行政院國家科學委員會專題研究計畫 成果報告
間質幹細胞與椎間盤本核細胞共同培養條件最佳化之研
究–對脊椎椎間盤再生之現行細胞移植療法改進之探討
研究成果報告(精簡版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2314-B-002-148- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學醫學院骨科 計 畫 主 持 人 : 楊曙華 共 同 主 持 人 : 王廷明、施庭芳、吳長晉 計畫參與人員: 學士級-專任助理:孫元蕙 報 告 附 件 : 國外研究心得報告 出席國際會議研究心得報告及發表論文 處 理 方 式 : 本計畫可公開查詢中 華 民 國 96 年 08 月 10 日
INTRODUCTION
The intervertebral disc (IVD) has a composite structure comprised of a nucleus pulposus (NP) core, a multilayered lamina of annulus fibrosus (AF), and cartilaginous end plates. Degeneration of the IVD and associated spinal disorders are a leading cause of morbidity resulting in substantial pain and increased health cost [1]. Some pathophysiological evidences indicate that IVD degeneration originates from the NP [2,3]. The most notable changes in the degenerative NP are a decrease in proteoglycan content and synthesis of collagen II as well as an increase in denaturation of collagen II and synthesis of collagen I [4]. As the NP loses its original osmotic properties and becomes more fibrotic, the IVD is not able to transmit intervertebral force optimally and various degenerative processes will subsequently happen in the IVD.
Regeneration of the NP tissues in the early stages of degeneration can theoretically retard or even reverse the degenerative process and possibly restore a healthy IVD. Since the approval of autologous disc chondrocyte transplantation (ADCT) in Germany in 1997, cell-based therapeutics has shown potential in the treatment of IVD degeneration [5]. The most clinically applicable cell source for disc cell transplantation would be the NP tissue obtained from surgeries for herniation of NP and degenerative disc disease. Cells harvested from degenerative IVD tissues are not completely healthy and have an altered phenotype, showing increased senescence [6], increased expression of both catabolic cytokines and degenerative enzymes [7], and decreased expression of matrix components [8,9]. Use of autologous NP cells from adjacent healthy IVDs is not clinically practical and the harvest of NP tissues would induce degeneration of the donor IVD [10].
Upregulating biologic and metabolic viability of NP cells and differentiating mesenchymal stem cells (MSCs) into NP-like cells are both potential alternative methods to achieve highly viable cells to facilitate regeneration of NP tissues. Growth factors such as fibroblast growth factor (FGF), endothelial growth factor (EGF), and transforming growth factor β(TGFβ) have been shown to enhance synthesis of proteoglycans by NP cells in vitro [11]. It has been demonstrated that MSCs could secrete a variety of growth factors and cytokines to upregulate the biologic and metabolic viability of IVD cells in co-culture systems [12]. On the other hand, MSCs have been induced into IVD-like cells through TGFβ-mediated differentiation [13,14]. However, studies have shown that a more effective proliferation and chondrogenesis may be obtained with proper combinations of growth factors and MSCs [15,16]. Moreover, a continuous delivery of
growth factors may be necessary to achieve maximal stimulatory influences. It has been demonstrated that human NP cells could manifest expression of TGFβ1 [17] as well as some other growth factors that could facilitate differentiation of MSCs into NP-like cells. When MSCs and NP cells are located in a closed environment such as the interior of the IVD or co-cultured in vitro, it can be expected that both cell types could secrete cytokines into the environment and achieve bidirectional influences in a paracrine fashion. The current study started from demonstrating separate expression profiles of cytokines in human NP cells and MSCs, and then a non-contact co-culture system was employed to examine the effects of secreted cytokines on cell proliferation and expression of key matrix components in MSCs and NP cells.
Materials and Methods
Sources and Cultivation of Human NP cells and MSCs
Under the regulation of the Research Ethical Committee, National Taiwan University Hospital, human NP tissues were obtained aseptically from a 23-year-old male patient who underwent a surgery for lumbar disc herniation. Human NP cells were isolated by incubating minced NP tissues in DMEM/F-12 medium (Sigma Co.; St. Louis, MO) containing 0.2% collagenase (Sigma) at 37˚C overnight.Theisolated human NP cells were re-suspended in phosphate-buffered saline (pH 7.4), washed, and their viability was determined using trypan blue dye exclusion. Human NP cells were expanded as monolayers in tissue flasks in a 5% CO2 atmosphereat37˚C in DMEM/F-12 medium
containing 10% fetal bovine serum (Biological Industries Ltd.; Kibbutz Beit Haemek, Isreal), 50 units/mL penicillin, 50 μg/mL streptomycin, 100 μg/mL neomycin (Sigma), and 25 μg/mL L-ascorbic acid (Sigma). The appearance of the monolayer cultured NP cells was observed regularly to monitor if any fibroblastic transformation occurred. Commercially available PoieticsTM human mesenchymal stem cells (Cambrex; East Rutherford, NJ) were expanded as monolayers in tissue culture flasks in a 5% CO2
atmosphereat37˚C in MSC medium (Cambrex).
RNA Expression Profiles of Cytokines in MSCs and NP cells
In order to understand the profiles of cytokine mRNA expression in human MSCs and NP cells, 1x106 monolayer-expanded cells were utilized for the following study. Extraction of total RNA was performed by using Qiagen RNeasy mini kit (Qiagen; Chatsworth, CA).
During the extraction process, RNA was treated with RNase-free DNase (Qiagen) to eliminate the influence of DNA in the PCR. An aliquot of RNA extract was used for RNA content determination, which was measured by absorbance at 260 nm measured with a DU 7500 spectrophotometer (Beckman Instrument Inc.; Fullerton, CA). Complement DNA was synthesized from extracted RNA with SuperScript III First-Strand Synthesis System for RT-PCR, oligo-(dT) primer, and dNTP mix (Invitrogen Corporation; Carlsbad, CA) at 50˚C for 50 min. Complement DNA synthesized from 10 ng RNA was used for each PCR primer. PCR was performed on the ABI PRISM 7700 Sequence Detection System by using random hexamers from the TaqMan Reverse Transcription Reagents and RT Reaction Mix (Applied Biosystems; Foster City, CA) to reverse transcribe the RNA, and TaqMan Universal PCR Master Mix and Assays-on-Demand Gene Expression probes (Applied Biosystems) for the PCR step. Target genes examined included glyceraldehydes-3-phosphate dehydrogenase (GAPDH), TGFß1, interleukin-1ß (IL1ß), bone morphogenetic protein (BMP) -2,4,6,7, insulin-like growth factor-1 (IGF1), and EGF. PCR products were analyzed by agarose gel electrophoresis.
Co-culture of MSCs and NP cells
Six-well culture plates (Orange Scientific, Braine-l'Alleud, Belgium) and 3 cm diameter culture inserts (Millipore; Carrigtwohill, Ireland) were used for co-culture of MSCs and NP cells. The culture insert used in this study consisted of a polyethylene terephthalate track-etched membranewith 0.4 μm poresatthe bottom,which prevented the exchange of cellular components. The initial co-culture preparations of MSCs and NP cells are summarized in Table 1 and Table 2. The cells on the insert were used for evaluating cell proliferation and mRNA expression after 72 hours of co-culture.
The growth medium used for the co-culture experiment was DMEM/F-12 medium containing 10% fetal bovine serum (Biological Industries Ltd.), 50 units/mL penicillin, 50 μg/mL streptomycin, 100 μg/mL neomycin (Sigma),and 25 μg/mL L-ascobic acid (Sigma). Evaluations of final cell numbers and mRNA expression were performed in six repeats in each experimental group.
Measurement of Cell Proliferation
Cell proliferation was determined by WST-8 assay using Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technology; Gaitherburg, MD) [16] with some modifications. 10%
working solution was made by mixing 1 volume of original CCK-8 solution with 9 volumes of DMEM/F-12 medium. After 72 hours of incubation, the culture inserts were transferred to new 6-well culture plates containing 2 mL of CCK-8 working solution in each well. They were incubated at 37˚C in ahumid atmospherecontaining 5% CO2for 2
hours. Then 100 μL of reacted solution was transferred to a 96-well microplate. Absorbance at 450 nm was measured using a VERSAmax microplate absorbance reader (Molecular Devices Corp.; Sunnyvale, CA). Cell counts were determined with a calibration curve of the tested cell type.
Quantatitive RT-PCR
The method for extraction of total RNA from MSCs or NP cells on the culture inserts was the same as above. Real-time PCR was performed on the ABI PRISM 7700 Sequence Detection System by using random hexamers from the TaqMan Reverse Transcription Reagents and RT Reaction Mix (Applied Biosystems) to reverse transcribe the RNA, and TaqMan Universal PCR Master Mix and Assays-on-Demand Gene Expression probes (Applied Biosystems) for the PCR step. The relative standard curve method (Applied Biosystems) was to calculate the amplication difference among untreated and co-cultured MSCs and NP cells. Target genes examined included GAPDH, collagen I (col1a1), collagen II (col2a1), aggrecan, Fas-associated death domain protein (FADD), and SOX-9. The relative expression of each target gene was calculated using the 2-△△CTmethod [17].
The mRNA expression of each target gene was normalized to the housekeeping gene GAPDH. △CT values of each target gene were calculated by subtracting the value of
cycle threshold (CT) for GAPDH from the CTvalue of each target gene. The △CTvalues
were further normalized to control samples (△△CT), which were MSCs and NP cells
cultured alone in all cases except the expression of collagen II in MSCs. Because RNA expression of collagen II was not detected in untreated MSCs, the △CT values of
collagen II expression in co-cultured MSCs were normalized to TGFβ1-treated MSCs. Finally the 2-△△CT values were obtained and showed the relative expression of target
genes in untreated, co-cultured and TGFβ1-treated cells.
Statistical Analysis
Statisticalsignificancewasdetermined using Student’st test, in which p < 0.05 was
RESULTS
Expression Profiles of Cytokines in MSCs and NP cells
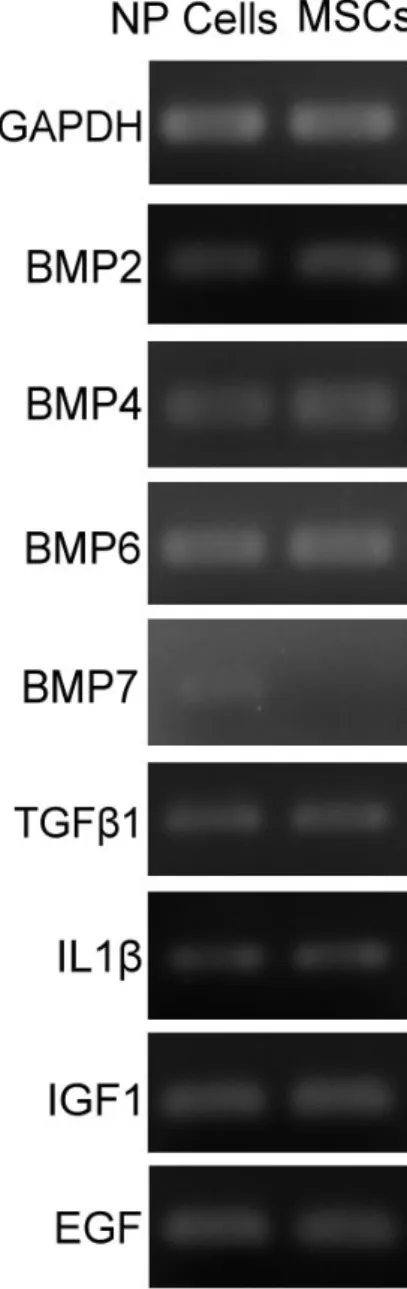
Monolayer-expanded human NP cells could manifest mRNA expression of BMP2, BMP4, BMP6, TGFß1, IL1ß, IGF1 and EGF. The expression of BMP7 was detectable in human NP cells but the level of expression was lower than the other cytokines. Monolayer-expanded MSCs expressed levels of BMP2, BMP4, BMP6, TGFβ1,IL1β, IGF1 and EGF comparable to NP cells. However, the expression of BMP7 could not be detected in MSCs (Fig. 1).
Proliferation of MSCs
The final cell number measured by WST-8 assay was 3.34±0.67x104 in Group M1, 3.78±0.78x104in Group M2, 4.02±0.86x104in Group M3, 4.09±0.94x104in Group M4, and 4.45±0.78x104in Group M5 (Fig. 1). Slight enhancement of MSC proliferation was observed in the co-culture conditions but was not statistically significant (p > 0.1). Proliferation of MSCs in the culture medium containing 10 ng/mL TGFß1 was significantly higher than that of untreated MSCs (p=0.025) but was only slightly higher than MSCs in other co-culture conditions (p > 0.1).
Relative mRNA Expressions of Selected Target Genes in MSCs
The relative mRNA expression of selected target genes in MSCs is summarized in Table 3. The mRNA expression of collagen I was significantly suppressed in the MSCs co-cultured with NP cells (Group M2: 76.6±12.3%, p=0.017; Group M3: 71.8±14.1%,
p=0.0011; Group M4: 72.4±10.1%, p=0.00016). However, the extent of suppression was
similar among different initial preparations (p > 0.5). The TGFβ1-treated MSCs showed a significantly higher level of collagen I mRNA in comparison with the untreated MSCs and co-cultured MSCs (247±99.8%, p <0.03).
There was no expression of collagen II mRNA in the untreated MSCs. Progressively higher levels of collagen II expression were detected as the number of co-cultured NP cells increased and the highest collagen II expression was found in the TGFß1-treated MSCs. In comparison with the TGFß1-treated MSCs, the level of collagen II expression was 3.8±7.7% in Group M2, 13.9±5.4% in Group M3, and 72.1±50.9% in Group M4. The mRNA expression of collagen II in the TGFß1-treated MSCs was significantly higher than that in Group M2 (p=0.00011) and M3 (p=0.00023), but was not significantly higher
than that in Group M4 (p=0.31).
The mRNA expression of aggrecan was suppressed in the MSCs co-cultured with NP cells, but the suppression was not statistically significant (Group M2: 87.5±49.2%,
p=0.69; Group M3: 89.9±33.8%, p=0.71; Group M4: 78.0±40.4%, p=0.45). On the other
hand, the level of aggrecan mRNA expression in the TGFβ1-treated MSCs was significantly lower than that in the untreated and the co-cultured MSCs (7.5±4.3%, p < 0.003).
The FADD expression of MSCs was slightly lower when co-cultured with fewer numbers of NP cells (Group M2: 89.0±28.3%, p=0.44), but was significantly suppressed when co-cultured with higher numbers of NP cells (Group M3: 75.4±19.3%, p=0.044; Group M4: 77.0±13.1%, p=0.027) or incubated in TGFβ1-containing culture medium (71.8±18.1%, p=0.021).
The expression of SOX9 in MSCs was slightly lower when co-cultured with NP cells (Group M2: 79.5±30.5%, p=0.25; Group M3: 88.9±26.4%, p=0.49; Group M4: 84.9±28.6%, p=0.37) and was slightly higher when treated with TGFß1 (138±68.7%,
p=0.24). However, none of the changes were statistically significant.
Proliferation of NP Cells
After 72 hours, the number of NP cells was 2.86±0.15x104 in the untreated group (N1) and 3.08±0.26x104in the TGFß1-treated group (N7) (Figure 2). There was no significant difference between these two groups (p=0.10). Proliferation of NP cells in every co-culture condition was significantly higher than that of untreated NP cells and TGFß1-treated NP cells (p < 0.003). The final number of NP cells was 3.87±0.63x104in Group N2, 4.22±0.78x104 in Group N3, 4.26±0.39x104 in Group N4, 4.33±0.70x104 in Group N5, and 4.27±0.62x104in Group N6. The final cell numbers were not significantly different among all co-culture groups (p > 0.2).
Relative mRNA Expression of Selected Target Genes in NP Cells
The relative mRNA expressions of selected target genes in NP cells are summarized in Table 4. The expression of collagen I in NP cells was slightly suppressed in every co-culture group, but the suppression was not statistically significant (Group N2: 91.0±7.3%, p=0.50; Group N3: 81.0±13.7%, p=0.19; Group N4: 90.2±27.8%, p=0.57; Group N5: 82.0±21.6%, p=0.27; Group N6: 87.0±23.5%, p=0.43). The TGFß1-treated
NP cells expressed a significantly higher level of collagen I mRNA than untreated NP cells (135±19.4%, p=0.037) as well as the NP cells co-cultured with MSCs (p < 0.002). The expression of collagen II mRNA was similar in untreated, co-cultured and TGFß1-treated NP cells (Group N2: 106±22.3%, p=0.68; Group N3: 108±27.4%, p=0.62; Group N4: 99.5±34.2%, p=0.98; Group N5: 97.6±34.2%, p=0.89; Group N6: 102±20.5%,
p=0.90; Group N7: 98.0±18.6%, p=0.88).
The mRNA expression of aggrecan was similar in untreated NP cells and NP cells co-cultured with fewer numbers of MSCs (Group N2: 100±35.6%, p=0.98; Group N3: 105±35.3%, p=0.78). When co-cultured with higher numbers of MSCs, the mRNA expression of aggrecan in NP cells was significantly enhanced (Group N4: 144±26.3%,
p=0.016; Group N5: 146±19.0%, p=0.0057; Group N6: 170±48.6%, p=0.011). The
TGFß1-treated NP cells also showed a significantly higher mRNA expression of aggrecan than untreated NP cells (163±41.9%, p=0.011). The enhancement of aggrecan mRNA expression was not significantly different among Groups N4, N5, N6 and N7 (p > 0.2).
The mRNA expression of FADD was similar in untreated and TGFß1-treated NP cells (102±28.1%, p=0.92). NP cells expressed lower levels of FADD mRNA in every co-culture condition, but the suppression was not statistically significant (Group N2: 83.3±24.3%, p=0.28; Group N3: 85.8±17.1%, p=0.29; Group N4: 88.7±20.9%, p=0.43; Group N5: 83.5±9.7%, p=0.17; Group N6: 83.4±30.5%, p=0.33).
The mRNA expression of SOX9 in NP cells was slightly enhanced when co-cultured with MSCs but not significantly (Group N2: 118±32.8%, p=0.23; Group N3: 113±28.6%,
p=0.34; Group N4: 119±32.9%, p=0.20; Group N5: 116±35.3%, p=0.31; Group N6:
114±24.9%, p=0.25).TheTGFβ1-treated NP cells showed a significantly lower level of SOX9 mRNA (73.6±10.7%) than untreated NP cells (p=0.002) and NP cells in every co-culture condition (p < 0.02).
DISCUSSION
Because of the restricted availability and suboptimal regenerative potential of autologous differentiated NP cells, uncommitted MSCs have caught special attention in tissue engineering therapy of IVD degeneration. Although the lineage of the different cell types in the NP is not known, it has been confirmed that cells of the NP and chondrocytes share many common features [8]. MSCs are undifferentiated cells found in small numbers in
the periosteum or in the bone marrow. They possess the unique ability to differentiate into varieties of cells found in connective tissues of mesenchymal origin including cartilage and bone [20]. Use of autologous MSCs can alleviate drawbacks resulting from the low quantity of autologous graft material, possible donor site morbidity and immunogenicity of allogenic cells [21]. Regarding use of MSCs for regeneration of IVD tissues, several approaches have been tested in the literature including differentiation of MSCs towards a NP-like cells by using growth factors [13,14], transplantation of MSCs to the IVD for tissue regeneration [22-26], and co-culture of MSCs and IVD cells to promote cell viability and matrix synthesis [27-29].
TGFβ-mediated induction of MSCs has been shown to result in a gene expression profile highly similar to that of native nucleus pulposus [13,14]. Nevertheless, the molecular and histological appearance of the TGFβ-treated MSCs is closer to that of fibrocartilage than that of hyaline articular cartilage because the expression of collagen I is also enhanced [14]. In the current study, TGFβ-treated MSCs showed some favorable changes including a lower probability of apoptosis by suppressing FADD expression, and improved chondrogenesis probability by upregulating SOX9 expression and inducing expression of collagen II. However, the enhanced expression of collagen I indicates more fibrotic changes might happen in the regenerative tissue and the reduced expression of aggrecan suggests a lower aggregation of glycosaminoglycans. For the purpose of achieving a more appropriate molecular phenotype and morphological features of hyaline cartilage, TGFβ-mediated chondrogenesis needs further optimization, for example, combining the use of several catabolic cytokines and growth factors [14]. It has been shown that proliferation and chondrogenesis of MSCs could be more effective if treated with proper combinations of growth factors [15,16]. It is reasonable to expect that MSCs can be stimulated by sustained secretion of cytokines from NP cells in an in vivo environment in the IVDs or an in vitro co-culture system. The current study showed that NP cells could manifest expression of BMP2, BMP4, BMP6, BMP7, TGFβ1, IGF1, and EGF. The current study also showed secreted cytokines from human NP cells could influence MSCs in many ways including slight promotion of cell proliferation, significant suppression of collagen I expression, slight suppression of aggrecan expression, significantly lower probability of apoptosis, and slightly lower expression of SOX9. The crucial finding signifying the success of induction of MSCs into NP-like cells is the appearance of collagen II expression. There was no collagen II expression detected in untreated MSCs
in the current study. After co-culture with NP cells, MSCs could start to manifest collagen II expression and the level of expression increased as the number of co-cultured NP cells increased.
Recent MSC studies in animal models evaluated the effects of transplanted MSCs in normal IVDs or the consequence of induced degeneration [22-26]. Transplantation of MSCs may not fully regenerate the disc but it could retard the overall degeneration process to some extent. Not only the TGFβ-treated autogenic MSCs could regenerate NP tissue in the degeneration-induced IVDs [22,24,26], but also transplanted untreated allogenic MSCs could survive and proliferate in nucleus pulposus and produce extracellular matrix [23,25]. It is hypothesized that the differentiation ability of MSCs is governed mainly by the environment in which they are placed [22], therefore even untreated MSCs could differentiate into NP-like cells when transplanted into the NP. For transplanted MSCs embedded inside scaffolds or carriers, direct cell-to-cell contact is not possible for MSCs residing deep in the center of the transplanted material. Therefore, cytokines secreted by the surrounding IVD tissues are the most possible mechanism to induce MSCs into NP-like cells. Furthermore, the tissue regeneration by MSC transplantation may perhaps simply require MSCs to have appropriately differentiated into “true”disc cells, or be efficient enough to act as helpers to induce endogenous disc cell proliferation and differentiation, which has not been sufficiently evaluated in animal models to date [30]. The non-contact co-culture system employed in the current study was intended to evaluate the interaction of MSCs and NP cells through secreted soluble factors such as cytokines.
It has been demonstrated that constitutively expressed cytokines in the MSCs include granulocyte colony stimulating factor (G-CSF), granulocyte-macrophage colony stimulating factor (GM-CSF), macrophage colony stimulating factor (M-CSF), leukemia inhibitory factor (LIF), IL6, IL11, and stem cell factor (SCF) [31]. Additional cytokines in MSCs including BMP2, BMP4, BMP6, TGFβ1, IGF1, EGF, IL1βhave been revealed in the current study. Through secreted cytokines, small numbers of MSCs could significantly promote proliferation of NP cells and, to a lesser extent, could slightly suppress apoptosis, slightly lower expression of collagen I, and slightly enhance chondrogenesis. Moreover, all these effects on NP cells could be achieved by co-culturing with a small number of MSCs and were not further altered by increasing the number of co-cultured MSCs. The only exception was the expression of aggrecan, which was
significantly higher in NP cells when co-cultured with higher numbers of MSCs. The expression of collagen II in NP cells was not affected in the co-culture system.
The results presented in the current study implicate a possible mechanism concerning interaction between MSCs and NP cells when both cell types are located in the same environment. Even though the current study showed a similar mRNA expression profiles of cytokines in both cell types, the secreted cytokines in the co-culture environment guided the uncommitted MSCs mainly toward the direction of differentiation and promoted the well-differentiated NP cells largely in the way of cell proliferation. Although the cell contact has been emphasized in previous studies co-culturing MSCs and IVD cells [27-29], the exact mechanism has not been proven yet. In addition, because MSCs and IVD cells possess very different cell phenotypes, the accuracy of determining phenotypic behaviors of each cell population before and after co-culture principally depends on a successful separation of two cell types. Any contamination, IVD cells exhibit in studies on MSCs for instance, will seriously compromise the conclusion obtained from those study results. Without conducting further studies regarding direct cell-to-cell contact, the results from the current study could reasonably assume that a stronger paracrine stimulation may exist when MSCs and IVD cells locate very closely to each other and generate a higher local concentration of excreted cytokines.
CONCLUSIONS
The current study utilized a non-contact co-culture system to reveal a possible interaction mechanism between MSCs and NP cells through paracrine stimulation. Through secreted cytokines, proliferation of NP cells could be significantly promoted even though the number of MSCs was relatively low in the environment. For the purpose of differentiating MSCs into NP-like cells with a higher level of collagen II expression, MSCs needed to be in an environment with high number of NP cells.
REFERENCES
1 Waddell G. Low back pain: a twentieth century health care enigma. Spine 1996;21:2820-2825.
2 Adler JH, Schoenbaum M, Silberberg R. Early onset of disk degeneration and spondylosis in sand rats (Psammomys obesus). Vet Pathol 1983;20:13-22.
4 Antoniou J, Stefen T, Nelson F, et al. The human lumbar intervertebral disc. J Clin Invest 1996;98:996-1003.
5 Ganey TM, Meisel HJ. A potential role for cell-based therapeutics in the treatment of intervertebral disc herniation. Eur Spine J 2002;11:S206-214.
6 Roberts S, Evans EH, Kletsas D, et al. Senescence in human intervertebral discs. Eur Spine J 2006;15:S312-316.
7 Le Maitre CL, Freemont AJ, Hoyland JA. Localization of degradative enzymes and their inhibitors in the degenerative human intervertebral disc. J Pathol 2004;204:47-54.
8 Sive JI, Baird P, Jeziorsk M, et al. Expression of chondrocyte markers by cells of normal and degenerative intervertebral discs. Mol Pathol 2002;55:91-97.
9 Pearce RH, Grimmer BJ, Adams ME. Degeneration and the chemical composition of the human lumbar intervertebral disc. J Orthop Res 1987;5:198-205.
10 Nomura T, Mochida J, Okuma M, et al. Nucleus pulposus allograft retards intervertebral disc degeneration. Clin Orthop 2001;389:94-101.
11 Thompson JP, Oegema TR Jr, Bradford DS. Stimulation of mature canine intervertebral disc by growth factors. Spine 1991;16:253-260.
12 Yamamoto Y, Mochida J, Sakai D, et al. Upregulation of the viability of nucleus pulposus cells by bone marrow-derived stromal cells. Spine 2004;29:1508-1514. 13 Risbud MV, Albert TJ, Guttapalli A, et al. Differentiation of mesenchymal stem
cells towards a nucleus pulposus-like phenotype in vitro: implications for cell-based transplantation therapy. Spine 2004;29:2627-2632.
14 Steck E, Bertram H, Abel R, et al. Induction of intervertebraldisc-like cells from adult mesenchymal stem cells. Stem Cells 2005;23:403-411.
15 Indrawattana N, Chen G, Tadokoro M, et al. Growth factor combination for chondrogenic induction from human mesenchymal stem cell. Biochem Biophys Res Commun 2004;320:914-919.
16 Im GI, Jung NH, Tae SK. Chondrogenic differentiation of mesenchymal stem cells isolated from patients in late adulthood: the optimal conditions of growth factors. Tissue Eng 2006;12:527-536.
17 Yang SH, Chen PQ, Chen YF, et al. An in-vitro study on regeneration of human nucleus pulposus cells by using gelatin/chondroitin-6-sulfate/hyaluronan tri-copolymer scaffold. Artif Organs 2005;29:806-814.
18 Kuhn DM, Balkis M, Chandra J, et al. Uses and limitations of the XTT assay in studies of Candida growth and metabolism. J Clin Microbiol 2003;41:506-508. 19 Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time
quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001;25:402-408. 20 Caplan AI. Mesenchymal stem cells. J Orthop Res 1991;9:641-650.
21 Risbud MV, Shapiro IM, Vaccaro AR, et al. Stem cell regeneration of the nucleus pulposus. Spine J 2004;4:S348-353.
22 Sakai D, Mochida J, Yamamoto Y, et al. Transplantation of mesenchymal stem cells embedded in Atelocollagen® gel to the intervertebral disc: a potential therapeutic model for disc degeneration. Biomaterials 2003;24:3531-3541.
23 Crevensten G, Walsh AJL, Ananthakrishnan D, et al. Intervertebral disc cell therapy for regeneration: mesenchymal stem cell implantation in rat interveretbral discs. Ann Biomed Eng 2004;32:430-434.
24 Sakai D, Mochida J, Iwashina T, et al. Differentiation of mesenchymal stem cells transplanted to a rabbit degenerative disc model. Potential and limitation for stem cell therapy in disc regeneration. Spine 2005;30:2379-2387.
25 Zhang YG, Guo X, Xu P, et al. Bone mesenchymal stem cells transplanted into rabbit intervertebral disc can increase proteoglycans. Clin Orthop 2005;430:219-226.
26 Sakai D, Mochida J, Iwashina T, et al. Regenerative effects of transplanting mesenchymal stem cells embedded in atelocollagen to the degenerated intervertebral disc. Biomaterials 2006;27:335-345.
27 Yamamoto Y, Mochida J, Sakai D, et al. Upregulation of the viability of nucleus pulposus cells by bone marrow-derived stromal cells. Spine 2004;29:1508-1514. 28 Richardson SM, Walker RV, Parker S, et al. Intervertebral disc cell-mediated
mesenchymal stem cell differentiation. Stem Cells 2006;24:707-716.
29 Le Visage C, Kim SW, Tateno K, et al. Interaction of human mesenchymal stem cells with disc cells. Changes in extracellular matrix biosynthesis. Spine 2006;31:2036-2042.
30 Leung VYL, Chan D, Cheung KMC. Regeneration of intervertebral disc by mesenchymal stem cells: potentials, limitations, and future direction. Eur Spine J 2006;15 (Suppl 3):S406-413.
marrow-derived mesenchymal progenitor cells in vitro: effects of dexamethasone and IL-1 alpha. J Cell Physiol 1996;166:585-592.
Figure 1. Expression Profiles of Cytokines in NP cells (left column) and MSCs (right column). Comparable levels of RNA expressions of selected cytokines were detected in NP cells and MSCs except for BMP7.
出 席 國 際 學 術 會 議 心 得 報 告
計畫編號 95-2314-B-002-148- 計畫名稱 間質幹細胞與椎間盤本核細胞共同培養條件最佳化之研究–對脊椎椎間盤再生之現行細胞移植療法改進之探討 出國人員姓名 服務機關及職稱 楊曙華 台大醫院骨科部 主治醫師 會議時間地點 2007年2月11至14日 美國加州聖地牙哥 會議名稱 第53屆骨科研究學會 發表論文題目3-DIMENSIONAL CULTURE OF HUMAN NUCLEUS PULPOSUS CELLS IN FIBRIN CLOT AND ALGINATE - An in-vitro study on feasibility of fibrin clot as a bioactive scaffold for regeneration of nucleus pulposus tissue INTERACTIONS BETWEEN HUMAN INTERVERTEBRAL DISC CELLS AND MESENCHYMAL STEM CELLS IN A NON-CONTACT
CO-CULTURE SYSTEM 一、參加會議經過
美國之骨科研究學會應是全世界最盛大的骨骼、肌肉、神經等骨科相關領域基礎研究之 會議,每年二、三月間輪流於美國南方及西岸城市舉行,與緊接在後舉行之美國骨科醫 學會(American Academy of Orthopaedic Surgeons)是每年年初全球骨科臨床及基礎研究 者的大事,能於大會中發表論文實為榮幸。本人此次是繼2005年後再一次獲得於此一會 議中發表研究論文。 於2月10日抵達會議地點美國加州聖地牙哥後,隨即至會場佈置參展論文之壁報,正式會 議於11日上午七點開始為期四天的緊湊議程。每天於五個會議廳共進行300篇口頭論文發 表,此外還有約1600篇壁報論文同時展示於大展覽廳。要於短短四天內瀏覽、消化如此 大量的研究資訊並非易事,但想起這些研究成果是由近萬篇投稿中篩選出來的傑作,不 得不盡力克服時差爭取有限時間吸收最新知識。 直接與椎間盤有關研究僅有一個口頭報告時段,與本研究團隊直接相關之椎間盤組織再 生之研究僅散見於壁報論文中,可見此一研究領域涉足者仍為數不多。為增廣研究寬度 及深度,會中本人積極參與Spine biology、Chondrocyte gene regulation、progenitors and stem cells、Tissue engineering、Intervertebral disc等口頭論文發表之觀摩。壁報論文部分擇著 重於Spine biology、Intervertebral disc、Tissue engineering、Cartilage、Stem cells等領域論 文之研讀。會中除吸收其他研究機構於口頭論文及壁報論文之構思及成果外,亦與來自 美國各地及瑞典之研究學者就彼此共同涉略之研究領域交換意見。
二、與會心得
由於本研究團隊著重於發展生醫活性材料以促進人類椎間盤細胞活性及細胞外基質之合 成,由近幾次與會之經驗發現完全近似發展方向之研究團對並不多,持續保持目前之研
(Mesenchymal stem cells)與椎間盤細胞交互作用之研究,因幹細胞研究數量較多,需 增進目前研究質量方可符合幹細胞研究之要求。至於本研究團隊最近致力發展有關以 Statins藥物調控椎間盤細胞表現之研究,此次會議有一篇壁報論文發表,再與其研究者 交談中發現彼此意見相同之處,一方面欣喜於研究方向之未來,一方面亦警惕自己需加 快研究腳步,以免錯失論文發表之先機。
出 席 國 際 學 術 會 議 心 得 報 告
計畫編號 95-2314-B-002-148- 計畫名稱 間質幹細胞與椎間盤本核細胞共同培養條件最佳化之研究–對脊椎椎間盤再生之現行細胞移植療法改進之探討 出國人員姓名 服務機關及職稱 楊曙華 台大醫院骨科部 主治醫師 會議時間地點 2007年2月11至14日 美國加州聖地牙哥 會議名稱 第53屆骨科研究學會 發表論文題目3-DIMENSIONAL CULTURE OF HUMAN NUCLEUS PULPOSUS CELLS IN FIBRIN CLOT AND ALGINATE - An in-vitro study on feasibility of fibrin clot as a bioactive scaffold for regeneration of nucleus pulposus tissue INTERACTIONS BETWEEN HUMAN INTERVERTEBRAL DISC CELLS AND MESENCHYMAL STEM CELLS IN A NON-CONTACT
CO-CULTURE SYSTEM 一、參加會議經過
美國之骨科研究學會應是全世界最盛大的骨骼、肌肉、神經等骨科相關領域基礎研究之 會議,每年二、三月間輪流於美國南方及西岸城市舉行,與緊接在後舉行之美國骨科醫 學會(American Academy of Orthopaedic Surgeons)是每年年初全球骨科臨床及基礎研究 者的大事,能於大會中發表論文實為榮幸。本人此次是繼2005年後再一次獲得於此一會 議中發表研究論文。 於2月10日抵達會議地點美國加州聖地牙哥後,隨即至會場佈置參展論文之壁報,正式會 議於11日上午七點開始為期四天的緊湊議程。每天於五個會議廳共進行300篇口頭論文發 表,此外還有約1600篇壁報論文同時展示於大展覽廳。要於短短四天內瀏覽、消化如此 大量的研究資訊並非易事,但想起這些研究成果是由近萬篇投稿中篩選出來的傑作,不 得不盡力克服時差爭取有限時間吸收最新知識。 直接與椎間盤有關研究僅有一個口頭報告時段,與本研究團隊直接相關之椎間盤組織再 生之研究僅散見於壁報論文中,可見此一研究領域涉足者仍為數不多。為增廣研究寬度 及深度,會中本人積極參與Spine biology、Chondrocyte gene regulation、progenitors and stem cells、Tissue engineering、Intervertebral disc等口頭論文發表之觀摩。壁報論文部分擇著 重於Spine biology、Intervertebral disc、Tissue engineering、Cartilage、Stem cells等領域論 文之研讀。會中除吸收其他研究機構於口頭論文及壁報論文之構思及成果外,亦與來自 美國各地及瑞典之研究學者就彼此共同涉略之研究領域交換意見。
二、與會心得
由於本研究團隊著重於發展生醫活性材料以促進人類椎間盤細胞活性及細胞外基質之合 成,由近幾次與會之經驗發現完全近似發展方向之研究團對並不多,持續保持目前之研
(Mesenchymal stem cells)與椎間盤細胞交互作用之研究,因幹細胞研究數量較多,需 增進目前研究質量方可符合幹細胞研究之要求。至於本研究團隊最近致力發展有關以 Statins藥物調控椎間盤細胞表現之研究,此次會議有一篇壁報論文發表,再與其研究者 交談中發現彼此意見相同之處,一方面欣喜於研究方向之未來,一方面亦警惕自己需加 快研究腳步,以免錯失論文發表之先機。