中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/46155
46
0
0
全文
(2) 目錄 第一章 前言………………………………………………………………1 第二章 文獻探討…………………………………………………………2 壹、疼痛概述…………………………………………………………2 貳、疼痛的分類………………………………………………………2 參、發炎疼痛反應……………………………………………………3 肆、痛覺神經纖維……………………………………………………4 伍、痛覺傳導路徑……………………………………………………5 陸、發炎疼痛的動物實驗模式………………………………………5 柒、疼痛行為評估方式………………………………………………6 捌、臨床疼痛治療方式………………………………………………8 玖、針刺的傳統醫學理論與現代研究………………………………8 一、針刺的傳統醫學理論………………………………………8 二、針刺的現代研究……………………………………………9 拾、針刺止痛的神經機轉………………… .. ………………………10 一、周邊性止痛機轉……………………………………………10 二、中樞性止痛機轉……………………………………………10 三、神經傳遞物質及調控因子等止痛機轉……………………11 四、其他機轉……………………………………………………11 拾壹、鈉離子通道概述………………………………………………11 一、疼痛訊息與動作電位………………………………………11 二、鈉離子通道與動作電位……………………………………12 三、鈉離子通道與疼痛的相關性………………………………12 四、鈉離子通道的結構與分類…………………………………13 拾貳、Nav1.7 與疼痛的相關性………………………………………13 拾參、Nav1.8 與疼痛的相關性………………………………………14 拾肆、Nav1.9 與疼痛的相關性………………………………………15 拾伍、其他鈉離子通道簡介…………………………………………16 拾陸、疼痛的性別差異………………………………………………16 第三章 材料與方法……………………………………………………… 17 IV.
(3) 壹、實驗動物…………………………………………………………17 貳、實驗動物分組……………………………………………………17 參、發炎疼痛模式……………………………………………………17 肆、電針治療…………………………………………………………18 伍、行為觀察…………………………………………………………18 一、機械力試驗…………………………………………………18 二、輻射熱試驗…………………………………………………19 三、冷熱板試驗…………………………………………………19 陸、動物犧牲…………………………………………………………19 柒、免疫螢光染色法…………………………………………………19 捌、西方墨點分析法…………………………………………………20 玖、統計分析…………………………………………………………21 第四章 結果 …………………………………………………………… 21 壹、疼痛行為觀察結果………………………………………………21 一、機械力試驗結果……………………………………………23 二、輻射熱試驗結果……………………………………………23 三、冷熱板試驗結果……………………………………………23 貳、免疫螢光染色結果………………………………………………26 參、西方墨點分析法結果……………………………………………26 第五章 討論 …………………………………………………………… 29 第六章 結論 …………………………………………………………… 33 第七張 參考文獻 ……………………………………………………… 33 Abstract …………………………………………………………………… 40. V.
(4) 圖目錄 圖 1 疼痛行為試驗結果 …………………………………………………22 圖 2 免疫螢光染色結果. ……………………………………………… 25. 圖 3 西方墨點分析法結果……………………………………………… 27. 表目錄 表 1 西方墨點分析法-濃度定量分析 ………………………………… 28. VI.
(5) 在小鼠發炎疼痛模式中研究電針足三里穴位對鈉離子通道表現的影響 中文摘要 陳香霓 1,2 林以文 2 1. 中國醫藥大學北港附設醫院 2 中國醫藥大學針灸研究所. 關鍵詞. 電針、發炎疼痛、鈉離子通道、足三里穴位. 目的 鈉離子通道與神經的興奮性有關,尤其是在初級傳入神經元的痛覺 傳導上扮演了重要的角色,在 10 種鈉離子通道的分型中以 Nav 1.7、Nav 1.8 及 Nav 1.9 與疼痛較具高度相關性,最近研究指出在 carrageenan 的 發炎模式下,背根神經節中可發現到 Nav 1.7 和 Nav 1.8 的表現量增加, 但 Nav 1.9 的變化則不明顯。電針已被廣泛運用在疼痛治療上,先前研 究指出電針可以引發鴉片肽、腺苷酸等神經傳導物質釋放來達到止痛的 效果,然而電針止痛與鈉離子通道的關係卻很少被探討,因此我們使用 小鼠發炎疼痛模式來研究電針足三里穴位的止痛效果以及電針對於鈉 離子通道表現的影響。 方法 我們將 18 隻 ICR 母鼠分成控制組、發炎組及電針組,連續 4 日在 足三里以 1mA、2Hz 電針 15 分鐘,觀察疼痛行為的變化,並在第 4 日 取下同側第 3、4、5 腰椎的背根神經節,分析鈉離子通道的表現。 結果與結論 研究結果顯示電針可明顯降低機械性及溫度性的疼痛行為,並可調 降 Nav 1.7 及 Nav 1.8 在發炎模式中的過度表現,但 Nav 1.9 則無顯著性 差異。推論電針足三里減輕發炎疼痛的療效機轉,和調控鈉離子通道中 Nav 1.7 與 Nav 1.8 的表現相關,但與 Nav 1.9 則較無相關性。. VII.
(6) 第一章. 前言. 疼痛是影響生活品質的主要原因之一,五分之ㄧ以上的人口伴隨有 疼痛的困擾,目前臨床對疼痛的治療主要有藥物和外科手術二種[1,2],但 即使疼痛症狀可因藥物或手術緩解,其所伴隨的副作用亦引起許多患者 的不適。鈉離子通道(Nav:voltage-gated sodium channel)為細胞膜上 的跨膜蛋白,鈉離子透過鈉離子通道在細胞內外流動繼而誘發動作電 位,帶動神經細胞的興奮,因此鈉離子通道在初級傳入神經元的痛覺傳 遞上扮演了重要的角色。臨床局部麻醉最常使用的麻醉劑,如lidocaine、 procaine即是透過阻斷鈉離子通道,阻止痛覺傳遞而達到止痛的效果。 近來相當多研究陸續發現鈉離子通道與發炎疼痛的相關性[17],就目前所 知,鈉離子通到一共有10個分型,研究指出其中Nav 1.7、Nav 1.8及Nav 1.9與疼痛具有一定的相關性[22]。 針刺在中國傳統醫學中應用於臨床治療已有二千多年歷史,並廣泛 應用在各種疼痛治療中。數十年來,相當多的學者不斷以實驗證實針刺 的療效[20],目前已知的針刺神經止痛機轉,包括周邊性神經機轉,例如 Melzack 及 Wall 所建立的閘門控制學說,中樞性調控機轉則與腦部多項 結構的抑制或興奮有關,另外鴉片肽(opioid peptides)、腺甘酸 (adenosine)等神經傳遞物質及調控因子皆已被報導與針刺療效相關 [17]. ,但對於針刺止痛機轉和鈉離子通道的相關性目前仍是未知。 因此本研究利用 carrageenan 誘發小鼠發炎疼痛模式,經電針足三里. 穴位治療後,以 von Frey test、Hargreaves’ test 及冷熱板試驗,觀察小鼠 機械性及溫度性疼痛行為的表現,並取下同側腰椎第 3、4、5 節的背根 神經節,以免疫螢光染色法及西方墨點分析法觀察背根神經節中 Nav 1.7、Nav 1.8 及 Nav 1.9 在發炎模式中及電針治療後的變化。我們推測, 在發炎模式中電針止痛的機轉與鈉離子通道的表現有相關性。. 1.
(7) 第二章 文獻探討 壹、疼痛概述 疼痛為病患就醫的主要原因之一,因此自古至今皆是很值得探討的 議題。根據統計,約 20%以上的人口有疼痛的困擾[1],因慢性疼痛尋求 醫療幫助的病患高達就醫總人數 20-50%,慢性疼痛更是全球三大健康 問題之一[2]。以急性疼痛的生理意義來說,疼痛是人體避開周遭危險環 境的保護反應,是維持生命所不可或缺的警報及防禦機制;然而對於慢 性疼痛而言,疼痛就不再是警報反應,反而是干擾生活品質的惱人問題。. 貳、疼痛的分類 疼痛根據發作的時間長短,可分為急性疼痛和慢性疼痛,亦可根據 病理的機轉不同而有其他的分類方式。 一、根據引起疼痛的病理原因不同,可將疼痛分為以下四大類[3]: (一)傷害性疼痛 例如溫度、機械力、電流電壓、化學等傷害性刺激所引起的痛覺。 又可分為體表性疼痛及內臟性疼痛二類。 (二)發炎性疼痛 因組織發炎而導致的疼痛。 (三)神經性疼痛 如神經截斷、壓迫或神經發炎等任何造成神經傷害的原因,皆能 引起神經性疼痛。 (四)疾病性疼痛 例如癌痛、複雜性區域疼痛綜合症(CRPS:complex regional pain syndrome)或精神疾患所造成的疼痛。. 2.
(8) 其中傷害性疼痛和發炎性疼痛通常持續的時間較短,神經性疼痛及 疾病性疼痛持續的時間較長,甚至多達好幾年[3]。 二、疼痛的感覺亦可區分為以下三大類[4]: (一)痛覺敏感(hyperalgesia) 當組織損傷後,若再次受到傷害性刺激,會比未受傷的組織來得 痛,以避免受傷的組織持續再受傷害,此現象稱為痛覺敏感。 痛覺敏感若發生在受傷害的部位上,稱之為原發性痛敏感 (primary hyperalgesia),若發生在遠離受傷害的部位上, 稱之為續發性痛覺敏感(secondary hyperalgesia)[5]。 (二)痛覺異常(allodynia) 是指對無傷害性的刺激也感覺到疼痛,例如對輕觸覺或無傷害性 的溫度亦感到疼痛。 (三)自發性疼痛(spontaneous pain or dysesthesia) 是指沒有受到任何的刺激,也有自發性疼痛感。 痛覺敏感多發生在發炎性疼痛中,與受傷害組織週邊的發炎物質釋 放有關;痛覺異常多發生在神經性疼痛中,例如糖尿病神經病變[1]。二 者與週邊痛覺神經敏感或中樞神經敏感有關[6]。. 參、發炎疼痛反應 在發炎的部位可見紅、腫、熱、痛等四大現象,稱為發炎反應,這 是因為受傷害的組織本身會釋出發炎前驅物(pro-inflammatory mediators),如組織胺(histamine)、緩激肽(bradykinin)、羥色胺(5-HT: serotonin,5-Hydroxytryptamine,也稱為血清素)、前列腺素(prostaglandin)、氫離子(H )、三磷酸腺苷(ATP)等物質[7];與發炎相關 +. 的肥大細胞(mast cell)亦會釋放組織胺、緩激肽、前列腺素[7];巨噬細 胞(macropgage)則會釋放白細胞介素(interleukin)、腫瘤壞死因子 α 3.
(9) (TNF-α:tumor necrosis factor α)、神經生長因子(NGF:nerve growth factor),以上諸多發炎調控物質會誘發局部發炎反應,並活化痛覺神經 元(nociceptive neuron 或稱傷害性神經元),引起疼痛感覺,而痛覺神 經元末梢的痛覺感受器(nociceptor 或稱傷害感受器)本身所釋放的 P 物質(substance P)會刺激肥大細胞及嗜中性血白球釋出更多的組織胺、 神經生長因子以放大發炎反應,另外痛覺神經元末梢釋出的 P 物質、降 鈣素基因相關肽(CGRP:calcitonin gene-related peptide)也會刺激周邊血 管擴張,同時加強疼痛訊號[6, 7]。. 肆、痛覺神經纖維 當溫度、機械性、化學性等刺激達到傷害周邊組織的程度後,痛覺 神經元周邊游離神經末稍(free nerve-ending)的痛覺感受器(nociceptor) 便會將傷害性刺激轉化(transduction)成神經衝動。這些神經末梢的痛 覺感受器多分布在皮膚、肌肉、骨膜、關節及內臟附近以便偵測刺激[8]。 這些感受外界刺激的痛覺神經元屬於初級感覺神經元(primary sensory neurons;或稱初級輸入神經元 primary afferent neurons)的一種[1]。 初級感覺神經元依據纖維的粗細不同及髓鞘包覆與否,主要可分為 三大類[9]: 一、Aα和 Aβ纖維(粗神經纖維) 此類神經元的纖維直徑最大,外層包覆有髓鞘,主要傳遞本體感覺 和非傷害性的機械刺激,與痛覺較無關。例如 Aβ 纖維連接皮膚表層的 Merkel 氏盤(Merkel's discs)、Pacinian 小體(pacinian corpuscles)及毛 囊,負責感受震動覺及輕觸覺[10]。 二、Aδ 纖維(粗神經纖維) 此類神經纖維的直徑大小介於三大類神經纖維之間,外層亦包覆有 髓鞘,負責傳遞位置明確、快速而持續時間短暫的銳痛(sharp pain,或 稱快痛 first or fast pain)感覺。在電生理的研究進展下,甚至可將 Aδ 4.
(10) 纖維再分為二大類,Type IAδ 纖維:主要負責反應機械性傷害刺激,對 傷害性高溫的反應閾值較高(>53 oC);Type II Aδ 纖維:對傷害性高 溫的反應閾值較低(>43oC),但對機械性傷害較不敏感。 三、C 纖維(細神經纖維) 此類神經纖維的直徑最小,外層無包覆髓鞘,負責傳遞位置不明 確、持續時間較長的鈍痛(dull pain,或稱慢痛 second or slow pain)感 覺。C 纖維中亦有許多分型,分別對溫度、機械力、化學性刺激有不同 的敏感度。但須注意的是,並不是所有的 C 纖維都具有痛覺感受器。. 伍、痛覺傳導路徑 痛覺神經元將傷害性訊息傳遞至背根神經節(DRG : dorsal root ganglia) 後,經由李氏徑(tract of Lissauer)傳入脊髓背角(dorsal horn), 將訊息交給次級輸入神經元(secondary afferent neurons),於背角再分 為兩條傳導路逕,一條傳至腹角(ventral horn),引起脊髓反射;另一 條經上行通路傳遞至視丘(thalamus)、皮質(cortex),引起痛覺。一 般而言傷害性刺激只要進入網狀結構視丘或較低等中樞即可引起疼痛 的感受,而大腦皮質則是掌管感覺的判斷,負責分辨痛覺的性質、強度 與位置[11]。. 陸、發炎疼痛的動物實驗模式 實驗室中可利用發炎疼痛模式來研究疼痛議題。發炎疼痛模式可依 誘發動物發炎部位不同,而分為表皮及皮下組織發炎、肌肉炎及關節炎 等。用於表皮及皮下組織發炎的藥劑主要有以下幾類[6]: 一、福馬林(formalin) 注射於鼠類的足底,幾分鐘後即可誘發神經與發炎疼痛,發炎反應 可持續數小時。 5.
(11) 二、鹿角膠(carrageenan)、完全佛蘭氏佐劑(CFA:complete Freund's adjuvant) 注射以上二種藥劑皆可誘發持續性的發炎疼痛及痛覺敏感,此類的 發炎模型也較相似於人體組織受傷後的發炎反應。注射 CFA 後,在 2 小時左右開始發生疼痛,6-8 小時後達到疼痛高峰,反應可持續至 1-2 周。施打 carrageenan 也有類似 CFA 的痛覺敏感反應,但持續時間較短, 約小於 1 周。. 柒、疼痛行為評估方式 當週邊組織受到傷害後,會產生痛覺敏感(hyperalgesia)的情形, 但在動物實驗模式中,因為無法直接得知動物對疼痛的感受程度,所以 透過觀察動物的行為表現,客觀評估動物的痛覺變化。 根據位於周邊的痛覺接受器主要可被機械性、溫度性(冷或熱)、 化學性三種刺激活化,針對這三種不同類型的刺激性質,發展出相對應 的痛覺評估方式。 一、機械性痛覺評估 機械性痛覺評估中最常採用的是 von Frey test,由 Maximilian von Fery 在 1896 年提出[12],使用不同粗細的細絲(filament)在動物表面施 以不同的機械性壓力刺激,觀察到動物抬起受刺激的肢體以避開刺激源 的動作,稱為縮足反應(withdrawal),藉以評估動物對機械性壓力刺 激的耐受程度。 二、溫度性痛覺評估 溫度性痛覺評估最常採用的方式包括二種,輻射熱痛刺激試驗 (Hargreaves’ test)及冷熱板試驗(hot/cold plate test)。 (一)輻射熱痛刺激試驗(Hargreaves’ test) 6.
(12) 由 Hargreaves 等人在 1988 年提出[13]。輻射熱痛刺激試驗是將動 物靜置在透明塑膠觀察盒內,使用輻射熱源透過下方的透明玻璃 ,照射動物足底,測量動物自開始給予輻射熱源至抬起被照射肢 體的時間,稱為潛伏期(latency),時間越短表示痛覺閾值 (nociceptic threshold)越低,可量化痛覺敏感的程度。 (二)冷熱板試驗(hot/cold plate test) 熱板試驗(hot plate test)由 Eddy 在 1950 年提出[14];冷板試 驗(cold plate test)由 Bennett 在 1988 年提出[15]。此二項痛 覺試驗是將動物放在冷或熱的金屬板上,利用傷害性溫度的刺激 引起疼痛,觀察自刺激至動物因疼痛抬起後肢的時間,即為潛伏 期(latency),另可記錄抬起後肢時所維持的時間,二者皆可用 於量化痛覺敏感的程度。 三、化學性痛覺評估 化學性痛覺由 Bennett 在 1988[15]年的研究中提出,以 50%芥子油 塗抹於動物腳掌,以誘發化學性刺激引起痛覺,紀錄自刺激給予到動物 因疼痛抬起後肢的時間,即為潛伏期(latency)。 小鼠疼痛行為評估除了縮足反應(withdrawal)外,還有舔足 (licking)、抬足(lifting)、抖腳(shaking)、跳躍(jumping)、自 殘(autotomy)、囓咬(biting)、學習逃避(learned escape)、警備 (guarding)、步態(gait)、姿勢(posture)、增加衝突(reinforcement conflict)、焦慮(anxiety)、注意力失能(attention disability)、社交 (sociability)、睡眠(sleep)等行為評估方式[16]。. 捌、臨床疼痛治療方式 目前臨床治療疼痛的方式大致有以下幾種:麻醉性鎮痛藥(narcotic analgesics) ,非麻醉解熱鎮痛藥(non-narcotic anti-pyretics and analgesics) 及非類固醇抗發炎藥物(NSAID:non-steroid anti-inflammatory drug), 7.
(13) 臨床上經常使用的 NSAID 類止痛藥,例如 aspirin 或 ibuprofen 即是透過 抑制環氧化酶(cyclooxygenases,Cox-1 and Cox-2)以降低發炎調控物質 prostaglandin 的合成,進而達到抗發炎的療效。止痛效果不佳時,可合 併使用精神科用藥,如抗焦慮劑、鎮靜劑等藥物輔助治療疼痛。藥物治 療效果受限者,則可考慮神經外科手術[7]。 上述疼痛治療的方式,仍舊無法緩解各種種類的疼痛,新的止痛方 法不斷的在研發,近期研究的新藥有作用在第一型瞬時受體電位離子通 道(TRPV1:transient receptor potential vanilloid subtype I)感受器的拮 抗劑、NMDA (N-methyl-D-aspartate)受體拮抗劑等[7],但即使疼痛症狀 可因藥物緩解,其所伴隨的副作用亦引起許多患者的不適,因此發展出 其他的輔助性療法,例如針刺、推拿、氣功等另類醫療(alternative medicine)[7]。. 玖、針刺的傳統醫學理論與現代研究 針刺在中國傳統醫學中應用於臨床治療已達二千多年歷史,1996 年 11 月召開的世界衛生組織義大利米蘭會議,提出 64 種針刺適應症,其 中以跟疼痛相關的項目居多,顯示針刺在疼痛治療領域的重要性。 一、 針刺的傳統醫學理論 (一)經絡與得氣 傳統中醫認為人體有經絡存在,身體的"氣"運行於經絡間以佈 施能量給組織及器官,當氣的運行因陰陽不平衡的疾病狀態而阻 滯經絡時,就會產生疼痛的症狀,針刺穴位有助於陰陽氣血的平 衡,達到疏通經絡的止痛效果。傳統中醫指出,針刺穴位重點在 於"得氣",可表現在施針者針下如魚吞鉤的沉緊感覺及被施針 者在針刺部位產生痠、麻、重、脹的感覺,現代研究則透過肌電 圖的紀錄,認為得氣或許和肌肉收縮相關[17]。. 8.
(14) (二)足陽明胃經的經絡循行 於《靈樞‧經脈》中記載足陽明胃經的循行路徑:「胃足陽明之 脈,起於鼻,交頞中,旁約太陽之脈,下循鼻外,入上齒中,還 出挾口,環唇,下交承漿,卻循頤後下廉,出大迎,循頰車,上 耳前,過客主人,循髮際,至額顱。 其支者:從大迎前,下人 迎,循喉嚨,入缺盆,下膈,屬胃,絡脾。 其直者:從缺盆下 乳內廉,下夾臍,入氣街中。其支者:起於胃口,下循腹裡,下 至氣街中而合。以下髀關,抵伏兔,下膝髕中,下循脛外廉,下 足跗,入中趾內間。 其支者,下膝三寸而別,下入中趾外間。 其 支者:別跗上,入大趾間,出其端。」[18],如同上述經絡循行路 線所示,足陽明胃經循行從頭走至足,經脈下行於大腿,下至 膝髕,沿著脛骨前脊外側,下至足背。 (三)足三里穴位的主治項目 足三里為足陽明胃經之合穴(合穴主治逆氣而洩),主治胃腸 病、頭面、目、鼻、口、齒痛,神志病及經脈循行部位的其 相關病症。故根據「經脈所過,主治所及」之治則,《醫宗金鑑》 中提到:「足三里主治中風、中溼、諸虛、耳聾、上牙疼、水腫、 心腹鼓脹、噎膈哮喘、寒溼腳氣,上、中、下三部痺痛等症。」; 《馬丹陽天星十二穴治雜病歌》記載:「三里膝眼下,三寸兩筋 間,能通心腹脹,善治胃中寒、腸鳴並洩瀉、腿腫膝胻痠…」; 《席弘賦》記載:「耳內蟬鳴腰欲折,膝下明存三里穴,髖骨腿 疼三里瀉。」。下肢發炎性疼痛的症狀相當於古代所謂的下肢痛、 下肢痺症的症狀,代表下肢關節或肌肉疼痛的臨床表現[19]。 二、針刺的現代研究 近數十年來,相當多的學者投入在動物實驗及臨床試驗中,不斷的 以實驗證實針刺的療效,2009 年一篇回顧臨床針刺鎮痛試驗的文章中提 到,針刺鎮痛多見於術後痛、下背痛、膝關節炎、慢性頭痛、肩痛頸痛、 癌痛等臨床症狀中[20]。 9.
(15) 統計研究指出,針刺後的 40 分鐘左右止痛效果會達高峰,止痛效 果可持續二小時左右[17]。針刺穴位後使用電針刺激較手捻針有更好的療 效[17],且因電針刺激的時間、強度、頻率的參數設定較為穩定,更適合 應用於基礎或臨床研究。. 拾、針刺止痛的神經機轉 2008 年趙氏在一篇回顧針刺止痛試驗的文章中指出,針刺止痛的神 經機轉大致可分為四大類[17]: 一、周邊性止痛機轉 1965 年 Melzack 及 Wall 建立閘門控制學說(gate control theory), 針刺所產生的銳痛感經由粗神經纖維(Aδ fibers)傳遞,透過刺激抑制 性神經元(inhibitory neuron)抑制細神經纖維(C fibers)所傳遞的慢性 疼痛感覺,並可抑制 projection cell 將疼痛訊息往中樞上傳[10] ,以達到 止痛效果。 二、中樞性止痛機轉 综合多項研究發現,針刺鎮痛的中樞性調控機轉與腦部多種結構的 抑制或興奮相關,例如延髓腹側髓質(rostral ventromedial medulla)、 細胞核中縫大核(mainly the nucleus raphe magnus)、導水管周圍灰質 (periaqueductal grey),藍斑核(locus coeruleus)及弓狀核(arcuate nucleus)等皆參與其中。 回顧文獻亦發現不同的電針頻率可誘發腦部分泌不同的鴉片肽 (opioid peptides)而達到止痛效果。例如 2Hz 的電針頻率以分泌 β-內啡 肽(β-endorphin)、內嗎啡肽(endomorphin)、腦啡肽(enkephalins) 為主;100Hz 的電針頻率以強啡肽 A (dynorphin A)為主;當 2Hz 與 100Hz 二種頻率交替使用,或使用介於上二者之間的 15Hz 頻率,則可同時產 生上述多樣的鴉片肽。. 10.
(16) 三、神經傳遞物質及調控因子等止痛機轉 對針刺鎮痛有正面性影響的神經傳遞物質及調控因子,包括阿片肽 (opioid peptides)、腺甘酸(adenosine)、5-羥色胺(5-HT: 5-hydroxytryptamine)、G-氨基丁酸(GABA:g-amino-butyric acid)、 生長抑素(SOM:somatostatin)、精氨酸加壓素(APV:arginine vasopressin)、神經降壓素(NT:neurotensin)等。 對針刺鎮痛有負面性影響的包括膽囊收縮素(CCK-8:cholecystokinin octapeptide)、P 物質(substance P)、血管緊張素Ⅱ(angiotensin I)、多巴胺(dopamine)、正腎上腺素(NA:noradrenalin)、穀氨酸 (glutamate)等。 四、其他機轉 研究證據顯示,中樞神經的膠質細胞,包括小膠質細胞(microglia) 和神經膠質細胞(astrocytes),都參與發炎性疼痛及神經性疼痛的表現。 其他與疼痛相關的可能分子機制尚包括:G 蛋白偶聯受體(GPCR:G protein-coupled receptors)、細胞外信號調節蛋白激酶(ERK)、核因子 -κB(NF-kB)及 cDNA 微陣列(cDNA microarray)等。 拾壹、鈉離子通道概述 一、疼痛訊息與動作電位 初級輸入性神經元的末梢有許多不同的受體及離子通道,可將感應 到的各類不同刺激轉換(transduction)成電位衝動,電位衝動繼而引發 神經軸(axon)上一連串的動作電位以產生訊息的傳遞(conduction)[1], 因此痛覺訊息從周邊傳遞至中樞神經系統有賴於動作電位的產生。 痛覺敏感可源自於週邊或中樞神經系統的反應,導致疼痛感覺增強 的主要原因之一為神經細胞的電氣興奮性增加,而鈉離子通道在週邊及 中樞神經系統的電氣興奮過程中扮演相當重要的地位[21]。. 11.
(17) 二、鈉離子通道與動作電位 鈉離子通道為哺乳動物細胞膜上的跨膜蛋白,能讓鈉離子快速的出 入細胞內外,產生細胞膜電位的變化,以誘發動作電位來傳遞訊息[7]。 動作電位處於靜止膜電位(resting membrane potential 或稱初始相 膜電位 prime phase)的階段,相對細胞膜外位電位約為-70mV,此時鈉 離子通道關閉,等候接受刺激活化,當刺激來臨時,鈉離子通道打開, 因細胞膜內外鈉離子濃度的懸殊差異,帶正電的鈉離子由細胞外向細胞 內湧入,頓時造成膜電位上升,若達到閾值(threshold),則觸發動作電位, 可見膜電位陡峭上升,此時稱為去極化階段(depolarizion or acived phase),而當細胞膜電位持續上升約至 15 mV 時,鈉離子通道的閘門 由於蛋白結構發生形變而塞住通道洞口,無法再通透鈉離子,造成鈉離 子通道進入不活化狀態,同時鉀離子通道打開,帶正電的鉀離子由細胞 內向細胞外湧出,膜電位逐漸往-80mV 的靜止膜電位下降,此時稱為再 極化階段(redepolarizion phase),於再極化階段時,無論刺激多頻繁多 強都不能再引起下一個動作電位,所以又稱為不反應期(inactivated phase)。直至整個膜電位到達-80mV 的靜止膜電位時,塞住鈉離子通道 口的變形蛋白結構解除,鈉離子通道才能接受下一個刺激,而產生下一 個動作的週期。整個動作電位產生的過程約歷時 10 毫秒,產生 2 nA 的 電流量。因此鈉離子通道經由靜止膜電位階段(resting membrane potential)、去極化階段(depolarizion or acived phase)及再極化階段 (redepolarizion phase)不斷的週期性循環中,產生一連串的動作電位[7]。 三、鈉離子通道與疼痛的相關性 近十幾年來鈉離子通道在痛覺領域已被廣泛的研究,證實鈉離子通 道存在中樞及周邊神經系統中,尤其是多樣化的鈉離子通道分布在痛覺 神經中,與痛覺具有一定程度的相關性[21]。Black 等人在 2004 年指出在 發炎模式下,鈉離子通道與疼痛感覺的傳輸有關,並可在背根神經節中 發現鈉離子通道表現的變化。臨床局部麻醉最常使用的麻醉劑,如 lidocaine、procaine 則是透過阻斷鈉離子通道而達到止痛的效果。研究發 現,針刺合谷穴所產生的止痛效果,可經由在合谷穴注射 procaine 後完 12.
(18) 全阻斷止痛效果[17]。以上眾多研究顯示鈉離子通道在疼痛機轉中佔有重 要的地位。 四、鈉離子通道的結構與分類 在感覺神經元細胞本體聚集的背根神經節中,所測得的再極化階段 鈉離子動作電位電流,比在海馬迴中測到的呈現更多樣化的電流表現, 表示周邊神經系統的鈉離子通道可能具有很多分型[8]。近一步研究發 現,鈉離子通道的結構主要由一個 α 亞基及四個 β 亞基(β1、β2、β3、 β4)所組成。α 亞基主要構成通道的孔洞,並決定鈉離子通道的主要功 能;β 亞基則可調控鈉離子通道的功能。目前在哺乳類身上發現的鈉離 子通道,因 α 亞基上的差異,可將鈉離子通道一共分為 10 個分型,其 中 Nav 1.1-1.9 與動作電位相關,Na X 為單純離子通道與電位無關[8]。 在成人的中樞神經元中可見Nav 1.1、Nav 1.2和 Nav 1.6,成人背根 神經節中的感覺神經元的則可見Nav 1.1、Nav 1.6、Nav 1.7、Nav 1.8和 Nav 1.9[22]。在鈉離子通道的10個分型中,以Nav 1.7、Nav 1.8及Nav 1.9 這3個分型已經被證實與疼痛較具有相關性,其餘6個分型的鈉離子通道 與疼痛較不相關或關係尚未明確[22]。 河豚毒素(tetrodotoxin,TTX)是一種對鈉離子通道專一性很高的 阻斷劑,依對河豚毒素的敏感程度,可將鈉離子通道區分為對河豚毒素 敏感的TTX-S及對河豚毒素較不敏感的TTX-R的二大分類[8]. 拾貳、Nav 1.7 與疼痛的相關性 Nav 1.7屬於TTX-S type,主要位在背根神經節細神經纖維的神經元 中,同時也存在交感神經節中[23]。Nav 1.7電流特性為快速激活(acti -vated)及失活(inactivated)[23],推論Nav 1.7與產生動作電位的閾值設 定相關[22]。 鈉離子通道的分型中最富盛名就屬Nav 1.7,原因來自於科學家發現 在巴基斯坦北部有一些家族遺傳性的失痛症,這群人生來即不具疼痛 感,例如熱刺激、穿刺傷甚至骨折皆不覺疼痛,也因此失去保護性的疼 13.
(19) 痛反射,但令人感興趣的是這群人除痛覺喪失外,其他感覺皆正常,且 身體其他功能亦與正常人無異。研究這些家族的基因發現,因為SCN9A 基因序列的突變導致Nav 1.7的功能完全喪失,以至對疼痛完全沒有感受 力,並將稱之為離子通道相關的對疼痛不敏感(CIP:channelopathyassociated insensitivity to pain)[8,40]。如果可研發Nav 1.7的抑制劑,將可 成功阻斷疼痛,又不影響其他生理功能,因此鈉離子通道與疼痛相關性 的研究自此大量展開[8]。 與Nav 1.7相關的痛覺異常疾病還有遺傳性的紅斑性肢痛症(IE: inherited erythromelalgia),起因於SCN9A基因的點突變,導致痛覺神經 的動作電位閾值變低且發生頻率變高,使整體動作電位增加,病人呈現 四肢末稍如火燒般的慢性疼痛[22]。另一個與Nav 1.7相關的體染色體顯性 遺傳疾病為陣發性劇痛症(PEPD:paroxysmal extreme pain disorder), 起因亦是SCN9A基因的點突變,但發生機轉與IE不同,PEPD乃因動作 電位再極化階段(不反應期)失常,動作電位持續時間延長,疼痛增加, 病人呈現直腸、眼睛、頜下火燒般的疼痛。因Nav 1.7基因點突變導致的 紅斑性肢痛症(IE)和陣發性劇痛症(PEPD),皆可發現有燒灼般的疼 痛感,且都伴隨者紅、腫、熱、痛的發炎症狀,因此推測Nav 1.7與發炎 性疼痛有密不可分的關係性。 已有相當多動物實驗發現,在發炎疼痛模式中Nav 1.7有增加的趨勢 [8]. 。在慢性發炎疼痛中可見Nav 1.7 mRNA及蛋白質表現增加,指出Nav. 1.7與發炎反應的疼痛機轉相關[23]。. 拾參、Nav 1.8 與疼痛的相關性 Nav 1.8 屬於 TTX-R type,亦多位在背根神經節細神經纖維的神經 元中[22] 。Nav 1.8 電流的特性為快速從非反應期回復,故能產生高頻率 動作電位[24]。過去研究亦指出,Nav 1.8 會受到發炎物質的調控,增加 激活及失活的速度及電流的強度,而增加感覺神經元的興奮[25, 26]。. 14.
(20) 在足底注射 carrageenan 和 CFA 發炎模式下,背根神經節中可見 Nav 1.8 mRNA 及蛋白質表現量的增加[27, 28] ,指出 Nav 1.8 與發炎反應 的疼痛機轉相關[23]。Nassar 於 2004 年發現在發炎模式下,Nav 1.8 基因 剔除的小鼠可見 TTX-sensitive 代償電流的增加,同時發現 Nav 1.7 mRNA 表現增加,顯示 Nav 1.7 可代償 Nav 1.8 基因剔除的小鼠的功能, 推論 Nav 1.7 與 Nav 1.8 在疼痛機轉的角色上彼此具有相關性[9]。. 拾肆、Nav 1.9 與疼痛的相關性 Nav 1.9 屬於 TTX-R type,亦多位在背根神經節的細神經纖維神經元 中,不同於 Nav 1.7、Nav 1.8,Nav 1.9 與靜止膜電位較有相關性[23]。Nav1.9 相對於其他鈉離子通道較晚被發現的原因,除了因為 Nav 1.9 的電流消 失速度快,因此不容易被記錄以外[29],Nav 1.9 產生的電流近似靜止膜 電位也是原因之ㄧ。Nav 1.9 的電流被發現的起緣來自於在 Nav 1.8 基因 剔除的小鼠仍可偵測到些餘的 TTX-R 電流,陸續研究發現 Nav 1.9 的動 作電位電流接近於靜止膜電位,因此推論 Nav 1.9 在靜止膜電位上扮演 重要角色[23]。 文獻指出發炎調控因子如bradykinin、5-HT和ATP,能影響 Nav 1.9 的表現[31]。研究發現注射CFA後的第七天,會引起Nav 1.9的mRNA增加 [30]. ,但有另一個研究發現在carrageenan誘發的發炎痛模型中,DRG中Nav. 1.9 mRNA及蛋白質並無明顯變化[20]。另有研究指出Nav 1.9基因剔除的 小鼠在formalin、carrageenan、CFA和prostaglandin E2 (PGE2)所誘發的發 炎模式中,皆可見痛覺敏感減輕的現象,且對發炎物質的敏感度亦降低 [31]. ,但有一相似的研究表示,Nav 1.9基因剔除的小鼠在formalin、. carrageenan、CFA和 prostanoids所誘發的發炎疼痛中,降低或喪失疼痛 的表現並不明顯[32]。以上文獻指出,Nav 1.9與發炎疼痛的相關機轉尚未 明確。. 15.
(21) 拾伍、其他鈉離子通道簡介 Nav 1.1-Nav 1.6這6個鈉離子通道的分型與疼痛較不相關。[23] 一、Nav 1.1(TTX-S type) 存在週邊及中樞神經系統中,對痛覺中的意義尚未明確[23]。 二、Nav 1.2(TTX-S type) 主要存在中樞神經系統中,在背根神經節裡的電流量相當低[23]。 三、Nav 1.3(TTX-S type) Nav 1.3被認為與動作電位的誘發及發生頻率相關,但Nav 1.3多發 現在未成熟的胎兒或初生兒的神經元中,在成人神經元相當少見。雖然 有研究指出疼痛時Nav 1.3的表現量會增加,但在另一研究中並無發現 類似的情形。Nassar 在2006年指出Nav 1.3基因完全剔除或選擇性剔除 的小鼠在發炎及神經痛模式中,與正常小鼠無異。因此Nav 1.3與疼痛的 相關性尚未明確[3] 。 四、Nav 1.4(TTX-S type) 多見在骨骼肌中[23]。 五、Nav 1.5(TTX-R type) 多存在心肌中[23]。 六、Nav 1.6(TTX-S type) 存在週邊及中樞等多種具髓鞘神經元中,但Nav 1.6在痛覺中所代表 的意義尚未明確[23]。. 拾陸、疼痛的性別差異 疼痛的感覺往往因人而異,因為影響疼痛認知的主觀因素很多,因 此疼痛的感受強度,不一定與刺激的大小成比例。此外疼痛亦有性別上 的差異性,根據流行病學的統計研究指出,慢性疼痛如筋肌膜疼痛、三 叉神經痛等發生在中年人和女性的族群比例較高。而在動物實驗的研究 中,母鼠對痛覺的敏感度高於公鼠,且母鼠對止痛劑的療效亦較公鼠明 顯[3]。 16.
(22) 第三章 材料與方法 壹、實驗動物 本研究所使用之 18 隻 ICR 母鼠均購自樂斯科生物科技股份公司 (Bio LASCO Taiwan Co.Ltd.),周齡 8-12 周,體重 20-30 公克之間。 飼養於每日 12 小時人工光照(早上 8 點至晚上 8 點),空調溫度 25±2 oC 之環境,並給予充足飼料及飲用水。所有研究過程皆符合動物實驗倫理 委員會之規範。. 貳、實驗動物分組 18 隻 ICR 母鼠隨機分為三組,包括: 一、控制組(Con:Control group) 注射生理食鹽水(normal saline),無接受電針治療。 二、鹿角膠組(Car:Carrageenan) 注射鹿角膠,無接受電針治療。 三、電針組(Acu:Electroacupuncture) 注射鹿角膠後,接受電針治療。 每組各有 6 隻小鼠。在注射 carrageenan 之前,先行記錄小鼠各項行 為反應測試的數值,包括機械力、輻射熱及冷熱板等,當作本研究的基 礎值(base line)。. 參、發炎疼痛模式 在 1-2 % isoflurane 氣體麻醉下(VIP 3000,Midmark,USA),將 20 µl 生理食鹽水 (pH 7.4,buffered with 20 mM HEPES)加入 3%. 17.
(23) carrageenan (lambda carrageenan,type IV;Sigma),注射在小鼠右後掌以 誘發發炎疼痛模式。. 肆、電針治療 電針治療在 2 % isoflurane 氣體麻醉下進行,針具為 0.5 寸針(32G, 宇光),連接多功能電刺激器(型號:Trio 300,Ito,Japan),針刺穴位 為雙側足三里,針刺身度 2-5 mm,刺激頻率為 2 Hz,刺激強度為 1 mA, 波型為連續性方波,刺激時間持續 15 分鐘,可見小鼠後肢的肌肉有微 小的顫動。 電針治療組的小鼠自注射 carrageenan 的隔天(第 1 天)開始電針治 療,一共連續第 4 天,每天皆接受電針治療 1 次,於第 4 天電針後將小 鼠犧牲。. 伍、行為觀察 在注射 carrageenan 之前及第四天電針後的 0.5-2 小時內,觀察各組 小鼠的行為表現。行為觀察皆在安靜的室溫環境中進行,且皆在小鼠處 於平和的狀態下給予刺激試驗,若小鼠在睡覺或理毛的狀態中則不給予 刺激試驗。 我們藉由以下行為試驗評估小鼠機械性及溫度性痛覺敏感的變化。 一、 機械力試驗 採用針刺回縮反應(von Frey test)。將小鼠置於鐵製網架上,以小型 透明壓克力盒侷限小鼠行動,等小鼠鎮靜後,以電子式的 von Frey filaments(型號:North Coast Medical)的尖端戳刺其右後足掌,儀器可 自動偵測小鼠因機械力致痛而抬起右後足(即縮足反應)的機械性壓力 值,每隻小鼠每天測試 5 次,每次間隔 30 秒,取 5 次測試所得的平均 值,藉以評估小鼠機械性痛覺敏感的閾值。. 18.
(24) 二、 輻射熱試驗 採用 Hargreaves’ test。將小鼠侷限至於無墊料之小型壓克力觀察盒 中,下方為控制溫度 32 oC 之加熱透明玻璃,給予至少 30 分鐘適應環境 後,以可移動式之穩定輻射熱源裝置(輻射熱源強度 30 %,型號:IITC Life Sciences,Series 8,Model 390 G),照射小鼠右後足掌,輻射熱源裝 置可顯示並記錄小鼠經多少秒數後因感覺疼痛而抬足的時間,每隻小鼠 每天測驗 5 次,每次照射不超過 20 秒,兩次照射間隔至少 5 分鐘,防 止造成組織損傷,5 次測試所得的平均值及為當日之潛伏期(latency), 藉此瞭解小鼠對輻射熱的痛覺敏感閾值。 三、 冷熱板試驗 將小鼠侷限至於無墊料之小型壓克力觀察盒中,下方為控制溫度之 金屬面板(型號:Panlab,HARVARD,APPARATUS),透過金屬面 板個別施與恆溫的冷刺激(4 oC)或熱刺激(50 oC),紀錄 5 分鐘內第 一次舔足的時間、縮足反應(withdrawal)的次數、跳離板面(jumping) 的次數,冷板測試加計因冷而站起顫慄(rearing)的次數。每隻小鼠每 天於冷熱板各測試 1 次。藉此瞭解小鼠對溫度性痛覺敏感的閾值及程度。. 陸、動物犧牲 自注射 carrageenan 後的第 4 天行為觀察結束後,以過量麻醉劑水化 氯醛(chloral hydrate,400mg/kg)注射小鼠腹腔,待小鼠麻醉昏迷後, 以仰臥姿勢開啟腹腔曝露出心臟,從左心室插入灌流針頭,另在右心房 剪一切口,以生理食鹽水灌流將體內血液沖洗乾淨。 控制組、鹿角膠組及電針組三個組別中皆有 6 隻小鼠,隨機分配其 中 3 隻作免疫螢光染色法,另外 3 隻作西方墨點分析法。. 柒、免疫螢光染色法(Immunohistochemistry). 19.
(25) 小鼠犧牲後用生理食鹽水將體內血液灌流沖淨,再用 4 %多聚甲醛 (paraformaldehyde)從心臟灌流將身體細胞固定,取下同側腰椎第 3、 4、5 節(L3、4、5)的背根神經節,放入 4 %多聚甲醛(paraformaldehyde) 固定一天,再置入 30 % 蔗糖(sucrose)中約 12 小時,以產生抗凍作用 (cryoprotection),再將已離心沉降的背根神經節組織放入至包埋劑 (O.C.T. Compound)中包埋,並快速冷凍至-20 oC,在低溫恆溫中,使 用冷凍切片機將組織切成 15 微米厚(15 μm)的組織切片。在室溫環境 下,組織切片置於磷酸鹽緩衝液(PBS:phosphate buffer saline,containing 3 % BSA,0.1 % Triton X-100 and 0.02 % sodium azide)中 blocking,於 室溫靜置 120 分鐘後,加入一級抗體 against Nav 1.7 (1:1000,Alomone)、 Nav 1.8 (1:1000,Alomone)及 Nav 1.9 (1:1000,Alomone),放在 4 °C 的環境中隔夜(約 16-24 小時),接著除去一級抗體並用 PBS 清洗後,將 組織切片放在玻片上,再加入螢光標記的二級抗體 6 mM Alexa Flour 594 goat anti-rabbit (Molecular Probes,Carlsbad,CA,USA) 2 小時後,除去 二級抗體並用 PBS 清洗,將組織蓋上玻片,用螢光顯微鏡觀察。. 捌、西方墨點分析法(Western blot analysis) 小鼠犧牲後用生理食鹽水將體內血液灌流沖淨,取下同側腰椎第 3、4、5 節(L3、4、5)的背根神經節,將取下的組織加入均質液(lysis buffer:containing 20 mmol/L imidazole-HCl (pH 6.8),100 mmol/L KCl, 2 mmol/L MgCl2,20 mmol/L EGTA (pH 7.0),300 mmol/L sucrose,1 mmol/L NaF,1 mmol/L sodium vanadate,1 mmol/L sodium molybdate, 0.2% Triton X-100 and proteinase inhibitor cocktail),在 4 oC 的環境中利 用振盪機將組織均質化 1 分鐘。以 13000 轉、10 分鐘高速離心後所得的 上清液即為組織蛋白質萃取液。在每份樣品中取蛋白質萃取液 30 μg, 以 8 %十二烷基硫酸鈉聚丙烯醯胺膠體電泳(SDS-Tris glycine gel electrophoresis),將電泳完成之膠取下,浸泡於 transfer buffer (tris, glycine,SDS,20 % methol)中,再放入 transfer 裝置(mini trans-blot electrophoretic transfer cell,BIO-RAD)將分離出的不同分子量蛋白質轉 20.
(26) 印(transfer)至 PVDF membrane 上,200 mA、120 分鐘。轉印完成後 取出 membrane,再利用含 5 %脫脂奶粉(non-fat milk power)的 TBST buffer (10 mmol/L Tris [pH 7.5],100 mmol/L NaCl,0.1 % Tween 20) blocking 60 分鐘,再加入各亞型的鈉離子通道一級抗體於 4℃隔夜反 應。反應完成後先以 TBST 沖洗三次,每次 10 分鐘,再加入二級抗體 Peroxidase-conjugated anti-rabbit antibody (1:5000)於室溫下作用 2 小時 反應,最後以冷光偵測試劑(ECL-Plus kit,T-Pro biotechnology)及冷 光螢光分析儀(LAS 3000,Fujifilm,Japan)顯影,再以 Image J v1.45i 軟體進行分析。. 玖、統計分析 實驗結果以 SPSS v17.0 軟體進行分析,數值以平均值±標準差(mean ± standard error)表示,當 p 值小於 0.05 時具有統計上的顯著差異。 控制組、發炎組、電針組及偽電針組的實驗數據則以 ANOVA 分析, 事後檢定(post hoc)採 Tukey's test,比較三組之間的療效差異。. 第四章. 結果. 壹、 疼痛行為觀察結果 我們觀察到注射 carrageenan 的右後足可見紅腫現象,類似於前人誘 發發炎疼痛模式的情形[33],此外發炎現象僅見於同側的右後足,對側的 左後足並無發炎現象,同於過去研究所見[34]。. 21.
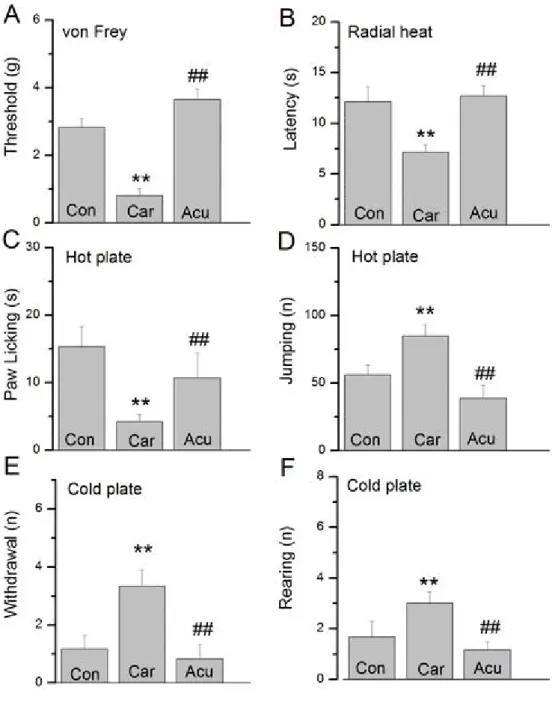
(27) 圖 1 疼痛行為試驗結果 A.電子式 von Frey test。B.輻射熱試驗Radial heat assay。C. 50 oC 熱板試驗之舔足時間(latency time for paw licking)。D. 50 oC熱板試驗 之跳躍次數(jumping)。E. 4 oC冷板試驗之縮足反應(withdrawal)次 數。F. 4 oC冷板試驗之冷而站起顫慄(rearing)次數。Con : 控制組(注 射生理食鹽水),Car:注射carrageenan組,Acu:注射carrageenan並以 22.
(28) 電針足三里治療組。**代表與Con組比較具統計學上的意義(p < 0.01) , ##代表與Car組比較具統計學上的意義(p < 0.01) 。 一、機械力試驗結果 在機械力試驗中,我們採用 von Frey filament test,使用電子式的 von Frey filament 刺激右後足掌,以儀器偵測縮足反應時的機械性壓力值當 作機械性痛覺敏感的閾值。注射生理食鹽水的控制組,並無引起機械性 痛覺敏感的現象(圖 1A,2.82±0.26,n=6),可視為良好的控制組。當我 們檢視注射 carrageenan 的組別時,縮足反應時的機械性壓力值較 control 組明顯降低,可見成功誘發機械性痛覺敏感的情形 (圖 1A,0.81±0.21, n=6,p<0.01),代表注射 carrageenan 誘發的發炎疼痛模式相當成功。接 下來我們觀察注射 carrageenan 並接受電針足三里治療的組別,發現縮足 反應的機械性壓力值在電針後提高,顯示機械性痛覺敏感有明顯降低的 情形 (圖 1A,3.64±0.31,n=6,p<0.01) ,代表電針足三里可降低 carrageenan 發炎模式所引起的機械性痛覺敏感現象,類似的痛覺減輕現象,亦可見 於下列溫度性痛覺敏感的試驗結果。 二、輻射熱試驗結果 接下來我們檢視溫度性痛覺敏感度的試驗結果。在輻射熱試驗中, 注射 carrageenan 的組別第一次抬足時間較 control 組明顯減少(圖 1B, 7.14±0.72 s and 12.1±1.49 s of carrageenan and control,n=6,p<0.01),代 表發炎造成溫度性痛覺敏感的增加。而在注射 carrageenan 並接受電針足 三里治療的組別,可見第一次抬足時間較 carrageenan 組明顯延長,(圖 1B,12.69±0.97,n=6,p<0.01),代表電針足三里可降低 carrageenan 發 炎模式所引起的溫度性痛覺敏感現象。 三、冷熱板試驗結果 溫度性痛覺敏感度的試驗上包括冷熱板試驗。在 50℃熱板的溫度性 痛覺敏感試驗中,我們發現注射 carrageenan 的組別在傷害性高溫刺激 下,第一次舔前足時間較 control 組明顯降低(圖 1C,4.17±1.05,n=6, 23.
(29) p<0.01),代表發炎造成高溫性痛覺敏感的增加。而接受電針治療的組 別,第一次舔足時間較 carrageenan 組明顯明顯延長(圖 3C,10.67±3.69, n=6,p<0.01)。而相似的試驗結果可見於跳躍(jumping)次數的分析中, 在注射 carrageenan 的組別跳躍次數較 control 組明顯增加(圖 1D, 84.83±8.37,n=6,p<0.01),電針治療組的跳躍次數則較 carrageenan 組 明顯減少(圖 1D,38.67±9.49,n=6,p<0.01)。代表電針足三里可降低 carrageenan 發炎模式所引起的高溫性痛覺敏感現象。 同樣的在 4℃冷板的溫度性痛覺敏感試驗中,我們發現注射 carrageenan 的組別在傷害性低溫刺激下,縮足反應的次數較 control 組 明顯增加(圖 1E,3.33±0.56,n=6,p<0.05),代表發炎造成低溫性痛覺 敏感的增加,而在電針治療的組別,縮足反應的次數則較 carrageenan 組明顯降低,(圖 1E,0.83±0.48,n=6,p<0.05)。相似的結果亦見於小 鼠 rearing 的次數觀察中(圖 1F)。代表電針足三里可降低 carrageenan 發炎模式下所引起的低溫性痛覺敏感現象。 根據我們行為的觀察結果,電針足三里可降低 carrageenan 發炎模式 下所引起的機械性及溫度性痛覺敏感現象。因此我們試著進一步證實鈉 離子通道的表現是否會受到電針治療的影響。. 24.
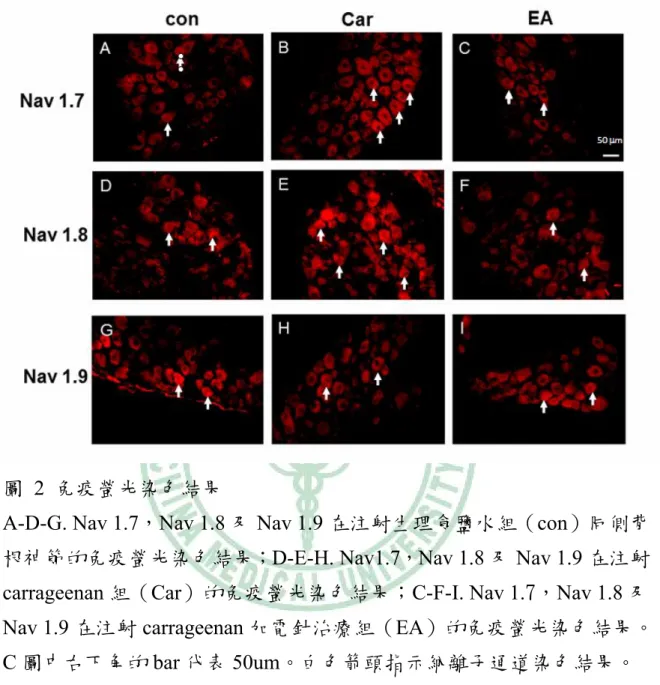
(30) 貳、免疫螢光染色結果. 圖 2 免疫螢光染色結果 A-D-G. Nav 1.7,Nav 1.8 及 Nav 1.9 在注射生理食鹽水組(con)同側背 根神節的免疫螢光染色結果;D-E-H. Nav1.7,Nav 1.8 及 Nav 1.9 在注射 carrageenan 組(Car)的免疫螢光染色結果;C-F-I. Nav 1.7,Nav 1.8 及 Nav 1.9 在注射 carrageenan 加電針治療組(EA)的免疫螢光染色結果。 C 圖中右下角的 bar 代表 50um。白色箭頭指示納離子通道染色結果。 免疫螢光染色下可見 Nav 1.7 的蛋白質表現在控制組的背根神經節 中(圖 2A)。Nav 1.7 的表現在 carrageenan 組(圖 2B)較 control 組有增加趨 勢,此項結果類似於 Blake 等人的研究結果[27]。值得注意的是當我們進 一步使用電針足三里治療後,Nav 1.7 的表現較 carrageenan 組明顯減少 (圖 2C)。 皆下來我們檢視 Nav 1.8 的表現。免疫螢光染色下同樣可見 Nav 1.8 的蛋白質表現在控制組的背根神經節中(圖 2G)。Nav 1.8 的表現在 25.
(31) carrageenan 組(圖 2E)較 control 組亦有增加趨勢,此項結果同樣類似於 Blake 等人的研究結果[27]。重要的是當我們進一步使用電針足三里治療 後, Nav 1.8 的表現亦明顯減少(圖 2F)。以上結果顯示在 carrageenan 的 發炎模式下,電針足三里能有效的調降 Nav 1.7、Nav 1.8 的過度表現。 當我們繼續檢視Nav 1.9的表現時,發現免疫螢光染色下仍然可見 Nav 1.9的蛋白質表現在控制組的背根神經節中(圖2G)。但Nav 1.9的表現 在carrageenan組與控制組比較並無明顯變化(圖2H),代表Nav 1.9在 carrageenan的發炎模式中並無顯著的免疫螢光染色變化。同樣的結果亦 見於電針治療組中,Nav 1.9的蛋白質表現在電針治療後亦無明顯的改變 (圖2I)。. 參、西方墨點分析法結果 皆下來我們進一步採用西方墨點分析法,分析 Nav 1.7、Nav 1.8 與 Nav 1.9 的蛋白質含量在發炎模式下經電針治療後的表現變化。. 26.
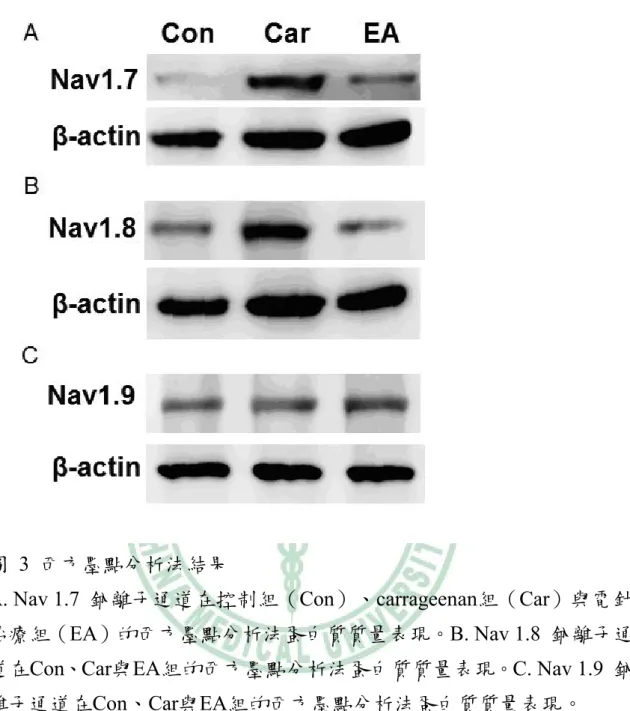
(32) 圖 3 西方墨點分析法結果 A. Nav 1.7 鈉離子通道在控制組(Con)、carrageenan組(Car)與電針 治療組(EA)的西方墨點分析法蛋白質質量表現。B. Nav 1.8 鈉離子通 道在Con、Car與EA組的西方墨點分析法蛋白質質量表現。C. Nav 1.9 鈉 離子通道在Con、Car與EA組的西方墨點分析法蛋白質質量表現。. 27.
(33) 表 1 西方墨點分析法-濃度定量分析. Quantification of Navs expression band densities (mean ± SEM, n=6). con. Car. Car+EA. Mean changes. Nav1.7. 100.0±0%. 129.9±10.2%*. 110.8±3.4%#. -19.1%a. Nav1.8. 100.0±0%. 122.8±4.7%*. 104.7±2.0%#. -18.1%a. Nav1.9. 100.0±0%. 87.1±15.0%. 100.7±5.0%. 13.6%. *p. < 0.05, compared to baseline; #p < 0.05, comparison between inflammation and EA groups. a Nav1.7 and Nav1.8 expression levels were attenuated by EA versus carrageenan inflammatory group; bNav1.7 and Nav1.8 expression levels were attenuated by EA versus CFA-induced inflammatory group. 2. 依據西方墨點分析法,我們發現在控制組的同側背根神經節中可發現 鈉離子通道的蛋白質表現。Nav 1.7 在 carrageenan 的發炎模式下,西方 墨點分析法的蛋白質質量較 control 組有增加的趨勢,且在電針治療後蛋 白質含量較 carrageenan 組明顯下降 (圖 3A,表 1 exhibited 19.1% decrease in signal compared with inflamed group,n=6,p<0.05)。相似的結果亦表 現在 Nav 1.8,Nav 1.8 的蛋白質在電針治療後亦有明顯下降趨勢 (圖 3B,表 1 exhibited 18.1% decrease in signal compared with inflamed group,n=6,p<0.05)。但 Nav 1.9 在 carrageenan 的發炎模式下,西方墨 點分析法的蛋白質含量較 control 組並沒有增明顯變化,且在電針治療後 蛋白質含量較 carrageenan 組亦無明顯改變 (圖 3C,表 1 exhibited 13.6% increase in signal compared with inflamed group,n=6,p>0.05)。 因此根據我們螢光免疫染色和西方墨點分析法的研究結果發現, Nav 1.7與Nav 1.8在carrageenan的發炎模式下,於同側的背根神經節中有 明顯增加的趨勢,但Nav 1.9則無明顯變化,代表Nav 1.7、Nav 1.8與發 28.
(34) 炎反應相關,但與Nav 1.9的蛋白質表現則無關。值得關注的是在電針治 療後,Nav 1.7與Nav 1.8的蛋白質表現有明顯減少的趨勢,但Nav 1.9則 無明顯變化,代表電針足三里能有效降低carrageenan發炎模式中過度表 現的Nav 1.7與Nav 1.8,但與Nav 1.9的蛋白質表現則無明顯相關性。 综合以上研究結果我們推測,電針足三里減輕發炎疼痛的機轉,或 許和調控 Nav 1.7 與 Nav 1.8 的表現相關,但與 Nav 1.9 則無關。. 第五章. 討論. 本研究發現在 carrageenan 所誘發的發炎疼痛模型下,經電針足三里 穴位治療後,觀察小鼠的疼痛行為表現,可見機械性及溫度性痛覺敏感 程度明顯減少。並在免疫螢光染色法及西方墨點分析法的研究結果中顯 示,電針足三里可降低因發炎引起的 Nav 1.7 及 Nav 1.8 過度表現現象。 週邊痛覺神經有許多不同的受體及離子通道,可感應不同的刺激並 轉換成電位衝動,電位衝動繼而引發一連串的動作電位,以將訊息從周 邊傳遞至中樞神經系統[1]。痛覺神經元表現著多種離子通道,例如鈣離 子通道(Cav:voltage-gated calcium channel),在發炎或神經損傷的動 物模型中,Cav 2.2 或 3.2 功能缺失的小鼠表現對機械力及溫度刺激的敏 感度降低[7],鈣離子通道已廣泛被應用在抗痙攣藥中,也被應用在疼痛 的治療上。鉀離子通道則與動作電位中的再極化階段相關,鉀離子通道 多扮演減緩神經興奮的角色[1]。鈉離子通道為哺乳動物細胞膜上的跨膜 蛋白,通道的開關與否,能讓鈉離子快速的出入細胞內外之間,產生細 胞膜電位的變化以發生動作電位傳遞訊息[7]。鈉離子通道與神經的興奮 性有關,尤其是在初級傳入神經元的痛覺傳遞上扮演了重要的角色。當 周邊組織受傷發炎後,週邊痛覺神經或中樞神經變得敏感而興奮性增 加,導致痛覺敏感的情形[4]。近來相當多研究陸續發現鈉離子通道與發 炎疼痛的相關性[5],就目前所知,鈉離子通到一共有 10 個分型,研究指 出其中 Nav 1.7、Nav 1.8 及 Nav 1.9 與疼痛具有相關性[22]。 我們的研究結果指出,在注射 carrageenan 的四天後,Nav 1.7、Nav 1.8 的表現皆有增加現象,Nav 1.9 則無明顯變化。值得注意的是電針足 29.
(35) 三里可明顯調降 Nav 1.7 及 Nav 1.8 在發炎模式中的過度表現,但 Nav 1.9 則無顯著性差異。因此我們根據研究結果推論電針足三里得以減輕發炎 疼痛的療效機轉,和調控鈉離子通道中 Nav 1.7 與 Nav 1.8 的表現相關, 但與 Nav 1.9 則無關。 我們的研究步驟相似於Black於2004年所作的實驗[27],Black等人採 用SD大鼠,在右後足掌注射4% carrageenan誘發發炎疼痛,並在同隻大 鼠的左後足掌注射生理食鹽水,控制組為右後足掌沒有注射carrageenan ,但仍在左後足掌注射生理食鹽水。第4天後取下同側第4、5腰椎的背 根神經節,結果發現在同側背根神經節中可發現大量鈉離子通道蛋白質 的表現,但在對側背根神經節及控制組中則無此現象。Black等人同樣採 用免疫螢光染色法及西方墨點分析法,觀察各型鈉離子通道的在發炎模 式下的變化,結果發現在carrageenan的發炎疼痛模式下,Nav 1.7與Nav 1.8的免疫螢光染色明顯增加,但Nav 1.9則無顯著性差異。進一步使用 西方墨點分析法的定量濃度分析發現,Nav 1.7和Nav 1.8在carrageenan 的注射下,定量濃度分析較控制組增加了12-25%,與我們的研究結果顯 示Nav 1.7的定量蛋白質濃度分析增加19.1%,以及Nav 1.8增加18.1%有 相類似的研究結果,顯示Nav 1.7和Nav 1.8與發炎疼痛有密切相關性。與 我們研究結果相似的部分還可見到Nav 1.9的定量蛋白質濃度分析在 carrageenan的發炎疼痛模式與控制組比較並無明顯變化,顯示Nav 1.9與 carrageenan所誘發的疼痛較無相關。但不同的是,我們在發炎疼痛的模 式中加入了傳統的電針治療,可進一步探討電針止痛與鈉離子通道的關 係。 Nav 1.7屬於TTX-S type[23],Nav 1.7表現在C纖維的神經末梢中且在 痛覺訊息傳遞中扮演中要角色[35,36]。基因剔除或基因抑制的技術亦運用 在目前的鈉離子通道研究中,觀察基因剔除或抑制的老鼠有哪些症狀的 變化,就可以進一步了解鈉離子通道所扮演的角色。近來在基因剔除及 基因抑制的小鼠實驗中,強烈證實阻斷鈉離子通道是相當有潛力的止痛 方式[37,38]。Nassar等人於2004年亦發表Nav 1.7基因抑制的小鼠在 Hargreaves’ test、hot-plate、Randall-Selitto test及von Frey test中,溫度痛 覺及機械性痛覺的閾值增加。另外再以formalin、carrageenan、CFA及 30.
(36) NGF注射Nav 1.7基因抑制的小鼠,觀察到溫度痛覺敏感及機械力痛覺異 常等疼痛行為皆有下降趨勢[37,38]。此結果和Yeomans等人在2005發表Nav 1.7基因抑制小鼠在CFA所誘發的發炎模式中,發炎疼痛的敏感度降低相 符合[39]。根據上述眾多研究可發現Nav 1.7與發炎性疼痛有高度相關性。 Nav 1.7基因剔除的小鼠在發炎疼痛模式中疼痛減輕的現象尤其明顯 [39]. 。此外,以Nav 1.7為標的的藥物能有效的治療多種類型的疼痛,例如. benzazepinone和imidazopyridine皆是透過阻斷Nav 1.7通道而達到疼痛治 療的效果[41]。而我們運用傳統的電針方式達到止痛的效果,並證實電針 能有效調控Nav 1.7鈉離子通的表現,因此根據我們的結果推論,電針治 療carrageenan發炎疼痛模式的止痛機轉,或許和調降Nav 1.7鈉離子通的 表現相關。 Nav 1.8屬於TTX-R type[42],在Black等人於2004年所作研究中,Nav 1.8的免疫螢光染色及西方墨點分析在carrageenan發炎疼痛模式皆有顯 著的增加,顯示Nav 1.8與發炎疼痛有密切相關性。在Nav 1.8基因剔除的 研究中發現,小鼠對傷害性低溫及機械力無反應[42],或對機械性痛覺異 常的表現有減輕趨勢[43]。另有Nav 1.8基因抑制的研究發現,小鼠對發炎 疼痛的行為表現減少,且對低溫的敏感度亦降低[44] 。Akopian等人在 1999年探討Nav 1.8功能被剔除的小鼠在carrageenan所誘發的發炎模式 中的行為表現,結果發現Nav 1.8功能被剔除的小鼠對非傷害性的高溫及 壓力失去敏感度,對低溫的敏感度有降低的現象,且對傷害性機械力的 刺激行為反應減少[45]。Nassar等人於2004年亦發現在發炎模式下,Nav 1.8基因剔除的小鼠可見TTX-sensitive代償電流的增加,同時發現Nav 1.7 mRNA表現增加,顯示Nav 1.7可代償 Nav 1.8基因剔除的小鼠的功能, 推論Nav 1.7與Nav 1.8在疼痛機轉的角色上彼此具有相關性[37]。但1999 年Akopian等人在Nav 1.7基因剔除的小鼠沒有發現TTX-resistant代償電 流的增加[45],已上研究證實Nav 1.8與發炎性疼痛的高度相關性。此外目 前已有二種Nav 1.8的阻斷劑進入試驗性治療中,其中之一是A-803467, 乃是對Nav 1.8有高度選擇性的阻斷劑,已經過實驗證實能有效降低大鼠 的發炎性疼痛[46]。另一個是MrVIB,從海洋錐形蝸牛分離出的 μO-conotoxin peptide是一種有效且具高度選擇性的Nav 1.8電流阻斷 31.
(37) 劑,研究證實能有效抑制感覺神經元的動作電位,並影響急性機械性疼 痛、發炎性疼痛、神經痛等多種動物疼痛模式[47]。我們的研究想要進一 步檢視電針止痛與Nav 1.8之間的關係性,研究結果發現,Nav 1.8在注射 carrageenan之後表現量會明顯增加,經電針治療後發現Nav 1.8的表現有 顯著降低,因此我們推測電針減輕發炎性疼痛的機轉和Nav 1.8相關。 Nav 1.9屬於TTX-R type[23],Nav 1.9亦多位在背根神經節的細神經纖 維神經元中,不同於Nav 1.7與Nav 1.8,Nav 1.9與靜止膜電位較有相關 性[19]。有文獻指出在鹿角膠誘發的發炎痛模型中,背根神經節中Nav 1.9 mRNA及蛋白質並無明顯變化,但在CFA誘發的發炎痛模型中,Nav 1.9 mRNA有明顯增加情形[20]。在Black於2004年的研究中發現注射 carrageenan並不影響背根神經節中Nav 1.9 mRNA或蛋白質的表現[27]。 Nassar在2004年Nav 1.9基因剔除的動物實驗中發現,NaV1.9在疼痛閾值 設定中扮演重要角色,但與動作電位無關[37],以上眾多文獻指出Nav 1.9 與發炎疼痛的相關機轉關尚未明確。但在我們的研究結果發現,Nav 1.9 的表現在注射carrageenan之後並無明顯增加趨勢,且經電針治療後觀察 Nav 1.9的表現亦無顯著變化,因此我們推測電針減輕發炎性疼痛的機轉 和Nav 1.9較無相關性。其它研究發現Nav 1.9基因剔除小鼠在內臟機械力 敏感度降低[21],Nav 1.9在腸繫膜神經元(myenteric neurons)的表現較 為明顯並與腸神經反射和腸胃症候較為相關[48]。 Wei Wang等人在2011一篇回顧性的研究指出,Nav 1.7在carrageenan 及CFA的發炎疼痛模式中可見Nav 1.7 mRNA及蛋白質的增加,並在Nav 1.7基因剔除及抑制的動物中發現發炎疼痛的減輕;Nav 1.8僅在 carrageenan的發炎疼痛模式中可見Nav 1.8 mRNA及蛋白質的增加,在 Nav 1.8基因剔除及抑制的動物中皆可發現發炎疼痛的減輕;在Nav 1.9 基因剔除的動物雖然發現與發炎性疼痛閾值相關,但在carrageenan的發 炎疼痛模式中,Nav 1.9 mRNA及蛋白質的變化並不明顯,僅在CFA的發 炎疼痛模式中,Nav 1.9 mRNA有增加趨勢. [8]. 。. 根據我們研究結果發現,在carrageenan發炎疼痛模式中,電針足三 里能有效減輕機械性及溫度性痛覺敏感現象,進一步以免疫螢光染色和 西方墨點分析法研究鈉離子通道的結果發現,Nav 1.7與Nav 1.8在 32.
(38) carrageenan的發炎模式下,於同側的背根神經節中有明顯增加的趨勢, 但Nav 1.9則無明顯變化,代表Nav 1.7、Nav 1.8與發炎疼痛相關,但與 Nav 1.9則無關。值得關注的是在電針治療後,Nav 1.7與Nav 1.8有明顯 減少的趨勢,但Nav 1.9則無明顯變化,代表電針足三里能有效降低 carrageenan發炎模式中Nav 1.7與Nav 1.8的過度表現,但與Nav 1.9則無明 顯相關性。 综合以上研究結果我們推測,電針足三里減輕發炎疼痛的療效機 轉,或許和調控 Nav 1.7 與 Nav 1.8 的表現相關,但與 Nav 1.9 則較無相 關性。. 第六章 結論 本研究在 carrageenan 所誘發的發炎疼痛模型下,探討以電針足三 里減輕小鼠發炎性疼痛與鈉離子通道之間的相關機轉。在電針足三里 後,藉由觀察小鼠的疼痛行為,可見機械性及溫度性痛覺敏感程度明顯 減少;在免疫螢光染色法及西方墨點分析法的研究結果顯示,電針可明 顯調降 Nav1.7 及 Nav1.8 在發炎模式中的過度表現,但 Nav1.9 則無顯著 性差異。因此我們根據研究結果推論電針足三里減輕發炎疼痛的療效機 轉,和調控鈉離子通道中 Nav 1.7 與 Nav 1.8 的表現相關,但與 Nav 1.9 則較無相關性。. 第七章. 參考文獻. 1. Foulkes T, Wood JN. Pain genes. PLoS genetics. 2008;4(7):e1000086. 2. Sharif-Naeini R, Basbaum AI. Targeting pain where it resides ... In the brain. Sci Transl Med. 2011 12;3(65):65. 3. Mogil JS. Animal models of pain: progress and challenges. Nature reviews Neuroscience. 2009;10(4):283-294. 4. Lakhmir S Chawla, Amirali Nader, Todd Nelson, Trusha Govindji, Ryan Wilson, Sonia Szlyk, Aline Nguyen,Christopher Junker, Michael G 33.
(39) Seneff. Utilization of base deficit and reliability of base deficit as a surrogate for serum lactate in the peri-operative setting. BCM Anesthesiol. 2010 Sep 9;10(1):16 5. Walder RY, Gautam M, Wilson SP, Benson CJ, Sluka KA. Selective targeting of ASIC3 using artificial miRNAs inhibits primary and secondary hyperalgesia after muscle inflammation. Pain. 2011;152(10): 2348-2356. 6. Dubner KRaR. Inflammatory Models of Pain and Hyperalgesia. Animal Models of Pain. 1999;40(3):111-118. 7. Basbaum AI, Bautista DM, Scherrer G, Julius D. Cellular and molecular mechanisms of pain. Cell. 2009;139(2):267-284. 8. Wang W, Gu J, Li YQ, Tao YX. Are voltage-gated sodium channels on the dorsal root ganglion involved in the development of neuropathic pain? Molecular pain. 2011;7:16. 9. Basbaum AI, Bautista DM, Scherrer G, Julius D. Cellular and Molecular Mechanisms of Pain. 2009;139(2):267-284. 10. Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature. 2001 Sep 13;413(6852):203-210. 11. 林佑穗、袁宗凡編譯:Guyton & Hall 新編醫用生理學. 2005:609-612. 12. KA M. Assessingmechanicalallodynia in the ratpaw with anewelectronicalgometer. Journal of Neuroscience Methods. 1896;84 (1-2):41-47. 13. K. Hargreaves RD, F. Brown, C. Flores, J. Joris. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain. 1988;32(1):77-88. 14. Eddt NB, Touchberry CF,Lieberman JE. Synthetic analgesics; methadone isomers and derivatives J Pharmacol Exp Ther. 1950 Feb;98(2):121-137. 15. Gary J, Bennett Y-KX. Aperipheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988; 34.
(40) 33( 1):87-107. 16. Mogil JS. Animal models of pain:progress and challenges. Nat Rev Neurosci. 2009 Apr;10(4):283-294. 17. Zhao ZQ. Neural mechanism underlying acupuncture analgesia. Progress in neurobiology. 2008;85(4):355-375. 18. 黃維三:針灸科學,2000:265-275. 19. 林昭庚:新編採圖針灸學,1999:739-754. 20. Chen J-GLW. acupuncture analgesia in clinical trials. American journal of chinese medicine. 2009;37(1):1-18. 21. Mark D, Baker and John N,Wood. Involvement of Na+ channels in pain pathway. TRENDS in Pharmacological Sciences. 2001;22(1) :27-31. 22. Momin A, Wood JN. Sensory neuron voltage-gated sodium channels as analgesic drug targets. Current opinion in neurobiology.2008;18(4): 383-388. 23. Cummins TR, Sheets PL, Waxman SG. The roles of sodium channels in nociception: Implications for mechanisms of pain. Pain. 2007;131(3): 243-257. 24. Rush AM, Cummins TR, Waxman SG. Multiple sodium channels and their roles in electrogenesis within dorsal root ganglion neurons. The Journal of physiology. 2007;579(Pt 1):1-14. 25. Gold MS, Reichling DB, Shuster MJ,Levine JD. Hyperalgesic agents increase a tetrodotoxin-resistant Na+ current in nociceptor. Proc Natl Acad Sci USA. 1996 Feb 6;93(3):1108-12. 26. Zhang YH, Vasko MR, Nicol GD. Ceramide, a putative second messenger for nerve growth factor, modulates the TTX-resistant Na+ current and delayed rectifier K+ current in rat sensory neurons. The Journal of physiology. 2002;544(2):385-402. 27. Black JA, Liu S, Tanaka M, Cummins TR, Waxman SG. Changes in the expression of tetrodotoxin-sensitive sodium channels within dorsal root ganglia neurons in inflammatory pain. Pain. 2004;108(3):237-247. 35.
(41) 28. Strickland IT, Martindale JC, Woodhams PL, Reeve AJ, Chessell IP, McQueen DS. Changes in the expression of NaV1.7, NaV1.8 and NaV1.9 in a distinct population of dorsal root ganglia innervating the rat knee joint in a model of chronic inflammatory joint pain. Eur J Pain. 2008;12(5):564-572. 29. Leffler A HR, Dib-Hajj SD, Waxman SG, Cummins TR. Pharmacological properties of neuronal TTX-resistant sodium channels and the role of a critical serine pore residue. Pflugers Arch. 2005;451(3): 454-463. 30. Tate S BS, Hick C, Trezise D, John V, Mannion RJ, et al. . Two sodium channels contribute to the TTX-R sodium current in primary sensory neurons. Nat Neurosci. 1998;1:653-655. 31. Amaya F, Wang H, Costigan M, Allchorne AJ, Hatcher JP, Egerton J, Stean T, Morisset V, Grose D, Gunthorpe MJ, Chessell IP, Tate S, Green PJ, Woolf CJ. The voltage-gated sodium channel Na(v)1.9 is an effector of peripheral inflammatory pain hypersensitivity. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2006;26(50):12852-12860. 32. Priest BT, Murphy BA, Lindia JA, Diaz C, Abbadie C, Ritter AM, Liberator P, Iyer LM, Kash SF, Kohler MG, Kaczorowski GJ, MacIntyre DE, Martin WJ. Contribution of the tetrodotoxin-resistant voltage-gated sodium channel NaV1.9 to sensory transmission and nociceptive behavior. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(26):9382-9387. 33.Tanaka M CT, Ishikawa K, Dib-Hajj SD, Black JA, Waxman SG. SNS Na+ channel expression increases in dorsal root ganglion neurons in the carrageenan inflammatory pain model. Neuroreport. 1998;20;9(6): 967-972. 34. Yen YT, Tu PH, Chen CJ, Lin YW, Hsieh ST, Chen CC. Role of acid-sensing ion channel 3 in sub-acute-phase inflammation. Molecular 36.
(42)
數據

相關文件
臺大機構典藏NTUR (National Taiwan University 二 Repository, http://ntur.lib.ntu.edu.tw) 經驗與協助推 動臺灣學術機構典藏TAIR (Taiwan Academic Institutional Repository,
The functional fall of the salivary secretion observed at 180 days post irradiation was not only associated with a reduction of gland mass but also to an alteration of the
Introduction: Eagle’s syndrome is a rare condition that refers to chronic recurrent pain in the oropharynx, face, and neck due to elongation of the styloid process or calcification
A prospective study of magnetic resonance and radiographic imaging in relation to symptoms and clinical findings of the temporomandibular joint in children with juvenile
The chairman/representative director, and at least 2/3 of the board of directors of a limited company or a company limited by shares providing the above services shall be the Republic
Chiun-Chuan Chen (National Taiwan University, NTU) Jann-Long Chern (National Central University, NCU) Yung-Fu Fang (National Cheng Kung University, NCKU) Yu-Chen Shu (National
For 5 to be the precise limit of f(x) as x approaches 3, we must not only be able to bring the difference between f(x) and 5 below each of these three numbers; we must be able
[This function is named after the electrical engineer Oliver Heaviside (1850–1925) and can be used to describe an electric current that is switched on at time t = 0.] Its graph
