行政院國家科學委員會專題研究計畫成果報告
水稻根部低溫耐性分子機制之探討
Molecular study of the cold-tolerance mechanism in the root
of rice (
Oryza sativa L.)
計畫編號:NSC 90-2313-B-002-284-
執行期限:90 年 08 月 01 日至 91 年 07 月 31 日
主持人:張孟基助理教授 執行機構及單位名稱國立台灣大學農藝學系
共同主持人:盧虎生教授 執行機構及單位名稱國立台灣大學農藝學系
計畫參與人員:楊蕓葦,陳維平執行機構及單位名稱國立台灣大學農藝學系
一、中文摘要 水 稻 經常因遭受非生物性之生理逆 境,如低溫而導致嚴重減產。因此極其重 要的是去瞭解水稻在低溫逆境下之耐冷機 制,尤其是其內部所有基因或蛋白質之表 現,找出與低溫反應相關之基因或蛋白 質。本計畫乃以水稻根部為系統,承續先 前已應用之基因差異顯示法,進一步以 PCR-selected cDNA substraction , cDNA-AFLP 及蛋白質雙向電泳等方法來研 究在低溫下其內部所有基因與蛋白質之表 現。實驗結果顯示,由先前基因差異顯示 法對低溫具耐性(台農 67 號, TNG67) 及對 低溫敏感(台中在來一號, TN1) 之水稻根 部所得之部份 cDNA 片段逕行篩庫而獲致新 增 基 因 , 如 Reversibly glycosylated polypeptide protein (RGP), Dna-J related protein, NADPH-dependent aldose reductase, inorganic pyrophosphatase, Myb-like protein 與 ICT , 將 進 一 步 分 析 其 生 理 功 能 。 在 PCR-selected cDNA substraction 之研究 上 亦 藉 由 反 向 北 方 墨 氏 法 (revrse Northern blot) 初步獲得近 20 個在低溫 下基因表現有差異之 cDNA 片段, 目前正從 事定序之進一步方析。另外在以 cDNA-AFLP 之方法來尋找早期與低溫訊息傳導有關之 基因,初步得到數個在低溫下可被誘導之 cDNA 片段,將繼續就其它不同引子之組合 進行篩選。最後本計畫亦針對水稻根部在 低溫或低溫處理後予以回溫其全部蛋白質 表現群相之差異利用蛋白質雙向電泳加以 分析,目前至少找到 8 個受溫度影響其表 現的蛋白質。綜合言之,本計畫未來將就 上述 cDNA 片段或蛋白質,進一步分析其與 低溫反應之關係,並檢測該基因如何在生 理功能上參與低溫耐性之調控。 關鍵詞:水稻,根,低溫耐性,聚合酉母選擇 性 cDNA 互補反應,cDNA-引子接合長度多型 性, 蛋白質雙向電泳 AbstractAbiotic environmental stresses, such as low temperature frequently result in significant reduction of the potential yield of rice. Therefore, to understand the cold tolerance mechanism in rice, it is essential to evaluate and determine the genes/ or proteins
expression profile in rice under chilling stress, and identify key transcripts or proteins that are associated with this process. To achieve this purpose, follows previous differential
display (DDRT-PCR) approach, several methods including PCR-selected cDNA substraction, cDNA-AFLP and protein 2-D gel analysis have been applied to address this problem. The result showed that with partial cDNA fragments derived from
DDRT-PCR using chilling tolerant (TNG 67) or chilling sensitive (TN1) rice cultivars as probes, several new genes have been found. These are reversibly glycosylated polypeptide protein (RGP), Dna-J related protein,
NADPH-dependent aldose reductase,
inorganic pyrophosphatase, Myb-like protein and ICT. The physiological function of these genes will be further characterized. Also about twenty cDNA fragments that expressed under chilling condition were identified by PCR-selected cDNA substraction and confirmed by reverse Northern blot. These cDNA fragments are now in sequencing. In addition, to look for genes involved in the early stage of cold signal transduction, another approach cDNA-AFLP was carried out in this study. So far a few cDNA fragments were obtained and other combination primer sets will be continuously used in searching for more genes. Finally, a protein 2-D gel analysis was used to reveal protein expression profiles difference in rice roots under chilling or re-warming condition. So far at least eight proteins were identified. In the future, genes or proteins identified above will be further analysis to dissect their relationship with cold tolerance in rice root.
Keywords: Rice, Root, Cold-tolerance, PCR
-selected cDNA substraction, cDNA-AFLP, protein 2-D gel analysis
二、緣由與目的 水稻(Oryza Sativa. L)為亞洲及台灣多數 人賴以維生的重要作物. 但在早春的幼苗 期經常遭受寒害而導致產量受損。因此對 於作物如何獲致低溫耐性之分子機制的瞭 解,一直是植物生理學、作物育種學上所 關心的共同研究課題。其研究結果亦可提 供未來對低溫具耐性之農作物在分子育種 上的考量。要達到此目的,極其重要的是 去研究植物在低溫逆境下其內部所有基因 之表現,找出與低溫反應相關之基因,並 檢測該基因如何在生理功能上參與低溫耐 性之調控. 本系作物生理研究室已初始用 基因差異顯示法,分別從對低溫具耐性(台 農 67 號, TNG67) 及對低溫敏感(台中在來 一號, TN1) 之水稻根部,篩選出超過 300 多個與低溫耐性相關之 cDNA 片段. 但因此 研究所得基因多在低溫逆境之較後期表 現,同時亦非所有與低溫耐性相關之基因 皆會受低溫所誘導,在低溫耐性之分子機 制的研究上並無法涵蓋所有主要基因。是 以在前述既有之基礎下,本計劃擬籍由其 他 PCR-selected cDNA substraction 及 cDNA-AFLP 的方法進一步尋找水稻根部與 早期低溫訊息傳遞相關之基因,並以蛋白 質體學(proteomics) 2D-gel 之分析,希望 盡可能概括水稻根部中與低溫耐性有關之 所有基因。 三、結果與討論 1. 分析由基因差異顯示法所獲得之 cDNA clones: 由於基因差異顯示法之局限,諸多癹現之 cDNA 片段多位於基因 3’ 端之非轉錄區 (UTR) 而無法比對到已知之基因序列。所 以我們以此些部份 cDNA 片段為探針,進行 水稻基因庫之篩選,所得結果如 table 1. 其中 Osic3 解碼 RGP-2 蛋白質。該蛋白質 已知與植物細胞壁及澱粉之形成相關。在 Northern blot 實 驗 中 證 實 該 基 因 之 表 現,在不耐冷之 TN1 水稻中會隨低溫處理 之延長而降低。相反的,在耐冷之 TNG67 水稻中則會增加。該基因之活性是否與低 溫下根部細胞壁之形成及耐冷性相關,則 仍 有 待 進 一 步 證 實 。 另 外 Osic4 解 碼 DnaJ-like 之蛋白質。該類蛋白質主要為
chaperon 而與蛋白質構形相關。再者 Osjc5 解 碼 細 胞 質 soluble inorganic pyrophosphatase. 先前許多實驗已證實植 物 在 低 溫 下 會 誘 導 液 泡 vacuolar inorganic pyrophosphatase 之增加,進而 促使細胞質內游離磷酸、ATP pool 提高, 增 加 sucrose phosphate synthase 等酵素之 活性, 而有 sucrose 或 proline 之累積, 提昇植物耐冷之程度。實驗顯示 Osjc5 基 因之表現在 TN1 及 TNG67 水稻根部,若以 低溫處理皆會增加。但 Osjc5 雖亦解碼 inorganic pyrophosphatase 蛋白質卻存 於 細 胞 質 內 , 與 前 述 液 泡 之 inorganic pyrophosphatase 不同。是以其雖可被低溫 誘導但是否與低溫耐性相關仍有待証明。 最後 Osim3 及 Osjm1 經確認為同一 cDNA 之 clone,為 ICT 基因。目前僅知 ICT 基因 可被乾旱及低溫誘導,功能未定。
2. 以 PCR-selected cDNA substraction 及 cDNA-AFLP 之方法尋找早期低溫訊息 傳遞相關之基因:
本實驗先前由基因差異顯示法所發現之 cDNA clones 主要是低溫或 ABA 處理水稻植 株 24 小時後所得,並無法涵概與早期低溫 訊息傳遞相關之基因。是以在本年度之計 畫中我們以低溫處理 TN1 及 TNG67 水稻 0.5, 1, 及 2 小時後之混合樣本(正常生長溫度 者為對照) 進行上述實驗,結果如 Fig.1 及 Fig.2。目前已成功的獲得一些 cDNA 片段, 正整理分析資料中。並將繼續此實驗尋找 更多相關基因。 3. 以蛋白質雙向電泳分析尋找與低溫耐 性表現相關之蛋白質: 由於水稻根部低溫耐性之表現可能不只是 轉錄層次上基因表現之差異,也可能經由 轉譯或後轉譯蛋白質表現之不同所致。所 以我們優化(optimize) 水稻根部蛋白質萃 取及雙向電泳之條件,對 TN1 及 TNG67 水 稻在低溫處理 1 天或隨後給予 1 天之回溫 處理,其蛋白質表現群相與正常溫度生長 者 之 異 同 進 行 分 析 , 結 果 如 Fig.3 及 Fig.4。目前我們己可初步挑選出 8 個在低 溫下表現量增加而回溫後卻減少之蛋白 質,如 table 2. 另外亦發現在低溫處理下 不耐冷之 TN1 水稻植株除了整体蛋白質表 現量降低外,亦有兩群主要蛋白質大幅減 少。將在後續計畫中深入探討。 四、計畫成果自評 本計畫之最終目標乃在闡明水稻根部低溫 耐性之分子機制,尤其是發現其中進行調 控之特殊基因或蛋白質(specific master regulatory transcripts or proteins) 。 目 前 本 研 究 室 已 順 利 的 建 立 起 PCR-selected cDNA substraction, cDNA-AFLP 及蛋白質雙向電泳的分析方法,初步 的找到一些與低溫表現相關的基因或蛋白 質,正積極進行其核甘酸或胺基酸序列之 分析比對。此與原計畫先前提出之研究內 容與進度相符。未來本實驗室將就上述已 建立之方法持續尋找更多與低溫耐性相關 的基因或蛋白質並探討其在水稻根部低溫 耐性所參與之生理角色。其研究結果亦可 提供未來對低溫具耐性之農作物在分子育 種上的考量。 五、參攷文獻
1. Boyer, J. S. (1982) Plant productivity and environment. Science 218, 443-448. 2. Chen, S. L., Lur, H. S., Yu, S. M., Chen,
H. H., and Chu, C. (2000) Identification of novel genes related to chilling tolerance in rice (Oryza sativa L.)
seedlings by differential display. Journal of A. A. C. 6, 614-633.
3. Chu, C. and Lee, T. M. (1992) Regulation of chilling tolerance in rice seedlings by plant hormones. Korean. J. Crop. Sci. 37, 288-298.
4. Diatchenko, L., Lau, Y. F., Campbell, A. P., Chenchik, A., Moqadam, F., Huang,
B., Lukyanov, S., Lukyanov, K., Gursakaya, N., Sverdlov, E. D., and Siebert, P. D. (1996) Suppression substractive hybridization: a method for generating differentially regulated or tissue-specific cDNA probes and libraries. Biochemistry 93, 6025-6030.
5. Durrant, W. E., Rowland, O., Piedras, P., Hammond-Knosack, K.E., and Jones, J. D. G. (2000) cDNA-AFLP reveals a striking overlap in Reca-specific resistance and wound response gene expression profile. The Plant Cell 12, 963-977.
6. Lee, T. M., Lur, H. S. and Chu, C. (1995) Role of abscisic acid in chilling-tolerant rice (Oryza sativa L.).seedlings. I.
Endogenous abscisic acid levels. Plant Cell Environ 16, 481-490.
7. Sandra, M. J. L., Marco, V., Marijke, K., Ringo, V. W., Ankie, B., Jan, W. K., and Sylvia, D. P. (2002) Glucosylation activity and complex formation of two classes of reversibly glycosylated polypeptides. Plant Physiology 129, 278-289.
8. Thomashow, M. F. (1999) Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant. Physiol. 50:571-599.
9. Wijk, K. J. (2001) Challenges and prospects of plant proteomics. Plant Physiology126, 501-508.
10.Karine, G., Claudette, J., Steven, P. C. G., Magda, P., Hans, D., Joel, V., and Dominique., J. (2001) Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiology 126, 835-848.
Table 1. Sequence analysis of newly identified differential-expressed cDNA clones. Clone IDAccession No. Sequence homology ( ) indicates homology Induced by cDNA length (bp) Osic1 AU089660 AU174915, 475 bps (98%) ABA 319 Osic3 AU089654 Rice reversibly glycosylated
polypeptide protein (RGP2)
LT 228
Osic4 AU089662 Zea Mays DnaJ-related proteinZMDJ1gene LT 422 Osia6 Osjc1 AU089656 AU089671 NADPH-dependent aldose reductase AU056518,503bps(98%) ABA LT 268 349 Osjc5 AU089668 Zea mays soluble inorganic
pyrophosphatase
LT 264
Osjc6 AU089674 Rice Myb-like protein LT 430 Osja2 AU089669 AU075555, 716bps ABA 289 Osim3
(Osjm1)
AU089676 AU089661
ICT mRNA MeJa 349
Osjm4 AU089665 AU172780, 445bps(98%) MeJa 214 Osjm5 AU089675 AU164834, 322bps(95%) MeJa 438
Accession numbers registered in DDBJ, EMBL and GenBank and mRNA sources of abscisic acid (ABA) treated, methyl jasmonate (MeJa) treated, or controls (CK) are indicated. Homology is given as percent matched and length in base pairs (bp) of nucleotide matched. Sequences showing a similarity score over 200 were considered homologous to the cDNA clones were presented. NA: data are not available.
Fig. 1. 以 cDNA-AFLP 之方法來篩選早期低溫訊息傳遞
相關之基因。AFLP (MseI/EcoRI adaptor) system II kit 購 自 Gibco-BRL 公司。圖中 CK 表示樣品來自正常溫度 30oC/25oC 下生長之水稻台農 67 號幼苗根部所合成的 cDNA。 mix 表示經 5℃ 低溫處理 30 分鐘、1 小時 2 小 時之水稻台農 67 號幼苗根部所合成 cDNA。 □處表示 基因表現量有所差異之 cDNA 片段。
Fig. 2. 以 PCR-selected cDNA substraction 方法
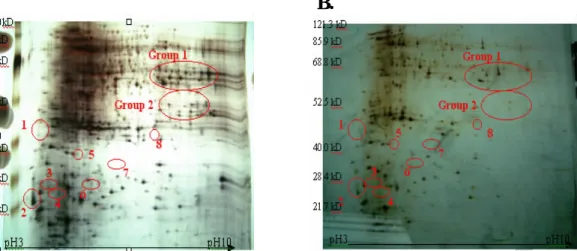
來釣取受低溫誘導或抑制之 TN1 及 TNG67 水稻 幼苗根部基因。1:以 TN1 cDNA (CK)做為 tester,及 TN1 cDNA (ABA 前處理 24 小時再經 過 5℃低溫處理 1 小時、2 小時及 3 小時之混合 樣品)做 driver;所釣取到會被低溫及 ABA 抑制 的基因 2:以 TN1 cDNA (ABA 前處理 24 小時 再經過 5℃低溫處理 1 小時、2 小時及 3 小時之 混合樣品)做 tester,再以 TN1 cDNA (CK) 做為 driver;所釣取到會被低溫及 ABA 誘導的基因 3:以 TNG67 cDNA (CK) 做為 tester,再以 TNG67 cDNA (5℃低溫處理 0.5 小時、1 小時及 2 小時之混合樣品)做 driver;所釣取到 在低溫下會被抑制的基因 4:將 3 的 tester 及 driver 各稀釋 10 倍後做一樣的處理所得到 的結果 5:以 TNG67 cDNA (5℃低溫處理 0.5 小時、1 小時及 2 小時之混合樣品)做 tester, 再以 TNG67 cDNA (CK) 做為 driver;所釣取到在低溫下會被誘導的基因 6:將 5 的 tester 及 driver 各稀釋 10 倍後做一樣的處理所得到的結果。CK 表示正常溫度 30o C/25oC 生 長,M 表示 100 bps DNA marker. A. B. C. Figur e. 3. 將台農 67 號水稻 (TNG67) 三 葉齡幼苗根部萃取之蛋白質, 以 pH 3-10 non-linear IPG 及 15% SDS-PAGE 進行雙 向 電 泳 之 蛋 白 質 表 現 群 相 圖 (proteins expression profile) 。 (A)TNG 67 正常溫度 30oC/25oC 生長。 (B)TNG 67 經低溫處理 24 小時。 (C)TNG 67 經低溫處理 24 小時後以回溫 處理 24 小時。 ○處表示在上述 A.B.C 之條件相較下,表
A. B.
Figur e. 4. 將台中在來 1 號水稻 (TN1) 三葉齡幼苗根部萃取之蛋白質, 以 pH 3-10
non-linear IPG 及 15% SDS-PAGE 進行雙向電泳之蛋白質表現群相圖(proteins expression profile) 。(A) TN 1 正常溫度 30oC/25oC 生長。(B) TN 1 經低溫處理 24 小時。 Table. 2. TNG 67 及 TN 1 水稻三葉齡幼苗於正常、低溫、低溫處理後逕行回溫 不同條件處理下,其根部蛋白質萃取液之雙向電泳分析比較所獲具表現差異性之 蛋白質。 Tr eate r ice cultivar s Pr otein spots TNG 67 CK TNG67 chilling TNG 67 rewarming TN1 CK TN1 chilling 1 - + - - - 2 + ++ ++ + + 3 - + + - - 4 + - + + - 5 + ++ - - - 6 - + - - - 7 - + - - - 8 - + - - - Group 1 + - Group 2 + - * (-)表示此蛋白質不表現或表現量較低 (+)表示蛋白質表現量增加之程度,+符號較多者表現量相對較高。
8