE, McMahon CW. Regulation of the natural killer cell receptor repertoire. Annu Rev Immunol. 2001;19:291-330.
Special NK- or T-subsets: KIR+ T-cell & NKT-cells
Mingari MC, Schiavetti F, Ponte M, Vitale C, Maggi E, Romagnani S, Demarest J, Pantaleo G, Fauci AS, Moretta L: Human CD8+ T lymphocyte subset that express HLA class I specific inhibitory receptors represent oligoclonally or monoclonally expanded cell populations. Proc Natl Acad Sci USA 1996, 93:2433-12438.
Bertrand H, Karlsson L. KIR expression on self-reactive CD8+ T cells is controlled by T-cell receptor engagement. Nature. 2000;403:321-328.
McMahon CW, Raulet D. Expression and function of NK cell receptors in CD8+ T cells. Current Opinion Immunol. 2001;13: 465-470.
Ugolini S, Arpin C, Anfossi N, et al. Involvement of inhibitory NKRs in the survival of a subset of memory-phenotype CD8+ T cells. Nat Immunol. 2001;2:430-435.
Godfrey, DI, Hammond JL, Poulton LD, Smyth MJ, Baxter AG. NKT cells: fact, functions and fallacies. Immunol Today. 2000;21:573-583.
Sinonasal lymphomas of NK- and T-lineages
Kanavaros P, Lescs MC, Briere J, Divine M, Galateau F, Joab I, Bosq J, Farcet JP, Reyes F, and Gaulard P. Nasal T-cell lymphoma: a clinicopathologic entity associated with peculiar phenotype and with Epstein-Barr virus. Blood 1993, 81:2688-2695.
Nagata H, Konno A, Kimura N, Zhang Y, Kimura M, Demachi A, Sekine T, Yamamoto K, Shimizu N: Characterization of novel natural killer (NK)-cell and gammadelta T-cell lines established from primary lesions of nasal T/NK-cell lymphomas associated with the Epstein-Barr virus. Blood 2001, 97:708-713.
1
CD94 1A transcripts characterize lymphoblastic
lym-phoma/leukemia of immature natural killer cell origin with
dis-tinct clinical features
Chung-Wu Lin,1Ting-Yun Liu,1 Shee-Uan Chen,2 Kun-Teng Wang,1 L. Jeffrey Medeiros3 and
Su-Ming Hsu1
Departments of Pathology,1 Obstetrics and Gynecology,2National Taiwan University Hospital,
Na-tional Taiwan University College of Medicine, Taipei, Taiwan and the Department of
Hematopa-thology,3The University of Texas M.D. Anderson Cancer Center, Houston, Texas
Key words: lymphoblastic lymphoma, natural killer cell, CD94
Contents: 29 text pages, 4 figures, 6 tables
Address correspondence to:
Su-Ming Hsu, M.D.
Department of Pathology,
National Taiwan University College of Medicine,
1-1 Jen-Ai Road, Taipei, Taiwan 10016
Blood First Edition Paper, prepublished online July 26, 2005; DOI 10.1182/blood-2005-02-0519
2
Author contribution statements:
Chung-Wu Lin proposed that CD94 1A transcripts might be used by immature NK cells and wrote
the initial manuscript. Ting-Yun Liu and Kun-Teng Wang carried out most of the experiments.
Shee-Uan Chen collected the cord blood and isolated the progenitor cells. L. Jeffrey Medeiros
re-viewed the data and participated in the writing of the manuscript. Su-Ming Hsu supervised the
3
Abbreviations:
2M: beta-2-microglobulin; DN: double negative; E2A: E2A immunoglobulin enhancer binding
factor; EBER: EBV-encoded small RNA; EBV: Epstein-Bar virus; ID2: inhibitor of differentiation
2; IL-2: interleukin 2; IL-2R : interleukin-2 receptor alpha; IL-15: interleukin-15; IL-15R :
inter-leukin-15 receptor alpha; KIR: killer immunoglobulin-like receptor; LBL: lymphoblastic
lym-phoma; LCM: laser capture microdissection; Lin: lineage; NK: natural killer; NK-LBL: LBL of the
NK lineage; rLCM: reverse LCM; LBL: LBL of the T lineage; TCF-1: cell factor 1; TCR:
T-cell antigen receptor; TdT: terminal deoxynucleotidyl transferase; WHO: World Health
4
Abstract
Most lymphoblastic lymphomas (LBLs) are regarded as neoplasms of immature T-cells because
they express cytoplasmic CD3 and frequently carry T-cell receptor (TCR) gene rearrangements.
Immature natural killer (NK)- and T-cells, however, have a common bipotent T/NK-cell precursor
in the thymus and NK-cells also express cytoplasmic CD3. Thus, some LBLs could arise from
im-mature NK-cells. Mature NK-cells express two CD94 transcripts: 1A, induced by IL-15, and 1B
constitutively. Since immature NK-cells require IL-15 for development, CD94 1A transcripts could
be a marker of NK-LBL. To test this hypothesis, we used laser capture microdissection to isolate
IL-15 receptor + lymphoid cells from the thymus and showed that these cells contained CD94 1A
transcripts. We then assessed for CD94 transcripts in 21 cases of LBL that were cytoplasmic CD3+,
nuclear TdT+, and CD56 , consistent with either the T- or NK-cell lineage. We found that 7 LBLs
expressed CD94 1A transcripts without TCR gene rearrangements, suggesting NK-cell lineage.
Pa-tients with NK-LBL were younger than paPa-tients with T-LBL (15 years versus 33 years; p=0.11) and
had a better 2-year survival (100% versus 27%; p<0.01). These results improve the current
5
Introduction
Precursor lymphoblastic lymphoma/leukemia (LBL) frequently presents as a mediastinal mass
in young patients. Immunophenotypically, most neoplasms are positive for cytoplasmic CD3 and
nuclear terminal deoxynucleotidyl transferase (TdT) and carry T-cell receptor (TCR)
rearrange-ments.1 Thus, most LBL cases are thought to be of T-lineage and represent lymphomas of
imma-ture T-cells from the thymus. However, it is difficult to distinguish immaimma-ture T-cells from immaimma-ture
NK-cells because both are derived from bipotent T/NK-precursors in the thymus and share similar
immunophenotypes.2-5 Cytoplasmic CD3 can be found in both immature T- and NK-cells.6
Simi-larly, nuclear TdT is expressed by lymphoid precursors of various lineages and is not specific for
T-cells.7In addition, TCR gene rearrangement, despite being a good marker for T-cells, can be found
in cases of acute myeloid leukemia and precursor B-cell lymphoblastic leukemia due to the
phe-nomenon of lineage promiscuity.8 These traditional criteria, therefore, do not exclude the possibility
that lymphoblastic lymphoma/leukemia of the NK-lineage exists.9 Although CD56 (neural cell
ad-hesion molecule; NCAM) has been recognized as a sensitive marker for NK-cells, particularly
ma-ture NK-cells,10 CD56 is not specific and can be expressed by neoplastic myeloid, lymphoid,
plas-macytoid dendritic,11-13 and plasma cells. Other than CD56, CD161 and CD44 could be used as
markers for NK-14-17 and T-cells,18,19respectively. but their sensitivity and specificity in
distinguish-ing LBLs from T-LBLs have yet to be tested. The lack of suitable markers for immature
NK-cells thereby makes it difficult to identify NK-cell LBL.
For the NK-cell lineage to diverge from the T-cell lineage, interleukin (IL)-15 and
transcrip-tion factor ID2 are essential. By contrast, IL-2 and transcriptranscrip-tion factor E2A are important for T-cell
6
family of transcription factors. ID2 is a dominant-negative inhibitor of E2A and promotes NK-cell
development at the expense of T-cell development.27,28 Likewise, IL-2 and IL-15 are highly
ho-mologous, and their receptors are heterotrimers that share the same and chains, but differ in the
chains.29 The IL-2 receptor (IL-2R ) is also known as CD25, whereas the IL-15 receptor
(IL-15R ) does not yet have a cluster designation (CD). Accordingly, NK-cell precursors most likely express IL-15R and ID2, and T-cell precursors are positive for CD25 and E2A.
T-cell precursors undergo TCR gene rearrangement and become immature T-cells that are
committed to the T-cell lineage.30Expansion and survival of immature T-cells depend upon another
transcription factor, TCF-1,31,32 that is also present in a subset of mature NK-cells.33 For NK-cell
precursors, their subsequent development and lineage commitment to immature NK-cells is unclear.
It is known only that mature human NK-cells use both the C-type lectin-like CD94 and killer
im-munoglobulin-like receptors (KIRs) as antigen receptors.34-36 Recently, it was found that CD94 has
both a distal 1A promoter and a proximal 1B promoter.37,38 These promoters generate transcripts
with identical coding regions. In mature NK-cells, the 1B promoter is constitutively active and
pro-duces the predominant transcript, whereas the 1A promoter is activated only by IL-15. The finding
of an IL-15-regulated CD94 1A promoter raises the possibility that CD94 1A might be the
pre-dominant form expressed in immature NK-cells.
In this study, we used laser capture microdissection to isolate IL-15R + lymphoid cells from
the thymus, and we confirmed that these immature NK cells are indeed CD94 1A+. We also studied
a group of LBL cases that were positive for cytoplasmic CD3 and nuclear TdT and distinguished
two immunophenotypic groups. One group that we propose is of immature NK-cell origin is
7
contrast, the second group had a CD94 1A immunophenotype and carries TCR gene
rearrange-ments supporting T-cell lineage. The identification of a subset of LBL cases of immature NK-cell
8
Materials and Methods
Tissue samples
The study was approved by the Institutional Review Board and informed consent was obtained
according to the Declaration of Helsinki. Twenty-one LBL samples were collected. Diagnoses were
made by histologic and immunophenotypic examination of lymph nodes or mediastinal biopsy
specimens in the Pathology Department of the National Taiwan University Hospital between 1993
and 2001. All diagnoses were established on pre-treatment biopsy specimens at the time the
pa-tients presented initially. Histologically, all specimens showed a characteristic diffuse infiltration of
lymphoblasts positive for cytoplasmic CD3 and nuclear TdT and were negative for CD56 and
CD20. All neoplasms were negative for Epstein-Barr virus (EBV) small-encoded small RNA
(EBER), and thus they were not associated with EBV infection. Clinical data were obtained from
the medical records.
For comparison, other specimens were examined including 10 cases of extranodal nasal-type
NK/T-cell lymphoma as defined according to the criteria specified in the WHO classification; 6
samples of the thymus obtained from thymectomy specimens of 2 adults who had myasthenia
gra-vis, and from autopsy specimens of 4 infants who died of prematurity; 3 spleens obtained removed
for cirrhosis and gastric cancer; 2 samples of endometrium obtained from curettage specimens for
dysfunctional uterine bleeding; 1 tonsil excised for chronic tonsillitis; and 4 peripheral blood
specimens obtained from volunteer normal donors.
Also studied were 4 samples of umbilical cord blood obtained from healthy newborns
9
were isolated by density gradient centrifugation with use of Ficoll-Paque at a density of 1.077.
CD34+ progenitor cells were then purified by an immunomagnetic cell separation system using
multisort microbeads conjugated to a monoclonal mouse anti-human CD34 antibody (QBENDD/10,
Miltenyi Biotec, Auburn, CA). The beads were dissociated from the purified CD34+ progenitors
after enzymatic release. The CD34+ progenitor cells were further incubated with a mixture of
bio-tin-conjugated antibodies against lineage (Lin) markers, including CD2 (clone LT2), CD3 (clone
UCHT-1), CD14 (clone UCHM-1), CD16 (clone LNK16), CD19 (clone LT19), CD33 (clone
WM53), CD41 (clone PM6/248), CD56 (clone MEM188), and glycophorin (clone AC107.3). All
antibodies were obtained from Serotec (Raleigh, NC), except for CD3 from BD Biosciences (San
Jose, CA) and glycophorin from Miltenyi Biotec (Auburn, CA). The CD34+Lin+ progenitors and
CD34+Lin progenitors were separated by streptavidin-conjugated microbeads. Flow cytometry
was used to check the purity of the cells. For flow cytometry, the CD34+Lin progenitors or
CD34+Lin+ progenitors were labeled with a PC5-conjugated CD34 antibody (clone 581, Coulter
Immunotech, Marseille, France).
CD94 transcripts by RT-PCR
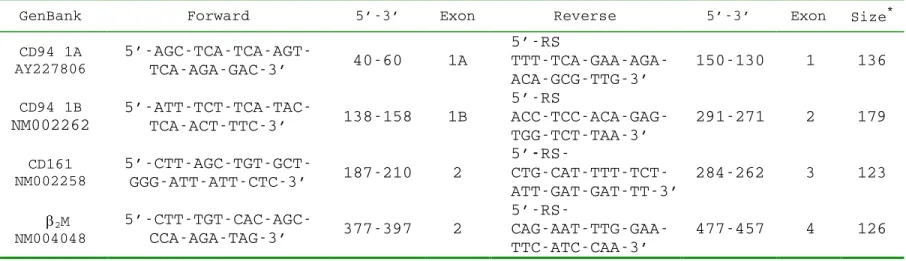
The sequences of the primers used for reverse transcription-polymerase chain reaction
(RT-PCR) analyses are listed in Table 1. The table also provides the GenBank accession number of the
CD94 cDNA, the positions of the primers, and the sizes of the PCR products. The “RS” in Table 1
is a random sequence, 5´-TGACAAACTGTGTTCACTAGC-3´, used for increased PCR specificity
and incorporation of fluorescent labels.
Reverse transcription was performed in a 20-µl mixture at 40o
10
included 0.1 µg of RNA purified from formalin-fixed, paraffin-embedded tissue blocks, 0.5 µM antisense primer, 200 units of reverse transcriptase, 50 mM Tris-HCl at pH 8.3, 75 mM KCl, 3 mM
MgCl2, 10 mM DTT, and 200 µM of each dNTP. The cDNA from 2 µl of the reverse transcription
mixture was amplified with the forward primer and a fluorescence-labeled primer of the random
sequence 5´-tet-CTTCTGACAAACTGTGTTCACTAGC-3´ in a standard PCR reaction.
PCR was done in a 20µl mixture including the DNA template, 0.3 µM of each primer, 15 mM Tris-HCl at pH 8.0, 1.5 mM MgCl2, 50 mM KCl, 200 µM of each dNTP, and 1 unit of Taq
poly-merase. The reaction mixture was subjected to 35 cycles of PCR. Each cycle consisted of
denatura-tion at 94oC for 45 seconds, annealing at 45oC for 45 seconds, and extension at 72oC for 45 seconds.
At the end of 35 cycles, a portion of the PCR products was separated by a high-resolution
electrophoretic system (ABI377 with GeneScan software, Perkin-Elmer, Foster City, CA). The
sizes of the RT-PCR products of CD94 are listed in Table 1. A reaction for beta-2-microglobulin
( 2M) was run as a positive control, and a reaction under the same conditions except for the
omis-sion of reverse transcriptase was run as a negative control. The RT-PCR is deemed positive if its
peak is higher than 10% of 2M.
Killer immunoglobulin-like receptor repertoire analysis
This analysis was done according to a protocol developed in our laboratory.39 Briefly, killer
immunoglobulin-like receptors (KIRs) can be categorized in one of three splicing groups,
KIR2DL4, KIR2D, or KIR3D. Three pairs of group-specific primers were designed and used in
RT-PCR reactions to evaluate the presence or absence of the KIR transcripts. The primers were 5´
11
2DL4, 5´-TGGGTGGGCCAGGAGGAAGGTTT-3´ and 5´-CATGGCGTGTGTTGGGTTCTTC
TTG-3´ for KIR2D, and 5´-TGGGTGGGCCAGGAGGAAGGTTT-3´ and 5´-CACTCCCCCCACT
GGGTGGTCGGC-3´ for KIR3D.
T-cell-receptor rearrangement
Standard PCR was performed to assess for T-cell receptor (TCR) chain gene rearrangements
according to a protocol developed in our laboratory.40 A monoclonal TCR rearrangement is
consid-ered to be present if a dominant clone, making up more than 30% of the total clones, is found.
CD161 transcripts by RT-PCR
RT-PCR for CD161 was performed using the primers listed in Table 1. The reaction
condi-tions were identical to those for CD94. The RT-PCR result is considered to be positive if the
ampli-fied peak is greater than 20% of 2M.
Immunohistochemistry
Immunohistochemical stains for CD44, E2A, ID2, and TCF-1 were performed on
formalin-fixed, paraffin-embedded tissue sections of all 21 LBL cases. The antibodies used were CD44
(clone 2C5, R&D Systems, Minneapolis, MN), E2A (polyclonal, Santa Cruz Biotech, Santa Cruz,
CA), ID2 (polyclonal, Santa Cruz Biotech, Santa Cruz, CA), and TCF-1 (7H3, Exalpha, Watertown,
MA). The staining was done using conditions recommended by the vendor.
12
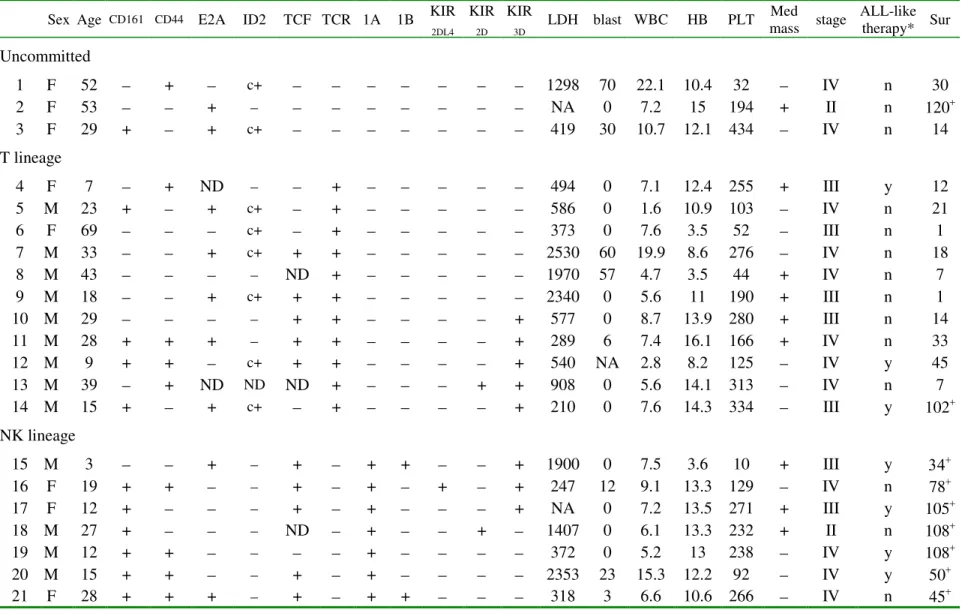
The 21 LBLs were divided into three groups, according tothe patterns of CD94 transcripts and
TCR gene rearrangement. The T-cell lineage was characterized by the presence of monoclonal
TCR gene rearrangement. The NK-cell lineage was characterized by the presence of CD94 1A
tran-scripts without monoclonal TCR gene rearrangement. The uncommitted cases had neither a
mono-clonal TCR gene rearrangement nor CD94 1A transcripts.
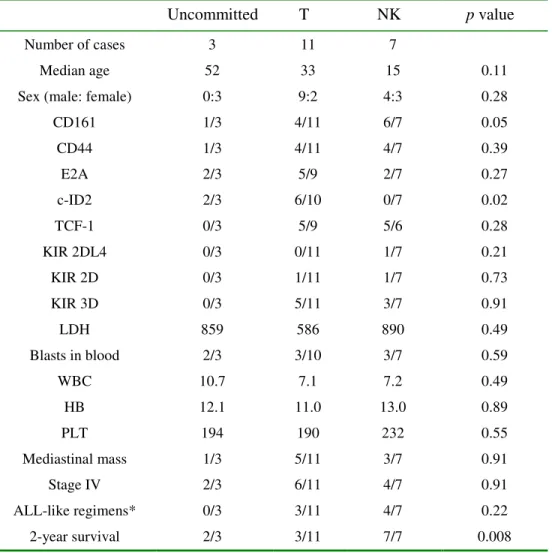
Comparison between LBL of T- and the NK-cell lineage
The uncommitted group was excluded from this comparison because of the small sample size
(n=3). Two-sample comparisons between the LBLs of T- and NK-cell lineage were performed with
the binomial test for categorical data and the non-parametric Mann-Whitney U-test for continuous
data.
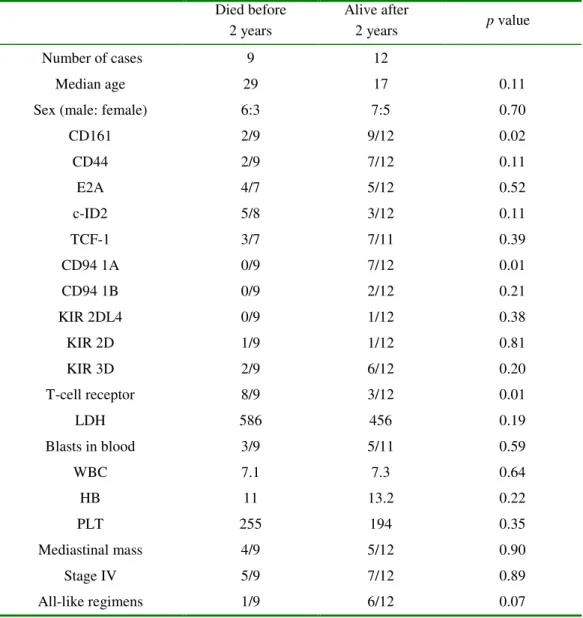
Correlation with 2-year survival
The 21 LBLs were divided into two groups according to the 2-year survival. Two-sample
comparisons were done with the binomial test for categorical data and with the non-parametric
Mann-Whitney U-test for continuous data.
Isolation of IL-15R + immature NK cells from thymus by reverse laser capture microdissec-tion
IL-15R + lymphoid cells, stained by a polyclonal goat antibody (R&D Systems, Minneapolis,
MN), were isolated from 5 µm frozen sections of the thymus of infants who died of prematurity. In conventional laser capture microdissection (LCM), the laser hits the target cells directly, thereby
13
circumvent these difficulties, we used a reverse LCM (rLCM) that removed IL-15R cells from
the slides. This approach preserved the original histopathology of the intact IL-15R + cells and
14
Results
Immature NK-cells from thymus are predominantly CD94 1A+, but mature NK-cells from
pe-ripheral tissues or extranodal nasal-type NK/T-cell lymphomas are predominantly CD94 1B+
We evaluated the distribution patterns of CD94 transcripts in benign and neoplastic tissues
known to have either mature or immature NK-cells, such as umbilical cord blood, peripheral blood,
tonsil, spleen, endometrium, thymus, and extranodal nasal-type NK/T-cell lymphomas (Table 2).
Umbilical cord progenitor cells, either CD34+Lin+ or CD34+Lin , had neither CD94 1A nor
CD94 1B transcripts. In 10 samples from peripheral blood, tonsil, spleen, and endometrium, 9 had
predominantly CD94 1B transcripts, including 2 samples with only CD94 1B transcripts and 7
samples in which CD94 1B transcripts were much more numerous than CD94 1A. Only 1 of these
samples, a spleen from a cirrhotic patient with hypersplenism and pancytopenia, showed only
CD94 1A transcripts. In 10 extranodal nasal-type NK/T-cell lymphomas assessed, CD94 transcripts
were identified in 7 neoplasms, and in 6 tumors CD94 1B transcripts were more numerous than
CD94 1A transcripts. By contrast, all 6 samples of thymus had predominantly CD94 1A transcripts,
including 5 with only CD94 1A transcripts and one in which CD94 1A transcripts were more
nu-merous than CD94 1B transcripts. Representative patterns of CD94 transcripts were shown in
Fig-ure 1.
These data suggest that immature NK-cells use predominantly CD94 1A transcripts, whereas
mature NK-cells use predominantly CD94 1B transcripts. This pattern also indicates that CD94 1A
transcripts can be used to identify LBLs of the NK lineage.
15 but not KIRs identify an NK-LBL
We used the presence of TCR gene rearrangements or expression of CD94 1A transcripts as
criteria for T-cell or NK-cell lineage commitment, respectively (Table 3). According to this scheme,
11 LBLs carried monoclonal TCR gene rearrangements and were classified as T-LBL, and 7 LBLs
had CD94 1A transcripts and were classified as NK-LBL. The remaining 3 cases had neither TCR
gene rearrangement nor CD94 1A transcripts and were considered to be of uncommitted lineage.
The T-LBLs were negative for CD94 1A and frequently expressed cytoplasmic ID2, whereas the
NK-LBLs were positive for CD94 1A and negative for ID2.
The immunohistochemical patterns of ID2 and E2A are shown in Figure 2. E2A exhibited
nu-clear staining in both normal thymus and T-LBL (A & B). For ID2, there is nunu-clear staining in the
normal thymus, but cytoplasmic staining in T-LBL (C and D).
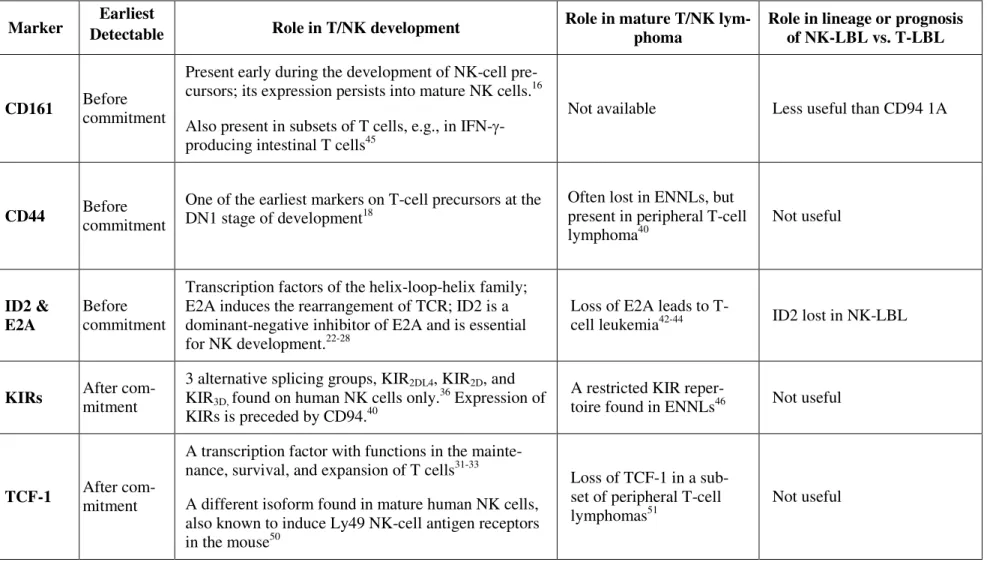
As shown in Table 4, except for CD161 which had borderline significance at p=0.05, the pres-ence or abspres-ence of CD44, E2A, TCF-1, and KIRs was not lineage-specific and did not correlate
with the lineage assignment based on the presence of CD94 1A transcripts. In particular, the
differ-ences in the distributions of KIR2DL4, KIR2D, and KIR3D between the T- and NK-cell lineages,
useful in lineage assignment in mature NK-cell lymphomas, were not significant statistically.
NK-LBL had a better 2-year survival rate than did T-LBL
Patients with NK-LBLs (CD 94 1A positive) had a better prognosis than did those with T-LBLs
(2-year survival, 100% vs 27%, p< 0.01, Table 4). NK-LBL patients also tended to be younger (15
yr vs 33 yr, p=0.11, Table 4), but, because of the small sample size, a firm conclusion cannot be drawn regarding the age difference. Lineage assignment based on antigen receptors, using the
pres-16
ence of CD94 1A or TCR gene rearrangement, therefore, had both biological and clinical
signifi-cance.
As a confirmatory test (Table 5), we divided the LBL patients into two groups, depending on
whether or not the patients survived beyond 2 years. Among various clinical and biological
parame-ters tested, we found that 2-year survival correlated most strongly with presence of CD94 1A
tran-scripts and absence of TCR gene rearrangement (p=0.01).
CD94 1A transcripts could be detected in immature IL-15R + lymphoid cells from thymus
The existence of a bipotent T/NK precursor in the thymus is well known, but has not been
well characterized histologically. Immunohistochemistry on the thymus from an infant of 29-week
gestational age who died of prematurity showed either focal aggregates of IL-15R + lymphoid cells
(Figure 3, A&B) or scattered interstitial staining (Figure 3, C&D). We isolated these cells by rLCM,
17
Discussion
In this study, we have attempted to improve the classification of LBLs by recognizing a subset
of neoplasms that are of immature NK-cell lineage. We started with 21 LBLs, all of them positive
for cytoplasmic CD3 and nuclear TdT, and negative for CD56. Because CD3 is part of the TCR
complex and TdT is the enzyme that creates junctional diversity during TCR gene rearrangement, it
is not surprising that immature T- or T/NK-cell precursors express cytoplasmic CD3 and TdT. It
should be noted, however, that neither cytoplasmic CD3 nor TdT implies commitment to T-cell
lineage. For example, immature or fetal NK-cells also express cytoplasmic CD3,6 and TdT can be
detected in the earliest Lin /Sca-1+/ILR7 /C-Kithi/TdT+ common lymphoid precursors.7 Therefore, the cytoplasmic CD3 and nuclear TdT immunophenotype is not lineage specific.
With the use of CD94 1A expression and TCR gene rearrangement, our data support a
reas-sessment of lineage assignment of LBLs, based on recent advances in the developmental biology of
and NK-cells, both of which are derived from a common T/NK-cell thymic precursor. For the
T-cell lineage,30 development proceeds from a bipotent T/NK-cell precursor through T-cell precursors
double negative (DN) stages 1 through 4, to CD4/CD8 double-positive immature T-cells, and
fi-nally to CD4 or CD8 single positive mature T-cells. The process of lineage commitment takes place
in CD44+/CD25+ T-cell precursors at stages DN2/DN3, characterized by TCR gene
rearrange-ment that is promoted by the transcription factor E2A. For the NK-cell lineage, developrearrange-ment
de-pends on IL-1520,21 and the transcription factor ID2.22,23 One of the earliest surface markers
associ-ated with the NK-cell lineage is CD161, but the intermediate stages preceding mature NK-cells are
still unknown. It is known only that mature NK-cells use the CD94/NKG2 family and KIRs in
lim-18
ited.
In accordance with these data, we present a model for NK-cell development that can be
inte-grated into the classification of LBLs (Fig 4). In this three-stage model, the bipotent T/NK-cell
pre-cursor develops into a uni-potent prepre-cursor, then to immature stages, and finally to mature cells.
CD94 1A transcripts and TCR rearrangements determine lineage commitment and have clinical
significance. The presence of CD94 1A without monoclonal TCR gene rearrangement identifies
NK-LBL, most of which are negative for cytoplasmic ID2.
The absence of ID2 in NK-LBLs and the presence of cytoplasmic ID2 in T-LBLs is of interest.
One might expect expression of nuclear ID2 to be associated with NK-LBL, and E2A to be
associ-ated with the T-LBL, because these molecules are required for NK- and T-cell development,
re-spectively.27,28 However, quite unexpectedly, we found that cytoplasmic ID2 was associated with
T-LBL, and E2A was not lineage-specific. How the loss of ID2 leads to NK-LBL is unknown, but it is
known that redistribution of ID2 from the nucleus to the cytoplasm is associated with
differentia-tion of murine hematopoietic cells,41 and that loss of E2A can cause T-lymphoblastic leukemia.42-44
Thus, the biologic significance of cytoplasmic ID2 is worthy of further investigation.
As summarized in Table 6, markers associated with precursors, such as CD161 and CD44, or
those associated with mature cells, such as TCF-1 and KIRs, are not useful clinically. The lack of
lineage specificity for CD161 is not totally unexpected, because CD161 is present in subsets of
T-cells, including those residing in the intestines.45 Markers associated with mature T- or NK-cells,
such as TCF-1 and KIRs,31-33,36 are not useful for delineating T- or NK-LBLs that are derived from
immature lymphoblasts. Because KIRs are related to T-cell receptors, and both belong to the
fami-19
lies could be used as criteria for lineage commitment in two developmentally related cellular
line-ages. We found, however, that KIRs are not useful in lineage assignment for LBLs (Table 4). As
KIRs are found only in human NK-cells, lineage commitment probably does not depend on KIRs.
This finding is also consistent with a previous report that CD94 expression precedes KIRs, and that
lineage commitment is independent of KIRs.46
LBL expressing NK cell-associated antigens was first recognized in 1987.47 By screening 38
cases of LBL, the authors found 6 cases expressing the NK-associated antigens CD16 (Leu11b) and
CD57 (Leu7). As a group, these cases of so-called “NK-LBL” were TdT+, CD2 (T11)+, CD4
(Leu3A)+, CD16+, and CD57+, and they had a worse prognosis than that of the T-LBL. However,
the true nature of these neoplasms was never further addressed at the molecular level. Subsequently,
CD56 became a popular marker of NK-cells even though CD56 is not specific and can be expressed
by many other neoplasms. This has led to confusion in the literature regarding neoplasms of
NK-cell lineage. For example, the entity described in the WHO classification as blastic NK-NK-cell
lym-phoma48 is, in fact, not a neoplasm of NK-cells and most likely is derived from type II plasmacytoid
dendritic cells.13For these reasons, we did not consider the absence of CD56 expression in LBLs as
evidence excluding NK-cell lineage, prompting us to search for other markers useful for
distin-guishing immature NK- from T-cells.
In conclusion, we propose that LBLs can be divided into those of NK- and T-cell lineage
us-ing CD94 1A and TCR gene rearrangement. Among 21 cases of LBL that were positive for
cyto-plasmic CD3 and nuclear TdT and negative for CD56, seven cases expressed CD94 1A transcripts
and did not carry monoclonal TCR gene rearrangements supporting immature NK-cell lineage.
20
after appropriate chemotherapy. The use of CD94 1A, instead of CD56, as a marker of NK-cell
lineage reflects recent advances in NK-cell biology and emphasizes the importance of complete
lineage commitment in the biology of NK-cells. Based on the identification of a true NK-LBL, we
present a revised model of NK-cell development. This model not only stresses the importance of
lineage commitment for clinical prognosis, but also implies further constraints on the ultimate
21
References
1. Pilozzi E, Muller-Hermelink HK, Falini B, de Wolf-Peeters C, Fidler C, Gatter K, Wainscoat J:
Gene rearrangement in T-cell lymphoblastic lymphoma. J Pathol 1999, 188:267-270
2. Michie AM, Carlyle JR, Schmitt TM, Ljutic B, Cho SK, Fong Q, Zuniga-Pflucker JC: Clonal
characterization of a bipotent T cell and NK cell progenitor in the mouse fetal thymus. J
Im-munol 2000, 164:1730-1733
3. Ikawa T, Kawamoto H, Fujimoto S, Katsura Y: Commitment of common T/natural killer (NK)
progenitors to unipotent T and NK progenitors in the murine fetal thymus revealed by a single
progenitor assay. J Exp Med 1999, 190:1617-1626
4. Sanchez MJ, Muench MO, Roncarolo MG, Lanier LL, Philips JH: Identification of a common
T/natural killer cell progenitor in human fetal thymus. J Exp Med 1994, 180:569-576
5. Sanchez MJ, Spits H, Lanier LL, Philips JH: Human natural killer cell committed thymocytes
and their relation to the T cell lineage. J Exp Med 1993, 178:1857-1866
6. Philips JH, Hori T, Nagler A, Bhat N, Spits H, Lanier LL: Ontogeny of human natural killer
(NK) cells: fetal NK cells mediate cytolytic function and express cytoplasmic CD3 epsilon,
delta proteins. J Exp Med 1992, 175:1055-1066
7. Melinda K, Garrett KP, Thompson LF, Rossi MID, Payne KJ, Kincade PW: Identification of
very early lymphoid precursors in bone marrow and their regulation by estrogen. Nature
Im-munol 2001, 2:718-724
8. Adriaansen HJ, Soeting PW, Wolvers-Tettero IL, van Dongen JJ: Immunoglobulin and T-cell
receptor gene rearrangements in acute non-lymphocytic leukemias. Analysis of 54 cases and a
22
9. Nakamura F, Tatsumi E, Kawano S, Tani A, Kumagai S, Nishikori M, Nagai T: Acute
lym-phoblastic leukemia/lymlym-phoblastic lymphoma of natural killer (NK) lineage: quest for another
NK-lineage neoplasm. Blood 1997, 89:4665-4666
10. Emile JF, Boulland ML, Haioun C, Kanavaros P, Petrella T, Delfau-Larue MH, Bensussan A,
Farcet JP, Gaulard P: CD5 CD56+ T-cell receptor silent peripheral T-cell lymphomas are
natu-ral killer cell lymphomas. Blood 1996, 87:1466-1473
11. Chaperot L, Bendriss N, Manches O, Gressin R, Maynadie M, Trimoreau F, Orfeuvre H,
Cor-ront B, Feuillard J, Sotto JJ, Bensa JC, Briere F, Plumas J, Jacob MC: Identification of a
leu-kemic counterpart of the plasmacytoid dendritic cells. Blood 2001, 97:3210-3217
12. Feuillard J, Jacob MC, Valensi F, Maynadie M, Gressin R, Chaperot L, Arnoulet C,
Brignole-Baudouin F, Drenou B, Duchayne E, Falkenrodt A, Garand R, Homolle E, Husson B, Kuhlein
E, Le Calvez G, Sainty D, Sotto MF, Trimoreau F, Bene MC: Clinical and biologic features of
CD4(+)CD56(+) malignancies. Blood 2002, 99:1556-1563
13. Leroux D, Mugneret F, Callanan M, Radford-Weiss I, Dastgue N, Feuillard J, Le Mee F,
Plessis G, Talmant P, Gachard N, Uettwiller F, Pages MP, Mozziconacci MJ, Eclache V, Sibille
C, Avet-Loiseau H, Lafage-Pochitaloff M: CD4(+), CD56(+) DC2 acute leukemia is
character-ized by recurrent clonal chromosomal changes affecting 6 major targets: a study of 21 cases by
the Groupe Francais de Cytogenetique Hematologique. Blood 2002, 99:4154-4159
14. Giorda R, Rudert WA, Vavassori C, Chambers WH, Hiserodt JC, Trucco M: NKR-P1, a signal
transduction molecule on natural killer cells. Science 1990, 249:1298-1300
15. Giorda R, Weisberg EP, IP TK, Trucco M: Genomic structure and strain-specific expression of
the natural killer cell receptor NKR-P1. J Immunol 1992, 149:1957-1963
16. Yokoyama WM, Seaman WE: The Ly-49 and NKR-P1 gene families encoding lectin-like
23
17. Khalturin K, Becker M, Rinkevich B, Bosch TC: Urochordates and the origin of natural killer
cells: identification of a CD94/NKR-P1-related receptor in blood cells of Botryllus. Proc Natl
Acad Sci USA 2003, 100:622-627
18. Schwarzler C, Oliferenko S, Gunthert U: Variant isoforms of CD44 are required in early
thy-mocyte development. Eur J Immunol 2001, 31:2997-3005
19. Lien HC, Lin CW, Huang PH, Chang ML, Hsu SM: Expression of cyclin-dependent kinase 6
(cdk6) and frequent loss of CD44 in nasal-nasopharyngeal NK/T-cell lymphomas: comparison
with CD56-negative peripheral T-cell lymphomas. Lab Invest 2000, 80:893-900
20. Liu CC, Perussia B, Young JD: The emerging role of IL-15 in NK-cell development. Immunol
Today 2000, 21:113-116
21. Mrozek E, Anderson P, Caligiuri MA: Role of interleukin-15 in the development of human
CD56+ natural killer cells from CD34+ hematopoietic progenitor cells. Blood 1996,
87:2632-2640
22. Yokota Y, Mansouri A, Mori S, Sugawara S, Adachi S, Nishikawa S, Gruss P: Development of
peripheral lymphoid organs and natural killer cells depends on the helix-loop-helix inhibitor
Id2. Nature 1999, 397:702-706
23. Ikawa T, Fujimoto S, Kawamoto H, Katsura Y, Yokota Y: Commitment to natural killer cells
requires the helix-loop-helix inhibitor Id2. Proc Natl Acad Sci USA 2001, 98:5164-5169
24. Leclercq G, Debacker V, de Smedt M, Plum J: Differential effects of interleukin-15 and
inter-leukin-2 on differentiation of bipotential T/natural killer progenitor cells. J Exp Med 1996,
184:325-336
25. Greenbaum S, Zhuang Y: Regulation of early lymphocyte development by E2A family proteins.
24
26. Langerak AW, Wolvers-Tettero IL, van Gastel-Mol EJ, Oud ME, van Dongen JJ: Basic
helix-loop-helix proteins E2A and HEB induce immature T-cell receptor rearrangements in
nonlym-phoid cells. Blood 2001, 98: 2456-2465
27. Morrow MA, Mayer EW, Perez CA, Adlam M, Siu G: Overexpression of the
Helix-Loop-Helix protein Id2 blocks T cell development at multiple stages. Mol Immunol 1999,
36:491-503
28. Engel I, Murre C: The function of E- and ID proteins in lymphocyte development. Nat Rev
Immunol 2001, 1:193-199
29. DiSanto JP: Cytokines: shared receptors, distinct functions. Curr Biol 1997, 7:R424-426
30. Rothenberg EV: T-lineage specification and commitment: a gene regulation perspective.
Semi-nar Immunol 2002, 14:431-440
31. Schilham MW, Wilson A, Moerer P, Benaissa-Trouw BJ, Cumano A, Clevers HC: Critical
in-volvement of Tcf-1 in expansion of thymocytes. Immunol 1998, 161:3984-3991
32. Ioannidis V, Beermann F, Clevers H, Held W: The B-catenin-TCF-1 pathway ensures
CD4+CD8+ thymocyte survival. Nature Immunol 2001, 2:691-697
33. Toor AA, Lund TC, Miller JS: T-cell factor-1 expression during human natural killer cell
de-velopment and in circulating CD56(+) bright natural killer cells. Exp Hematol 2001,
29:499-506
34. Chang C, Rodriguez A, Carretero M, Lopez-Botet M, Phillips JH, Lanier LL: Molecular
char-acterization of human CD94: a type II membrane glycoprotein related to the C-type lectin
su-perfamily. Eur J Immunol 1995, 9:2433-2437
35. Lopez-Botet M, Perez-Villar JJ, Carreterro M, Rodriguez A, Melero I, Bellon T, Liano M,
rec-25
ognition of HLA class I molecules. Immunol Rev 1997, 155:165-174
36. Long EO, Rajagopalan S: HLA class I recognition by killer cell Ig-like receptors. Semin
Im-munol 2000, 12:101-108
37. Lieto LD, Borrego F, You CH, Coligan JE: Human CD94 gene expression: Dual Promoters
dif-fering in responsiveness to IL-2 and IL-15. J Immunol 2003, 171:5277-5286
38. Wilhelm BT, Landry JR, Takei F, Mager DL: Transcriptional control of murine CD94 gene:
differential usage of dual promoters by lymphoid cell types. J Immunol 2003, 171:4219-4226
39. Lin CW, Lee WH, Chang CL, Yang JY, Hsu SM: Restricted killer cell immunoglobulin-like
receptor repertoire without T-cell receptor gamma rearrangement supports a true natural
killer-cell lineage in a subset of sinonasal lymphomas. Am J Pathol 2001, 159:1671-1679
40. Lin CW, Yang JY, Chuang YC, Chen YH, Albitar M, Hsu SM: Presence of restricted killer
im-munoglobulin-like receptor repertoire and monoclonal T-cell receptor gamma rearrangement as
evidence of mixed NK/T-cell differentiation in a subset of sinonasal lymphomas. Lab Invest
2003, 83:55-64
41. Tu X, Baffa R, Luke S, Prisco M, Baserga R: Intracellular redistribution of nuclear and
nucleo-lar proteins during differentiation of 32D murine hemopoietic cells. Exp Cell Res 2003,
288:119-130
42. Engel I, Murre C: Disruption of pre-TCR expression accelerates lymphomagenesis in
E2A-deficient mice. PNAS 2002, 99:11322-11327
43. Park ST, Nolan GP, Sun XH: Growth inhibition and apoptosis due to restoration of E2A
activ-ity in T cell acute lymphoblastic leukemia cells. J Exp Med 1999, 189:501-508
44. Hunger, SP: Chromosomal translocations involving the E2A gene in acute lymphoblastic
26
45. O’Keeffe J, Doherty DG, Kernna T, Sheahan K, O’Donoghue DP, Hyland JM, O’Farrelly C:
Diverse populations of T cells with NK cell receptors accumulate in the human intestine in
health and in colorectal cancer. Eur J Immunol 2004, 34:2110-2119
46. Miller JS, McCullar V: Human natural killer cells with polyclonal lectin and
immunoglobu-linlike receptors develop from single hematopoietic stem cells with preferential expression of
NKG2A and KIR2DL2/L3/S2. Blood 2001, 98:705-713
47. Sheibani K, Winberg CD, Burke JS, Nathwani BN, Blayney DW, Van de Velde S, Swartz WG,
Rappaport H: Lymphoblastic lymphoma expressing natural killer cell-associated antigens: a
clinicopathologic study of six cases. Leuk Res 1987, 11:371-377
48. Chan JKC, Jaffe ES, Ralfkiaer E. Blastic NK-cell lymphoma. In: Jaffe ES, Harris NL, Stein H,
Vardiman JW (eds). World Health Organization classification of tumours. Pathology and
genet-ics of haematopoietic and lymphoid tissues. IARC Press: Lyon 2001. 214-215
49. Yokoyama WM, Kim S, French AR: The dynamic life of natural killer cells. Annu Rev
Immu-nol 2004, 22:405-429
50. Held W. Clevers H, Grosschedl R: Redundant functions of TCF-1 and LEF-1 during T and NK
cell development, but unique role of TCF-1 for Ly49 NK cell receptor acquisition. Eur J
Im-munol 2003, 33:1393-1398
51. Dorfman DM, Greisman HA, Shahsafaei A: Loss of expression of the
WNT/beta-catenin-signaling pathway transcription factors lymphoid enhancer factor-1 (LEF-1) and T cell factor-1
27
Acknowledgments
Research funding was provided through Grants NTUH 94-S001 and NSC93- 2320-B002-077 to
CW Lin, and NSC-93-2314-B-002-005 and NSC-93-3112-B-041-Y to SM Hsu, from the National
Taiwan University Hospital and the National Science Council, Taiwan. We thank Mr. C.C. Lu for
the artwork, and the National Taiwan University Center for Genomic Medicine for technical
28
Figure legends
Figure 1. High-resolution electrophoretic tracings of mature and immature CD94 patterns. From the top to the bottom panels, two distinct patterns of CD94 transcripts are shown. The
imma-ture CD94 1A pattern, either CD94 1A alone or CD94 1A>> CD94 1B, was found in the thymus or LBLs. The mature CD94 1B pattern, either CD94 1B alone or CD94 1A <<1B, was found in pe-ripheral tissues or extranodal nasal-type NK/T-cell lymphomas. (X-axis: in base pairs; Y-axis:
in-tensity of the fluorescence.)
Figure 2. Immunohistochemistry for E2A and ID2 from LBLs and thymus of premature in-fants. Note the nuclear E2A stain in both the thymus (A) and a T-LBL (B). For ID2, the staining is nuclear in the normal thymus (C), but cytoplasmic in a case of T-LBL (D). (A-D, 400X,
immuno-histochemistry using formalin-fixed, paraffin-embedded tissue sections and hematoxylin
counter-stain.)
Figure 3. IL-15R + immature lymphoid cells isolated from thymus of a premature infant by reverse laser capture microdissection. The figure shows a nodular pattern of IL-15R +
lymphoid
cells in the thymus before and after reverse laser capture microdissection (rLCM) (A & B, with
hematoxylin counterstain), and an interstitial pattern of IL-15R +lymphoid cells in the thymus
be-fore and after rLCM (C & D, phase-contrast view without hematoxylin counterstain). Note the
well-preserved histopathology of IL-15R + lymphoid cells isolated from a 5 µm frozen section of thymus with use of rLCM.
29
T/NK-cell precursor may develop into either immature T-cells under the actions of E2A and IL-2,
or immature NK-cells under the actions of ID2 and IL-15. Lineage commitment is fully established
when immature T-cells complete TCR gene rearrangement, or, as our data support, immature
NK-cells express CD94 1A transcripts. The model shows where the cytokines and transcription factors
are critical, but does not exclude their possible roles at other developmental stages. For example,
the assignment of ID2 before IL-15 is based on previous data23 and a recent review.49According to
this model, the loss of ID2 in NK-LBLs is due to abnormal differentiation. Whether the expression
of IL-15R or, equivalently, the role of IL-15, might precede that of ID2 in the development of
30 Figure 1
31 Figure 2
32 Figure 3
33 Figure 4
34
Table 1. Primers for CD94 and CD161
GenBank Forward 5’-3’ Exon Reverse 5’-3’ Exon Size*
CD94 1A AY227806 5’-AGC-TCA-TCA-AGT-TCA-AGA-GAC-3’ 40-60 1A 5’-RS TTT-TCA-GAA-AGA-ACA-GCG-TTG-3’ 150-130 1 136 CD94 1B NM002262 5’-ATT-TCT-TCA-TAC-TCA-ACT-TTC-3’ 138-158 1B 5’-RS ACC-TCC-ACA-GAG-TGG-TCT-TAA-3’ 291-271 2 179 CD161 NM002258 5’-CTT-AGC-TGT-GCT-GGG-ATT-ATT-CTC-3’ 187-210 2 5’ -RS- CTG-CAT-TTT-TCT-ATT-GAT-GAT-TT-3’ 284-262 3 123 2M NM004048 5’-CTT-TGT-CAC-AGC-CCA-AGA-TAG-3’ 377-397 2 5’-RS- CAG-AAT-TTG-GAA-TTC-ATC-CAA-3’ 477-457 4 126 RS=5’-TGA-CAA-ACT-GTG-TTC-ACT-AGC-3’ *Size: Size of the RT-PCR product in base pairs
35
Table 2. Tissue distribution of CD94 transcripts
CD34 progenitors and thymus Peripheral tissues ENNLs
Age Sex Tissue Diagnosis CD94 Age Sex Tissue Diagnosis CD94 Age Sex Tissue Diagnosis CD94 1 0 d M CD34
+
Lin+ UC progenitors
normal
newborn neg 1 44 M spleen
cirrhosis with hypersplenism &
pancytopenia
1A 1 60 M nasal cavity ENNL neg 2 0 d M CD34
+Lin
UC progenitors
same
case above neg 2 32 F
endo-metrium
reactive
endometrium 1B 2 65 M nasal cavity ENNL neg 3 0 d F CD34
+
Lin+ UC progenitors
normal
newborn neg 3 75 M spleen
gastric cancer without splenic
involvement
1B 3 45 F nasal cavity ENNL neg 4 0 d F CD34
+Lin
UC progenitors
same
case above neg 4 45 F
endo-metrium
reactive
endometrium 1B>>1A 4 61 M nasal cavity ENNL 1A 5 1 d F thymus premature infant
(GA: 29 weeks) 1A 5 74 M spleen
cirrhosis with
splenomegaly 1B>>1A 5 42 M nasal cavity ENNL 1B>>1A 6 1 d F thymus premature infant
(GA: 37 weeks) 1A 6 65 M tonsil
chronic
tonsillitis 1B>>1A 6 62 M nasal cavity ENNL 1B>>1A 7 1 d M thymus premature infant
(GA: 27 weeks) 1A 7 43 M PB
normal
volunteer 1B>>1A 7 55 M nasal cavity ENNL 1B>>1A 8 1 d M thymus premature infant
(GA: 27 weeks) 1A 8 24 M PB
normal
volunteer 1B>>1A 8 39 F
nasal cavity ENNL 1B>>1A
9 40 F thymus MG with LH 1A 9 28 F PB normal
volunteer 1B>>1A 9 69 M nasal cavity ENNL 1B>>1A 10 17 F thymus MG with LH 1A>>1B 10 27 F PB normal
volunteer 1B>>1A 10 38 M nasal cavity ENNL 1B>>1A Abbreviations: d: day; ENNL: extranodal nasal-type NK/T-cell lymphoma; F: female; GA: gestational age; Lin: lineage; M: male; MG with LH: myasthenia gravis with
36
Table 3. Pathologic and clinical data for lymphoblastic lymphomas
Sex Age CD161 CD44 E2A ID2 TCF TCR 1A 1B KIR
2DL4 KIR 2D KIR 3D LDH blast WBC HB PLT Med mass stage ALL-like therapy* Sur Uncommitted 1 F 52 + c+ 1298 70 22.1 10.4 32 IV n 30 2 F 53 + NA 0 7.2 15 194 + II n 120+ 3 F 29 + + c+ 419 30 10.7 12.1 434 IV n 14 T lineage 4 F 7 + ND + 494 0 7.1 12.4 255 + III y 12 5 M 23 + + c+ + 586 0 1.6 10.9 103 IV n 21 6 F 69 c+ + 373 0 7.6 3.5 52 III n 1 7 M 33 + c+ + + 2530 60 19.9 8.6 276 IV n 18 8 M 43 ND + 1970 57 4.7 3.5 44 + IV n 7 9 M 18 + c+ + + 2340 0 5.6 11 190 + III n 1 10 M 29 + + + 577 0 8.7 13.9 280 + III n 14 11 M 28 + + + + + + 289 6 7.4 16.1 166 + IV n 33 12 M 9 + + c+ + + + 540 NA 2.8 8.2 125 IV y 45 13 M 39 + ND ND ND + + + 908 0 5.6 14.1 313 IV n 7 14 M 15 + + c+ + + 210 0 7.6 14.3 334 III y 102+ NK lineage 15 M 3 + + + + + 1900 0 7.5 3.6 10 + III y 34+ 16 F 19 + + + + + + 247 12 9.1 13.3 129 IV n 78+ 17 F 12 + + + + NA 0 7.2 13.5 271 + III y 105+ 18 M 27 + ND + + 1407 0 6.1 13.3 232 + II n 108+ 19 M 12 + + + 372 0 5.2 13 238 IV y 108+ 20 M 15 + + + + 2353 23 15.3 12.2 92 IV y 50+ 21 F 28 + + + + + + 318 3 6.6 10.6 266 IV n 45+
Abbreviations: 1A: CD94 1A; 1B: CD94 1B; age in years; blast: percentage of blasts in peripheral blood; c: cytoplasmic; F: female; HB: hemoglobin in g/dl; KIR: killer immunoglobulin-like receptor; LDH: lactate dehydrogenase, IU/L; M: male; Med: mediastinal; n: no; NA: not available; ND: not determined; PLT: platelets, 103/µl; Sur: survival in months, where a + sign indicates a censored case; TCF: T-cell factor; TCR: T-cell receptor; WBC: white blood cells, 103/µl; y: yes. * The patients were treated with either ALL-like or CHOP-based regimens.
37
Table 4. Lineage assignment according to CD94 1A and TCR rearrangement
Abbreviations: age in years; c-ID2: cytoplasmic ID2; HB: hemoglobin in g/dl; KIR: killer immunoglobulin-like receptor; LDH: lactate dehydrogenase IU/L; PLT: platelets 103/µl; TCF-1: T-cell factor; WBC: white blood cells, 103/µl.
* The patients were treated with either ALL-like or CHOP-based regimens.
Uncommitted T NK p value
Number of cases 3 11 7
Median age 52 33 15 0.11
Sex (male: female) 0:3 9:2 4:3 0.28
CD161 1/3 4/11 6/7 0.05 CD44 1/3 4/11 4/7 0.39 E2A 2/3 5/9 2/7 0.27 c-ID2 2/3 6/10 0/7 0.02 TCF-1 0/3 5/9 5/6 0.28 KIR 2DL4 0/3 0/11 1/7 0.21 KIR 2D 0/3 1/11 1/7 0.73 KIR 3D 0/3 5/11 3/7 0.91 LDH 859 586 890 0.49 Blasts in blood 2/3 3/10 3/7 0.59 WBC 10.7 7.1 7.2 0.49 HB 12.1 11.0 13.0 0.89 PLT 194 190 232 0.55 Mediastinal mass 1/3 5/11 3/7 0.91 Stage IV 2/3 6/11 4/7 0.91 ALL-like regimens* 0/3 3/11 4/7 0.22 2-year survival 2/3 3/11 7/7 0.008
38
Table 5. 2-year survival correlates with CD94 1A & TCR rearrangement
See Table 4 for abbreviations.
Died before 2 years Alive after 2 years p value Number of cases 9 12 Median age 29 17 0.11
Sex (male: female) 6:3 7:5 0.70
CD161 2/9 9/12 0.02 CD44 2/9 7/12 0.11 E2A 4/7 5/12 0.52 c-ID2 5/8 3/12 0.11 TCF-1 3/7 7/11 0.39 CD94 1A 0/9 7/12 0.01 CD94 1B 0/9 2/12 0.21 KIR 2DL4 0/9 1/12 0.38 KIR 2D 1/9 1/12 0.81 KIR 3D 2/9 6/12 0.20 T-cell receptor 8/9 3/12 0.01 LDH 586 456 0.19 Blasts in blood 3/9 5/11 0.59 WBC 7.1 7.3 0.64 HB 11 13.2 0.22 PLT 255 194 0.35 Mediastinal mass 4/9 5/12 0.90 Stage IV 5/9 7/12 0.89 All-like regimens 1/9 6/12 0.07