LPS、PGN刺激巨噬細胞產生TNFα 是透過iNOS所調控的sustained ERK activation; Sustained ERK activation mediated by iNOS is required for LPS- and PGN-induced TNFα
77
0
0
全文
(2) 目錄 目錄 ........................................................................................................................... I 致謝 ........................................................................................................................ IV ABSTRACT ........................................................................................................... VI 中文摘要 ...............................................................................................................VII 壹、. 序論 (INTRODUCTION) ......................................................................... 1. 第一節. 免疫系統 (IMMUNE SYSTEM) 與巨噬細胞 (MACROPHAGES) ............ 2. 第二節. 內毒素 (ENDOTOXIN) ........................................................................ 2. 第三節. TOLL-LIKE RECEPTOR (TLR) ............................................................ 4. 第四節. INOS (INDUCIBLE NITRIC OXIDE SYNTHASE) ...................................... 5. 第五節. SRC KINASE ........................................................................................ 6. 第六節. 活化蛋白質激酶 (MITOGEN-ACTIVATED PROTEIN KINASE,MAPK. KINASE). ………………………………………………………………………….8. 貳、. 實驗材料及方法 ....................................................................................... 10. 第一節. 實驗設計流程 .................................................................................. 11. 第二節. 實驗材料 .......................................................................................... 12. 第三節. 實驗方法 .......................................................................................... 16. 參、 一、. 結果 (RESULTS)..................................................................................... 32 AG 及 ODQ 皆可同步抑制 PGN 所產生的 TNF,且 SNAP 及. 8-BR-CGMP 處理 RAW 264.7 巨噬細胞,皆可增加 TNF的釋放。 ............. 33 二、. SNAP 與 8-BR-CGMP 可誘導 INOS KNOCKOUT MICE PERITONEAL. MACROPHAGE. 產生 TNF ................................................................................ 34 I.
(3) 三、. AG 及 ODQ 皆能有效抑制 PGN 誘導 ERK 的活化 .......................... 35. 四、. LPS 與 PGN 誘導 ERK 的活化有兩個不同的階段............................ 36. 五、. LPS 與 PGN 所調控的 SUSTAINED ERK 的活性是會受 AG 的影響 . 36. 六、. INOS. 七、. SRC FAMILY KINASES 參與在 LPS 與 PGN 所誘導的 SUSTAINED ERK. 為 PGN-MEDIATED SUSTAINED ERK ACTIVATION 所需 ............ 37. ACTIVATION。..................................................................................................... 八、. 38. SRC 參與在 LPS、PGN 與 SNAP 所誘導巨噬細胞的 SUSTAINED ERK. ACTIVATION 及. TNFPRODUCTION。 .............................................................. 39. 肆、. 討論 ( DISCUSSION )............................................................................. 41. 伍、. 參考文獻(REFERENCE)......................................................................... 46. 圖表目錄 陸、. 圖表(FIGURE) ......................................................................................... 52. FIGURE 1. NO/CGMP IS INVOLVED IN PGN-INDUCED TNF SECRETION. ........ 53 FIGURE 2. INOS IS REQUIRED FOR TNFSECRETION MEDIATED BY LPS AND PGN. .................................................................................................................. 55 FIGURE 3. PGN-INDUCED ERK ACTIVATION IS INHIBITOR BY AG OR ODQ. ..... 56 FIGURE 4. SUSTAINED ERK ACTIVATION MEDIATED BY LPS AND PGN. ........... 57 FIGURE 5. LPS- AND PGN-MEDIATED SUSTAINED ERK ACTIVATION WAS AG-SENSITIVE. ................................................................................................... 58 FIGURE 6. INOS IS INVOLVED IN PGN-MEDIATED ERK ACTIVATION. ............... 59 FIGURE 7. INVOLVEMENT OF SRC FAMILY KINASES IN THE LATE STAGE OF LPSAND PGN-INDUCED TNF SECRETION AND ERK ACTIVATION. .......................... FIGURE 8. LPS- AND PGN-MEDIATED TNF SECRETION WAS INHIBITED BY II. 60.
(4) SRC-SPECIFIC SIRNA. .......................................................................................... 61. FIGURE 9. LPS- AND PGN-MEDIATED TNF SECRETION WAS INHIBITED BY SRC-SPECIFIC SIRNA, WHICH COULD BE REVERSED BY ECTOPIC SRC. ................ 62. FIGURE 10. SNAP MEDIATED TNF SECRETION WAS INHIBITED BY SRC-SPECIFIC SIRNA BUT ECTOPIC SRC COULD REVERSE THIS PHENOMENON. ......................... 63. FIGURE 11. THE PROPOSED NO/SGC/SRC/ERK SIGNALING PATHWAY FOR TNF SECRETION IN LPS- AND PGN- STIMULATED MACROPHAGES. ............... 64 附圖一、STRUCTURE OF LIPOPOLYSACCHARIDE AND PEPTIDOGLYCAN. ........... 65 附圖二、LPS-INDUCED TNF TRANSCRIPT IS SENSITIVE TO INHIBITORS OF INOS AND SGC. ............................................................................................................ 66. 附圖三、MEK INHIBITOR PD98059 SUPPRESSED THE SNAP- AND 8-BR-CGMP-INDUCED TNFPRODUCTION. ...................................................... 67 附圖四、LPS-MEDIATED ERK ACTIVATION WAS ABROGATED IN INOS NULL MACROPHAGES, WHICH COULD BE RESTORED BY SNAP AND 8-BR-CGMP. ........ III. 68.
(5) 致謝 兩年的時間一眨眼就匆匆過去了,在這期間我從一個對生命科學 領域懵懂無知的大學生,變成即將畢業的研究生。回顧這兩年走來的 路程,內心充滿幸福與感謝,。首先要感謝的是我的指導老師馬明琪 老師,因為有老師總是把我們當作自己的小孩,無私的付出、拉拔以 及帶領我們在學涯大海中朝著正確的方向前進,也時常告誡我們做事 要嚴謹、要有計劃,才能達到事半功倍的功效。還有我的共導老師張 乃文老師,因為動物實驗的機緣才有機會跟您學習許多實驗上以及學 術上的知識與技巧,不厭其煩的抽出時間教導我動物技術,回顧一路 走來,兩位老師的指導與敦敦教誨都深深的印在我心中。感謝已經畢 業的佳融學長、宜倫學姊、郁芸學姊、冠妤學姊、還有妙瑩學姊,在 我剛開始接觸這個領域時,給我許多實驗技術上以及知識上的指導, 即使在你們畢業後,也不時的給予我們許多幫助。感謝一路與我走來 的青昭,雖然不同系所但同在馬老師的實驗室裡一起成長,有著許多 甘苦與共的回憶。班上的同學俞凱、姵岑以及基醫所的佳嫻、光荻、 承雋、欣珊、政澤、松源、凡妮、碧玉、沐恩、志豪、佩怡,因為有 你們的陪伴才有許多的回憶。永國與瑞青,雖然只有一年的相處,但 也因為你們的出現,讓實驗室熱鬧許多。奇君、倉裕、容慈、巧雯、. IV.
(6) 佩珊、景程、玉鳳,在第二年裡有了你們的參與讓生活更加熱鬧。還 有麗如姊與所秘育青,在學涯中幫了我許多忙。以及所有幫助過我的 貴人,因為有了你們,才能讓我不斷成長。最後要感謝的就是我的父 母,因為他們無私的付出與支持,才能讓我無憂無慮的追求我的興趣, 一路走來到今天。. V.
(7) Abstract Toll-like receptors (TLRs) are important for macrophages to recognize foreign insults and induce the secretion of proinflammatory cytokines such as tumor necrosis factor-alpha (TNF). While TLR4 can recognize lipopolysaccharide (LPS), a Gram-negative bacterial component; TLR2 acts as a signaling receptor for peptidoglycan (PGN), a major component of the cell wall of Gram-positive bacteria. Concomitant induction of nitric oxide synthase (iNOS) and TNF in LPS- and PGN-stimulated macrophages raised the possibility that iNOS was involved in TNFgeneration. Indeed, pharmacological blockade or knockout of iNOS reduced LPS- and PGN-mediated TNF secretion. Meanwhile, SNAP (a NO donor) or 8-br-cGMP (a cGMP analogue) induced the production of TNF in both wild type and iNOS null macrophages. Since (1) LPS- and PGN-increased TNF generation was PP2-sensitive, (2) NO could upregulate the expression of Src, and (3) ERK was required for transcription of tnf gene; therefore, one might wonder NO-mediated Src enhancement was involved in generation of TNF in macrophages exposed to LPS and PGN. Indeed, LPS-, PGN-, SNAP-, and 8-br-cGMP-mediated activation of ERK as well as induction of TNF were greatly suppressed in macrophages treated with PD98059 (the MEK inhibitor) and attenuation of Src by src specific siRNA reduced LPS- and PGN-evoked ERK activation and TNF release in Raw264.7 macrophages and reintroduction of siRNA-resistant Src could reverse these events. With these results, the strong association between ERK activation and TNFsecretion in macrophages devoid of iNOS exposed to LPS, PGN, SNAP and 8-br-cGMP indicated that via activation of Src and ERK, iNOS was required for LPS-and PGN-elicited TNF generation.. VI.
(8) 中文摘要. Toll-like receptors (TLRs)對於巨噬細胞來說扮演著重要的角色, 其主要功能有接受外來的訊息以及誘發發炎相關的激素,如 tumor necrosis factor-alpha (TNF)。TLR4 可辨識 lipopolysaccharide (LPS)、 革蘭氏陰性菌的細胞壁成分之一,TLR2 則被 peptidoglycan (PGN)活 化,革蘭氏陽性菌細胞壁的主要成分之一。在先前的研究中認為由 LPS 與 PGN 刺激巨噬細胞所產生的 iNOS 與 TNF是並行互不干擾的, 但 iNOS 有可能參與在調控 TNF基因的路徑中。透過藥物的抑制以 及基因剃除 iNOS 的表現的確使 LPS 與 PGN 刺激巨噬細胞產生 TNF 的釋放量降低,另一方面,SNAP (a NO donor) or 8-br-cGMP (a cGMP analogue)皆可以促使 WT 或是 iNOS null mice 巨噬細胞的 TNF表現 量增加。(1) PP2 可以抑制 LPS 或 PGN 刺激巨噬細胞產生 TNF,(2) NO 可使 Src 的表現量增加,(3) TNF基因的轉錄是需要 ERK,上述 的原因,存在著一個可能性, NO 促進 Src 的增加是與 LPS、PGN 刺激巨噬細胞所產生的 TNF表現量有關。由實驗結果可知,PD98059 (MEK 的抑制劑) 以及 Src siRNA 的細胞皆可有效的抑制 LPS、PGN、 SNAP、8-br-cGMP 刺激巨噬細胞所造成的 ERK activation 以及 TNF VII.
(9) 的釋放量。基於實驗結果指出 iNOS、ERK activation 與 TNF存在著 重要的關聯性,可能是 LPS、PGN、SNAP、8-br-cGMP 透過 iNOS 活化 Src 和 ERK 去調控 TNF的表現量。. VIII.
(10) 壹、 序論 (Introduction). 1.
(11) 第一節. 免疫系統 (immune system) 與巨噬細胞 (macrophages). 人體的免疫反應,可分為兩個階段:先天性免疫 (Innate immunity) 及後天性免疫 (adaptive immunity)。先天性免疫不具專一性,利用非 特異性的作用,來阻隔多種不同感染性病原。動物體內阻隔感染的有 皮膚、黏膜等屏障,而唾液、胃液、淚腺中的酵素可以有效分解致病 菌。但若這些屏障都無法有效阻隔致病菌的入侵時,生物體內則會啟 動第二道屏障,參與者多為吞噬性的細胞,包括單核球. (monocyte)、. 噬中性球 (neutrophil)。單核球活化後會移行到周邊組織中,就稱為 巨噬細胞 (macrophage),其可藉由吞噬作用 (phagocytosis) 來清除致 病菌,且在進行吞噬作用的同時也會釋放出細胞激素 (cytokines) 和 化學激素 (chemokines) 引發發炎反應。另一方面,巨噬細胞吞噬致 病菌並加以修飾之後表現於細胞膜上,此過程稱為抗原呈現作用 (antigen presentation)。透過抗原呈現作用,巨噬細胞可以活化 T 細胞 釋放細胞激素 (cytokines);另外一方面,活化的 B 細胞會產生專一 性抗體使巨噬細胞更能有效率的辨識病原體。所以,巨噬細胞在發炎 及免疫反應中,扮演很重要的角色 (Janeway, 2001)。. 第二節. 內毒素 (Endotoxin). 2.
(12) 內毒素的定義為:存在於活體微生物細胞壁或細胞質內的毒素,除非 細胞裂解,否則這毒素不會分泌到培養基中(Myhre et al., 2006)。酯多 醣 (lipopolysaccharide, LPS)是屬於格蘭氏陰性菌細胞外膜的主要成 分之ㄧ,屬於內毒素 endotoxin 的一種。LPS 在結構上主要可區分為 三部份:O 抗原、脂質 A 及核心多醣 (Caroff et al., 2002)。位於外側 的 outer-polysaccharide 是一段具有抗原特異性的多聚醣,內含可誘發 專一抗體的 O 抗原(O-specific chain),可應用於臨床診斷,做為辨 識不同格蘭氏陰性菌的依據;位於最內側的部份是由一段脂肪酸所組 成的長鏈結構,稱為脂質 A,則是 LPS 誘發發炎反應產生的主要部 位;位於中央的核心多醣可以利用共價鍵的結合方式連接 O-specific chain 及脂質 A (Caroff et al., 2002)。在細菌入侵宿主細胞時由細胞壁 所釋放的 LPS 會刺激體內包括 T 細胞、單核球細胞、巨噬細胞、 上皮細胞、血管內皮細胞等多種不同細胞的生理反應 (如: 調節物質 的產生、胞飲作用、增生及分化等),嚴重時會造成宿主嚴重的發炎、 水腫、以及敗血性休克等症狀(Woltmann et al., 1998)。peptidoglycan (PGN) ,它是細菌細胞壁的主要成份,在格蘭氏陽性菌細胞壁中, 但是少量存在於陰性菌的細胞壁,含有多層的 PGN。PGN 主要由 GlcNAc (N-acetylgalactosamine) 與 MurNAc (N-acetylmuramic acid) 以 β(1-4)連接的聚合物構成,聚合物的 MurNAc 可連接四個 L 或 D 3.
(13) 構形的胺基酸,通常第三個為 m-diaminopimelic acid (m-DAP,lysine 的前驅物),且大多數格蘭氏陽性菌的第三個胺基酸為 L-lysine。以此 m-DAP 或 lysine 與另一個聚合物所連接的第四個胺基酸連結,反覆 下來形成 PGN (Dziarski and Gupta, 2006)。. 第三節. Toll-like receptor (TLR). Toll-like receptor (TLRs). 是一個 pattern-recognition receptors. family 的一員 (Akira et al., 2006),到目前為止,已有 16 種 TLRs 被 發現 (Keestra et al., 2007)。除了 TLRs 3,7,8,和 9 是細胞內位於 endosome 的受體,其餘的 TLRs 都位於細胞膜上。雖然目前 TLR10, 12,和 13 的 ligands 尚不清楚,但多數 TLRs 的 ligands 都已被發現。 TLRs 信號傳遞主要是透過兩條路徑來活化 NF-κB 促其進入核內增加 NF-κB 下游基因的表現。除了 TLR3 之外,大部分的 TLRs 透過 MyD88 傳遞訊息 (Wong and Wen, 2008)。而內毒素活化巨噬細胞就是透過 TLRs 的路徑,如 TLR4 可以辨識革蘭氏陰性菌的 Lipopolysaccharide (LPS);TLR2 辨識 peptidoglycan (PGN);TLR3 可以辨識病毒的雙股 RNA;TLR5 可以辨識鞭毛蛋白;TLR7 跟 TLR8 是同源的類似物, 可辨識單股 RNA 或是 siRNA;TLR9 可辨識病毒 DNA 上的 CpG motif (Hayden et al., 2006; Kawai and Akira, 2006)。LPS 與 PGN 的活化路徑 4.
(14) 很相似,透過 TLR-4 與 TLR-2 的 cytosolic domain 和細胞內的訊息傳 遞蛋白結合,活化下游一連串的訊息傳遞路徑,如:MAPK pathway、 NF-κB。TLR4 將 LPS-induced signal 傳遞至細胞內時,經由 MyD88 (Toll-interacting protein) 及 IL-1R-associated kinase (IRAK) 及 TNF-receptor-associated factor 6 (TRAF6) 等訊息蛋白,而活化 TGF-activated kinase-1 (TAK1),引發 NF-inducing kinases (NIK) 活化並與 IKK形成複合物,磷酸化 ,造成其分解進而活化轉 錄因子 NF-。NF-活化後由細胞質進入細胞核,調控包括 tnf、. inos 等基因在內的轉錄作用 (Thanos and Maniatis, 1995)。. 第四節. iNOS (inducible nitric oxide synthase). 一氧化氮(NO) ,是一種獨特可擴散的訊號分子,透過稱為氮氧化物 合成酶的酵素(NOSs)對 L-arginine 的氧化作用所產生 (Nathan, 1994)。NOS具有三種不同的 isoforms,依照被鈣離子影響與否可區 分為 Ca2+ -dependent 與 Ca2+ -independent兩大類,而 Ca/calmodulin 能調節neuronal (nNOS) 以及endothelial (eNOS) 的活性並且持續的 表現,而 inducible (iNOS) 的活性是不受 Ca/calmodulin 所調控並且 只透過致病菌的產物以及 cytokines 引起。而 NO 在人體內維持著. 5.
(15) 一定的濃度,調控著人體的生理狀況,例如:傳導神經訊息,舒張血 管,鬆弛平滑肌, 抑制血小板凝集。相反地,由巨噬細胞以及其他 宿主細胞所產生的大量的 NO 則調控著宿主的免疫能力,包括抗細 菌和抗癌功能(Bogdan, 2001; Duffield, 2003)。但是,也有研究指出緩 慢並持續性表現的 NO 會誘導發炎以及癌症的產生(Knowles and Moncada, 1994; Moncada and Martin, 1993)。而 soluble guanylate cyclase (sGC) 是 NO 主要鍵結的目標之一,當 NO 鍵結到 sGC, 會活化 sGC 促使guanosine 5-triphosphate 轉變成 Cyclic guanosine monophosphate (cGMP),而 cGMP 也是人體內的二級訊號分子 (Krumenacker et al., 2004)。. 第五節. Src kinase. cellular Src (c-Src) 分類上屬於 nonreceptor tyrosine kinase (NRTK) (Levy et al., 1984),是 Src family kinases (SFKs) 的其中一員,這群 protein-tyrosine kinases 在結構上有幾個高度保留的區域。由 N 端到 C 端分別為可 myristylation 的 SH4 domain,接下來是 unique domain、 SH3 domain、SH2 domain、linker、kinase domain、以及 regulatory domain。 SH4 的 myristylation 可使 Src 透過長鏈脂肪酸與細胞內 膜結合、unique domain 可特異性的辨別整個家族中的成員、SH3 可 6.
(16) 與 proline-rich 的胺基酸序列結合、 SH2 則可與 phosphorylated tyrosine 有很強的鍵結、kinase domain 與 regulatory domain 則可調 控 Src 的酵素活性。 Src 分子量為 60 kDa,它的特性在於胺基酸 C 端末端有個短尾巴 (regulatory domain) ,位於其間的 Tyr-527 若被 磷酸化則可使 Src 活性降低。當 Tyr-527 磷酸化後會造成 SH2、 SH3 與 kinase domain 聚集,形成結構上的抑制。至於 kinase domain 上的 Tyr-416 的自體磷酸化則可增加 Src 活性。因此藉由 Tyr-416 與 Tyr-527 兩個位置的磷酸化可調控 Src 的活性 (Boggon and Eck, 2004)。目前在人體內的 SFKs 總共有 11 種,分別為 Blk、Brk、Fgr、 Frc、Fyn、Hck、Lck、Lyn、Src、Srm、Yes,其中 Src、Fyn、Yes 是廣泛存在真核細胞中的 (Roskoski, 2004),而 Lyn、Hck、Fgr、Blk、 Lck 則表現在造血細胞。PGN 處理巨噬細胞之下對 Src family 的研 究最早在 1995 年,證明了巨噬細胞在 PGN 處理下可使 Lyn 的磷酸化 表現量增加 (Gupta et al., 1995)。過去在 LPS 處理下,巨噬細胞的 Lyn、 Hck、Fgr 的酵素活性也都上升。但 Fanying Meng 在 1997 年證明了, 即便把這三種蛋白 knockout 之後,LPS 刺激下其巨噬細胞仍舊可以 分泌 IL-6、TNF-α這些發炎的前驅物質 (Meng and Lowell, 1997)。 所以此意味著在 LPS 刺激下的巨噬細胞有尚未被發現的 tyrosine kinase(s) 主導 pro-inflammatory cytokines 的產生。 7.
(17) 第六節. 活化蛋白質激酶 (Mitogen-activated protein kinase,. MAPK kinase). The mitogen-activated protein kinases (MAPKs)是一群蛋白質結構類 似的 serine/threonine kinases,存在於真核細胞中;當細胞外接受到刺 激時,MAPKs 可啟動細胞內訊息傳遞路徑。MAPKs 可區分成三大類: (一) extracellular responsive kinases (簡稱 ERK1/2 或 p44/42 isoforms MAPK) (Davis, 1994; Nishida and Gotoh, 1993);(二) c-jun N-terminal kinase 及 stress-activated protein kinases (簡稱 JNKs/SAPKs) (Derijard et al., 1994); (三)p38 MAPKs (Han et al., 1994)。雖然這三大類的 MAPKs 在結構上及功能上都很類似,但卻由不同的刺激、由不同上游的 kinases 來調控活性,架構出不同的 MAPK cascade 傳送訊息 (Cano et al., 1995)。ERK cascade 由三個層次的 kinases 組成。第一個層次是 MAP kinase kinase kinase(簡稱 MAPKKK) ,包括有 Raf-1 和 B-Raf; 第二個層次是 MAP kinase kinase(簡稱 MAPKK),就是 MEK;而 第三個層次是 MAPK,即 ERK1/2。當 MAPKKK 受到上游訊息活化 之後,會將 MEK 上的 serine/threonine 磷酸化,造成 MEK 活性增加 而磷酸化 ERK1/2,使之活化。此一 ERK cascade 包括 Ras、GEF、. Rap 在內等多種蛋白調控(Grewal et al., 1999)。之前的研究指出, 8.
(18) ERK cascade 會受 growth factors 和 tumor promoters 所誘導而活化, 進而調節轉錄因子及基因的表現參與細胞的增生、分化、移行、凋 亡(Kumar et al., 1998)。. 9.
(19) 貳、 實驗材料及方法. 10.
(20) 第一節 實驗設計流程. 11.
(21) 第二節 實驗材料 細胞: 本實驗所使用老鼠巨噬細胞株 (murine macrophage-like cell line) Raw264.7,購於食品工業研究所,取自公的 BALB/c 老鼠的巨噬細胞, 經過 c-Abl (abelson) transform。第二種是從活體老鼠取得之巨噬細胞 (rat peritoneal macrophages),簡稱為 PEM 細胞。. 動物: 使用的動物有 C57BL/6 or SD 兩種品系。 I. C57BL/6 mice:約五到七週大的 mice,公鼠,購於財團法人國家 實驗動物中心。 II. SD rat:約 250 g,公鼠,購於樂斯科生物科技股份有限公司。. 試劑: A. 實驗藥品 1. Lipopolysaccharides from Escherichia coli serotype 0111:B4 和 thioglycollate (Sigma) (St. Louis, Missouri, USA) 2. Peptidoglycan from Staphylococcus aureus (Sigma) 3. S-Nitroso-N-acetyl-DL-penicillamine (SNAP) (Sigma) 12.
(22) 4. 8-Bromoguanosine 3′,5′-cyclic monophosphate sodium salt (8-Br-cGMP) (Sigma) 5. 1H-[1,2,4]Oxadiazole[4,3-a]quinoxalin-1-one (ODQ) (Sigma) 6. N-([3-(Aminomethyl)phenyl]methyl)ethanimidamide dihydrochloride ;1400W dihydrochloride (1400W) 7. 2’–amino-3’-methoxyflavone (PD98059) (Merck). B. 細胞培養試劑 1. RPMI-1640 (invitrogen) 2. fetal bovine serum (FBS) (JRH Biosource) 3. Penicillin-Streptomycin (Sigma) 4. L-Glutamine (Sigma). C. 電泳及西方墨點轉漬試劑 1. Protein assay reagent (Bio-Rad) 2. Ammonium persulfate (APS) (Gibco BRL) 3. N,N,N,N-Tetramethyl ethylene diamine (TEMED) (Gibco BRL) 4. Bis-acrylamide (AMRESCO) 5. Tween 20 (Merck) 6. Prestained marker (Bio-Rad) 7. SDS-PAGE molecular weight standard[Bio-Rad (Bio-Rad Pacific Ltd., Hong Kong)] 13.
(23) 8. Sodium dodecylsulfate (SDS) (USB) 9. Tris-base (USB) 10. Glycine (USB) 11. Western Blot detection reagent (ECL kit) [Amersham Pharmacia 12. Thoglycolate (MERCK) 13. Methanol (Fisins Scientific Equipment). D. 抗體 1. Phospho-p44/42 MAP kinase (Thr202/Tyr204) (E10) Mouse mAb (Cell Signaling) 2. Anti-iNOS/NOS II (rabbit polyclonal IgG) (upstate cell signaling solutions) 3. ERK-1 (K-23): sc-94 (Santa cruz biotechnology, inc) Anti-actin (anti-actin mouse monoclonal antibody) (Sigma) 4. HRP-rabbit anti-goat antibody (Jackson) 5. HRP-goat anti-mouse antibody (Jackson) 6. HRP-goat anti-rabit antibody (Jackson). 14.
(24) 實驗儀器與耗材. 儀器. 廠牌. Centrifuge. Eppendrof 5424. Centrifuge. Beckman Allegra. Hemacytometer. BOECO. Hoefer semiPhor. Pharmacia Biotech. TE70PH meter. JENCO 6071. Power supply. EPS 1001. Shaker. TKS RS01. Universal Microplate. BIO-TEK. Spectrophotometer Automated Strip Washer. BIO-TEK. Stirrer/Hot plate. CORNING PC640. Vertical slab gel unit. Hoefer SE400. Vortex. GENIE SI-2 G560. Water bath. TKS ZX-400. Boyden chamber. Neuro probe. 15.
(25) 耗材. 廠牌. Protein Assay Kit. Bio-Rad. Polycarbonate filter (8 μm). Neuro probe. PVDF transfer membrane. NEN. X-15RDigital analysis system. Kodak EDAS 120. 第三節 實驗方法. (一). 細胞培養 (cell culture). A. 培養液 培養細胞的培養液為 RPMI,配置前將滅好菌且烘乾過的玻璃 瓶拿到無菌操作台裡面照 UV 一個晚上。配置時將一單位的 RPMI powder、2.0 g 的 NaHCO3 和 5.957 g 的 HEPES 溶於滅菌的二次水 中,將 pH 值調至 pH 7.3~pH 7.4,補二次水至總體積 1 liter。 再以 0.22 μm 的濾膜過濾分裝,保存於 4℃。. B. 培養液添加物 1. Fetal bovine serum 16.
(26) 將冷凍的 FBS 靜置於水中,使其慢慢溶解後,於無菌操作檯 中分裝至 50 ml 無菌離心管中,再用 parafilm 包覆瓶口,保存 於-20℃。 2. Penicillin-Streptomycin. (10X;10000 unit/ml-10000 μg/ml). 每 500 ml 的 RPMI 加 5 ml 的 Penicillin-Streptomycin。 3. L-Glutamine (10X;200 mM) 每 500 ml 的 RPMI 加 5 ml 的 L-Glutamine。. C. PBS (phosphate balanced solution) 1X PBS 以 8 g NaCl、1.2 g NaH2PO4.H2O 加二次水配製,將 其 pH 調成 7.4 後補二次水到 1L,滅菌後保存於 4℃。. D. 冷凍培養液 以 1 ml 的 DMSO 加上 3 ml 的 RPMI 配製成 25% DMSO,這 裡的 RPMI 必須是沒添加過血清但有添加 P/S 與 L-Glu,以上混 何均勻後存放於-20℃。冷凍培養液,以 4 ml 的 25% DMSO 與 6 ml FBS 混合,最後的 DMSO 濃度為 10%. 17.
(27) E. 細胞培養步驟 1. 細胞分盤 (subculture) 將細胞培養於 culture RPMI medium (10% FBS、1% P/S、1% L-Glu),放置於細胞培養箱 (37℃,5%. CO2),等細胞成長約八. 至九分滿時,進行分盤培養。分盤前務必在新的培養皿上標明細 胞株的名稱、分盤時間及代數,分盤時將舊的培養液吸走,每次 以 37℃回溫後的滅菌 1X PBS 7~10 ml 沖洗細胞,清洗一次。再 加入 8~10 ml 37℃回溫後的培養液,將細胞全部沖下來,並吸放 數次直到均勻沖散細胞以 1:3 或 1:4 的比例分盤培養。. 2. 冷凍細胞: 培養好約八分滿的細胞,且冷凍細胞前一天需更換成新鮮的 培養液。首先將舊的培養液吸走,每次以 7~10 ml 滅菌 1X PBS 沖洗細胞,清洗一次。加入 8~10 ml 的培養液,將細胞全部沖下 來,再將細胞液吸取置於 50ml 離心管,離心 (1000 rpm,10 分 鐘或 2000 rpm,5 分鐘) 之後將細胞沉澱下來。去除上清液之後, 以 1 ml 冷凍培養液將細胞上下吸、放數次,直到細胞被均勻打 散,置於冷凍管,先存放於 4℃冰箱 30 分鐘,再移置於-80℃冰 箱,12 小時之後再存放於液態氮桶。. 18.
(28) 3. 解凍細胞: 由液態氮桶中取出細胞,將細胞溶於適量的 RPMI 培養液, 離心 2000 rpm 五分鐘。倒掉 RPMI 培養液,以 10 ml 的 RPMI 將 細胞打散,隔天細胞貼附後再換培養液,約一個禮拜細胞穩定後 才可以做實驗。. (二). 收集 cell. lysates. A. 試劑: 主要溶液 Lysis buffer (RIPA buffer),可將細胞裂解。以 4.38 g NaCl、3.0285g Tris-base、1.25g Deoxycholate、5 ml IGEPAL CA-630 (相 當的粘稠,要緩慢的取) 來配製。以上藥品溶於適量的二次水後,將 溶液配成 500 ml pH 7.4,高溫高壓滅菌後保存於 4℃。每當要使用前, 必須將它配置成 Modified RIPA buffer,若要配製成 1 ml,則需要 1 ml RIPA、10 μl 200 mM Sodium orthovannadate (final conc.= 1 mM)、5 μl 200 mM EGTA (final conc. = 1 mM)、10 μl 0.5% Aprotinin (final conc. = 0.0025%)、4 μl 200 mM PMSF (final conc. = 1 mM)。後面追加的是蛋 白質水解酶抑制劑,包括 Sodium orthovannadate 是 phosphatase 19.
(29) inhibitor,EGTA 是鈣離子的螯合劑, Aprotinin 是 serine protease inhibitor,PMSF 也是 serine protease inhibitor。. B. 步驟 : 1. 將 medium 吸走,每次以 1ml 1X PBS 清洗細胞,共兩次,之後再 將 PBS 吸乾。 2. 加入 500 μl modified RIPA buffer(6 cm dish),利用刮勺來回將 細胞刮下 (置於冰上操作),再將細胞液置於 eppendrof,將 eppendrof 在 rack 上左右來回刮數次。 3. 以 4℃、10000 rpm,離心 10 分鐘,取上清液到新的 eppendorf, 保存於 -80℃。. (三). 蛋白質濃度測定. A. 試劑 : 1. BSA (1 μg/μl) 2. Protein Assay Kit. B. 步驟 : 1. 以不同濃度的 BSA 畫出蛋白質濃度測定的 standard curve。其方 20.
(30) 法如下 : 取 0、5、10、15、20、25、30 μl BSA (1 μg/μl) eppendorfs。 2. 加入不同量的 d.d H2O 到 eppendorfs 中,使其總體積達 800 μl。 3. 加 200 μl Protein Assay Kit (dye) 到 eppendorfs,以 vortex 混合均 勻。 4. 在 sample 的測定上,取 10 μl cell lysates,790 μl d.d H2O,200 μl dye,混合均勻。 5. 採用 Bradford Protein Assay,測各種不同量之 BSA 在波長 595 nm 的吸光值,畫出 standard curve。再測 sample 的 O.D.值,求出 sample 的蛋白質濃度。. (四). 蛋白質電泳 (SDS-PAGE). A. 試劑 : 1. Acryamide. 150 g. N’N’-Methylene bisacryamide. 4g. 將所需之化學藥品溶於 350 ml d.d H2O 再加 d.d H2O 至 500 ml, 保存於 4℃。. 2. Tris-HCl (2 M,pH 8.8). 500 ml. Tris-HCl. 121.14g 21.
(31) 將 Tris-HCl 溶於 350 ml d.d H2O,pH 調成 8.8 再加 d.d H2O 至 500 ml。. 3. Tris-HCl (2 M,pH 6.8). 500 ml. Tris-HCl. 121.14g. 將 Tris-HCl 溶於 350 ml d.d H2O,pH 調成 6.8 再加 d.d H2O 至 500 ml。. 4. 10% APS. 10 ml. APS. 1g. 將 APS 溶於 8 ml d.d H2O 再加 d.d H2O 至 10 ml, 保存於 4℃。. 5. Resolving gel (8%). 30 ml. H2O. 15.8 ml. 2M Tris-HCl, pH 8.8. 6 ml. Acrylamide/bis (30%/0.8%). 8 ml. 20% SDS. 150 μl. 10% APS. 150 μl. TEMED. 20 μl. 6. Stacking gel (8%). 10 ml 22.
(32) H2O. 8 ml. 2M Tris-HCl, pH 6.8. 0.625ml. Acrylamide/bis (30% / 0.8%). 1.33 ml. 20% SDS. 50 μl. 10% APS. 50 μl. TEMED. 10 μl. 7. 10X Sample buffer 50 ml 1M Tris-HCl, pH 6.8. 1 ml. SDS. 5g. Sucrose. 25 g. Bromophenol blue. 10 mg. 2-mercaptoethanol. 5 ml. 將所需之化學藥品溶於 350 ml d.d H2O,再加 d.d H2O 至 500 ml, 保存於室溫。. 8. Running buffer. 500 ml. Tris-HCl. 7.5 g. Glycine. 36 g. 10 % SDS. 2.5 g. 再加 d.d H2O 至 500 ml, 保存於室溫。. B. 步驟 : 1. 將兩塊玻璃板、兩條 spacer 及灰色夾組合成一個製膠架,然後利 23.
(33) 用黑色旋鈕固定在底座。 2. 將下層膠配方所需的藥劑依序加入燒杯,攪拌均勻,利用 pipette 加到電泳槽至適當位置。 3. 加入 1 ml d.d H2O,隔絕空氣,待凝膠 (約 25 分鐘)。 4. 將 d.d H2O 倒掉,插入 comb,將上層膠配方所需的藥劑依序加入 燒杯,攪拌均勻,利用 pipette 加到電泳槽。 5. 等上層膠凝固之後,將 comb 拿走,倒入上、下層各 200 ml 1X running buffer。 6. 將 sample 和 10X sample buffer (10:1) 混和均勻,稍微 spin 一下, 然後煮沸 5 分鐘。 7. 利用 micropipette,將 sample 加到 well 內,進行膠體電泳法 (50 V、 20 mA,時間 18 小時;200 V、30 mA,時間 5 小時;如果要跑 開 Src-416,則需 4℃、70V、時間 20 小時或室溫、60V、時間 23 小時)。. (五). Western blot analysis. A. 試劑 : 1. Transfer buffer. 500 ml. Tris-HCl. 1.5 g 24.
(34) Glycine. 7.2 g. SDS. 0.5 g. Methanol. 100 ml. 將所需之化學藥品溶於 350 ml d.d H2O 再加 d.d H2O 至 500 ml, 隔夜去除氣泡,保存於室溫。. 2. 10X TBS. 500 ml. Tris-HCl. 30.3 g. NaCl. 43.83 g. 將所需之化學藥品溶於 350 ml d.d H2O,pH 調成 8.8 再加 d.d H2O 至 500 ml,保存於室溫。. 3. Blotting buffer. 50 ml. 10X TBS. 5 ml. BSA. 1.5 g. Tween 20 (USB). 25 μl. 將所需之化學藥品溶於 35 ml d.d H2O 再加 d.d H2O 至 50 ml,保存於室溫。. 4. Washing buffer. 500 ml. 10X TBS. 50 ml. Tween 20. 500 μl. 將所需之化學藥品溶於 350 ml d.d H2O 25.
(35) 再加 d.d H2O 至 500 ml,保存於室溫。. B. 步驟 : 1. 剪取適當大小的 PVDF transfer membrane,以 methanol 浸泡約 30 秒後,再以 d.d H2O 浸泡。 2. 利用 semi-phor transblotter 將電泳膠中的蛋白質 transfer 到 PVDF transfer membrane (25V、300 mA,1.5 小時)。 3. 將 PVDF transfer membrane 取出,浸泡於 30 ml blotting buffer 中,在室溫下置於 shaker 上 blotting 3 小時(shaker 擺動速度: 5rpm)。 4. 將 blotting buffer 倒掉,加入 10 ml 含有 primary antibody (1: 1000)的 blotting buffer,置於 4℃作用 12 小時或室溫 1~2 小時。 5. 隔天取出之後,在室溫下先置於 shaker 上作用 30 分鐘(shaker 擺動速度:5rpm),再以 washing buffer 清洗 membrane 三次,5 分鐘;5 分鐘;10 分鐘 (shaker 擺動速度: 50~60 rpm)。 6. 加入 10 ml 含 secondary antibody 的 blotting buffer (1:2000),室溫 blotting 1 小時 (shaker 擺動速度: 5 rpm)。 7. 再以 washing buffer 清洗 membrane 五次, 5 分鐘;5 分鐘;10 分鐘;15 分鐘;15 分鐘 (shaker 擺動速度: 50~60 rpm)。. 26.
(36) 8. 依 membrane 大小 (0.125 ml/cm2),加入適量的 ECL kit solution I 與 II 等比例的均勻混和液。在 membrane 與之作用 5 分鐘後, 以 X-ray film 感光。. (六). 一氧化氮濃度測定. (Nitrite concentration. measurement). A. 試劑 : Reagent A4 與 A5 Reagent A4 以 500 mg Sulfanilamine、2.5 mg H3PO4 溶於 500 ml, 避光保存於 4℃。Reagent A5 50 mg N-(1-naphthll-)-ethylenediamine (NED) 溶於 500 ml 二次水,避光保存於 4℃。主要的作用在於 A4 的 Sulfanilamine 會與溶於水的 NO (NO2-) 作用產生化合物,這化合 物會與 A5 的 NED 形成紅色的產物,藉此有色產物的量來推測溶液 中有多少 NO 溶於水。. B. 步驟 首先為了將樣品標準化,我們測量 culture dish 中培養液的總量, 與其中蛋白質總量。然後自 culture dish 先取 50 μl 的培養液加至 96. 27.
(37) well dish 中,加入 45 μl 的 A4 混合均勻,利用 ELISA reader 讀取波 長 540 nm 的吸光值 (A1)。再加入 45 μl A5 呈色、避光十分鐘。再 用相同波長讀取第二次的吸光值 (A2)。測定吸光值的時候要避免產 生氣泡。濃度的計算,是以 Nitrite 濃度計算公式為 Total Nitrite = (A2-A1 ) nitrite-( 0.006 ) blank nitrite,利用 NaNO2 做標準曲線:3.5361× A3Total Nitrite - 0.001 =. ( NO2 濃度 μg / ml)。將所測的的數值以內. 差法算出濃度,再以培養液總量與蛋白質總量來標準化。. (七). 腹腔巨噬細胞製備及培養. A. 動物 1. 4-6 weeks 的 SD rat (體重約 250~300g) 2. C57BL/6 mice (5-6 weeks) 體重約 20g 3. iNOS-deficient mice (5-6 weeks) 體重約為 20g. B. 試劑 1. 3% Brewer thioglycolate medium 10 ml/rat 2. 3% Brewer thioglycolate medium 10 ml/mice. C. 步驟 28.
(38) 將老鼠腹腔注射 3% Brewer thioglycolate medium,劑量 10 ml,誘 發老鼠腹腔的巨噬細胞引起發炎反應。4 天後,使用 CO2 犧牲老鼠。 並將 37℃已回溫 PBS,以無菌注射空針,注射 3% TG 至老鼠腹腔中 且不斷按摩老鼠腹部區域約 2-3 分鐘後,使用手術剪刀 Operating Scissors str,將老鼠腹腔剪開,以無菌塑膠吸管,將腹部內的 PBS 緩 緩回抽至 PBS 呈現白色透明混濁;mice 部分則是直接剪開腹腔,利 用玻璃滴管尖端細頭處緩慢將腹腔內 PBS 抽吸出,放入 50 ml 離心 管 (過程中應避免氣泡產生及避免抽取到腹腔內組織器官引起出血)。 將抽吸好 PBS 使用 centrifuge 離心 (2000 rpm,20 min),離心後,將 上清液丟棄。以適量 RPMI (已添加 10% FBS) 將細胞輕輕打散,待 細胞計數後,培養在 culture dish 內,放入 37℃ 5% CO2 下細胞培養 箱,待細胞貼附約 5hrs 後,去除舊的 RPMI 培養液,重新更換新的 RPMI medium,以利細胞繼續生長,培養細胞至 24 hrs 準備待做實 驗。. (八). TNF-測定. A. 試劑 1. Assay buffer 配製方式為 8.0 g NaCl、1.13 g Na2HPO4、0.2 g KH2PO4、0.2 g 29.
(39) KH2PO4、0.2 g KCl、5.0 g BSA、1 ml Tween20,加二次水到 1.0 L, PH 調至 7.4。主要的功能在於稀釋檢體 (包括 standards)與抗體 (coating Ab 除外)。 2. Other buffer Coating buffer B,配製法 4.3 g NaHCO3、5.3 g Na2CO3,加二次 水到 1.0 L,pH 調至 9.4。主要功能用於稀釋 Coating Ab 。Washing buffer,以 9.0 g NaCl、1 ml Tween20,加二次水到 1.0 L,PH 調至 7.4。 主要功能用來洗去未連接的抗體或蛋白。Stop solution,配製 1.8 N H2SO4,主要功能用來終止呈色反應。. B. Plate coating 以coating buffer B稀釋coating Ab (1.25 μg/ml),例如:10 ml則以 12.5 μl coating Ab + 9.988 ml coating buffer。於96孔盤中每個well放入 100 μl coating solution,輕拍盤子讓溶液均勻,再用保鮮膜或蓋子封 好,於4℃下放置12~18 hrs。將多餘的coating solution到掉後,倒放在 擦手紙上盡量把液體吸乾。每個well加入300 μl assay buffer來做block 的動作 (依kit不同有些為特殊的blocking buffer),於室溫下放置1 hr, 也可放在2~8℃下可保存五天。倒掉assay buffer,倒放在擦手紙上。 在配置好樣品後,以300~400 μl washing buffer 洗3~6次,再倒放在擦 手紙上吸乾水分。 30.
(40) C. ELISA method 用 assay buffer 序列稀釋來稀釋 standards。standards 的濃度依序為 1000 pg、500 pg、250 pg、125 pg、62.5 pg、31.25 pg、15.625 pg、0 pg, 共八組濃度。同樣的以 assay buffer 稀釋欲待測 samples。第一次實驗 欲稀釋倍數,可由一倍到五千倍來做大範圍的稀釋,往後則挑選落在 標準曲線內的稀釋倍數來做多重複的實驗以求誤差值。首先將先前 plate 300 μl assay buffer 倒掉,再以 300~400 μl washing buffer 洗滌 3 次後,倒放在擦手紙上以利吸乾多餘液體,再至已事先設計好 wells 內依序加入 100 μl standards 或 sample,立即馬上加入 50 μl Detection Antibody 反應,於室溫下作用 2 小時,之後再加 300~400 μl washing buffer 洗滌 3 次,再加入 assay buffer 稀釋後的 streptavidin-HRP 100μl 於室溫下作用 30 mins。再使用 300~400 μl washing buffer 洗滌 3 次, 依序每 well 加入 100μl chromogen TMB 反應,避光 30 mins 之後每 well 依序加入 100 μl stop solution (需於 30 分鐘內測定吸光值)。使用 ELISA reader 測得 450/650 nm 的吸光值後,使用電腦將所得的 standarts 做成標準曲線,將 sample 測得的數值帶入標準曲線後,依照 稀釋倍數與蛋白質濃度定量換算出每毫升所含的數值。. 31.
(41) 參、 結果 (Results). 32.
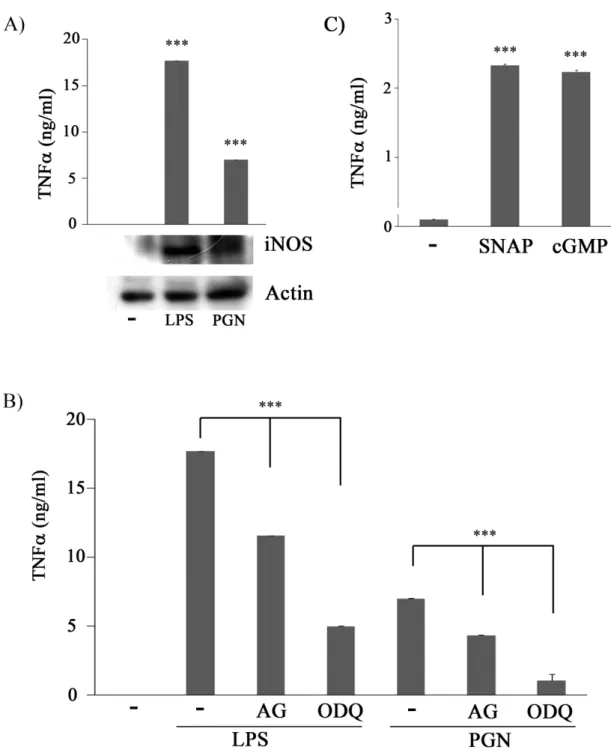
(42) 一、. AG 及 ODQ 皆可同步抑制 PGN 所產生的 TNF,且 SNAP. 及 8-br-cGMP 處理 Raw 264.7 巨噬細胞,皆可增加 TNF的釋 放。. 過去實驗室証明 LPS 會經由 TLR4 活化巨噬細胞,活化轉錄因子 NF-;活化後 NF-可由細胞質進入到細胞核中,與其標的基因的 5-flanking region 結合後進而開啟下游基因表現。而 iNOS 及 TNF- 都是 NF-的標的基因,因此 RAW264.7 巨噬細胞株在 LPS 的刺激 之下會促使 iNOS 及 TNF-的表現。有趣的是先前實驗室研究發現 LPS 刺激巨噬細胞促進 TNF-的表現是受到 iNOS 的調控。而 PGN 是透過 TLR2 活化巨噬細胞,相似的路徑下,我們進一步探討 LPS&PGN-induced macrophage activation 中,iNOS 扮演了何種角色, 由 Fig 1A 所知 iNOS、TNF-會受到 LPS 與 PGN 的刺激表現,因此 我們想問在 PGN 刺激巨噬細胞釋放 TNF的過程中,iNOS 是否參與 其中。要回答這個問題,我們利用 AG (為 iNOS 抑制劑;2mM) 和 ODQ (sGC 抑制劑;100 M),前處理 RAW 264.7 巨噬細胞 30 分鐘後 再處理 PGN (5 μg/ml)。48 hrs 後,收集 cell conditioned medium 測量 TNF的釋放量。結果如 Fig 1B 所示,與 control 組比較,LPS 與 PGN 刺激後的細胞其 TNF釋放量有顯著增加;但合併處理 AG 或 ODQ 33.
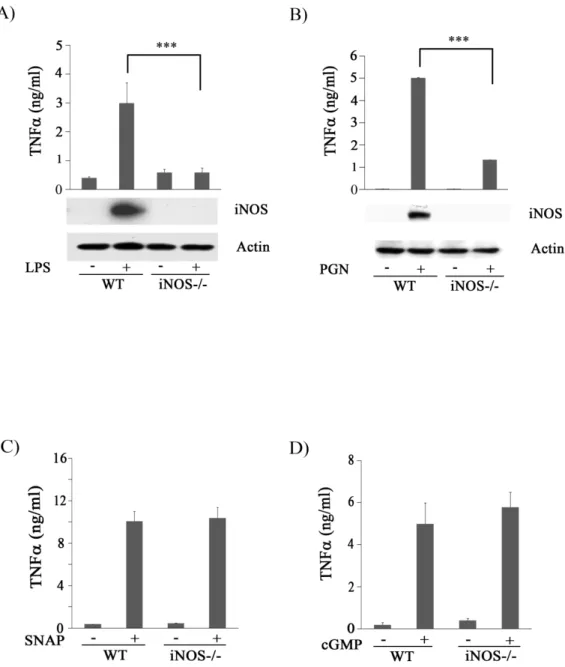
(43) 的細胞,其 TNF的釋出就大幅降低。因此我們合理的懷疑 PGN 刺 激巨噬細胞所造成 TNF釋放的過程中,NO 及 sGC 可能參與其中。 接下來我們直接給予 SNAP (NO 的 donor;100 M)及 8-br-cGMP (cGMP 的 analog;100 M) 來刺激巨噬細胞。結果顯示 SNAP 及 8-br-cGMP 都可以誘導 Raw264.7 巨噬細胞產生 TNF分泌(Fig 1C), 這意味著 LPS 與 PGN 誘導 TNF分泌的過程中,iNOS 及 sGC 參與 其中。. 二、. SNAP 與 8-br-cGMP 可誘導 iNOS knockout mice peritoneal. macrophage 產生 TNF. 我們證實了 AG 及 ODQ 可抑制 PGN-induced TNF secretion。 為了證明 iNOS 為巨噬細胞分泌 TNF所需要的重要蛋白,於是我們 使用 iNOS knockout (KO) mice,來檢視 iNOS 在 PGN 誘導 TNF表現 的重要性。我們收集 wild type (WT) 及 iNOS-/- 老鼠的腹腔巨噬細胞 (PEM) 處理 PGN (5 μg/ml),48 hrs 後收集 cell conditioned medium 測 量 TNF的分泌的量。結果發現與 WT 巨噬細胞相比,iNOS-/- 的巨 噬細胞在 PGN 刺激後的 TNF分泌量明顯下降 (Fig 2A)。接下來, 我們以 SNAP 與 8-br-cGMP 來刺激 WT 與 iNOS-/- 的 PEMs。48 hrs 34.
(44) 後,收集 cell conditioned medium 測量 TNF的分泌的量。由實驗結 果得知 SNAP (NO 的 donor;100 M)及 8-br-cGMP (cGMP 的 analog; 100 M)皆能誘導 WT 與 iNOS-/- PEMs TNF的產生 (Fig 2B、C)。結 果顯示巨噬細胞的 iNOS 及其所啟動的 NO/cGMP pathway,對於 PGN 所誘增的 TNF,有著決定性的影響。. 三、. AG 及 ODQ 皆能有效抑制 PGN 誘導 ERK 的活化. 先前實驗室已經證明 NO/sGC 會影響 tnf mRNA 的表現量(附圖 二),也有文獻指出 TLR4 能透過活化 ERK 傳遞訊號,增加 tnf 基 因的轉錄及 TNF的釋放。而實驗室之前也證明了 1400W 及 ODQ 能 抑制 LPS 誘導 ERK 的活化,且更進一步證實 SNAP 及 8-br-cGMP 能 活化 ERK,增加 TNF的釋出 (附圖三)。由於 TLR2 活化後,TNF 的量會提高,故我們好奇在 TLR2 對 tnf 基因的轉錄及 TNF 釋放 的調控上也跟 TLR4 相類似。為回答這個問題我們使用 AG (2 mM) 和 ODQ (100 M),前處理 RAW264.7 巨噬細胞 30 分鐘後再處理 PGN (5 μg/ml)。48 小時後,收集 cell lysates,分析 ERK 的活性。實驗結果與 TLR4 的結果相同 (Fig 3);意味著,NO 及 sGC 參與在 LPS 和 PGN 誘導 ERK 的活化過程中。 35.
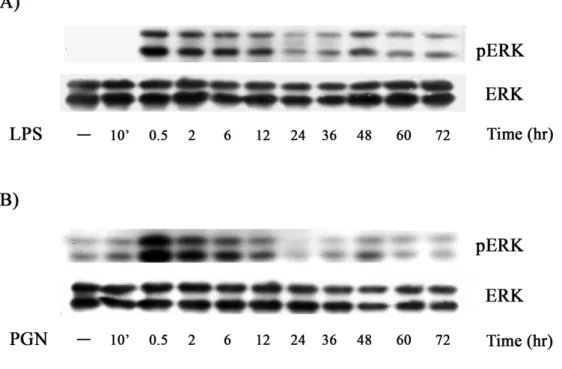
(45) 四、. LPS 與 PGN 誘導 ERK 的活化有兩個不同的階段. 許多文獻指出在 M-CSF 透過活化 ERK 來傳遞刺激巨噬細胞的訊息 路徑中,觀察到 ERK 有兩個不同時間點的活化階段;其分別為 transient activation 和 sustained activation。而文獻的研究皆指出 persistent activation 對於巨噬細胞的分化有較大的影響(Ebisuya et. al., 2005; Guha and Mackman, 2002; Marshall, 1995),因此我們 想觀察 LPS 與 PGN 活化巨噬細胞其 ERK 的活性是否也具有相同的 情形。我們以 LPS (100 ng/ml)和 PGN (5 μg/ml)處理細胞後分別在 10 mins、0.5、2、6、12、24、36、48、60、72 hr 的時間點收集 cell lysates, 分析 ERK 的活性。結果發現在 0.5 hr 左右,ERK 活性明顯增高,但 在 0.5 小時到達高峰後,其活性逐漸下降。24 小時的 ERK 活性較於 0.5 小時的 ERK 活性明顯降低許多。故第一個 ERK activation,我們 稱之為 transient ERK activation。有趣的是,第二波的 ERK 活化於 48 小時時出現了,我們稱之為 sustained ERK activation (Fig 4)。. 五、. LPS 與 PGN 所調控的 sustained ERK 的活性是會受 AG 的. 影響 36.
(46) 在確定 LPS、PGN 處理的巨噬細胞,其 ERK 的活化有 transient 與 sustained ERK activation 之分,因此我們想問的問題是 LPS 與 PGN 活 化 ERK 的路徑中,iNOS 是參與在哪個階段。因此我們利用 AG (2 mM) 前處理 RAW264.7 巨噬細胞 30 分鐘後再處理 LPS (100 ng/ml)和 PGN (5 μg/ml),分別在 10 mins、0.5、2、12、24、48 的時間點收集 cell lysates, 分析 ERK 的活性。結果發現在 iNOS 活性被抑制的情形下,transient (30 mins)的 ERK 活性幾乎不受影響,但 sustained ERK activation 卻明 顯下降,故 sustained ERK activation 有可能受到 iNOS 的調控。. 六、. iNOS 為 PGN-mediated sustained ERK activation 所需. 上述實驗 Fig 3 暗示 NO/cGMP pathway 可能參與 PGN-induced sustained ERK activation。為了證實 iNOS 對 sustained ERK 的重要性, 我們使用 iNOS-/- mice,來觀察其 PEM 在缺乏 iNOS 之下,其 PGN-induced sustained ERK activation 是否受到影響?我們以 WT 及 iNOS-/- mice 的 PEMs 處理 PGN (5 μg/ml) 48 小時後,收集 cell lysates, 分析 ERK 的活性。由數據可知,與 WT PEMs 相比,以 PGN 處理的 iNOS-/- PEMs 其 sustained ERK 的活性明顯被抑制 (Fig 6) 。由此我 37.
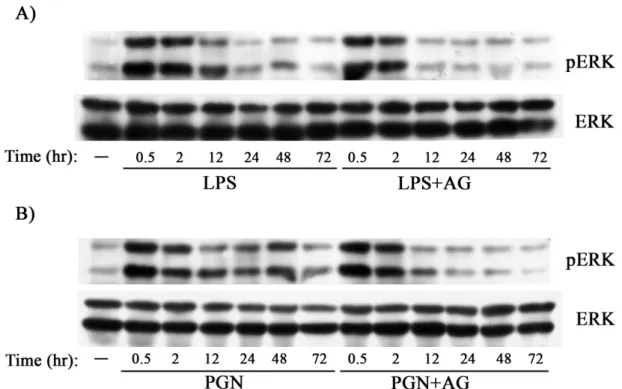
(47) 們推論 LPS 與 PGN 刺激巨噬細胞釋放 TNF的路徑中,iNOS 造成 sustained ERK activation 是很重要的。. 七、. Src family kinases 參與在 LPS 與 PGN 所誘導的 sustained. ERK activation。. 在先前的研究中指出 Src family kinases 在許多不同的細胞型態中都 有一個重要的功能,就是做為訊息傳遞的 co-transducers (Leu and Maa, 2003; Lowell and Berton, 1999),而巨噬細胞中主要的 SFK 為 Lyn、 Fgr 和 Hck。基於文獻中相關的實驗都是以 LPS 或 PGN 對巨噬細胞 做短時間的處理,且也都只看到 transient ERK activation;由於 SFKs 活性增加可促使 ERK 活化,因此我們懷疑巨噬細胞中 sustained ERK activation 係因 LPS 或 PGN 長期刺激而來。為回答這個問題,我們先 將巨噬細胞以 LPS (100 ng/ml)或 PGN (5 μg/ml) 處理兩小時後,讓 transient ERK activation 回到原點,加入 PP2 (Src family kinases inhibitor;10 μM) 後再經 46 hr,收集 cell lysates 與 conditioned medium 來分析 SFK 對 sustained ERK activation 及 TNF production 的影響。 由 Fig 7 的結果顯示,在合併處理 PP2 的組別中,sustained ERK activation 與 TNF的釋放量都有明顯的被抑制,因此我們認為 Src 38.
(48) family kinases 參與在 LPS 與 PGN 所主導巨噬細胞之 TNF釋放過程 中。 八、. Src 參與在 LPS、PGN 與 SNAP 所誘導巨噬細胞的 sustained. ERK activation 及 TNFproduction。. 由 Lyn、Fgr、Hck 不存在時,巨噬細胞因 LPS 刺激而產生的 TNF 並未明顯降低的情況來看,應有 Lyn、Fgr 及 Hck 以外的 tyrosine kinases 參與在 LPS 誘導巨噬細胞釋放 TNF的路徑中 (Meng and Lowell, 1997)。另一方面,我們實驗室也提出了在 LPS 刺激巨噬細胞活化移 行的路徑中,Src 扮演著一個重要的角色 (Maa et al., 2008)。由於 Src family kinases 參與在 LPS 與 PGN 所誘導的 sustained ERK activation, 因此我們想知道 Src 是否參與在 LPS 或 PGN 刺激下之巨噬細胞釋放 TNF的路徑中。我們利用 Raw264.7 及表達 vector control (ctrl cell), 表達 src-specific siRNA 的細胞 Src-attenuated Raw264.7 (siRNA) 以及 表達 ectopic Src 的 Src-attenuated Raw264.7 (siRNA/Src) 來做實驗。這 些不同的細胞以 LPS 或 PGN 處理 48 小時後觀察 Src 與 ERK 的活性 與 TNF 分泌的量。由 Fig 8 及 Fig 9 顯示,當 Src-attenuated Raw264.7 (siRNA) 受到 LPS 或 PGN 刺激,Src 的活性 (Src-pY416) 與 Src 的表 現量與 vector control (ctrl cell) 相較下明顯的降低,此外 ERK 的活性 39.
(49) (pERK) 還有 TNF 的表現量也都顯著的降低,而在 LPS 或 PGN 刺 激 ectopic Src 的 Src-attenuated Raw264.7 (siRNA/Src) 後,Src 的活性 (Src-pY416) 與 Src 的表現量與 Src-attenuated Raw264.7 (siRNA) 相較 明顯的回升,ERK 的活性 (pERK) 還有 TNF 的表現量也隨著 Src 的表現量上升而增加 (Fig 9) ,這也意味著在 LPS 與 PGN 誘導產生 TNF 的路徑中,Src 扮演一個重要的角色。在前面的實驗我們已經 知道 LPS、PGN iNOS NO/cGMP ERK TNF,而 Src 又受 到 NO/cGMP 調控,進而我們利用 SNAP 處理 Src-attenuated Raw264.7 (siRNA) 以及表達 ectopic Src 的 Src-attenuated Raw264.7 (siRNA/Src) 去觀察 ERK 與 TNF的表現情形。實驗結果得知,在 Src 缺乏的情形 下,ERK 的活性與 TNF的表現量都被明顯的抑制 (Fig 10),這樣的 結果指出 Src 不僅在 LPS 與 PGN 活化巨噬細胞 migration 中扮演重要 的角色,相對的在 LPS 與 PGN 誘導 TNF的表現也具有重要的意義。 . . 40.
(50) 肆、 討論 ( Discussion ). 41.
(51) TLRs 可辨識革蘭氏菌細胞壁的成分的 LPS 以及 PGN。TLR-4 目 前是人類被發現的第一個 Toll 相關蛋白,隨著對 TLR-4 分子結構、 識別受體、訊號傳導途徑及基因缺陷型動物模型的深入探討,我們益 形了解 TLR-4 在人體免疫系統的重要性。當 LPS 與 PGN 鍵結到 TLRs 後,會增加 NF-基因轉錄活性,促使 IL-1、IL-2、IL-6 、IL-8、 TNF及 iNOS 的產生,引發發炎反應(Lee et al., 2005; Wright et al., 1990)。 LPS已經被證實能誘導許多細胞內的訊息傳遞而導致發炎反應, 包括活化NF-及MAPKs family的成員如 ERK 1/2 (Liu et al., 1994; van der Bruggen et al., 1999)、JNKs (Hambleton et al., 1996),和 p38 (Casey et al., 1994)。文獻也指出 LPS 可透過活化 ERK1/2 訊息傳遞 途徑,來促進細胞轉錄因子 Elk-1 的磷酸化,使 Elk-1 和 SRF 結 合在一起,進一步活化 egr-1 promoter 增加 Egr-1 來增加 TNF 基因的表現 (Guha et al., 2001)。文獻也指出當使用 MEK 抑制劑 U0126 時,可減少 LPS 所誘導發炎細胞激素 (如 IL-1,IL-8,TNF) 的分泌及 iNOS 的蛋白表現。而類似的情形也發生在TLR2的活化過 程。由此可知ERK1/2在誘發細胞內訊息傳遞中,扮演一個重要性的 角色 (Scherle et al., 1998)。 以我們的實驗結果,我們釐清了在 LPS 刺激巨噬細胞後,所產生 42.
(52) 的 iNOS、ERK、Src、TNF 彼此之間的關聯性。最初我們以 LPS 與 PGN 處理巨噬細胞,同時發現 iNOS 的產生與 TNF的釋出明顯 增加,這固然可以 inos 和 tnfgene 皆為 NF-的標的基因,故 iNOS 與 TNF的量同步上升來解釋。但由於 NO 為一 second messenger, 故我們不排除有一個可能,那就是 iNOS 產生的 NO 會主導 TNF的 產生。我們利用 1400W 與 AG 抑制 iNOS,利用 ODQ 抑制 sGC,實 驗結果顯示這些藥物皆能抑制 LPS- and PGN-induced TNFsecretion, 確認了 iNOS 與 sGC 參與在 LPS 與 PGN 誘導 TNF-分泌的過程中 (Fig 1)。而文獻也指出於 human neutrophil 中,NO donors (SNAP) 可 以調節內毒素誘導 TNFsynthesis (Van Dervort et al., 1994);在 iNOS transfected human U937 cells 中,NO 可增加 TNF產生,當 U937 cells 缺少了 sGC,U937 cells 則無法被 NO 誘導產生 TNF(Yan et al., 1997)。 另 外 NO 及 cGMP 都 可 增 加 human peripheral blood 的 TNF synthesis(Wang et al., 1997)。 sGC 是 NO 主要的 target,當 NO 作用於 sGC 可促使細胞內訊息 傳遞(Bogdan, 2001)。我們使用 (1) NO donor, SNAP 及 cGMP 類似物, 8-br-cGMP,同樣的都可以增加 TNF的釋放 (Fig 1C),證明了 NO/cGMP 參與了 TNF釋出。(2) 我們也利用了 iNOS 有缺陷老鼠 的巨噬細胞,發現與正常巨噬細胞相較,其在給予 LPS 與 PGN 刺激 43.
(53) 後 TNF分泌量大幅減少 (Fig 2),但 SNAP 與 8-br-cGMP 皆可誘導 WT 與 iNOS-/- PEMs 產生 TNF。另外以 reverse transcripation-PCR 分 析,看 RNA 的結果,我們也證明了先前實驗的結果。使用 AG 及 ODQ 分別都可抑制 LPS 所誘導的 Raw 264.7 巨噬細胞 tnfmRNA表現量; SNAP 和 8-br-cGMP 的確可以使 tnfmRNA 表現量增加 (附圖二)。 這也證明了 cGMP 確實會參與 NO 誘導 Raw 264.7 巨噬細胞表現 tnfmRNA 的過程。cGMP effector proteins 已知有 phosphodiesterases (PDEs)、cGMP-dependent protein kinases (PKGs) 、cGMP-gated ion channels。究竟何者參與了 NO/cGMP-induced TNF來增加發炎反應, 值得進一步探討。 也有文獻指出 LPS 透過活化 ERK 的訊息傳遞途徑,增加了 tnf 基因的轉錄及 TNFsecretion (Luyendyk et al., 2008; van der Bruggen et al., 1999; Yao et al., 1997)。和此結果互相一致的是在我們的實驗裡 PD98059 (MEK 抑制劑) 可以有效抑制 LPS、SNAP、8-br-cGMP 產生 TNF (附圖三) ,這意味著 iNOS 及 NO/cGMP pathway 參與在 LPS 誘導 ERK 的活化,進而造成 TNF的釋出。而這樣的情形也在 PGN 的處理下觀察到,我們更進一步的用 wild-type (WT) 及 iNOS-/- PEMs 來檢視。我們發現以 LPS 與 PGN 處理的 iNOS-/- PEMs 其 ERK 不能 活化 (Fig 6、附圖四),但 SNAP 和 8-br-cGMP 則皆可促進 ERK 活 44.
(54) 化 (附圖四)。有趣的是,我們觀察到了 sustained ERK 的活化是影響 TNF釋放的主要因素之一,也說明了在巨噬細胞的 iNOS 為 LPS及 PGN-induced sustained ERK activation 所必需。根據這些實驗結果 我們推論 LPS 與 PGN 刺激巨噬細胞後會因 NF-活化而產生 iNOS。 iNOS 的產物 NO 與及下游的 sGC 產物 cGMP 會去啟動訊息傳遞路徑。 藉由 Src 的蛋白量及其活性的增加,細胞內會發生 sustained ERK activation,增加 tnf基因的轉錄及 TNF的釋出。 一般認為 iNOS 及 TNF在 LPS 與 PGN 刺激後,是蛋白量 的增加是並行、互不干擾的。但我們實驗顛覆此一傳統的看法。我們 發現LPS and PGN-induced TNF是需要 iNOS 的參與。因 iNOS 不 存在或其活性受阻時,TNF的產生就會下降。因此這麼一個LPS、 PGN/iNOS/sGC/Src/ERK/TNF-的pathway就被確立 (Fig 10)。我們不 排除這樣的pathway有可能存在巨噬細胞的其他TLR訊息路徑中,也 不排除其有可能也發生於巨噬細胞外其他種類的細胞。. 45.
(55) 伍、 參考文獻(reference). 46.
(56) Akira S, Uematsu S, Takeuchi O (2006). Pathogen recognition and innate immunity. Cell 124: 783-801. Bogdan C (2001). Nitric oxide and the immune response. Nat Immunol 2: 907-16. Boggon TJ, Eck MJ (2004). Structure and regulation of Src family kinases. Oncogene 23: 7918-27. Cano E, Hazzalin CA, Kardalinou E, Buckle RS, Mahadevan LC (1995). Neither ERK nor JNK/SAPK MAP kinase subtypes are essential for histone H3/HMG-14 phosphorylation or c-fos and c-jun induction. J Cell Sci 108 ( Pt 11): 3599-609. Caroff M, Karibian D, Cavaillon JM, Haeffner-Cavaillon N (2002). Structural and functional analyses of bacterial lipopolysaccharides. Microbes Infect 4: 915-26. Casey JR, Petranka JG, Kottra J, Fleenor DE, Rosse WF (1994). The structure of the urokinase-type plasminogen activator receptor gene. Blood 84: 1151-6. Davis RJ (1994). MAPKs: new JNK expands the group. Trends Biochem Sci 19: 470-3. Derijard B, Hibi M, Wu IH, Barrett T, Su B, Deng T et al (1994). JNK1: a protein kinase stimulated by UV light and Ha-Ras that binds and phosphorylates the c-Jun activation domain. Cell 76: 1025-37. Duffield JS (2003). The inflammatory macrophage: a story of Jekyll and Hyde. Clin Sci (Lond) 104: 27-38. Dziarski R, Gupta D (2006). Mammalian PGRPs: novel antibacterial proteins. Cell Microbiol 8: 1059-69. Ebisuya M, Kondoh K, Nishida E (2005). The duration, magnitude and compartmentalization of ERK MAP kinase activity: mechanisms for providing signaling specificity. J Cell Sci 118: 2997-3002. 47.
(57) Grewal SS, York RD, Stork PJ (1999). Extracellular-signal-regulated kinase signalling in neurons. Curr Opin Neurobiol 9: 544-53. Guha M, Mackman N (2002). The phosphatidylinositol 3-kinase-Akt pathway limits lipopolysaccharide activation of signaling pathways and expression of inflammatory mediators in human monocytic cells. J Biol Chem 277: 32124-32. Guha M, O'Connell MA, Pawlinski R, Hollis A, McGovern P, Yan SF et al (2001). Lipopolysaccharide activation of the MEK-ERK1/2 pathway in human monocytic cells mediates tissue factor and tumor necrosis factor alpha expression by inducing Elk-1 phosphorylation and Egr-1 expression. Blood 98: 1429-39. Gupta D, Jin YP, Dziarski R (1995). Peptidoglycan induces transcription and secretion of TNF-alpha and activation of lyn, extracellular signal-regulated kinase, and rsk signal transduction proteins in mouse macrophages. J Immunol 155: 2620-30. Hambleton J, Weinstein SL, Lem L, DeFranco AL (1996). Activation of c-Jun N-terminal kinase in bacterial lipopolysaccharide-stimulated macrophages. Proc Natl Acad Sci U S A 93: 2774-8. Han J, Lee JD, Bibbs L, Ulevitch RJ (1994). A MAP kinase targeted by endotoxin and hyperosmolarity in mammalian cells. Science 265: 808-11. Hayden MS, West AP, Ghosh S (2006). NF-kappaB and the immune response. Oncogene 25: 6758-80. Janeway CA, Jr. (2001). How the immune system protects the host from infection. Microbes Infect 3: 1167-71. Kawai T, Akira S (2006). TLR signaling. Cell Death Differ 13: 816-25. Keestra AM, de Zoete MR, van Aubel RA, van Putten JP (2007). The central leucine-rich repeat region of chicken TLR16 dictates unique ligand specificity and species-specific interaction with TLR2. J Immunol 48.
(58) 178: 7110-9. Knowles RG, Moncada S (1994). Nitric oxide synthases in mammals. Biochem J 298 ( Pt 2): 249-58. Krumenacker JS, Hanafy KA, Murad F (2004). Regulation of nitric oxide and soluble guanylyl cyclase. Brain Res Bull 62: 505-15. Kumar A, Middleton A, Chambers TC, Mehta KD (1998). Differential roles of extracellular signal-regulated kinase-1/2 and p38(MAPK) in interleukin-1beta- and tumor necrosis factor-alpha-induced low density lipoprotein receptor expression in HepG2 cells. J Biol Chem 273: 15742-8. Lee SH, Lee SY, Son DJ, Lee H, Yoo HS, Song S et al (2005). Inhibitory effect of 2'-hydroxycinnamaldehyde on nitric oxide production through inhibition of NF-kappa B activation in RAW 264.7 cells. Biochem Pharmacol 69: 791-9. Leu TH, Maa MC (2003). Functional implication of the interaction between EGF receptor and c-Src. Front Biosci 8: s28-38. Levy BT, Sorge LK, Meymandi A, Maness PF (1984). pp60c-src Kinase is in chick and human embryonic tissues. Dev Biol 104: 9-17. Liu MK, Herrera-Velit P, Brownsey RW, Reiner NE (1994). CD14-dependent activation of protein kinase C and mitogen-activated protein kinases (p42 and p44) in human monocytes treated with bacterial lipopolysaccharide. J Immunol 153: 2642-52. Lowell CA, Berton G (1999). Integrin signal transduction in myeloid leukocytes. J Leukoc Biol 65: 313-20. Luyendyk JP, Schabbauer GA, Tencati M, Holscher T, Pawlinski R, Mackman N (2008). Genetic analysis of the role of the PI3K-Akt pathway in lipopolysaccharide-induced cytokine and tissue factor gene expression in monocytes/macrophages. J Immunol 180: 4218-26. 49.
(59) Maa MC, Chang MY, Chen YJ, Lin CH, Yu CJ, Yang YL et al (2008). Requirement of inducible nitric-oxide synthase in lipopolysaccharide-mediated Src induction and macrophage migration. J Biol Chem 283: 31408-16. Marshall CJ (1995). Specificity of receptor tyrosine kinase signaling: transient versus sustained extracellular signal-regulated kinase activation. Cell 80: 179-85. Meng F, Lowell CA (1997). Lipopolysaccharide (LPS)-induced macrophage activation and signal transduction in the absence of Src-family kinases Hck, Fgr, and Lyn. J Exp Med 185: 1661-70. Moncada S, Martin JF (1993). Evolution of nitric oxide. Lancet 341: 1511. Myhre AE, Aasen AO, Thiemermann C, Wang JE (2006). Peptidoglycan--an endotoxin in its own right? SHOCK 25: 227-35. Nathan C (1994). Nitric oxide and biopterin: a study in Chiaroscuro. J Clin Invest 93: 1875-6. Nishida E, Gotoh Y (1993). The MAP kinase cascade is essential for diverse signal transduction pathways. Trends Biochem Sci 18: 128-31. Roskoski R, Jr. (2004). Src protein-tyrosine kinase structure and regulation. Biochem Biophys Res Commun 324: 1155-64. Scherle PA, Jones EA, Favata MF, Daulerio AJ, Covington MB, Nurnberg SA et al (1998). Inhibition of MAP kinase kinase prevents cytokine and prostaglandin E2 production in lipopolysaccharide-stimulated monocytes. J Immunol 161: 5681-6. Thanos D, Maniatis T (1995). NF-kappa B: a lesson in family values. Cell 80: 529-32. van der Bruggen T, Nijenhuis S, van Raaij E, Verhoef J, van Asbeck BS (1999). Lipopolysaccharide-induced tumor necrosis factor alpha 50.
(60) production by human monocytes involves the raf-1/MEK1-MEK2/ERK1-ERK2 pathway. Infect Immun 67: 3824-9. Van Dervort AL, Yan L, Madara PJ, Cobb JP, Wesley RA, Corriveau CC et al (1994). Nitric oxide regulates endotoxin-induced TNF-alpha production by human neutrophils. J Immunol 152: 4102-9. Wang S, Yan L, Wesley RA, Danner RL (1997). Nitric oxide increases tumor necrosis factor production in differentiated U937 cells by decreasing cyclic AMP. J Biol Chem 272: 5959-65. Woltmann A, Hamann L, Ulmer AJ, Gerdes J, Bruch HP, Rietschel ET (1998). Molecular mechanisms of sepsis. Langenbecks Arch Surg 383: 2-10. Wong FS, Wen L (2008). Toll-like receptors and diabetes. Ann N Y Acad Sci 1150: 123-32. Wright SD, Ramos RA, Tobias PS, Ulevitch RJ, Mathison JC (1990). CD14, a receptor for complexes of lipopolysaccharide (LPS) and LPS binding protein. Science 249: 1431-3. Yan L, Wang S, Rafferty SP, Wesley RA, Danner RL (1997). Endogenously produced nitric oxide increases tumor necrosis factor-alpha production in transfected human U937 cells. Blood 90: 1160-7. Yao J, Mackman N, Edgington TS, Fan ST (1997). Lipopolysaccharide induction of the tumor necrosis factor-alpha promoter in human monocytic cells. Regulation by Egr-1, c-Jun, and NF-kappaB transcription factors. J Biol Chem 272: 17795-801. Zhou X, Yang W, Li J (2006). Ca2+- and protein kinase C-dependent signaling pathway for nuclear factor-kappaB activation, inducible nitric-oxide synthase expression, and tumor necrosis factor-alpha production in lipopolysaccharide-stimulated rat peritoneal macrophages. J Biol Chem 281: 31337-47. 51.
(61) 陸、 圖表(Figure). 52.
(62) Figure 1. NO/cGMP is involved in PGN-induced TNFsecretion. (A) Raw264.7 cells were treated with or without LPS (100 ng/ml) or PGN (5g/ml) for 48 hrs. Equal amounts of lysates (80 g) from each sample were resolved by SDS-PAGE and probed with anti-iNOS and anti-actin antibodies. (B) Raw264.7 cells were pretreated with AG (2 mM) or ODQ (100 M) for 30 m and then stimulated with or without PGN 53.
(63) (5g/ml) for 48 h. (C) Raw264.7 cells were treated without or with SNAP (100 M) and 8-br-cGMP (cGMP, 100M) for 48 h. The concentration of TNFin culture medium of each sample was determined by ELISA ***,P<0.001.. 54.
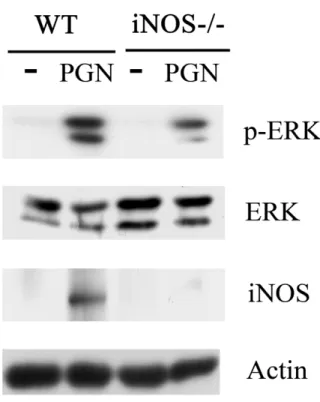
(64) Figure 2. iNOS is required for TNF secretion mediated by LPS and PGN. Peritoneal macrophages from wild type (WT) and iNOS-/- mice were treated without or with LPS (100 ng/ml) (A), PGN (5g/ml) (B), SNAP (100 M) (C), or 8-bro-cGMP (cGMP, 100M) (D) for 48 h, then the concentration of TNFin culture medium of each sample was determined by ELISA.. Meanwhile, total lysates (80 g) from each. group in part (A) and (B) were resolved by SDS-PAGE and probed with antibodies against iNOS and actin. ***, P<0.001. 55.
(65) Figure 3. PGN-induced ERK activation is inhibitor by AG or ODQ. Raw264.7 cells were pretreated without or with AG (2mM) and ODQ (100 M) for 30 m and then were stimulated without or with and PGN (2 g/ml) for 48hr. Equal amounts of lysates (80 g) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated.. 56.
(66) Figure 4. Sustained ERK activation mediated by LPS and PGN. Raw264.7 cells were stimulated with LPS (100 ng/ml) (A) and PGN (2 g/ml) (B) for various time as indicated.. Equal amounts of lysates (80. g) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated.. 57.
(67) Figure 5. LPS- and PGN-mediated sustained ERK activation was AG-sensitive.. Raw264.7 cells pretreated without or with AG were stimulated with LPS (100 ng/ml) (A) and PGN (2 g/ml) (B).. Equal amounts of lysates (80. g) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated.. 58.
(68) Figure 6. iNOS is involved in PGN-mediated ERK activation. Primary peritoneal macrophages from wild type (WT) and iNOS-/- mice were treated without or with PGN (2 g/ml) for 48 h, then equal amounts of lysates (25 g) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated.. 59.
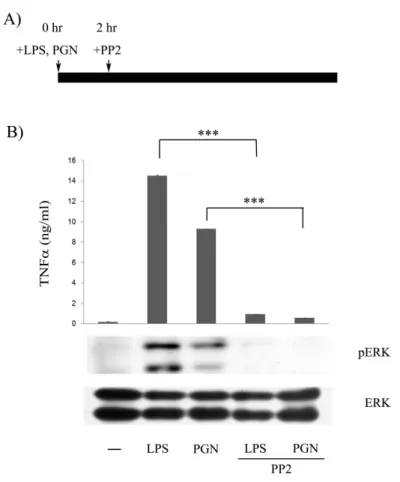
(69) Figure 7. Involvement of Src family kinases in the late stage of LPSand PGN-induced TNF secretion and ERK activation. (A) Flow diagram describing the administration of PP2 and LPS. (B) Raw264.7 cells were incubated with PP2 (10 M) after 2 h LPS (100 ng/ml) and PGN (2 g/ml) stimulation. Equal amounts of lysates (80g) from each group were separated by SDS-PAGE and probed with antibodies as indicated. The concentration of TNFin culture medium of each group was determined by ELISA. ***, P<0.001.. 60.
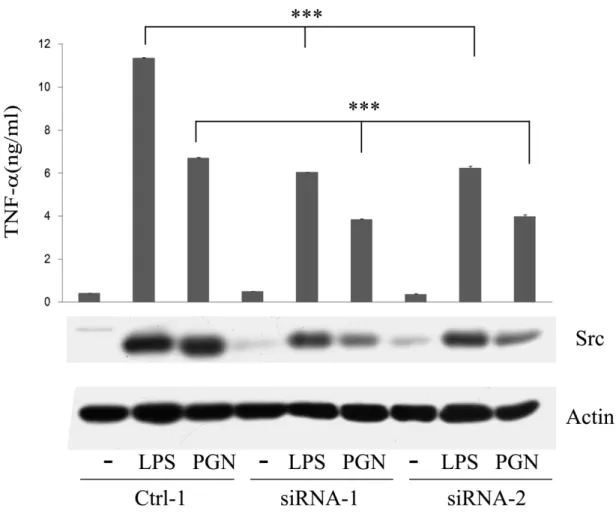
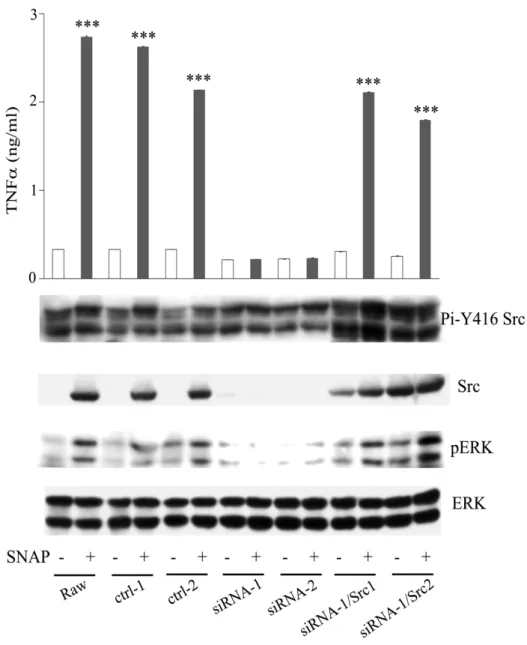
(70) Figure 8. LPS- and PGN-mediated TNF secretion was inhibited by src-specific siRNA. Raw264.7 cell and its derived control (ctrl-1), Src attenuated cells (siRNA-1, -2) were stimulated without or with LPS and PGN for 48h. Equal amounts of lysates (80g) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated. The concentration of TNFin culture medium of each group was determined by ELISA. ***, P<0.001.. 61.
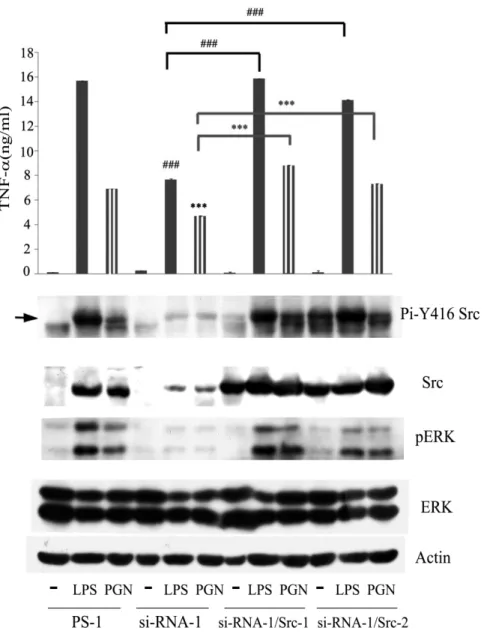
(71) Figure 9. LPS- and PGN-mediated TNF secretion was inhibited by src-specific siRNA, which could be reversed by ectopic Src. Raw264.7 cell and its derived control (ctrl-1, -2), Src attenuated cells (siRNA-1, -2) and Src attenuated cells harboring plasmid encoding avian Src (siRNA-2/Src1, siRNA-2/Src2) were stimulated without or with LPS and PGN for 48h. Equal amounts of lysates (80g) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated. The concentration of TNFin culture medium of each group was determined by ELISA. The arrow indicates the position of Src.***, P<0.001.. 62.
(72) Figure 10. SNAP mediated TNF secretion was inhibited by src-specific siRNA but ectopic Src could reverse this phenomenon. Raw264.7 cell and its derived control (ctrl-1, -2), Src attenuated cells (siRNA-1, -2) and Src attenuated cells harboring plasmid encoding avian Src (siRNA-2/Src1, siRNA-2/Src2) were stimulated without or with SNAP. for 48h.. Equal amounts of lysates (80g) from each sample. were resolved by SDS-PAGE and probed with antibodies as indicated. The concentration of TNFin culture medium of each group was determined by ELISA.***, P<0.001. 63.
(73) Figure 11. The proposed NO/sGC/Src/ERK signaling pathway for TNF secretion in LPS- and PGN- stimulated macrophages. Following LPS and PGN treatment, the signal is transduced into macrophages via Toll-like receptor 4 and Toll-like receptor 2, which leads to NF-activation. Traditional model (A) indicated TNF and iNOS were independent. But our lab study indicated activated NF- increases the expression of iNOS. NO produced by iNOS contributes to Src and sustained ERK activation, TNF generation (B).. 64.
(74) 附圖一、structure of lipopolysaccharide and peptidoglycan. a. structure of lipopolysaccharide b. structure of peptidoglycan. 65.
(75) 附圖二、LPS-induced tnf transcript is sensitive to inhibitors of iNOS and sGC. Raw264.7 cells were pretreated without or with AG (2 mM), ODQ (100 M) for 30 min, and then cells were stimulated without or with LPS, SNAP and 8-br-cGMP for 48 hrs. The amount of tnf transcript was analysis by RT-PCR. gapdh was utilized as an internal control for amplification efficiency.. 66.
(76) 附圖三、MEK inhibitor PD98059 suppressed the SNAP- and 8-br-cGMP-induced TNFproduction. (A) Raw264.7 cells were pretreated without or with 1400W (100 M), ODQ (100 M) for 30 min and then were stimulated without or with LPS (100 ng/ml). Lysates from each group were separated by SDS-PAGE and probed. with. antibodies. against. either. phosphorylated. or. nonphosphorylated ERK. The concentration of TNF-in culture medium was determined by ELISA. Raw264.7 cells were preincubated without or with PD98059 (10 M) for 1 hr, then stimulated without or with (B) LPS (100 ng/ml), (C) SNAP (100 M) and 8-bro-cGMP (cGMP, 100M) for 48 hrs. The content of phosphorylated ERK, ERK and the concentration of TNF-in culture medium in each group were determined by Western blot analysis and ELISA, respectively.. 67.
(77) 附圖四、LPS-mediated ERK activation was abrogated in iNOS null macrophages, which could be restored by SNAP and 8-br-cGMP. (A) Primary peritoneal macrophages from wild type (WT) and iNOS knockout (iNOS-/-) mice were treated without or with (A) LPS (100 ng/ml), (B) SNAP (100 M) and 8-bro-cGMP (cGMP, 100M) for 48 hrs, then equal amounts of lysates (25 g) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated.. 68.
(78)
數據

+7
相關文件
• 由於細胞代謝、紫外線的電離輻射、不良的生 活習慣 ( 煙、酒、肥胖 ) 、各種感染物所引起 的中性粒細胞、巨噬細胞的激活, 會催化分子 氧發生單價還原產生的 "
A genre is more dynamic than a text type and is always changing and evolving; however, for our practical purposes here, we can take genre to mean text type. Materials developed
3.16 Career-oriented studies provide courses alongside other school subjects and learning experiences in the senior secondary curriculum. They have been included in the
1.4 For education of students with SEN, EMB has held a series of consultative meetings with schools, teachers, parents and professional bodies to solicit feedback on
There were 216 MICE events held in hotels, of which 210 were meetings & conferences and 6 were exhibitions, drawing a total of 23,878 participants and attendees. Meanwhile, 94
straight red hair long brown hair gray hair curly red hair.. What does she
According to Japan’s transport ministry, there were 613 cases of train services suspended or delayed for at least 30 minutes resulting from collisions with deer and other
[r]
