Amyloid-Degrading Ability of Nattokinase from
Bacillus subtilis Natto
R
UEI-L
INH
SU,
†,‡K
UNG-T
AL
EE,
§J
UNG-H
AOW
ANG,
§L
ILYY.-L. L
EE,
† ANDR
ITAP.-Y. C
HEN*
,†,‡Institute of Biological Chemistry, Academia Sinica, Taipei 115, Taiwan, R. O. C., Institute of Biochemical Sciences, National Taiwan University, Taipei 106, Taiwan, R. O. C., and Department of
Biochemical Science and Technology, National Taiwan University, Taipei 106, Taiwan, R. O. C.
More than 20 unrelated proteins can form amyloid fibrils in vivo which are related to various diseases, such as Alzheimer’s disease, prion disease, and systematic amyloidosis. Amyloid fibrils are an ordered protein aggregate with a lamellar cross-β structure. Enhancing amyloid clearance is one of the targets of the therapy of these amyloid-related diseases. Although there is debate on whether the toxicity is due to amyloids or their precursors, research on the degradation of amyloids may help prevent or alleviate these diseases. In this study, we explored the amyloid-degrading ability of nattokinase, a fibrinolytic subtilisin-like serine protease, and determined the optimal conditions for amyloid hydrolysis. This ability is shared by proteinase K and subtilisin Carlsberg, but not by trypsin or plasmin. KEYWORDS: Nattokinase; amyloid; natto; subtilisin NAT; amyloid degradation; fibril; amyloidosis INTRODUCTION
Natto, a fermented food made from boiled soybeans, has been eaten for more than 1000 years in Asia. The fermenta-tion microbe isolated from natto is the Gram-positive endospore-forming bacterium Bacillus subtilis natto (formerly designated Bacillus natto) (1). Nattokinase (formerly desig-nated Subtilisin NAT) (2) is an extracellular enzyme secreted by B. subtilis natto (3) and belongs to the alkaline serine protease family, the catalytic center of which contains three conserved residues, Asp-32, His-64, and Ser-221 (4). It has a molecular mass of 27.7 kDa and an isoelectric point of 8.7 (5). Nattokinase is composed of 275 amino acids, and the gene sequence is homologous to those of other members of the subtilisin family (99.5% homology with subtilisin E, 86% with subtilisin BPN′, and 72% with subtilisin Carlsberg) (6). It not only degrades fibrin in thrombi (7) but also cleaves plasminogen activator inhibitor type I (7, 8). Nattokinase has greater thrombolytic activity than plasmin (5, 7, 9), a natural thrombolytic protease in blood, and increases the production of plasmin from plasminogen due to its action on plasminogen activator. These observations, together with the fact that it can be absorbed across the intestinal tract after oral administration (10, 11) and induce fibrinolysis (11), make nattokinase a potential clot-dissolving agent for the treatment
of cardiovascular disease. Dietary supplementation with natto suppresses the intimal thickening of arteries and leads to the lysis of mural thrombi seen after endothelial injury (12). Other clinically thrombolytic agents, such as urokinase and streptokinase, are costly and unstable in the intestinal tract (13). The use of oral administration of nattokinase in fibrinolytic therapy for thrombosis and the prevention of atherosclerosis is therefore of interest. Nattokinase is cur-rently used as a nutrient supplement to improve circulation in the body (3, 11, 12).
Although much research has been carried out on nattoki-nase, there has been no interest in whether it can degrade amyloids, which are also highly insoluble and protease-resistant. Here, we tested its ability to degrade amyloid fibrils formed from three different proteins and peptides. The first sample was Aβ40 fibrils; amyloid plaque formation is one of the pathological characteristics of Alzheimer’s disease, and Aβ40 is one of the major cleaved products of human amyloid precursor protein and the major component of amyloid plaques. The second sample was insulin fibrils, as repeated insulin injection of diabetes patients can cause injection-localized amyloidosis (14). The third sample was prion peptide fibrils, as prion is the agent responsible for prion diseases. We synthesized the prion peptide correspond-ing to human prion sequence (108-144) to produce amyloid fibrils, as previous observations have shown that it is the most likely segment to form an amyloid structure (15, 16). Human prion protein has either a Met or Val at position 129; we choose the Val-containing sequence, as the resulting amyloid fibrils gave a stronger fluorescence signal with the test agent, thioflavin T (ThT).
* Author to whom correspondence should be addressed: Rita P.-Y. Chen, Institute of Biological Chemistry, Academia Sinica, No 128, Sec 2, Academia Rd, Nankang, Taipei, 115, Taiwan. Tel: +886-2-2785-5696. Fax: +886-2-2788-9759. E-mail: pyc@gate.sinica.edu.tw.
†Academia Sinica.
‡Institute of Biochemical Sciences, National Taiwan University. §
Department of Biochemical Science and Technology, National Taiwan University.
10.1021/jf803072r CCC: $40.75 2009 American Chemical Society Published on Web 12/31/2008
MATERIALS AND METHODS
Production and Purification of Nattokinase. Bacillus
sub-tilis natto was isolated from the commercial product and
maintained on an NB slant at 4 °C (17). Production of nattokinase was performed in a 7-L fermentor (Bioflo 110 Modular Benchtop Fermentor, New Brunswick Scientific, NJ) based on the method described in our previous report (18). The working volume was 5 L of 5% (w/v) soymilk (250 g of soybean powder in 5 L of H2O), the aeration rate 1.0 volume per volume
per minute, the agitation speed 800 rpm, and fermentation was performed at 37°C. All subsequent steps were at 4 °C. After 28 h, the supernatant was recovered by centrifugation at 12000g and concentrated on an Amicon Ultra membrane with a 10 kDa cutoff. The concentrated enzyme solution was applied to a gel filtration column (2.6× 60 cm) packed with 320 mL of HiPrep 26/60 Sephacryl S-100 high resolution gel (Amersham Biosci.) with a mobile phase of 50 mM sodium phosphate and 150 mM NaCl, pH 7.0, and a flow rate of 0.5 mL/min. One milliliter fractions were collected, and those with nattokinase activity and showing a single band on 15% SDS-PAGE were pooled. Nattokinase activity was determined by a chromogenic method using S2251 (H-D-Val-Leu-Lys-pNA; Sigma) as the substrate (19).
Peptide Synthesis. Peptide Aβ40
(DAEFRHDSGYEVH-HQKLVFFAEDVGSNKGAIIGLMVGGVV) and human prion peptide sequence 108-144 with Val at residue 129 (abbreviated as huPrP) (Ac-NMKHMAGAAAAGAVVGGLGGYVLGSAM-SRPIIHFGSD-NH2) were synthesized by the Fmoc-polyamide
method on a PS3 peptide synthesizer (Rainin). The N-terminal end of the huPrP peptide was acetylated and the C-terminal end amidated in order to mimic the configuration in the full-length protein. Preloaded Fmoc-Val-Wang resin (substitution 0.47 mmol/g) was purchased from Anaspec Inc. and used in the synthesis of Aβ40, while Rink Amide AM resin (substitution 0.74 mmol/g) was purchased from Novabiochem and used in the synthesis of huPrP. Fmoc-amino-acid derivatives (0.4 mmol) were coupled to 0.1 mmol of resin using 0.4 mmol of benzotriazole-1-yl-oxy-tris-pyrrolidino-phosphonium hexafluo-rophosphate in dimethylformamide (DMF) containing 4.45% (v/v) of N-methylmorpholine. Acetylation of the N-terminal of huPrP was performed using 0.4 mmol of acetic anhydride
instead of amino-acid derivative in the synthetic procedure. Fmoc cleavage was performed using 20% (v/v) piperidine in DMF. The peptides were cleaved from the resin by stirring at room temperature for 1-2 h with a mixture of 9.4 mL of trifluoroacetic acid, 0.1 mL of triisopropylsilane, 0.25 mL of water, and 0.25 mL of ethanedithiol and precipitated with three volumes of ice-cold methyl t-butyl ether by centrifugation at 2000g for 10 min at 4°C, then the pellet was washed another two times with methyl t-butyl ether and dried under vacuum. The resulting white powder was purified by reverse-phase HPLC using a Vydac C18 column (10 mm × 250 mm) and acetonitrile-water mixtures containing 0.1% trifluoroacetic acid (v/v). The final products were analyzed by matrix-assisted laser desorption ionization (MALDI) mass spectrometer. Fractions containing the desired product were lyophilized and stored at -20 °C.
Preparation of Amyloid Fibrils. For Aβ40 fibril formation,
a 500 µM Aβ40 stock solution was prepared in 75% trifluoro-ethanol. The peptide concentration was quantified by the absorbance at 275 nm. To form fibrils, the stock solution was diluted to 25 µM in 20 mM sodium phosphate buffer, 150 mM KCl, pH 7, and incubated at 25°C for about two weeks (20). The huPrP fibrils were prepared by dissolving the huPrP peptide in 20 mM NaOAc, 140 mM NaCl, pH 3.7 to a final concentra-tion of 50 µM and incubating the soluconcentra-tion at 25°C for about one week (15, 16). To obtain insulin fibrils, bovine insulin (Sigma) was dissolved in dilute HCl (pH 2.1) at a concentration of 2.5 mg/mL and incubated at 60°C for about two days (21, 22). Fibril formation was monitored by the appearance of negative ellipticity at 218 nm using circular dichroism spectroscopy or fluorescence emission on binding ThT using fluorescence spectroscopy.
Circular Dichroism (CD) Spectroscopy. The sample was
placed in a 1-mm quartz cell and the CD spectrum between 200 and 250 nm recorded on a J-715 CD spectrometer (JASCO, Japan). The bandwidth was set to 2 nm and the step resolution was 0.1 nm. Two scans were averaged for each sample.
Thioflavin T Binding Assay. ThT binding assay measures
the fluorescence intensity change in ThT on binding to amyloid fibrils. This method has been extensively used for identifying the presence of amyloid fibrils. A stock solution of 5 mM ThT (Sigma) was prepared by dissolving 2 mg of the dye in 1.25 mL of 140 mM NaCl, 100 mM phosphate buffer, pH 8.5, and passing the solution through a 0.22 µm Millipore filter. A fresh working solution was prepared by adjusting the final dye concentration to 200 µM. A 30 µL aliquot of the sample was mixed with 30 µL of 200 µM ThT dye solution for 1 min at room temperature, then the fluorescence emission between 460 and 600 nm was measured in a 3-mm path-length rectangular cuvette on a FP-750 spectrofluorometer (JASCO, Japan) with excitation at 442 nm.
Protease Solutions. Stock solutions of all proteases (all
except nattokinase from Sigma) used in this study were prepared at a concentration of 49 µM and quantified by the Bradford assay (Bio-Rad). The protease solutions were prepared in different buffers according to the manufacturer’s instructions; nattokinase and subtilisin Carlsberg were dissolved in 50 mM phosphate buffer containing 150 mM NaCl, pH 7, proteinase K was dissolved in 10 mM Tris-HCl buffer, pH 7, trypsin was dissolved in 1 mM HCl, and human plasmin was dissolved in distilled water.
Degradation of Different Kinds of Amyloid Fibrils by Nattokinase. The prepared Aβ40, huPrP, and insulin fibrils were
collected by centrifugation at 14000g for 20 min at room
Figure 1. 15 % SDS-PAGE of the purified nattokinase, indicated by the arrow.
temperature and suspended in 50 mM phosphate buffer contain-ing 150 mM NaCl, pH 7. In order to obtain more homogeneous fibrils, the fibril solutions were incubated in an ultrasonic bath for 10 min before use. For the enzymatic reaction, 1 µL of nattokinase solution was mixed with 179 µL of each kind of fibril solution (final nattokinase concentration: 0.27 µM) and the mixture incubated at 40 °C for 1 h, then fibril digestion was examined by CD spectroscopy and the ThT binding assay.
Effects of Temperature and pH on the Amyloid-Degrading Activity of Nattokinase. In this experiment, only insulin fibrils
were used as the substrate for the enzymatic reaction, as a larger amount of fibrils was needed (insulin is available commercially and fibril formation takes only 2 days). The effect of temperature on nattokinase activity was measured by digestion at various temperatures from 30 to 60 °C at pH 7, degradation being monitored using the ThT binding assay. To examine the effect of pH on nattokinase activity, insulin fibrils were suspended in the following buffers: 50 mM NaOAc and 150 mM NaCl (pH 5), 50 mM phosphate and 150 mM NaCl (pH 6 and 7), 50 mM Tris-HCl and 150 mM NaCl (pH 8 and 9), and 50 mM glycine-NaOH and 150 mM NaCl (pH 10). The reaction was carried out at 40 °C. Nattokinase activity was measured by CD spectroscopy, as the ThT binding assay is pH-sensitive. The nattokinase concentration used was 0.27 µM.
Comparison of the Aβ Fibril-Degrading Activity of Dif-ferent Proteases. The enzymatic reaction was carried out at
37°C and pH 7 for 1 h using nattokinase, proteinase K, subtilisin Carlsberg, trypsin, or plasmin. The reaction mixture comprised 1 µL of protease stock (49 µM) and 179 µL of Aβ fibril solution (final protease concentration: 0.27 µM), and 30 µL of the mixture was removed every 15 min for testing in the ThT binding assay.
Measurement of Rate of Aβ Fibril Degradation by Nattokinase. The Aβ peptide concentration in the fibril form
was determined by subtracting the nonaggregated Aβ peptide concentration in the supernatant fraction after centrifugation from the overall Aβ concentration. The fibrils were suspended in 50 mM phosphate buffer containing 150 mM NaCl, pH 7.4. Nattokinase was added into the fibril solution at different enzyme concentrations and the mixture was incubated at 37°C. After different incubation times, the amount of remaining fibril was measured by the ThT binding assay. The rate of fibril degrada-tion was calculated from the time course of fibril degradadegrada-tion.
RESULTS
Degradation of Amyloid Fibrils by Nattokinase.
Nattoki-nase was purified from liquid culture by gel filtration chroma-tography. Fractions containing nattokinase were identified using
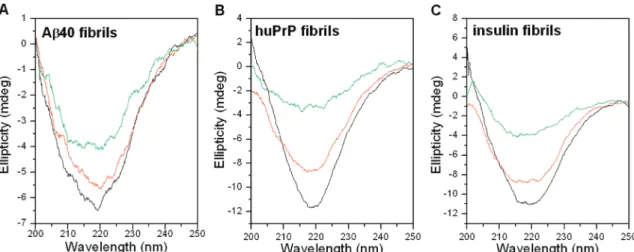
Figure 2. CD spectra of Aβ40 fibrils (A), huPrP fibrils (B), and insulin fibrils (C) before and after digestion by nattokinase at 40°C, pH 7. The spectrum immediately after enzyme addition is shown by the black line. The spectra of samples digested for 1 or 48 h are shown by the red or green lines, respectively.
Figure 3. Fluorescence emission spectra of Aβ40 fibrils (A), huPrP fibrils (B), and insulin fibrils (C) before and after digestion by nattokinase at 40°C, pH 7, using ThT binding assay. The spectrum immediately after enzyme addition is shown by the black line. The spectra of samples digested for 1 or 48 h are shown by the red or green lines, respectively.
the activity assay, then were tested for the presence of a 27.7 kDa band on SDS-PAGE. The band was characterized by N-terminal sequence analysis. Those fractions showing a pure protein were pooled and lyophilized (Figure 1).
To determine whether nattokinase has amyloid-degrading activity, Aβ40 fibrils formed from synthetic Aβ40 peptide, huPrP fibrils formed from synthetic human prion peptide (containing human prion protein amino acids 108-144 with valine at residue 129), and insulin fibrils formed from purified bovine insulin were used. At 40 °C and pH 7, nattokinase degraded all three fibrils, as shown by the decrease in the amplitude of the negative ellipticity at 218 nm in the CD spectra after digestion, indicating loss of β-sheet structure (Figure 2). Similar results were obtained using the ThT binding assay, in which the decrease in ThT fluorescence intensity at 487 nm indicated loss of amyloid structure after nattokinase digestion (Figure 3). These results demonstrated that nattokinase was able to degrade all three different amyloid fibrils.
Optimization of Amyloid Digestion by Nattokinase. The
fibrinolytic activity of nattokinase is known to be stable at neutral and alkaline pH (7-12), but labile below pH 5 (3). To examine the effect of pH on the amyloid-degrading activity of nattokinase, we carried out proteolytic digestion of insulin fibrils over a broad range of pH values (pH 5-10) at 40°C. As shown in Figure 4, the activity increased gradually with increasing pH, maximal activity being seen at pH 10 in the range tested. At pH 5, nattokinase did not show any activity.
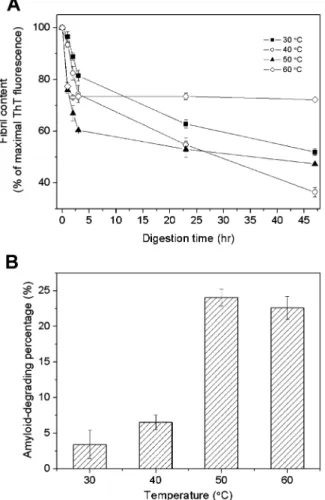
In addition, it has been reported that the fibrinolytic activity of nattokinase is gradually lost above 60°C (3). We therefore tested the amyloid-degrading ability of nattokinase at different temperatures and pH 7. As shown in Figure 5, at 60°C, insulin fibrils were degraded rapidly in the first few minutes, but then
Figure 4. Degradation of insulin fibrils by nattokinase at different pHs. The digestion was carried out at 40°C and monitored by CD spectroscopy. (A) Time-course of degradation monitored by measuring the remaining amyloid fibrils. (B) Percentage amyloid degradation at different pHs calculated using the data for one hour of digestion. 100% was defined as the CD ellipticity at 218 nm of the sample before treatment. The values are the mean ( SEM for the results of two experiments.
Figure 5. Degradation of insulin fibrils by nattokinase at different temperatures between 30 and 60°C. The digestion was carried out in phosphate buffer, pH 7, and monitored using the ThT binding assay. (A) Time-course of degradation monitored by measuring the remaining amyloid fibrils. (B) Percentage amyloid degradation at different temperatures calculated using the data for one hour of digestion. 100% was defined as the fluorescence intensity at 487 nm of the sample before treatment. The values are the mean ( SEM for the results of two experiments.
Figure 6. Comparison of Aβ40 fibril degradation by proteinase K, nattokinase, subtilisin Carlsberg, trypsin, and plasmin. The digestion was carried out at 37°C and pH 7 and monitored by the ThT binding assay every 15 min. 100% was defined as the fluorescence intensity at 487 nm of the control before incubation.
digestion stopped, probably due to the inactivation or self-digestion of nattokinase. At 50°C, rapid initial digestion was seen, followed by slow digestion, while, at 40 °C, the initial release was less rapid, but digestion continued and was more complete than in the other cases. The percentage of amyloid degradation measured at 40°C and pH 7 in the ThT binding assay (Figure 5B) is lower than that measured by CD spectroscopy under the same condition (Figure 4B). We noted that ThT fluorescence intensity was higher in the insulin fibrils incubated at 40 °C (data not shown). Since the maximal fluorescence intensity was measured before incubation, these inconsistent data might result from the underestimation of the maximal fluorescence intensity at 40°C.
Comparison of Nattokinase with Other Proteases.
Nat-tokinase and several other serine proteases were tested for their ability to digest Aβ fibrils at 37 °C and pH 7. The proteases used were proteinase K (a keratinolytic enzyme and subtilisin-like serine protease used in research on prion diseases), subtilisin Carlsberg, trypsin, and plasmin (a fibrinolytic serine protease). The same protease concentration (0.27 µM) was used.
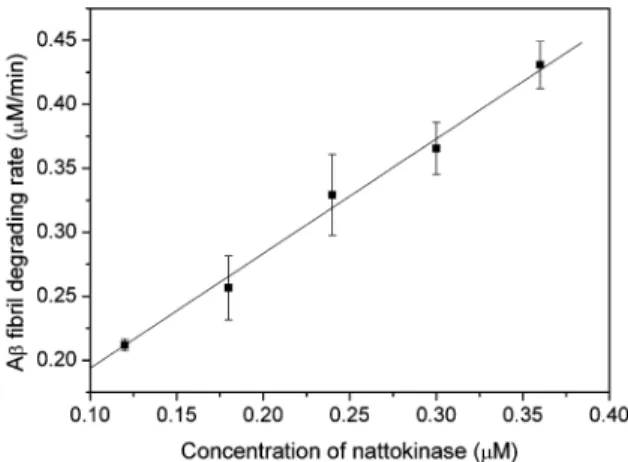
As shown in Figure 6, at 37°C and pH 7, proteinase K and subtilisin Carlsberg degraded Aβ fibrils at the highest rates. The hydrolysis rate was slower for nattokinase and much slower for trypsin, while plasmin was ineffective. Although the digestion was not performed at the optimal temperatures for proteinase K, subtilisin Carlsberg, or nattokinase in order to mimic body temperature, the amyloid-degrading activities of these proteases were still much greater than those of trypsin and plasmin. Tucker et al. (23) have shown that plasmin can degrade Aβ monomer and that it can also degrade aggregated Aβ, but the rate (0.003 s-1) is 20 times lower than that for degrading aggregated fibrin (0.064 s-1) and 160 times lower than that for degrading Aβ monomer (0.48 s-1). In our study, under the same reaction conditions, nattokinase was more efficient than plasmin in cleaving amyloid. Here, the rate of Aβ fibril degradation by nattokinase was 0.015 s-1at 37 °C and pH 7.4 (Figure 7).
DISCUSSION
In this study, we used three kinds of in vitro prepared amyloid fibrils, Aβ40 amyloid fibrils, insulin amyloid fibrils, and huPrP (consisting prion sequence 108-144) amyloid fibrils, to test the amyloid-degrading ability of nattokinase. Aβ40 and huPrP peptide have a random coil structure when first dissolved in a
buffer with a physiological salt concentration, but, after a few days, undergo a coil-to-β structure conversion. The native structure of insulin contains many R-helices, but an R-to-β structural conversion occurs when it is dissolved at acidic conditions and heated at 60°C. The structural conversion can be confirmed by the appearance of negative ellipticity at 218 nm in the CD spectrum, fibril morphology under electron microscopy, and fluorescence emission at 487 nm in the ThT binding assay. The characteristic cross-β structure can be monitored by CD spectroscopy. The ThT binding assay is the most sensitive and widely used method for the detection and quantification of amyloid fibrils. It has been proposed that ThT binds with its long axis parallel to the axis of the amyloid fibrils (24) and that the side chains of the amino acid residues restrict the rotation of ThT, so the fluorescence quantum yield of ThT increases (21). After treatment of the three different amyloid fibrils with nattokinase, the amyloid signal was decreased (Figures 1 and 2), indicating the general amyloid-degrading ability of nattokinase.
This amyloid-degrading ability of nattokinase suggests that it may be useful in the treatment of amyloid-related diseases. The feather-degrading enzyme keratinase (Versazyme), pro-duced by Bacillus licheniformis, can degrade preheated prion (25) and is useful for inactivating prion in meat and bone meal (26) and for decontaminating medical instruments (27), but cannot be taken orally. Many proteins and peptides, such as immunoglobin light chain, transthyretin, β2-microglobulin, serum amyloid A protein, Aβ peptides, and insulin, can form amyloid deposits in the body (28). Aβ peptides can move from the brain into the blood with the aid of low density lipoprotein receptor-related protein and p-glycoprotein (29). In addition, prion disease can be transmitted through blood transfusion (30), suggesting the existence of prion in the blood. Amyloid fibrils are insoluble and not easily digested by proteases. The discovery of an enzyme which can be safely taken orally and can degrade amyloid fibrils could be very useful in the therapy of amyloid-related diseases. Nattokinase not only dissolved blood clots (9) but also degraded amyloid fibrils. Our amyloid-degrading studies demonstrated that it is active at neutral pH and body temperature. Previous results in rats, dogs, and humans have suggested that nattokinase can enter the circulation when taken orally (11, 12), so it has the potential to clear amyloid deposits in various parts of the body. Moreover, prion diseases can be transmitted by the use of contaminated surgical instruments. Nattokinase can tolerate a temperature of 50°C and functions even better under basic conditions, such as pH 10, suggesting that it might be useful in instrument decontamination.
Further animal studies are needed in order to evaluate the therapeutic potential of nattokinase. Various amyloid fibrils can be prepared in vitro and injected into mice. Nattokinase can be given by directly feeding the mice with natto or nattokinase or by injecting nattokinase solution into the body. The amyloid-degrading ability can be tested after an incubation period by detecting the remaining amyloid. Moreover, since natto has been ingested by humans for a long time, it would be worthwhile to carry out an epidemiological study on the rate of occurrence of various amyloid-related diseases in a population regularly consuming natto.
ABBREVIATIONS USED
huPrP, human prion peptide (sequence108-144 with Val at residue 129); Aβ40, amyloid β peptide (residues 1 to 40).
Figure 7. Nattokinase concentration dependence of Aβ fibril degradation. Aβ fibril (44 µM) was incubated with nattokinase (final enzyme concentra-tion of 0.12, 0.18, 0.24, 0.30, or 0.36 µM) at 37°C in phosphate buffer, pH 7.4. The values are the mean ( SEM for the results of two separate experiments.
LITERATURE CITED
(1) Ueda, S. Industrial applications of B. subtilis: utilization of soybean as natto, a traditional Japanese food. In Bacillus subtilis: molecular biology and industrial applications; Maruo, B., Yoshikawa, H., Eds.; Elsevier Science B. V.: Amsterdam, the Netherlands, 1989; pp 143-161.
(2) Peng, Y.; Yang, X.; Zhang, Y. Microbial fibrinolytic enzymes: an overview of source, production, properties, and thrombolytic activity in vivo. Appl. Microbiol. Biotechnol. 2005, 69, 126–132. (3) Sumi, H.; Hamada, H.; Tsushima, H.; Mihara, H.; Muraki, H. A novel fibrinolytic enzyme (nattokinase) in the vegetable cheese Natto; a typical and popular soybean food in the Japanese diet. Experientia 1987, 43, 1110–1111.
(4) Kamata, H.; Yamagata, Y.; Nakamura, T.; Nakajima, T.; Oda, K.; Murao, S.; Ichishima, E. Characterization of the complex between R2-macroglobulin and a serine proteinase from Bacillus natto. Agri. Biol. Chem. 1989, 53, 2695–2702.
(5) Fujita, M.; Nomura, K.; Hong, K.; Ito, Y.; Asada, A.; Nishimuro, S. Purification and characterization of a strong fibrinolytic enzyme (nattokinase) in the vegetable cheese natto, a popular soybean fermented food in Japan. Biochem. Biophys. Res. Commun. 1993, 197, 1340–1347.
(6) Nakamura, T.; Yamagata, Y.; Ichishima, E. Nucleotide sequence of the subtilisin NAT gene, aprN, of Bacillus subtilis (natto). Biosci., Biotechnol., Biochem. 1992, 56, 1869–1871.
(7) Fujita, M.; Hong, K.; Ito, Y.; Fujii, R.; Kariya, K.; Nishimuro, S. Thrombolytic effect of nattokinase on a chemically induced thrombosis model in rat. Biol. Pharm. Bull. 1995, 18, 1387–1391. (8) Urano, T.; Ihara, H.; Umemura, K.; Suzuki, Y.; Oike, M.; Akita, S.; Tsukamoto, Y.; Suzuki, I.; Takada, A. The profibrinolytic enzyme subtilisin NAT purified from Bacillus subtilis cleaves and inactivates plasminogen activator inhibitor type 1. J. Biol. Chem. 2001, 276, 24690–24696.
(9) Chang, C. T.; Fan, M. H.; Kuo, F. C.; Sung, H. Y. Potent fibrinolytic enzyme from a mutant of Bacillus subtilis IMR-NK1. J. Agric. Food Chem. 2000, 48, 3210–3216.
(10) Fujita, M.; Hong, K.; Ito, Y.; Misawa, S.; Takeuchi, N.; Kariya, K.; Nishimuro, S. Transport of nattokinase across the rat intestinal tract. Biol. Pharm. Bull. 1995, 18, 1194–1196.
(11) Sumi, H.; Hamada, H.; Nakanishi, K.; Hiratani, H. Enhancement of the fibrinolytic activity in plasma by oral administration of nattokinase. Acta Haematol. 1990, 84, 139–143.
(12) Suzuki, Y.; Kondo, K.; Matsumoto, Y.; Zhao, B. Q.; Otsuguro, K.; Maeda, T.; Tsukamoto, Y.; Urano, T.; Umemura, K. Dietary supplementation of fermented soybean, natto, suppresses intimal thickening and modulates the lysis of mural thrombi after endothelial injury in rat femoral artery. Life Sci. 2003, 73, 1289– 1298.
(13) Banerjee, A.; Chisti, Y.; Banerjee, U. C. Streptokinase - a clinically useful thrombolytic agent. Biotechnol. AdV. 2004, 22, 287–307. (14) Uversky, V. N.; Fink, A. L. Conformational constraints for amyloid fibrillation: the importance of being unfolded. Biochim. Biophys. Acta 2004, 1698, 131–153.
(15) Chen, P. Y.; Lin, C. C.; Chang, Y. T.; Lin, S. C.; Chan, S. I. One O-linked sugar can affect the coil-to-β structural transition of the prion peptide. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 12633– 12638.
(16) Zhang, H.; Kaneko, K.; Nguyen, J. T.; Livshits, T. L.; Baldwin, M. A.; Cohen, F. E.; James, T. L.; Prusiner, S. B. Conformational transformations in peptides containing two putative R-helices of the prion protein. J. Mol. Biol. 1995, 250, 514–526.
(17) Kuo, L. C.; Cheng, W. Y.; Wu, R. Y.; Huang, C. J.; Lee, K. T. Hydrolysis of black soybean isoflavone glycosides by Bacillus subtilis natto. Appl. Microbiol. Biotechnol. 2006, 73, 314–320. (18) Kuo, L. C.; Lee, K. T. Cloning, expression, and characterization of two β-glucosidase from isoflavone glucoside-hydrolyzing Bacillus subtilis natto. J. Agric. Food Chem. 2008, 56, 119–125. (19) Friberger, P.; Knos, M.; Gustavsson, S.; Aurell, L.; Claeson, G. Methods for determination of plasmin, antiplasmin and plasmi-nogen by means of substrate S-2251. Haemostasis 1978, 7, 138– 145.
(20) Chang, E. S.; Liao, T. Y.; Lim, T. S.; Fann, W.; Chen, R. P. A new amyloid-like β-aggregate with most of the amyloid charac-teristics except fibril morphology. J. Mol. Biol. 2008, 10.1016/ j.jmb.2008.11.009.
(21) Groenning, M.; Norrman, M.; Flink, J. M.; van de Weert, M.; Bukrinsky, J. T.; Schluckebier, G.; Frokjaer, S. Binding mode of Thioflavin T in insulin amyloid fibrils. J. Struct. Biol. 2007, 159, 483–497.
(22) Nielsen, L.; Khurana, R.; Coats, A.; Frokjaer, S.; Brange, J.; Vyas, S.; Uversky, V. N.; Fink, A. L. Effect of environmental factors on the kinetics of insulin fibril formation: elucidation of the molecular mechanism. Biochemistry 2001, 40, 6036–6046. (23) Tucker, H. M.; Kihiko, M.; Caldwell, J. N.; Wright, S.;
Kawara-bayashi, T.; Price, D.; Walker, D.; Scheff, S.; McGillis, J. P.; Rydel, R. E.; Estus, S. The plasmin system is induced by and degrades amyloid-beta aggregates. J. Neurosci. 2000, 20, 3937– 3946.
(24) Krebs, M. R. H.; Bromley, E. H. C.; Donald, A. M. The binding of thioflavin-T to amyloid fibrils: localisation and implications. J. Struct. Biol. 2005, 149, 30–37.
(25) Langeveld, J. P. M.; Wang, J. J.; Van de Wiel, D. F. M.; Shih, G. C.; Garssen, G. J.; Bossers, A.; Shih, J. C. H. Enzymatic degradation of prion protein in brain stem from infected cattle and sheep. J. Infect. Dis. 2003, 188, 1782–1789.
(26) Coll, B. A.; Garcia, R. A.; Marmer, W. N. Diffusion of protease into meat & bone meal for solubility improvement and potential inactivation of the BSE prion. PLoS ONE 2007, 2, e245. (27) Gupta, R.; Ramnani, P. Microbial keratinases and their prospective
applications: an overview. Appl. Microbiol. Biotechnol. 2006, 70, 21–33.
(28) Chiti, F.; Dobson, C. M. Protein misfolding, functional amyloid, and human disease. Annu. ReV. Biochem. 2006, 75, 333–366. (29) Wang, Y. J.; Zhou, H. D.; Zhou, X. F. Clearance of
amyloid-beta in Alzheimer’s disease: progress, problems and perspectives. Drug DiscoVery Today 2006, 11, 931–938.
(30) Hunter, N.; Foster, J.; Chong, A.; McCutcheon, S.; Parnham, D.; Eaton, S.; MacKenzie, C.; Houston, F. Transmission of prion diseases by blood transfusion. J. Gen. Virol. 2002, 83, 2897–2905. Received for review October 2, 2008. Revised manuscript received November 23, 2008. Accepted November 23, 2008.