Analysis of the AXIN2 Gene Expression in Non-small Cell Lung
Cancer
Ruo-Chia Tseng
1, Chieh Tseng
1,2, Han-Shui Hsu
3, Yi-Ching Wang
1*
1Department of Life Sciences, National Taiwan Normal University Taipei, Taiwan
2 Institute of Bioinformatics and Structure Biology, National Tsing Hua University Hsinchu, Taiwan
3Division of Thoracic Surgery, Taipei Veterans General Hospital Taipei, Taiwan
(Received: 2 May 2005, accepted: 2 June 2005)
ABSTRACT
Lung cancer is one of the most common malignancies in the world and is the leading cause of cancer deaths in industrial countries, including Taiwan. Therefore, it is important to identify the etiologically associated gene(s) involved in lung cancer. Wingless (Wnt) signaling is involved in cellular proliferation, motility, and tumorigenesis. Axis inhibition protein 2 (AXIN2) is a scaffold protein for the degradation complex in the Wnt signaling pathway. Therefore, AXIN2 could act as a tumor suppressor. To determine whether the AXIN2 expression is altered in non-small cell lung cancer (NSCLC), we analyzed the AXIN2 mRNA and protein expression by reverse transcriptase polymerase chain reaction and immunohistochemistry in 56 NSCLC and non-cancerous matched tissues. Our results indicated that 45% of NSCLC patients showed low mRNA expression. Chi-squared analysis indicated that low mRNA expression appeared to be more frequent in early stage (54%) than in late stage (26%) (P = 0.048). A more significant correlation of low AXIN2 mRNA expression in elderly patients (57%) than in young patients (31%) (P = 0.052) was observed. In addition, 36% of NSCLC patients exhibited reduction or complete loss of AXIN2 protein expression. Chi-squared analysis indicated a highly significant correlation between aberrant AXIN2 expression and differentiation, recorded as negative staining in 52% (16/31) of NSCLC with well and moderate differentiation, compared with 20% (4/20) of NSCLC with poor differentiation (P = 0.023). Our data suggested that the alteration of AXIN2 expression might play an etiological role in NSCLC tumorigenesis.
Key words: AXIN2, NSCLC
Introduction
Lung cancer is one of the most common malignancies in the world and is the leading cause of cancer deaths in industrial countries, including Taiwan (Minna et al., 1997). Lung cancer is often classified into two major types: small-cell lung cancer (SCLC) and non-small cell lung cancer (NSCLC) (Minna et al., 1997). Human cancers develop through multi-step processes involving mutations in several types of genes, including those inactivating recessive tumor suppressor genes (TSGs), those activating dominant
oncogenes, and those inactivating genes involved in DNA repair or replication (Fearon, 1997). However, many of the lung cancer-related TSGs remain undefined.
Wingless (Wnt) signaling via β-catenin is involved in vertebrate development, morpho- genesis, cellular proliferation, differentiation, motility and tumorigenesis (Wodarz and Nusse, 1998; Salahshor and Woodgett, 2005). Wnt signaling is initiated by secreted Wnt proteins, which bind to a class of seven-pass trans- membrane receptors encoded by the frizzled genes (Polakis, 2000). Activation of receptor leads to the
phosphorylation of the disheveled (DVL) protein. DVL antagonizes the function of the glycogen synthase kinase 3 (GSK3)/Axis inhibition protein 1 (AXIN1)/Axis inhibition protein 1 (AXIN2)/ adenomatous polyposis coli (APC) complex (Boutros et al., 2000), blocking β-catenin degradation, so that the phosphorylation of β-catenin is reduced. The unphosporylated β-catenin accumulates in the cytoplasm and translocates into the nucleus. Nuclear β-catenin interacts with the T-cell specific factor/lymphoid enhancer binding factor 1 transcription factor, and thereby activates the expression of the specific Wnt responsive genes such as c-jun, c-myc, and cyclin D1 (Morin et al., 1997). Therefore, regulation of cytoplasmic β-catenin levels is crucial, because deregulation of this system plays a critical role in cellular proliferation, differentiation, motility and possibly tumorigenesis.
The levels of β-catenin in cytoplasm is controlled by the negative regulators AXIN1 and AXIN2, which interact with the APC tumor suppressor and GSK3 (Behrens et al., 1998; Hart
et al., 1998). In the absence of Wnt signaling, the
cytoplasmic concentrations of β-catenin are kept low through degradation. This degradation depends on the phosphorylation of β-catenin by GSK3. The phosphorylated β-catenin is recognized rapidly by the proteasome and destroyed (Kitagawa et al., 1999). AXIN2 is upregulated in response to increased β-catenin concentrations and thus serves to limit the duration and intensity of the Wnt signal (Woodgett, 2005). The role of AXIN2 in this signaling pathway predicts that it could function as a tumor suppressor. The AXIN2 gene loci can be mapped to chromosome 17q24.3 (Mai et al., 1999). Molecular studies revealed that AXIN2 contains 10 exons spanning more than 2.5kb (Dong et al., 2001). If the function of the AXIN2 gene is lost, β-catenin is no longer degraded. This will lead to β-catenin accumulation in the cytoplasm and may result in tumorigenesis.
The analysis of AXIN2 mRNA was only reported for colon, ovarian, and liver cancers (Yan
et al., 2001; Lustig et al., 2002; Koch et al., 2004).
However, reports for mRNA expression of the
AXIN2 gene are quite limited regarding primary
lung cancers. To better define the alterations in AXIN2 expression in the pathogenesis of NSCLC,
we have recruited 56 primary surgically resected NSCLC tumors and analyzed the mRNA and protein expression in these tumors. The data indicated that 45% and 36% of patients showed low mRNA and protein expressions, respectively. In addition, the alteration of AXIN2 expression was associated with lung cancers in early stages. Materials and Methods
Sample preparation and clinical characterization of patients
Surgically resected tumors and surrounding normal tissues from 56 patients with NSCLC were collected between the years 2002 and 2004. Of these patients, 39 had adenocarcinomas (ADs) and 17 squamous carcinomas (SQs). The histology of the tumor types and their stages were determined according to the World Health Organization classification and the TNM system, respectively. Information on the age, sex, and smoking history of the patients was obtained from hospital records (Table 1).
Surgically resected tumor samples were immediately snap-frozen, and subsequently stored in liquid nitrogen. For the RNA expression assay, total RNA was prepared from tumors and normal lung tissues, using Trizol reagent (Invitrogen, Carlsbad, California). The synthesized cDNA was made using SuperScriptTM reverse transcriptase
(Invitrogen) with the protocols provided by the manufacturer.
Analysis of protein expression: immunohistochemistry (IHC) assay
Paraffin blocks of tumors were cut into 5 µm slices and then processed using standard deparaffinization and rehydration techniques. Polyclonal antibody for AXIN2 (1:500) (Santa Cruz Biotechnology, CA) was used as the primary antibody to detect the protein expression. The primary antibody was detected using biotinylated secondary antibody (DAKO, Carpinteria, California) according to the manufacturer’s instructions. The sections were then counterstained with haematoxylin. The evaluation of the IHC was conducted blindly, without knowledge of the clinical and pathologic characteristics of the cases. Staining for AXIN2 was scored as 3 if >70% tumor cells were positive; 2 if 41-70% tumor cells were positive; 1 if 5-40% cells were positive; and 0 if <5% cells were
positive. The stains were graded negative when the score was 1 or 0, with adequate staining in surrounding normal cells.
Statistical analysis
Analysis of mRNA expression: multiplex reverse transcriptase–polymerase chain reaction (RT–PCR) assay
AXIN2 mRNA expression was assayed by a multiplex RT–PCR analysis using the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene as an internal control. The coding regions of exons 5–7 of the AXIN2 and the GAPDH genes were amplified. Primers for the RT-PCR were the AXIN2F 5´- CAGCGAGTATTACTGCTAC TCGAAA-3´ (forward), AXIN2R 5´- TTTTTTGT GCTTTGGGCACTATG-3´ (reverse). Reactions were carried out in a volume of 25 µL with 1 µL cDNA and 0.25 pmol primers on a DNA thermal cycler. PCR was performed for 32 cycles with annealing temperature of 70°C. The number of cycles and the amount of primers and cDNA used were determined to provide quantitative amplification during multiplex RT-PCR. To quantify the relative levels of gene expression in the multiplex RT–PCR assay, the value for the internal standard (GAPDH) in each test tube was used as the baseline value for gene expression in that sample, and a relative value was calculated for the target gene (AXIN2) transcript amplified from each tumor and matched normal sample. Tumor cells that exhibited mRNA expression below 50% of that of normal cells were deemed to have a low expression pattern, as suggested by a previous study (Hsu et
al., 2004).
Pearson’s χ2 test was used to compare the
frequency of AXIN2 alterations in NSCLC patients with different characteristics, including age, sex, and smoking status, and various clinicopathological parameters, such as tumor type, tumor stage, and differentiation status. We defined two age groups by median age. Smokers included both current smokers and ex-smokers. Results
AXIN2 protein expression in primary NSCLC tumors and its correlation with clinical parameters in NSCLC patients
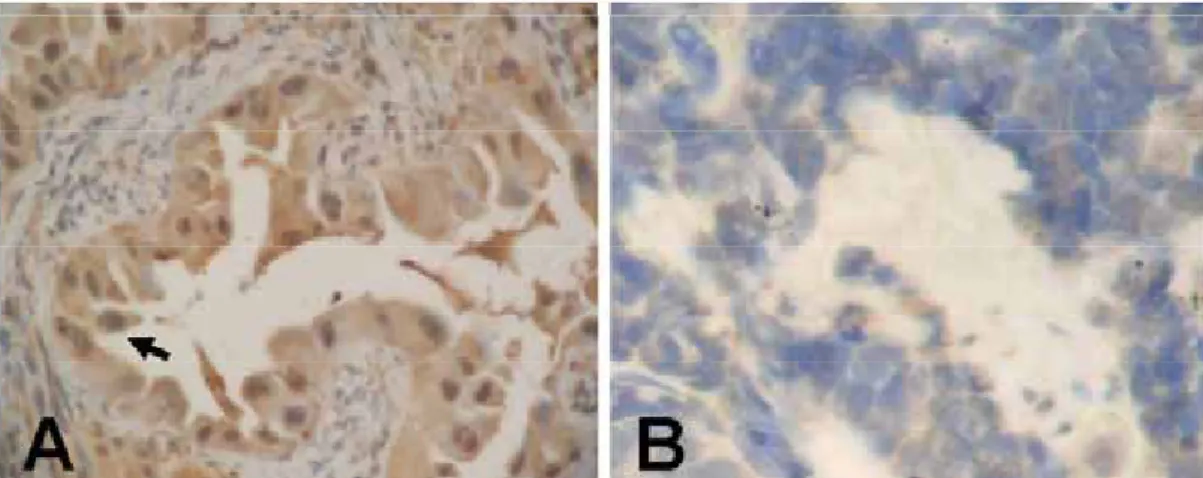
Immunohistochemical staining was performed on 56 tumor samples. Positive immunoreactivity was visible as a brown precipitate (Figure 1A). Twenty lung tumors (36%) showed either low or no staining (Figure 1B). Chi-squared analysis indicated a highly significant correlation between aberrant AXIN2 protein expression and differentiation, recorded as negative staining in 52% (16/31) of NSCLC which were well and moderately differentiated, compared with 20% (4/20) of NSCLC with poor differentiation (P = 0.023) (Table 1).
AXIN2 mRNA expression and its correlation with clinical parameters and protein expression in NSCLC patients
RNAs of high quality were extracted from56 tumors and used for multiplex RT–PCR (Figure 2). Decreased AXIN2 transcripts were shown by
Figure 1. Immunohistochemical analysis of AXIN2 proteins in formalin-fixed paraffin-embedded sections of resected tumor
specimens of the NSCLC. Positive immunoreactivity is visible as a brown precipitate as indicated by an arrow (A). A patient with negative immunoreactivity is shown in (B). Original magnification ×200.
Table 1. Alteration of protein and mRNA expressions of AXIN2 gene in relation to the clinicopathological parameters of 56
NSCLC tumors and molecular data.
AXIN2
Characteristics Total
n Low protein n (%) Low mRNA n (%)
Overall 56 20 (36%) 25 (45%) Clinicalpathological parameters Age <72 26 10 (38) 8 (31) >72 30 10 (33) 17 (57)0.052 Sex Male 44 17 (39) 21 (48) Female 12 3 (25) 4 (33) Smoke No 13 3 (23) 4 (31) Yes 43 17 (40) 21 (49) Tumor type SQ 17 7 (41) 8 (47) AD 39 13 (33) 17 (44) Tumor stage a I+II 37 15 (41) 20 (54)0.048 III+Ⅳ 19 5 (26) 5 (26) Differentiation b Well+Moderate 31 16 (52)0.023 15 (48) Poor 20 4 (20) 9 (45)
Concordant analysis between protein and mRNA expression
mRNA expression
Total
n Low protein n (%)
Wild type 31 2 (6)
Low 25 18 (72)0.000
a. The P values with signification (P < 0.05) were shown as superscripts.
b Total number of sample (n) in tumor differentiation grade category is less than 56. Only the data with defined differential grades were included.
RT–PCR to occur in 25 tumors (45%) (Table 1). Chi-squared analysis showed that low mRNA expression appeared to be more frequent in early stage (54%) than in late stage (26%) (P = 0.048) (Table 1). In addition, our results revealed a significant correlation between elderly patients (57%) and young patients (31%) (P = 0.052) (Table 1).
To investigate whether the low AXIN2 mRNA and protein expressions were correlated, both protein and RNA expression were analyzed using the Chi-squared test. Low protein expression was significantly associated with low mRNA expression (P < 0.001) (Table 1).
Discussion
AXIN2 is believed to act as a scaffold protein, which binds several components of the Wnt/β-catenin pathway and promotes the
phosphorylation of β-catenin by GSK3 and its consequent degradation. Hence, AXIN2 serves as a negative regulator of the signaling pathway. AXIN2 has been shown to act as a tumor suppressor in humans (Lustig et al., 2002). In addition, prior study showed that AXIN2 can
Figure 2. Representative figures for AXIN2 mRNA
expression analysis by the RT-PCR assay. M, molecular marker; N, normal lung tissue; T, tumor tissue of the lung. Patient 1 showed a lower mRNA expression level in the tumor tissue than in its normal tissue. Patient 2 showed a normal expression level for mRNA expression in the tumor tissue compared to its normal tissue.
repress Wnt signaling, leading to reduced cell growth and increased cell death (Hughes and Brady, 2005). Here, we analyzed the alteration of AXIN2 mRNA and protein expression in lung cancer. Immunohistochemical and semi- quantitative RT-PCR analyses showed low mRNA and protein expression in 45% and 36% of patients with NSCLC, respectively. A highly significant correlation was observed between low protein expression and low mRNA expression.
High levels of AXIN2 mRNA expression have been detected in human colon, ovarian and liver cancers (Koch et al., 2004; Lustig et al., 2002; Salahshor and Woodgett, 2005). However, mutations in the AXIN2 gene rarely occur in human cancers. For example, mutations in AXIN2 were found in approximately 20% of colorectal tumors (Liu et al., 2000). A fraction of hepatocellular carcinomas (~10%) with wild-type β catenin have mutations in AXIN1 and AXIN2 (Taniguchi et al., 2002). In addition, no significant overexpression of AXIN2 mRNA was observed in breast, lung, and uterus tumors (Lustig et al., 2002; Yan et al., 2001). Lusting et
al. (2002) previously showed a decrease of
AXIN2 mRNA expression in uterine and lung carcinomas. Their findings were consistent with our data that low AXIN2 mRNA expression occurred in lung cancer. However, no data for AXIN2 protein expression has been reported in lung cancer. To our knowledge, the present study is the first report to show the expression of AXIN2 at both the RNA and protein levels in lung cancer.
In RT–PCR analysis, 25 patients with NSCLC exhibited low or absent AXIN2 mRNA expression. In addition, a high frequency of low AXIN2 mRNA or protein can be detected in early-stage and well and moderately differentiated lung tumors, indicating that loss of AXIN2 might be an early event during lung cancer initiation. The meaning of altered expression of AXIN2 has to be studied in more detail with a larger cohort of patients. In addition, the expression analysis of the
AXIN2 gene may be examined in precancerous
samples such as dysplasic and metaplasic lesions from patients. Note that decreased mRNA and protein expression of AXIN2 was lowered significantly at early stage, but not at stage III and IV. A relationship might exist between the expression of AXIN2 and poor survival, and only cancer patients without AXIN2 alteration could be
alive with the advanced stages of the disease. How the expression of AXIN2 influences the survival of lung cancer patients is a question that warrants further analysis.
In conclusion, our finding with NSCLC tumors corroborates decreased mRNA expression with low protein level in the AXIN2 gene. Our recent published data showed that a frequent deletion region was located in 17q24.3, which is the chromosomal region harboring the AXIN2 gene (Tseng et al., 2005). In addition, the preliminary data indicated that promoter hypermethylation was occurred in ~35% of 20 lung tumor tissues analyzed (data not shown). More studies are needed to analyze whether chromosome deletion or promoter hypermethylation of AXIN2 leads to altered gene expression.
References
Behrens J, Jerchow BA, Wurtele M, Grimm J, Asbrand C, Wirtz R, Kuhl M, Wedlich D, Birchmeier W. 1998. Functional interaction of an axin homolog, conductin, with beta-catenin, APC, and GSK3beta. Science 280:596-599.
Boutros M, Mihaly J, Bouwmeester T, Mlodzik M. 2000. Signaling specificity by Frizzled receptors in Drosophila. Science 288: 1825-1828.
Dong X, Seelan RS, Qian C, Mai M, Liu W. 2001. Genomic structure, chromosome mapping and expression analysis of the human AXIN2 gene. Cytogenet Cell Genet 93:26-8.
Fearon ER. 1997. Human cancer syndromes: clues to the origin and nature of cancer. Science 278:1043-50.
Hart MJ, de los Santos R, Albert IN, Rubinfeld B, Polakis P. 1998. Downregulation of beta-catenin by human Axin and its association with the APC tumor suppressor, beta-catenin and GSK3 beta. Curr Biol 8:573-81.
Hsu HS, Wang YC, Tseng RC, Chang JW, Chen JT, Shih CM, Chen CY. 2004. 5' cytosine-phospho-guanine island methylation is responsible for p14ARF inactivation and inversely correlates with p53 overexpression in resected non-small cell lung cancer. Clin Cancer Res 10:4734-41.
pRb/E2F and Wnt/beta-catenin pathways: E2F1 induces axin2 leading to repression of Wnt signalling and to increased cell death. Exp Cell Res 303:32-46.
Kitagawa M, Hatakeyama S, Shirane M, Matsumoto M, Ishida N, Hattori K, Nakamichi I, Kikuchi A, Nakayama K. 1999. An F-box protein, FWD1, mediates ubiquitin-dependent proteolysis of beta- catenin. Embo J 18:2401-10.
Koch A, Weber N, Waha A, Hartmann W, Denkhaus D, Behrens J, Birchmeier W, von Schweinitz D, Pietsch T. 2004. Mutations and elevated transcriptional activity of conductin (AXIN2) in hepatoblastomas. J Pathol 204:546-54.
Liu W, Dong X, Mai M, Seelan RS, Taniguchi K, Krishnadath KK, Halling KC, Cunningham JM, Boardman LA, Qian C and others. 2000. Mutations in AXIN2 cause colorectal cancer with defective mismatch repair by activating beta-catenin/TCF signalling. Nat Genet 26:146-7.
Lustig B, Jerchow B, Sachs M, Weiler S, Pietsch T, Karsten U, van de Wetering M, Clevers H, Schlag PM, Birchmeier W and others. 2002. Negative feedback loop of Wnt signaling through upregulation of conductin/axin2 in colorectal and liver tumors. Mol Cell Biol 22:1184-93.
Mai M, Qian C, Yokomizo A, Smith DI, Liu W. 1999. Cloning of the human homolog of conductin (AXIN2), a gene mapping to chromosome 17q23-q24. Genomics 55: 341-4.
Minna JD, Sekido Y, Fong K, Gazdar AF. 1997. Cancer. 5 ed: Lippincott: Philadelphia. 849-57 p.
Morin PJ, Sparks AB, Korinek V, Barker N, Clevers H, Vogelstein B, Kinzler KW. 1997. Activation of beta-catenin-Tcf signaling in colon cancer by mutations in beta-catenin or APC. Science 275:1787-90.
Polakis P. 2000. Wnt signaling and cancer. Genes Dev 14:1837-51.
Salahshor S, Woodgett JR. 2005. The links between axin and carcinogenesis. J Clin Pathol 58:225-36.
Taniguchi K, Roberts LR, Aderca IN, Dong X, Qian C, Murphy LM, Nagorney DM, Burgart LJ, Roche PC, Smith DI and others. 2002.
Mutational spectrum of beta-catenin, AXIN1, and AXIN2 in hepatocellular carcinomas and hepatoblastomas. Oncogene 21:4863-71. Tseng RC, Chang JW, Hsien FJ, Chang YH, Hsiao
CF, Chen JT, Chen CY, Jou YS, Wang YC. 2005. Genomewide loss of heterozygosity and its clinical associations in non small cell lung cancer. Int J Cancer 17:17-24.
Wodarz A, Nusse R. 1998. Mechanisms of Wnt signaling in development. Annu Rev Cell Dev Biol 14:59-88.
Yan D, Wiesmann M, Rohan M, Chan V, Jefferson AB, Guo L, Sakamoto D, Caothien RH, Fuller JH, Reinhard C and others. 2001. Elevated expression of axin2 and hnkd mRNA provides evidence that Wnt/beta -catenin signaling is activated in human colon tumors. Proc Natl Acad Sci U S A 98:14973-8.
在非小細胞肺癌中 AXIN2 基因表現之分析
曾若嘉
1曾傑
1,2許瀚水
3王憶卿
1 1國立臺灣師範大學生命科學系 2國立清華大學生物資訊與結構生物研究所 3台北榮民總醫院胸腔外科 (收稿日期:2005.5.2,接受日期:2005.6.2) 摘 要 肺癌在世界上是最普遍的惡性腫瘤之一,並且在包括台灣的工業化國家是癌症死亡的主要 原因;因此,探索哪些基因參與肺癌之形成是很重要的。Wingless (Wnt) 訊號傳遞與細胞增生、 活動及癌化生成是有關連的,在Wnt 訊號傳遞途徑中,Axis inhibition protein 2 (AXIN2) 蛋白的 功能如同骨架蛋白,在β-catenin 蛋白質降解過程中扮演重要角色,推測 AXIN2 基因在此訊號傳 遞途徑中似具有腫瘤抑制功能。為瞭解在非小細胞肺癌中,AXIN2 基因表現是否發生變異,我 們利用反轉錄-聚合酶及免疫組織化學染色法偵測 56 位非小細胞肺癌及非腫瘤配對肺組織中AXIN2 基因之異常。反轉錄-聚合酶分析的結果顯示,45%的病人 AXIN2 基因之 mRNA 表現有降
低的情形。卡方分析發現mRNA 低表現經常發生於腫瘤分期之早期病人,其比例為 54%(P 值 為0.048),達顯著相關;另外,亦發現 mRNA 低表現與年老病人是有相關性的 (P 值為 0.052)。 免疫組織化學染色法分析結果顯示36%的非小細胞肺癌樣本有 AXIN2 蛋白質低表達之情形。卡 方分析發現AXIN2 蛋白質低表達經常發生於良好或中度之腫瘤分化分期 (P 值為 0.023),達顯 著相關。我們的研究結果顯示,非小細胞肺癌中 AXIN2 基因之 mRNA 及蛋白質表現的變異是經 常發生的,而且可能與肺癌早期形成有關。 關鍵詞:AXIN2,非小細胞肺癌