行政院國家科學委員會專題研究計畫 成果報告
利利利利利利利利利利利利利利利/核酸 Aptamer 之研究
計畫類別: 個別型計畫
計畫編號: NSC93-2113-M-009-012-
執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日
執行單位: 國立交通大學生物科技研究所
計畫主持人: 吳東昆
計畫參與人員: 張程翔,劉媛婷,虞美婷,葉佳宜,張晉豪,劉妍希,王庭翊,
陳令宗,林宏明
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 94 年 10 月 20 日
行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※
※ ※
利用氧化鯊烯環化酵素作為篩選胜肽/核酸 Aptamer 之研究
※ ※
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:■個別型計畫 □整合型計畫
計畫編號:NSC 93-2113-M-009-012-
執行期間:93 年 08 月 01 日至 94 年 07 月 31 日
計畫主持人:吳東昆
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立交通大學生物科技學系
中 華 民 國 94 年 10 月 20 日
行政院國家科學委員會專題研究計畫成果報告
計畫編號:NSC 93-2113-M-009-012-
利用氧化鯊烯環化酵素作為篩選胜肽/核酸 Aptamer 之研究
執行期限:93 年 08 月 01 日至 94 年 07 月 31 日
主持人:吳東昆 國立交通大學生物科技學系
計畫參與人員:張程翔,劉媛婷,虞美婷,葉佳宜,張晉豪,劉妍希,王庭翊,陳令宗,
林宏明
一、中文摘要 膽固醇生合成途徑中負責關鍵中間環化步驟 反應之酵素-氧化鯊烯環化酵素,由於其在生化及 醫學中之學術重要性與作為發展降低膽固醇及抗 黴菌藥物之重要標的之潛力,而成為學術與藥廠 中研發之重要題目。本研究利用氧化鯊烯環化酵 素作為標的以尋找可與其進行交互作用之核酸片 段與胜肽序列,,希望未來發展成為藥物。核酸 片段之篩選經由經由配位體指數遞增演化之技 術,結合聚合脢連鎖反應與膠體遲滯電泳技術, 經過數個重複篩選、放大、清洗之過程,已得到 數段核酸片段,經由 DNA 定序儀已測定其序列。 另外在胜肽序列之篩選,利用兩種噬菌體胜肽 庫,七個及十二個隨機胜肽的胜肽庫,對氧化鯊 烯環化酵素進行配位基的篩選。經由五次循環之 鍵結、洗滌,亦已獲得五個對氧化鯊烯環化酵素 具有親和性的噬菌體胜肽序列,由西方墨點法測 定得到 1 個七胜肽序列和 4 個十二胜肽序列具有 反應。 關鍵詞:配位體指數遞增演化,氧化鯊烯環化酵 素,羊毛硬脂醇,重複質譜,聚合脢連鎖反應,膠 體遲滯電泳,隨機胜肽庫 AbstractOxidosqualene cyclase fulfills a unique role in designing new antifungal, hypocholesteremic, and phytotoxic drugs, due to its great biological and medicinal significance and potential. In this study, the mechanistic insights and structure-function relationships of oxidosqualene cyclase enzyme is extended and the enzyme is further explored for protein-DNA and protein-peptide interactions study to search for OSC-binding DNA and peptide search. We have applied systematic evolution of ligands by exponential enrichment (SELEX) technology to identify OSC-binding aptamer from a DNA pool of more than 1015 individual nucleic acid molecules. Following the amplification of the selected oligonucleotides by PCR and successive iteration of the selection procedure, oligonucleotides with high binding affinity and specificity were obtained. In
biopanning for OSC binding ligand identification. Following five biopanning process of OSC-binding selection and DNA sequencing of selected phages, five OSC-bound phages were selected. Among them, one heptapeptide and four dodecapeptides, showing OSC-binding affinity were identified from Western blotting experiments.
Keywords: systematic evolution of ligands by exponential enrichment (SELEX), oxidosqualene-lanosterol cyclase, lanosterol, tandem
mass, electrophoresis mobility shift assay (EMSA), random peptide library
二、緣由與目的
The world’s population continues to be seriously plagued by pathogenic organisms, which cause serious health problems. On the other hand, the relationships between increase of coronary heart disease and elevated plasma LDL cholesterol are well established. The development of cholesterol lowering drugs mainly focused on inhibiting HMG-CoA reductase activity will be very beneficial for prevention of this disease. Both cholesterol and fungal sterol biosyntheses share a common biosynthetic pathway until oxidosqualene, the substrate which is catalyzed by oxidosqualene cyclase to form the sterol and related triterpenoid core structure.
Oxidosqualene cyclase enzymes (OSCs) constitute a family of enzymes which catalyze the enzymatic conversion of oxidosqualene into a diverse array of sterols and related triterpenes. The multiple pathways for cyclization, rearrangement and oxidation of (oxido)squalene into structural diverse sterols and triterpenoids represent one of the most remarkable and fascinating biotransformations found in nature. The structure-function relationships and product profiles of (oxido)squalene cyclase enzymes have fascinated both chemists and biochemists for more than half a century because of the complexity of the enzymatic reactions and the diverse products distribution as well as their unique role in designing antifungal, hypocholesterolemic, and phytotoxic drugs.
characterize the putative functional motif, important in
determining substrate recognition, cyclization/rearrangement cascade, or changing product specificity for all cyclases. Several critical amino acid residues (Tyr410Cys, Ala464Val, His477Tyr, Ile481Thr, and Tyr532His derived from oxidosqualene-cycloartenol cyclase (CAS) gene of A.
thaliana) involved in determining product specificity,
especially in the final termination of carbocation between lanosterol and cycloartenol formation, have been identified. The result suggested that a single amino acid change is sufficient to alter the product specificity in the course of evolution. We have also successfully purified OSC from bovine liver to homogeneity as a single band under silver stain determination. These results provide bases for further understanding the biological origins and structure-function relationships of OSC as well as for developing novel OSC inhibitors for antifungal and hypocholesteremic purposes. Moreover, three independent regions on ERG7 of S. cerevisiae and twenty-nine non-alanine residues located on the putative active site surface of CAS from A. thaliana were also mutated and assayed for their ability to complement the cyclase-deficient, ERG7 knockout, S.
cerevisiae strain. Three amino acid residues (Trp443,
Phe445, and Lys448 in yeast ERG7 gene) were also identified using alanine scanning coupled with plasmid shuffle strategies and demonstrated the catalytic or structural role of aforementioned region in determining the substrate's entrance, cyclization/rearrangement mechanism and/or product specificity on
oxidosqualene-lanosterol cyclase, oxidosqualen-cycloartenol synthase, and other cyclase
enzymes. In addition, Phe-104-Thr, Trp-232-Arg, Trp-390-Ala, Phe-510-Arg, Trp-587-Ala, Phe-699-Ala, and Tyr-707-Ala failed to complement the ERG7 disruption were identified from the ERG7 counterselection process, indicating that these residues are also crucial for the catalytic function of the enzyme. For the last decade, exploitation of nucleic acid’s shape- or sequence-specific recognition properties as well as the in vitro polymerase chain amplification has greatly contributed to the identification of novel nucleic acid moleculeas for specific bio-molecular interactions with a variety of binding partners. SELEX, systematic evolution of ligands by exponential enrichment, is now widely applied to screen RNA or DNA oligos for binding against target small molecule organic dyes, nucleic acid binding proteins, or nucleic acids. The technology involves the synthesis of random oligonucleotide library, in vitro hybridization between target and library, enrichment of binding sequence via PCR amplification, iteration of selection cycle for binding affinity improvement, and identification of binding sequence. The novel SELEX technology is gaining more and more importance as an extremely useful screening tool, since a far larger population of molecules ( >1013 ) can be rapidly screened in short time. Moreover, functional
molecules can be selected from the mainly non-functional pool of RNA or DNA by column chromatography or other selection techniques that are suitable for the enrichment of any desired property.
In parallel, peptide aptamer, oligopeptides that can specifically recognize and strongly bind to a protein of interest, has become another new tools for molecular medicine search. Peptide aptamers are small oligopeptide, which have the potential to dominantly interfere with specific activity of their target proteins and, therefore, could be used as in vivo inhibitors. In addition, peptide aptamers also provide a basis for the development of novel diagnostic and therapeutic strategies, with implications for a broad variety of different disease entities, including metabolic disorders, infections, and cancer. For the diagnostic purposes, peptide aptamers could be employed as antibody-like molecules for the detection of the target, such as in western blot analyses. Following the immobilization of the target protein on a membrane, the peptide aptamer which specifically interact with the target protein could be visualized with secondary antibodies against the scaffold or against a tag sequence fused to the aptamer, which will become an alternative to the development of monoclonal antibodies.
三、結果與討論
Generation of Degenerate Single Strand DNA Library
A pool of 72-mer oligonucleotides which contain a degenerate 36-nucleotide sequence flanked by two 18-nucleotide regions was synthesized using the solid-phase phosphoramidite chemistry. The nucleotide sequences for the flanking regions are as follow: 5’-CGTACGGTCGACGCTAGC-3’ and 5’-biotin-GGATCCGAGCTCCACGTG -3’. The synthetic DNA was purified by PAGE and amplified by PCR to generate DNA library. The resulting biotinylated double stranded DNA was applied to an avidin-agarose column pre-equilibrated with buffer A. After the binding of the biotinylated DNA to the avidin beads, the unbound DNA was washed with five times of buffer A. Matrix-bound double strand DNA denatured with NaOH and the free DNA was precipitated. Following centrifugation, the DNA library was re-dissolved in the selection buffer and stored at -20 ℃ for selection.
Selection Using Proteins Immobilized on Microtiter Plate
To perform the aptamer selection, the ssDNA library was “panned” against protein immobilized on the surface of a microtiter plate. Different concentration of OSC was loaded onto the surface of the microtiter plate and incubated overnight at room temperature. After overnight incubation, linkers that are not derivatized with OSC were blocked with bovine serum albumin and incubated at 37 ℃ for 3
hr, followed by thrice washes with KPi/Tween 20. DNA library was first denatured at 95 ℃ for 5 min and then placed on ice for 5 min. The ssDNA pool was added to a BSA-only well and then incubated at room temperature for 1 hr. Next, the solution was transferred to the “OSC target” well and incubated for 1 hr before washed thrice with selection buffer. The bound aptamer was eluted with guanidine thiocyanate after the plate was heated at 80 C for 15 min and precipitated. The eluted DNA was PCR amplified and subjected for next round of selection. After 3 to 5 rounds of selection the recovered PCR products were visualized by gel electrophoresis and compared to the ones obtained from the control wells.
Selection Using Protein Immobilized on Filters
To ensure the most accessible folding or stable structure of DNAs, samples were pre-thermally equilibrated and then passed through filters to remove species not bound to the filters. Bound DNAs were extracted from the filters by 7M urea and heat incubation. Following the incubation of DNA with OSC-bound filter, proteinase K was added to digest the protein portion. The digested protein was extracted with phenol/chloroform and the DNA portion was precipitated with NaOAc/Ethanol solution. The collected DNA was PCR amplified and subjected for next round of selection. Again, after 3 to 5 rounds of selection the recovered PCR products were visualized by gel electrophoresis and compared to the ones obtained from the control wells.
Filter Binding Assay
Results from measurement of the amount of radioactively labeled DNA showed that only the DNA interact with the OSC was observed from filter binding assay. DNA did not interact with BSA as shown in Figure 1.
Figure 1. Filter binding assay of DNA with various concentrations of OSC ranging from 0.25, 0.5, 1.0, 2.0 to 4.0 µmol.
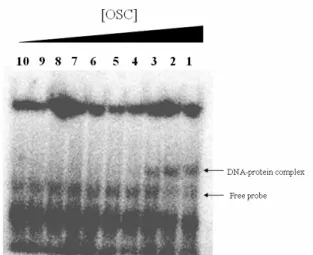
Electrophoretic Mobility Shift Assay (EMSA)
EMSA was applied to determine the later rounds of SELEX. A specific DNA-protein complex was observed as an up-shift band at position of 156 kDa as shown in Figure 2. DNA was eluted from DNA-protein complex and cloned into pBluescript II
Figure 2. EMSA results of OSC-bound DNA.
DNA Sequencing
DNA sequencing of SELEX plasmids isolated through the OSC-bound targets was carried out by the dideoxy chain-termination method using ABI PRISM BigDye Terminator Cycle Sequencing Reaction kits on an Applied Biosystems 3100 DNA Sequencer. After several rounds of selection, 9 aptamers which showed binding affinity to OSC target were selected, cloned, and sequenced. Following analysis of the sequences using the NCBI Blast program, the results were shown in Figure 3.
Figure 3. DNA sequences of OSC aptamers.
Biopanning Screening of OSC-Binding Peptide
Prepare a solution of 100 µg/mL of OSC in 5 mM KPi (add 1 mM DTT and 0.2 %Trition X-100)(pH 7.4). Add 150 µL of OSC solution to each microtiter well. Store plates at 4°C overnight. At the time, inoculate 10 mL LB medium with ER2738 at 37 ℃. Next day pour off the coating solution form each well and washing each well rapidly 6X with TBST (TBS + 0.1% [v/v] Tween-20). Fill each well completely with blocking buffer. Incubate at least 1 hour at 4°C. Discaed the blocking solution and wash each well rapidly 6X with TBST buffer. Dilute 4 x 1010 phage (10 µL of original library) with 100 µL of TBST. Pipet onto coated plate and rock gently for 10–60 minutes at room temperature. Discard nonbinding phage by pouring off and slapping plate face-down onto a clean paper towel. Wash plates 20 times with TBST. Elute bound phage with 200 µL of an elution buffer and rock gently for 10 minutes at room temperature. Pipet eluate into a microcentrifuge tube of 15 mL 1 M Tris-HCl (pH 9.1). Titer a small amount
ELISA Assay of OSC-Binding Peptide
For each clone to be characterized, inoculate 20 mL of LB medium with ER2738 and incubate at 37°C until slightly turbid. Alternatively, dilute an overnight culture of ER2738 1:100 in 20 mL LB. Add 5 µL of phage to each culture and incubate at 37°C with vigorous aeration for 4.5 hours. Transfer the culture to a centrifuge tube and spin 10 minutes at 10,000 rpm. Transfer supernatant to a fresh tube and re-spin. Pipet the upper 80% of the supernatant to a fresh tube and add 1/6 volume of PEG/NaCl. Allow phage to precipitate at 4°C for overnight. Spin PEG precipitation 15 minutes at 10,000 rpm at 4°C. Decant supernatant, re-spin briefly, and remove residual supernatant with a pipette. Suspend the pellet in 1 mL TBS. Transfer the suspension to a microcentrifuge tube and spin for 5 minutes at 4°C to pellet residual cells. Transfer the supernatant to a fresh microcentrifuge tube and re-precipitate with 1/6 volume of PEG/NaCl. Incubate on ice 60 minutes. Microcentrifuge for 10 minutes at 4°C. Discard supernatant, re-spin briefly, and remove residual supernatant with a micropipet. Suspend the pellet in 50 µL TBS. Titer the sample.
Coat one row of ELISA plate wells for each clone to be characterized with 100–200 µL of 100 µg/mL of OSC in 5 mM KPi buffer. Incubate at 4°C overnight. Shake out excess target solution and wash 6X with TBST ( TBS+0.1% tween-20 ). Fill each well completely with blocking buffer. Additionally, one row of uncoated wells per clone to be characterized should also be blocked in order to test for binding of each selected sequence to BSA-coated plastic. Incubate the blocked plates at 4°C, 1 hours. Shake out the blocking buffer and wash each plate 6 times with 1X TBST, slapping the plate face-down onto a clean section of paper towel each time. Carry out ten serial dilutions of the phage in 200 µL of TBS/Tween per well, starting with 1012 virions in the first well of a row and ending with 105 virions in the well. Incubate at room temperature for 1 hours with agitation. Wash plate 6 times with 1X TBST. Dilute HRP-conjugated anti-M13 antibody 1:5,000 in blocking buffer. Add 200 µL of diluted conjugate to each well and incubate at room temperature for 1 hour. Wash 6 times with 1X TBST. Add 1 µL 30% H2O2 to 10 mL of ABTS stock solution per plate to be analyzed. Add 200 µL substrate solution to each well, incubate at room temperature for 10–60 minutes. Read plates using a microplate reader set at 405 nm.
(a)
(b)
Figure 4. Biopanning results of (a) 7-mer peptide phage library and (b) 12-mer peptide phage library
Figure 5. OSC-binding peptide sequences
四、計劃成果自評
We have successfully accomplished the preliminary objective to select the OSC binding aptamers. The microtiter plate binding selection and electrophoretic mobility shift assay were proven to be useful for binding affinity selection. New materials have been obtained to regulate the oxidosqualene cyclization reactivity. The obtained result will be used to design new OSC inhibitor and regulate the OSC activity though interfering the binding affinity and specificity. Besides, the application of phage display libraries for OSC-binding peptide screening have resulted in obtaining one heptapeptide and four dodecaheptides which showed binding affinity to OSC enzyme. Further experiments on OSC affinity binding and specificity will be examined using the
monoclonal antibody generated from the OSC enzyme. Therefore, this research result opens new avenue for future development of DNA or peptide-based antifungal and hypocholesteremic drug for therapeutical purposes.
五、參考文獻
1. Barth, M. M.; Binet, J. L.; Thomas, D. M.; de Fornel, D. C.; Samreth, S.; Schuber, F. J.; Renaut, P. P. J. Med. Chem. 1996, 39, 2302-2312.
2. Bacher, J. M., and Ellington, A. D. (1998) DDT
3, 265-273.
3. Bock, L. C., Griffin, L. C., Latham, J. A., Vermaas, E. H., and Toole, J. J. (1992) Nature
355, 564-566.
4. Brody, E. N., and Gold, L. (2000) Reviews Mol.
Biotechnol. 74, 5-13.
5. Colin Cox, J., and Ellington, A. D. (2001)
Bioorg. & Med. Chem. 9, 2525-2531.
6. Gold, L., Brown, D., He, Y.-Y., Shtatland, T., Singer, B. S., and Wu, Y. (1997) Proc. Natl.
Acad. Sci. USA 94, 59-64.
7. Golden, M. C., Collins, B. D., Willis, M. C., and Koch, T. H. (2000) J. Biotechnol. 81, 167-178.
8. Hermann, T., and Dinshaw, J. P. (2000)
Science 287, 820-825.
9. Hesselberth, J., Robertson, M. P., Jhaveri, S., and Ellington, A. D. (2000) Reviews Mol.
Biotechnol. 74, 15-25.
10. Hoppe-Seyler, F., Crnkovic-Mertens, I., Denk, C., Fitscher, B. A., Klevenz, B., Tomai, E., and Butz, K. (2001) J. Steroids Biochem. & Mol.
Biol. 78, 105-111.
11. Iqbal, S. S., Mayo, M. W., Bruno, J. G., Bronk, B. V., Batt, C. A., and Chambers, J. P. (2000)
Biosensors & Bioelectronics 15, 549-578.
12. James, W. (2001) Current Opinion in
Pharmacology 1, 540-546.
13. Jayasena, S. D. (1999) Clinical Chem. 45, 1628-1650.
14. Kusser, W. (2000) Reviews Mol. Biotechnol. 74, 27-38.
15. Mark, M.; Muller, P.; Maier, R.; Eisele, B. J.
Lipid Res. 1996, 37, 148-158.
16. Meisenheimer, K. M., Meisenheimer, P. L., Willis, M. C., and Koch, T. H. (1996) Nucleic
Acids Res. 24, 981-982.
17. Meisenheimer, K. M., and Koch, T. H. (1997)
Critical Reviews Biochem. Mol. Biol. 32,
101-140.
18. Morris, K. N., Jensen, K. B., Julin, C. M., Weil, M., and Gold, L. (1998) Proc. Natl. Acad. Sci.
USA 95, 2902-2907.
19. Norris, C. L., Meisenheimer, P. L., and Koch, T. H. (1996) J. Am. Chem. Soc. 118, 5796-5803.
20. Willis, M. C., LeCuyer, K. A., Meisenheimer, K. M., Uhlenbeck, O. C., and Koch, T. H. (1994) Nucleic Acids Res. 22, 4947-4952.