Cloning and expression of carp cathepsin Z:
Possible involvement in yolk metabolism
Chi-Min Kao
a, Fore-Lien Huang
a,b,⁎
a
Institute of Molecular and Cellular Biology, National Taiwan University, Taipei, Taiwan
b
Institute of Biological Chemistry, Academia Sinica, Taipei, Taiwan
Received 27 October 2005; received in revised form 15 May 2006; accepted 16 May 2006 Available online 26 January 2008
Abstract
A cDNA encoding cathepsin Z (CTPZ) was cloned from a carp ovarian cDNA library. It is homologous to mammalian CTPZ. The amino acid residues important for protein folding and enzymatic activity of mammalian CTPZ are conserved in carp CTPZ. It is widely expressed in a variety of carp tissues as revealed by Western blot and reverse transcription-polymerase chain reaction. The CTPZ mRNA was transiently accumulated during oocyte maturation. In oocytes, CTPZ is localized in cortical granules and in the cytoplasm surrounding the yolk granules. After fertilization, CTPZ remained associated with the yolk granules while the cortical granular CTPZ was discharged to plasma membrane, perivitelline space, and fertilization envelope. Carp cathepsin Z has proteolytic activity toward vitellogenin that could be inhibited by inhibitors specific for the proteases of papain family. The potential roles of cathepsin Z in carp eggs are discussed.
© 2008 Published by Elsevier Inc.
Keywords: Carp; Cathepsin Z; Cortical granule; Fertilization envelope; Oocyte maturation; Vitellogenin
1. Introduction
Cathepsins are divided into four families according to the identity of the critical residue in the active site, cysteine-, aspartyl-, serine- and metal ions in metalloprotease. Cathepsin Z belongs to the group of cysteine proteases, which represent the major component of lysosomal proteolysis system (Kirschke and Wiederanders, 1987). Cathepsins are found in yeast (Gazith et al., 1968), baculovirus (Vernet et al., 1990), eubacteria (Gething and Davidson, 1976), and practically all protozoa (Rautenberg et al., 1982), plants (Lynn et al., 1980) and mammals (Banon et al., 1964). In human, the expression of cathepsin B, L, H, O, C, Z, and F are ubiquitous (Chan et al., 1986; Troen et al., 1987; Fuchs and Gassen, 1989; Velasco et al., 1994; Paris et al., 1995; Nagler and Menard, 1998; Santamaria et al., 1998b; Wang et al., 1998), whereas expression of cathepsin S, K, W, L2 and J are more
restricted (Shi et al., 1992; Inaoka et al., 1995; Linnevers et al., 1997; Santamaria et al., 1998a; Tisljar et al., 1999).
Cathepsins are thought to function primarily in intracellular protein degradation and turnover. Recent studies suggest that these enzymes may also play specific functions such as polypeptide chain processing (Riese et al., 1996; Villadangos et al., 1997), bone remodeling (Tezuka et al., 1994), proenzyme activation (Azaryan and Hook, 1994) and allergic reactions (Kalsheker et al., 1996). In some cases, cathepsins can escape from the normal targeting pathway and can be secreted into the extracellular space, and function extracellularly in pathophysio-logical conditions such as arthritis (Esser et al., 1993, 1994), pulmonary emphysema (Mason et al., 1986), muscular dystro-phy (Katunuma and Kominami, 1987), Alzheimer's disease (Cataldo et al., 1991), rheumatoid arthritis (Hashimoto et al., 2001), tumor invasion and metastasis (Sloane and Honn, 1984). A majority of cathepsins identified in mammals were also found in fish. Some information about their functions is now available. In sockeye salmon (Oncorhynchus nerka), the proteo-lysis activity of cathepsin D in muscle was significantly increased during the anorexic freshwater migration (Mommsen et al., 1980)
Comparative Biochemistry and Physiology, Part B 149 (2008) 541–551
www.elsevier.com/locate/cbpb
⁎ Corresponding author. Institute of Molecular and Cellular Biology, National Taiwan University, No. 1 Sec. 4, Roosevelt Road, Taipei, Taiwan 10617. Tel.: +886 2 33662481; fax: +886 2 33662478.
E-mail address:[email protected](F.-L. Huang). 1096-4959/$ - see front matter © 2008 Published by Elsevier Inc. doi:10.1016/j.cbpb.2006.05.017
and these protein-derived amino acids are used for gonad maturation, oxidation and gluconeogenesis (Mommsen, 2004). In the white muscle of chum salmon (Oncorhynchus keta), cathepsins B, D, H and L activities were 3–7 times higher in the fish during spawning migration than those during feeding migration (Yamashita and Konagaya, 1990). In female Atlantic salmon (Salmo salar), cathepsin D also promoted the proteolysis of white muscle proteins during spawning and the muscle protein degradation was inferred to be connected with gonad maturation (von der Decken, 1992). In addition to its role in muscle de-gradation, fish cathepsin D could process vitellogenin into yolk proteins (Wallace and Selman, 1985; Brooks et al., 1997). Similar function of cathepsin D on vitellogenin processing was observed in amphibia (Opresko and Karpf, 1987) and bird (Retzek et al., 1992). Interestingly, a liver- and female-specific aspartic protease (LAP) of zebrafish (Danio rerio), which is closely related to cathepsin D, is involved in maturation and vitellogenesis (Riggio et al., 2002). LAPs were also identified from pufferfish (Takifugu rubripes) and two Antarctic species (Mommsen, 2004).
Cathepsin B, S and L were also found in fish. The cathepsin B-subfamily comprises cathepsin B, C and Z. In killifish, the lipovitellin heavy chain of 122 kDa was degraded during oocyte maturation, concomitant with an increased cathepsin B activity (LaFleur et al., 2005). In Atlantic salmon, cathepsin B is localized in epithelial cells of the proximal tubules of trunk kidney and mucosal epithelial cells of the esophagus and intestine. In head kidney, cathepsin B is present in large macrophage-like cells (Tahtinen et al., 2002). Similarly, in chum salmon, cathepsin B was demonstrated to be present in macrophage-like phagocytes near/in the necrotic muscle fibers in the white muscle. These observations suggest that cathepsin B activity may be related to muscle protein metabolism and immune response (Yamashita and Konagaya, 1991). Cathepsin B has also been suggested to be involved in the proteolysis of yolk proteins during fish and amphibian embryogenesis (Yoshizaki et al., 1998). In carp, another cysteine protease, cathepsin S-like protease forms an apparently monophyletic group with the mammalian S cathepsins by phylogenetic analysis (Uinuk-ool et al., 2003). In fact, cathepsin S is highly expressed in spleen, B-lymphocytes, macro-phages and dendritic cells in mammals (Kirschke et al., 1989; Morton et al., 1995), implicating the functional specificity of antigen processing in immune system.
The cathepsin L-like subfamily comprises cathepsin H, K, S, L, L2, W, F, O and the rodent placentally expressed cathepsins (PECs) (Uinuk-ool et al., 2003). Cathepsin L is abundant in fish. Its function has been reported to be associated with muscle protein degradation (Yamashita and Konagaya, 1990), antigen processing (Uinuk-ool et al., 2003) and pathogen defense
(Aranishi, 1999). Furthermore, in seabream (Sparus aurata), cathepsin L is responsible for the proteolytic cleavage of lipovitellin components during oogenesis (Carnevali et al., 1999). In killifish (F. heteroclitus), cathepsin L and F have been suggested to play a role in yolk proteins processing, by which the free amino acids facilitate oocyte to acquire buoyancy (Wallace and Selman, 1985; Fabra and Cerda, 2004).
Recently, the mRNA levels of cathepsin Z and other seven cathepsins were investigated in the ovarian follicles of killifish (Fabra and Cerda, 2004). The expression of cathepsins L, S, H, and F remained unchanged during vitellogenesis but was transiently increased during oocyte maturation, suggesting that these four cathepsins may be involved in oocyte maturation-associated yolk metabolism. The level of cathepsin Z mRNA of killifish remained almost constant during vitellogenesis and oocyte maturation. No specific function had been ascribed to cathepsin Z in killifish. However, cathepsin Z was found to be involved in molting in Caenorhabditis elegans and in Oncho-cerca volvulus (Hashmi et al., 2004; Lustigman et al., 2004), and in host tissue invasion in Myxobolus cerebralis (Kelley et al., 2003). In addition, cathepsin Z is highly expressed in human prostate carcinoma (Nagler et al., 2004). These findings indicate that cathepsin Z has certain important functions in a variety of animals.
Previously, we cloned a partial cDNA encoding cathepsin Z from a carp ovarian cDNA library (Chang et al., 1996). In order to understand the structural properties and the possible functions of cathepsin Z in carp oocytes, we first tried to obtain a full-length cDNA of carp cathepsin Z then investigated its subcellular localization in oocytes, its expression during oocyte maturation, and its proteolytic activity. In this study, we found that carp cathepsin Z has proteolytic activity toward vitello-genin and the mRNA level of cathepsin Z was transiently increased during oocyte maturation. Furthermore, cathepsin Z is bound to yolk granules. These results suggest that cathepsin Z may be involved in yolk metabolism, particularly during oocyte maturation. In addition, cathepsin Z is also localized in other organelles of eggs, suggesting that cathepsin Z may have additional functions in fish eggs.
2. Materials and methods
2.1. Cloning and sequencing of carp cathepsin Z cDNA A cDNA homologous to human cathepsin Z was found in a carp (Cyprinus carpio) ovarian cDNA library constructed previously (Chang et al., 1996). It lacks the 5′ region. The 5′ region was
obtained by 5′ rapid amplification of cDNA ends (5′-RACE) using
Table 1
The oligonucleotide sequence of the primers used for the rapid amplification of cathepsin Z cDNA ends and RT-PCR
Primers Sequence Length (bp) Temperature (°C)
916AA1 5′-GTTCAACATGGCTCGCGTGGTC-3′ 22 61
916MP1 5′-CCTCCCCAAAGCATGGGACTGGAG-3′ 24 65
916EE1 5′-TCAAGGCTTCACGTGTTGACATGG-3′ 24 61
916GSP1 5′-CATCAGGAATGCCATTGCTGTGTGCA-3′ 26 60
the gene-specific primer (916GSP1) and AP1 primer. All primer sequences used in this study are listed inTable 1.
Briefly, total RNA was extracted from fully-grown oocytes using the Ultraspec™ RNA isolation system (Biotecx Labora-tories, Inc., TX, USA) according to the manufacturer's instructions. The polyA+ RNA was purified on oligo(dT)-cellulose column (Invitrogen, CA, USA) for synthesis of the single-stranded cDNA by AMV reverse transcriptase (Clontech, CA, USA). The resultant cDNA was used directly for the second-stranded cDNA synthesis. The double-stranded cDNA was ligated with the commercial adaptor (AP) and used as template for subsequent 5′-RACE according to the manufacturer's instructions (Marathon cDNA Amplification Kits, Clontech). PCR amplification was performed as follows: denaturation at 94 °C for 30 s and extension at 68 °C for 2 min for 30 cycles, followed by an extension for 5 min at 72 °C using a Perkin-Elmer 2400 thermal cycler. The PCR products were analyzed on a 0.8% agarose gel. The fragment of expected size was excised and further purified by phenol/chloroform extraction. After the 5′ cDNA end was obtained, the full sequence of cathepsin Z cDNA was obtained by PCR using the primers (forward: 916AA1; reverse: 916EE1) and cloned into pGEM T-easy vector (Promega, WI, USA). The nucleotide sequence was determined by the dideoxy chain termination methods (Sanger et al., 1977) using a Thermo Sequenase Kit purchased from Amersham Life Science (Buckinghamshire, UK). Computer analysis of the cDNA and deduced amino acid sequence was performed with the software of DNASIS (Hitachi Software Engineering Co., Japan).
2.2. Expression of recombinant cathepsin Z and induction of antiserum
To prepare an expression vector suitable for production of functional cathepsin Z in E. coli, the coding sequence contained the mature carp cathepsin Z was amplified by PCR using the 916MP1 (forward) and 916EE1 (reverse) primers. The PCR was carried out for 30 cycles of denaturation at 95 °C for 30 s, annealing at 58 °C for 30 s, and extension at 68 °C for 1 min, followed by a final 5 min extension at 72 °C. The amplified product was gel-purified and cloned into the pGEMT-easy vector. The recombinant plasmid was further transformed into E. coli (DH5α™) (Life Technologies, Inc.). The insert was excised by EcoR I digestion and subcloned into the pGEX-2T vector (Pharmacia LKB Biotechnology) for protein expression. The expression vector was transformed into E. coli (BL21) (Novagen) and cultured in 100 mL medium (10 g/L bacto-tryptone, 5 g/L yeast extract, 5 g/L NaCl, 2% glucose and 100 µg/mL ampicillin, adjusted to pH 8.0) at 37 °C. At an OD600 of 0.8, isopropyl-1-thio-β-D-galactopyranoside (IPTG) was added to a final concentration of 0.1 mM, and the incubation was shifted to 28 °C for 10 h.
The recombinant protein was analyzed by SDS-polyacryla-mide gel electrophoresis (SDS-PAGE) as described byLaemmli (1970)and the protein band of expected size was cut-out. The gel-purified protein was used as antigen to induce antiserum. Antiserum was induced in adult New Zealand white rabbits (Oryctolagus caniculus) by subcutaneous injection with 100 µg of recombinant protein in Freund's complete adjuvant
(Sigma-Aldrich). Subsequent boosts with Freund's incomplete adjuvant were performed at intervals of 2 weeks.
2.3. Purification of recombinant protein by high performance liquid chromatography
The IPTG-induced cells were harvested, washed and subjected to three freeze-thaw cycles. The treated cells were sonicated and the lysate was filtered using a 0.45 µm syringe filter (PALL Life Sciences, Germany). The glutathione S-transferase-cathepsin Z (GST-CTPZ) fusion protein was affinity purified at 4 °C by a GSTrap™ FF column (Amersham Biosciences) and eluted with 20 mM reduced glutathione (Calbiochem). Then, the affinity-purified proteins were dialyzed against 0.05 M Tris–HCl, pH 8.0, concentrated using a YM-10 membrane (Amicon, USA), and further purified by high performance liquid chromatography (HPLC) Mono Q HR 5/5 anion-exchange column (Amersham Biosciences). The column was eluted with a flow rate of 1 mL/ min within 22 min with the following gradient program: 0–2 min, buffer A (0.05 M Tris–HCl, pH 8.0); 2–22 min, 0–100% buffer B (0.05 M Tris–HCl, pH 8.0, 1 M NaCl). Protein was monitored by absorbance at 280 nm. Each peak was collected and analyzed electrophoretically.
2.4. Northern blot
Oocytes maturation in carp was induced with ovaprim™ (Vancouver, Canada)– a GnRH analog – as described previously (Chang et al., 1996). After induction, the ovaries were removed at various time intervals and stored in liquid nitrogen. Twenty micrograms of total RNA were electrophoresed on a 1% agarose gel as described elsewhere (Esch et al., 1986). After electrophor-esis, RNA was transferred onto a nylon membrane (Amersham Biosciences) by capillary blotting using 20× SSC (0.3 M sodium citrate, pH 7.0, 3 M NaCl) for 16 h. Then, RNA was immobilized on a nylon membrane by a UV cross-linker (Hoefer, CA). The DNA probe was prepared using the DIG (digoxigenin) DNA labeling kit (Boehringer Mannheim, Germany) according to the manufacturer's instructions. Pre-hybridization was carried out in 5× SSC, 50% deionized formamide, 0.1% SDS, 0.1% N-lauroylsarcosine, 5× Denhardt's solution (1 × Denhardt's solu-tion = 0.02% bovine serum albumin, 0.02% polyvinylpyrrolidone, 0.02% Ficoll), 6.28 mg/mL yeast tRNA and 1% blocking reagent (Roche) for 4 h at 42 °C. Hybridization was performed at 42 °C overnight with DIG-labeled cathepsin Z cDNA (25 ng/mL). After hybridization, the membrane was washed with 0.1× SSC buffer containing 0.1% SDS for 30 min at 50 °C, followed by incubation with alkaline phosphatase-conjugated anti-DIG antibody (Pierce, IL, USA) for 1 h at room temperature. The membrane was washed twice for 1 h in washing buffer (0.1 M maleic acid, pH 7.5, 0.3% Tween-20, 0.15 M NaCl). After rinsing with detection buffer (0.1 M Tris–HCl, pH 9.5, 0.1 M MgCl2, 0.1 M NaCl), nitroblue tetrazolium salt and 5-bromo-4-chloro-3-indolyl phosphate (NBT/ BCIP) were used for color development as described previously (Chang et al., 1996). The hybridization signal was analyzed using the Chemilmager 4000™ image analysis system (Alpha Innotech Corporation, San Leandro, CA, USA).
2.5. Reverse transcription-polymerase chain reaction (RT-PCR) RT-PCR was performed using the Ready-To-Go™ RT-PCR Beads (Amersham Biosciences). Briefly, first-stranded cDNA was synthesized from 1 µg of total RNA extracted from various tissues. The RNA was subjected to reverse-transcription using oligo(dT)12–18primer by Moloney Murine leukemia virus (M-MuLV) reverse transcriptase (Invitrogen) at 42 °C for 15 min. The product was further amplified by PCR using gene-specific primers (916MP1 and 916EE1). The reaction was performed as follows: initial denaturation at 95 °C for 5 min, followed by 30 cycles of denaturation at 95 °C for 45 s, annealing at 56 °C for 30 s, and extension at 72 °C for 40 s, followed by a final 5 min extension at 72 °C. Aliquots of the PCR mixtures were analyzed on a 0.8% agarose gel.
2.6. Western blot
For Western blotting analysis, tissues were homogenized in extraction buffer (0.05 M Tris–HCl, pH 7.2, 0.1 M NaCl, 1 mM PMSF). Five microgram of protein was resolved on a 12% SDS-PAGE gel. The proteins were transferred onto a nitrocellulose
membrane (Schleicher & Schuell, Dassel, Germany) using electrophoresis as described previously (Chang et al., 1996). The blot was incubated with rabbit anti-cathepsin Z polyclonal antiserum at 4 °C for 16 h. After washing with PBST (1× PBS, 0.05% Tween-20), it was further incubated with horseradish peroxidase conjugated goat anti-rabbit IgG (Pierce, IL, USA) at room temperature for 1 h. Then, 3, 3′-diaminobenzidine (DAB) was used to develop the blot as previously described (Chang et al., 1996).
2.7. Immunohistochemistry
The ovarian tissues were prepared as previously described (Wang and Huang, 2002). Paraffin sections were cut into 10 µm in thickness. Deparaffined sections were boiled in 0.01 M citrate buffer, pH 6.0, for 15 min. After washing with PBST, the sections were incubated with rabbit anti-cathepsin Z polyclonal antiserum at 4 °C for 16 h and further incubated with horseradish peroxidase conjugated goat anti-rabbit IgG for 1 h at room temperature. The procedures of immunostaining were the same as previously described (Chang et al., 1996). The pre-immune rabbit serum was used as control.
Fig. 1. Nucleotide sequence and deduced amino acid sequence of carp cathepsin Z cDNA. A putative signal peptide is boxed. The cleavage site of the mature enzyme is indicated by arrowhead, and the active site residues Cys84, His233and Asn254are underlined. The putative N-linked glycosylation site is marked by a dotted line. The
2.8. Preparation of carp vitellogenin
Mature females were purchased from local market and maintained in laboratory for 1 week prior to use. The fish were injected intraperitoneally on day 1 and 7 with 17β-estradiol as previously described (Chang et al., 1996). After treatment for 14 days, blood was taken from the caudal vessel. The serum was dialyzed against 0.1 M sodium phosphate, pH 7.8, containing 0.2 M NaCl at 4 °C for 16 h and separated by a gel-filtration column of Sephacryl™ S-200 HR (Amersham) (1.6×90 cm) with a flow rate of 0.4 mL/min. The purified vitellogenin was analyzed on a 10% SDS-PAGE gel and stored at−70 °C. 2.9. Enzymatic activity assay of recombinant cathepsin Z
Protease activity was assayed by gelatin substrate gel elec-trophoresis as described elsewhere (Keene et al., 1986). One-tenth
microgram of recombinant cathepsin Z was electrophoresed on a 10% SDS-PAGE gel copolymerized with 0.1% gelatin (Sigma) under non-reducing condition. After electrophoresis, the gel was washed twice in 2.5% Triton X-100 for 20 min each, subjected to rinse twice in distilled water, and further incubated in reaction buffer (0.1 M sodium acetate, pH 5.0, 5 mM EDTA, 8 mM DTT, 0.15 M NaCl) for 20 h at 37 °C. After staining with 0.1% Coomassie brilliant blue and several cycles of destaining, the digested band was visualized as non-staining area on the gel. The enzymatic activity was also assayed by degradation of carp vitellogenin. Ten microgram of vitellogenin was incubated with 100 ng GST-CTPZ in digestion buffer (0.1 M sodium acetate, pH 5.5, 5 mM EDTA, 2 mM DTT, 0.1 M NaCl) at 37 °C for 2 h and then subjected to electrophoresis on an 8% SDS-PAGE gel. Proteases inhibitors were used to examine their effects on the enzymatic activity of recombinant cathepsin Z. The recombinant enzyme was pre-incubated with either 5 µM
trans-epoxysuccinyl-Fig. 2. Alignments of cathepsin Z. Data were obtained from GenBank: C. carpio, AY949988; F. heteroclitus, AY226449; H. sapiens, NM_001336; M. musculus, NM_022325; U. caupo, U30877; C. elegans, NM_058622; O. volvulus, AY591516; T. canis, AF143817, and M. cerebralis, AY224078. The multiple sequence alignment was performed using the Clustal W program. Identical amino acid residues were shaded. Gaps indicated by dotted lines are introduced for maximal alignment.
L-leucyl-amido-(4-guanidino)butane (E-64) (Calbiochem) or 1 mM phenylmethylsulfonylfluoride (PMSF) (Roche) or 1 µM pepstatin (Calbiochem) at 37 °C for 15 min prior to the addition of vitellogenin (10 µg). Incubation was further performed at 37 °C for 2 h and then analyzed electrophoretically.
2.10. Statistical analysis
The data were statistically analyzed by one-way ANOVA and followed by Duncan's multiple-range test.
3. Results
3.1. cDNA nucleotide and deduced amino acid sequences of carp cathepsin Z
A partial cathepsin Z cDNA was screened from a carp ovarian cDNA library. The full-length cDNA was obtained by 5′-RACE. The nucleotide sequence and the deduced amino acid sequence are presented inFig. 1. It is 1369 bp in length and contains an open reading frame of 903 bp that encodes a protein of 301 amino acid residues. The deduced amino acid sequence comprises a signal peptide of 17 residues, a pro-peptide of 36 residues and a mature enzyme of 248 residues with a calculated molecular mass of 28,082 Da. The putative signal sequence of cathepsin Z contains 76% hydrophobic amino acid that suggests it may be destined for secretion or transported to cell organelles. The pro-peptide of carp cathepsin Z contains two lysine residues that are thought to be the recognition site for mannose phosphorylation required for tar-geting the protein to the secretory pathway via endoplasmic
reticulum (Erickson, 1989). Carp cathepsin Z contains twelve cysteine residues including one (Cys27) in the pro-peptide. The presence of cysteine residue in the pro-peptide is an unique feature of cathepsin Z, which is critical for enzymatic activity regulation (Sivaraman et al., 2000). Carp cathepsin Z contains only one potential N-glycosylation site (NFTL, position 176–179), rather than two as found in human cathepsin Z. The mature enzyme contains the characteristic active site (Cys84, His233, Asn254) as found in proteases of the papain family. A His-Ile-Pro insertion was present in front of the cysteine residue (Cys84) of the active site in the cathepsin Z of carp and other species (Fig. 2). Such an insertion is unique to cathepsin Z (Nagler et al., 1999). Therefore there is a HIP-subfamily of cysteine proteases among the papain-like family (Santamaria et al., 1998b; Falcone et al., 2000). 3.2. Cathepsin Z is widely expressed in various tissues of carp
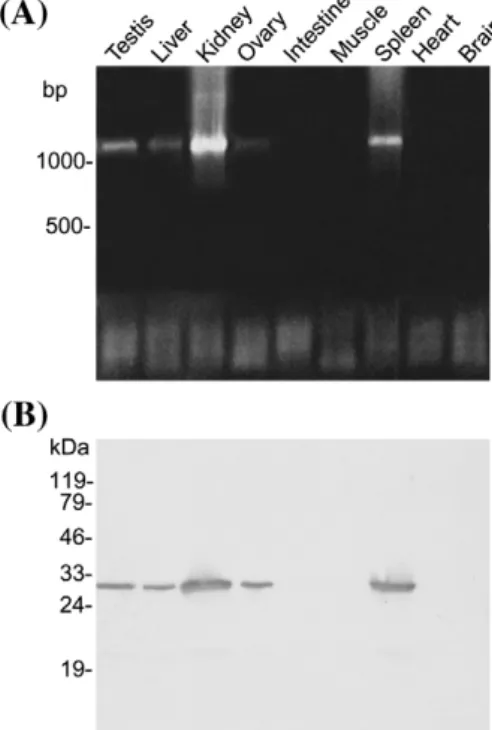
To investigate the distribution of cathepsin Z in carp tissues, PCR and Western blotting were employed. The results of RT-PCR showed that a higher expression level was found in testis, kidney and spleen, and a lower level was found in liver and the fully-grown oocytes (Fig. 3A), while no transcripts could be detected in intestine, muscle, heart and brain. Western blotting confirmed these results. Kidney and spleen contained a higher amount of cathepsin Z than testis, liver and oocyte. In contrast, the
Fig. 3. Expression of cathepsin Z in carp tissues. (A) RT-PCR. One microgram total RNA prepared from each sample was used for RT-PCR by a pair of specific primers that amplified a fragment of 1.1 kb DNA. The amplified DNA was subjected to electrophoresis on a 0.8% agarose gel and stained with ethidium bromide. (B) Western blot. Five microgram protein from various tissues was analyzed by the antiserum against carp cathepsin Z.
Fig. 4. Change in cathepsin Z transcripts during oocyte maturation. Twenty microgram of total RNA prepared from oocytes treated with GnRH analog for varying period of time was analyzed on a 1% agarose gel. After blotting, the membrane was stained by ethidium bromide to visualize rRNA (A), and probed with DIG-labeled cathepsin Z cDNA to detect cathepsin Z transcripts (B). The hybridization signals were normalized against the intensity of 18S rRNA. The normalized hybridization intensity of 8-h sample was used as reference to calculate the relative intensity of the sample of other time intervals (C). The sample of each time interval consisted of three carps. Duplicate RNA samples from each carp were analyzed. Shown are means ± SE. The different letter indicates significant difference (pb0.001). Ov, ovulated egg.
protein concentration in intestine, muscle, heart and brain is below the detection level (Fig. 3B). The molecular mass of carp cathepsin Z was estimated to be 28 kDa by Western blotting, which is close to the expected molecular mass of the mature enzyme.
3.3. Expression of cathepsin Z mRNA during oocyte maturation Cathepsins were suggested to be involved in yolk protein metabolism. Therefore, we investigated the expression pattern of carp cathepsin Z during oocyte maturation. The ovarian RNA from fish treated with GnRH (gonadotropin-releasing hormone) analog for various length of time was analyzed by Northern blotting. The cathepsin Z mRNA was detected as a single transcript of 1.4 kb (Fig. 4B). The intensity of hybridization signals of cathepsin Z transcripts increased during the early phase of oocyte maturation, reaching peak at 8 h post-treatment, then decreased to the level of untreated oocytes in ovulated eggs. The ratio of 18S rRNA to total RNA also changed as maturation progressed, which is higher in the oocytes of the early phase of oocyte maturation (from 4 to 8 h post-treatment) and lower in the oocytes of the late phase of oocyte maturation (12 h post-treatment) and in the ovulated eggs (Fig. 4A). When normalized to the amount of 18 S rRNA, the relative intensity of hybrid-ization signals of cathepsin Z transcripts increased as maturation progressed and reached peak (at 8 h post-treatment) by 9-folds compared to that of untreated oocytes, then declined near to the level of untreated oocytes in ovulated eggs (Fig. 4C).
3.4. Expression of recombinant cathepsin Z
In order to obtain a functional cathepsin Z, the coding sequence of the predicted mature enzyme was fused to glutathione S-transferase (GST) cDNA and expressed in E. coli. The recombinant protein was efficiently induced with 0.1 mM IPTG at 37 °C (Fig. 5A), but virtually all the protein thus produced was insoluble and could not be renatured. When cultured at 28 °C, a
Fig. 5. Purification of recombinant cathepsin Z. (A) Induction of recombinant protein was performed by adding 0.1 mM IPTG to the bacteria cultured at 28 °C for 10 h. The bacterial extracts were analyzed on a 12% SDS-PAGE gel and stained with Coomassie brilliant blue. Lane 1, host with vector cDNA encoding glutathione S-transferase (GST); lane 2, host with recombinant cDNA encoding the fusion protein of GST and cathepsin Z (GST-CTPZ). (B) Purification of the recombinant cathepsin Z by a Mono Q anion-exchange chromatography. The details of purification procedure are described in Materials and methods. (C) Electrophoresis. Three microgram of purified recombinant cathepsin Z was analyzed on a 12% SDS-PAGE gel.
Fig. 6. Immunohistochemical detection of cathepsin Z in the unfertilized and fertilized carp egg. Cathepsin Z is localized in cortical granules (CG) and the cytoplasm surrounding the yolk granules (YG) of unfertilized egg (A). (B) Higher magnification of the yolk granules in (A). After fertilization, the cathepsin Z stored in the cortical granules was discharged into plasma membrane (PM), perivitelline space (PVS), and fertilization envelope (FE) yet still associated with yolk granules (C). (D) Higher magnification of the boxed area in (C). CH, Chorion. Bar represents 45 µm in (A) and 300 µm in (C), respectively.
moderate amount of recombinant protein was expressed, in which about 3% recombinant protein was in soluble form. The soluble GST-CTPZ was affinity purified by a GSTrap™ FF column, then by a HPLC Mono Q HR 5/5 column (Fig. 5B). The protein appeared as a single band of 55 kDa as analyzed by SDS-PAGE (Fig. 5C).
3.5. Subcellular localization of cathepsin Z in oocytes The localization of cathepsin Z in carp ovary was examined by immunohistochemistry. Only the oocytes showed positive signals. Inside the oocyte, cathepsin Z was localized in cortical granules (Fig. 6A) and in the cytoplasm surrounding the yolk granules (Fig. 6B). During cortical reaction induced either by fertilization or by immersing eggs in hypotonic solution, the cortical granular cathepsin Z was discharged to the plasma membranes and the perivitelline space (Fig. 6C). The cathepsin Z in the perivitelline space was then gradually recruited to the outmost layer of chorion to become new components of fertilization envelope (Fig. 6D). Note that cathepsin Z remained associated with the yolk granules of cortical-reacted eggs. 3.6. Enzymatic assay of cathepsin Z
Zymography was employed to examine whether the recombi-nant cathepsin Z has enzymatic activity. As shown inFig. 7A, the recombinant cathepsin Z had proteolytic activity toward gelatin. Because cathepsins were suggested to be involved in yolk metabolism, therefore the proteolytic activity of cathepsin Z on vitellogenin was examined. The carp vitellogenin prepared in this study had a major band of 200 kDa and several minor bands of lower molecular weights in SDS-PAGE under reducing condition.
When treated with recombinant cathepsin Z, the major band of vitellogenin disappeared, whereas the minor bands were not affected. The molecular weights of the cleavage products were 95 and 87 kDa, respectively. The enzymatic activity of recombinant cathepsin Z was inhibited by PMSF or E-64, a specific inhibitor of thiol protease (Fig. 7B), but not by pepstatin, an inhibitor of aspartic protease (data not shown). These results indicate that carp cathepsin Z is a protease of papain family.
4. Discussion
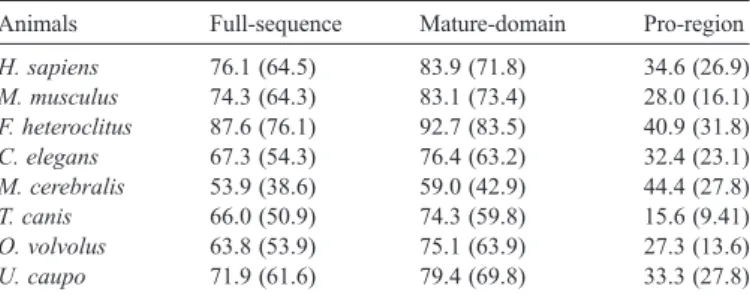
A full-length cDNA encoding carp cathepsin Z was obtained in this study. Its amino acid sequence is homologous to the cathepsin Z of other species (Table 2) and other types of cathepsins. Phylogenetic analysis reveals that cathepsin Z is more closely related to cathepsin W and B than to cathepsin S, K, L, and H (data not presented). Moreover, the fusion protein of carp cathepsin Z and glutathione S-transferase had proteolytic activity toward gelatin and vitellogenin (Fig. 7A) that could be inhibited by E-64 and PMSF (Fig. 7B) but not by pepstatin (data not shown). E-64 is an inhibitor of the protease of papain family, PMSF could inactivate the enzyme that comprises serine or cysteine as a residue of active site, and pepstatin is an inhibitor of aspartic proteinases. These results indicate that the cDNA we cloned did encode a protease belonging to papain family.
Carp cathepsin Z shares several common features with the cathepsin Z of other species. These include (1) The cleavage site of mature protein at Leu54 that generates an active enzyme of 28 kDa (Santamaria et al., 1998b), (2) The residues of active site, Cys84, His233and His254, are conserved in all cathepsins and other proteases of papain family, (3) The amino acid residues important for protein folding and enzymatic activity are conserved in the sequences of cathepsin Z of all species. The prepro-enzyme contains twelve cysteine residues, in which one residue is located in the pro-peptide of procathepsin Z, whereas the other eleven residues are located in the mature protein of the cathepsin Z of human, mouse, killifish and carp (Santamaria et al., 1998b; Deussing et al., 2000; Fabra and Cerda, 2004). Six disulfide bonds are formed in human procathepsin Z (Sivaraman et al., 2000). Of these six bonds, two (Cys81–Cys124 and Cys118–Cys156) are structurally conserved in human cathepsin B, H, K, L, papain and caricain, a third (Cys146–Cys162) matches a disulfide bond in cathepsin B, while the remaining are unique to procathepsin Z. According to the resolution of crystal structure of human
Fig. 7. Enzymatic assay of recombinant cathepsin Z. (A) Zymography. One-tenth microgram of recombinant cathepsin Z was loaded on a 10% gel containing 0.1% of gelatin. (B) Enzymatic activity of recombinant cathepsin Z (CTPZ) on vitellogenin. Ten microgram crude vitellogenin was incubated under various conditions for 2 h at 37 °C and then analyzed on a 8% SDS-PAGE gel. The amount of reagent used for assay was 0.1 µg for CTPZ, 1 mM for PMSF, and 5 µM for E-64, respectively.
Table 2
Percent homology in different regions of carp procathepsin Z with other species Animals Full-sequence Mature-domain Pro-region H. sapiens 76.1 (64.5) 83.9 (71.8) 34.6 (26.9) M. musculus 74.3 (64.3) 83.1 (73.4) 28.0 (16.1) F. heteroclitus 87.6 (76.1) 92.7 (83.5) 40.9 (31.8) C. elegans 67.3 (54.3) 76.4 (63.2) 32.4 (23.1) M. cerebralis 53.9 (38.6) 59.0 (42.9) 44.4 (27.8) T. canis 66.0 (50.9) 74.3 (59.8) 15.6 (9.41) O. volvolus 63.8 (53.9) 75.1 (63.9) 27.3 (13.6) U. caupo 71.9 (61.6) 79.4 (69.8) 33.3 (27.8) Similarity and identity (in parentheses) were calculated using EMBOSS pairwise alignment algorithms at EBI.
cathepsin Z, the cysteine residue (Cys27) in the pro-peptide contributes to the inhibition of enzymatic activity by forming a disulfide bond with the cysteine (Cys84) located in the active site (Sivaraman et al., 2000). Thus, the removal of the cysteine residue from the pro-peptide region is required for the activation of cathepsin Z. Such a mechanism offers explanation that human procathepsin Z cannot be autoactivated by acid treatment as observed in other types of cathepsins. This situation may also occur in carp cathepsin Z. Consistent with the above notion, we found that the fusion protein containing the full length of carp procathepsin Z had no enzymatic activity (data not shown) while the fusion protein containing the mature cathepsin Z did show enzymatic activity (Fig. 7).
In oviparous animals, synthesis, accumulation, storage, and utilization of yolk is critical to embryonic development. Many types of cathepsins including cathepsin B, F, and L were reported to be involved in yolk metabolism. In killifish, cathepsin Z was constitutively expressed and its expression remained almost unchanged during vitellogenesis and oocyte maturation (Fabra and Cerda, 2004). In the present study, we found that cathepsin Z was associated with yolk granules (Fig. 6) and had proteolytic activity on vitellogenin (Fig. 7B). In addition, the mRNA of cathepsin Z was transiently accumulated during the maturation of carp oocytes (Fig. 4). These results suggest that cathepsin Z may be involved in yolk metabolism in carp oocytes, at least during oocyte maturation. In killifish, a transient accumulation of the mRNAs of cathepsins L, S, H, and F were also observed during oocyte maturation (Fabra and Cerda, 2004). Apparently, oocyte maturation-associated yolk metabolism is a complicated and well-coordinated process in which multiple cathepsins including cathepsin Z are involved. Yet, the mechanism is totally unknown. Apparently, more studies are needed to gain the detail of the processing of fish yolk proteins.
In addition to yolk granules, cathepsin Z is also present in the cortical granules of carp eggs. The cortical granular cathepsin Z was discharged to the plasma membrane and the perivitelline space after fertilization. The cathepsin Z was then gradually recruited from the perivitelline space to the chorion to become new components of fertilization envelope but remained bound to the plasma membrane (Fig. 6). The functions of cathepsin Z present in the plasma membrane, perivitelline space, and fertilization envelope are unknown and remain speculative. In sea urchin, a cortical granule serine protease (CGSP1) discharged during fertilization was proposed for the cleavage of sperm receptors to prevent polyspermy and for the dissolution of the proteins connecting the plasma membrane and the vitelline membrane to elevate the fertilization envelope (Haley and Wessel, 1999). Whether the cathepsin Z in the cortical granules has similar function as the CGSP1 in sea urchin is deserved for further studies. Several components of carp cortical granules were found to be released to the perivitelline space and then added to the fertilization envelope. They are cystatin, fibroin-like substance, and cathepsin B-like substance (Chang et al., 1998). They were cross-linked by transglutaminase to form high-molecular weight conjugates. Once added to the fertilization envelope, the fertilized eggs acquired adhesiveness (Chang and Huang, 2002). Whether cathepsin Z was conjugated with these three
substances had not investigated yet. It is highly probable because cystatin is an inhibitor and can bind proteases of the papain family. If this is the case, cathepsin Z may also act as a structural unit for the generation of the adhesiveness in the fertilized eggs of carp.
Different animals apply different measures to prevent poly-spermy. In carp, cystatin had been demonstrated to agglutinate sperm by electrostatic interaction (Wang and Huang, 2002). In the perivitelline space of eggs, cystatin, fibroin-like substance, cathepsin B-like substance, and cathepsin Z (Fig. 6) appeared in a pattern of meshwork probably due to conjugation with each other. The meshwork organization of cystatin may be more efficient than the free form of cystatin to trap sperm. It is therefore assumed that cathepsin Z may play some role in preventing polyspermy through its interaction with cystatin and other cortical granular substances.
In conclusion, cathepsin Z may have different functions in eggs. None is clearly defined yet. More studies are needed to gain insight into the biological significance of cathepsin Z. References
Aranishi, F., 1999. Lysis of pathogenic bacteria by epidermal cathepsin L and B in the Japanese eel. Fish Physiol. Biochem. 20, 37–41.
Azaryan, A.V., Hook, V.Y., 1994. Unique cleavage specificity of‘prohormone thiol protease’ related to proenkephalin processing. FEBS Lett. 341, 197–202. Banon, P., Brandes, D., Frost, J.K., 1964. Lysosomal enzymes in the rat ovary
and endometrium during the estrous cycle. Acta Cytol. 8, 416–425. Brooks, S., Tyler, C.R., Carnevali, O., Coward, K., Sumpter, J.P., 1997. Molecular
characterisation of ovarian cathepsin D in the rainbow trout, Oncorhynchus mykiss. Gene 201, 45–54.
Carnevali, O., Carletta, R., Cambi, A., Vita, A., Bromage, N., 1999. Yolk formation and degradation during oocyte maturation in seabream Sparus aurata: involvement of two lysosomal proteinases. Biol. Reprod. 60, 140–146. Cataldo, A.M., Paskevich, P.A., Kominami, E., Nixon, R.A., 1991. Lysosomal
hydrolases of different classes are abnormally distributed in brains of patients with Alzheimer disease. Proc. Natl. Acad. Sci. U. S. A. 88, 10998–11002. Chan, S.J., San, S.B., McCormick, M.B., Steiner, D.F., 1986. Nucleotide and
predicted amino acid sequences of cloned human and mouse preprocathep-sin B cDNAs. Proc. Natl. Acad. Sci. U. S. A. 33, 7721–7725.
Chang, Y.S., Huang, F.L., 2002. Fibroin-like substance is a major component of the outer layer of fertilization envelope via which carp egg adheres to the substratum. Mol. Reprod. Dev. 62, 397–406.
Chang, Y.S., Wang, S.C., Tsao, C.C., Huang, F.L., 1996. Molecular cloning, structural analysis, and expression of carp ZP3 gene. Mol. Reprod. Dev. 44, 295–304.
Chang, Y.S., Weng, J.W., Li, C.C., Huang, F.L., 1998. Identification of cystatin as a component of carp chorion. Mol. Reprod. Dev. 51, 430–435. Deussing, J., von Olshausen, I., Peters, C., 2000. Murine and human cathepsin Z:
cDNA-cloning, characterization of the genes and chromosomal localization. Biochim. Biophys. Acta 1491, 93–106.
Erickson, A., 1989. Biosynthesis of lysosomal endopeptidases. J. Cell. Biochem. 40, 31–41.
Esch, F.S., Mason, A.J., Cooksey, K., Mercado, M., Shimasaki, S., 1986. Cloning and DNA sequence analysis of the cDNA for the precursor of the beta chain of bovine follicle stimulating hormone. Proc. Natl. Acad. Sci. U. S. A. 83, 6618–6621.
Esser, R.E., Watts, L.M., Angelo, R.A., Thornburg, L.P., Prior, J.J., Palmer, J.T., 1993. The effects of fluoromethyl ketone inhibitors of cathepsin B on adjuvant induced arthritis. J. Rheumatol. 20, 1176–1183.
Esser, R.E., Angelo, R.O., Murphey, M.D., Watts, L.M., Thornburg, L.P., Palmer, J.T., Talhouk, J.W., Smith, R.E., 1994. Cysteine proteinase inhi-bitors decrease articular cartilage and bone destruction in chronic inflam-matory arthritis. Arthritis Rheum. 37, 236–247.
Fabra, M., Cerda, J., 2004. Ovarian cysteine proteinases in the teleost Fundulus heteroclitus: molecular cloning and gene expression during vitellogenesis and oocyte maturation. Mol. Reprod. Dev. 67, 282–294.
Falcone, F.H., Tetteh, K.K., Hunt, P., Blaxter, M.L., Loukas, A., Maizels, R.M., 2000. The new subfamily of cathepsin-Z-like protease genes includes Tc-cpz-1, a cysteine protease gene expressed in Toxocara canis adults and infective stage larvae. Exp. Parasitol. 94, 201–207.
Fuchs, R., Gassen, H.G., 1989. Nucleotide sequence of human preprocathepsin H, a lysosomal cysteine proteinase. Nucleic Acids Res. 17, 9471. Gazith, J., Schulze, I.T., Gooding, R.H., Womack, F.C., Colowick, S.P., 1968.
Multiple forms and subunits of yeast hexokinase. Ann. N. Y. Acad. Sci. 151, 307–331.
Gething, M.J., Davidson, B.E., 1976. Chorismate mutase/prephenate dehydra-tase from Escherichia coli K12. 2. Evidence for identical subunits catalysing the two activities. Eur. J. Biochem. 71, 327–336.
Haley, S.A., Wessel, G.M., 1999. The cortical granule serine protease CGSP1 of the sea urchin, Strongylocentrotus purpuratus, is autocatalytic and contains a low-density lipoprotein receptor-like domain. Dev. Biol. 211, 1–10. Hashimoto, Y., Kakegawa, H., Narita, Y., Hachiya, Y., Hayakawa, T., Kos, J., Turk,
V., Katunuma, N., 2001. Significance of cathepsin B accumulation in synovial fluid of rheumatoid arthritis. Biochem. Biophys. Res. Commun. 283, 334–339. Hashmi, S., Zhang, J., Oksov, Y., Lustigman, S., 2004. The Caenorhabditis elegans cathepsin Z-like cysteine protease, Ce-CPZ-1, has a multifunctional role during the worms' development. J. Biol. Chem. 279, 6035–6045. Inaoka, T., Bilbe, G., Ishibashi, O., Tezuka, K., Kumegawa, M., Kokubo, T.,
1995. Molecular cloning of human cDNA for cathepsin K: novel cysteine proteinase predominantly expressed in bone. Biochem. Biophys. Res. Commun. 206, 89–96.
Kalsheker, N.A., Deam, S., Chambers, L., Sreedharan, S., Brocklehurst, K., Lomas, D.A., 1996. The house dust mite allergen Der p1 catalytically inactivates alpha 1-antitrypsin by specific reactive centre loop cleavage: a mechanism that pro-motes airway inflammation and asthma. Biochem. Biophys. Res. Commun. 221, 59–61.
Katunuma, N., Kominami, E., 1987. Abnormal expression of lysosomal cysteine proteinases in muscle wasting diseases. Rev. Physiol., Biochem. Pharmacol. 108, 1–20.
Keene, W.F., Petitt, M.G., Allen, S., McKerrow, J.H., 1986. The major neutral proteinase of Entamoeba histolytica. J. Exp. Med. 163, 539–549. Kelley, G.O., Adkison, M.A., Leutenegger, C.M., Hedrick, R.P., 2003. Myxobolus
cerebralis: identification of a cathepsin Z-like protease gene (MyxCP-1) ex-pressed during parasite development in rainbow trout, Oncorhynchus mykiss. Exp. Parasitol. 105, 201–210.
Kirschke, H., Wiederanders, B., 1987. Lysosomal proteinases. Acta Histochem. 82, 2–4.
Kirschke, H., Wiederanders, B., Brommed, D., Rinne, A., 1989. Cathepsin S from bovine spleen. Purification, distribution, intracellular localization and action on proteins. Biochem. J. 264, 467–473.
Laemmli, U., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680–685.
LaFleur Jr., G.J., Raldua, D., Fabra, M., Carnevali, O., Denslow, N., Wallace, R.A., Cerda, J., 2005. Derivation of major yolk proteins from parental vitellogenins and alternative processing during oocyte maturation in Fundulus heteroclitus. Biol. Reprod. 73, 815–824.
Linnevers, C., Smeekens, S.P., Bromme, D., 1997. Human cathepsin W, a putative cysteine protease predominantly expressed in CD8+ T-lymphocytes. FEBS Lett. 405, 253–259.
Lustigman, S., Zhang, J., Liu, J., Oksov, Y., Hashmi, S., 2004. RNA interference targeting cathepsin L and Z-like cysteine proteases of Onchocerca volvulus confirmed their essential function during L3 molting. Mol. Biochem. Parasitol. 138, 165–170.
Lynn, K.R., Brockbank, W.J., Clevette, N.A., 1980. Multiple forms of the asclepains. Cysteinyl proteases from milkweed. Biochim. Biophys. Acta 612, 119–125.
Mason, R.W., Johnson, D.A., Barrett, A.J., Chapman, H.A., 1986. Elastinolytic activity of human cathepsin L. Biochem. J. 233, 925–927.
Mommsen, T.P., 2004. Salmon spawning migration and muscle protein metabolism: the August Krogh principle at work. Comp. Biochem. Physiol. B 139, 383–400.
Mommsen, T.P., French, C.J., Hochachka, P.W., 1980. Sites and pattern of protein and amino acid utilization during the spawning migration of salmon. Can. J. Zool. 58, 1785–1799.
Morton, P.A., Zacheis, M.L., Giacoletto, K.S., Manning, J.A., Schwartz, B.D., 1995. Delivery of nascent MHC class II-invariant chain complexes to lyso-somal compartments and proteolysis of invariant chain by cysteine proteases precedes peptide binding in B-lymphoblastoid cells. J. Immunol. 154, 137–150.
Nagler, D.K., Menard, R., 1998. Human cathepsin X: a novel cysteine protease of the papain family with a very short proregion and unique insertions. FEBS Lett. 434, 135–139.
Nagler, D.K., Zhang, R., Tam, W., Sulea, T., Purisima, E.O., Menard, R., 1999. Human cathepsin X: a cysteine protease with unique carboxypeptidase activity. Biochemistry 38, 12648–12654.
Nagler, D.K., Kruger, S., Kellner, A., Ziomek, E., Menard, R., Buhtz, P., Krams, M., Roessner, A., Kellner, U., 2004. Up-regulation of cathepsin X in prostate cancer and prostatic intraepithelial neoplasia. Prostate 60, 109–119. Opresko, L.K., Karpf, R., 1987. Specific proteolysis regulates fusion between
endocytic compartments in Xenopus oocytes. Cell 51, 557–568.
Paris, A., Strukelj, B., Pungercar, J., Renko, M., Dolenc, I., Turk, V., 1995. Molecular cloning and sequence analysis of human preprocathepsin C. FEBS Lett. 369, 326–330.
Rautenberg, P., Schadler, R., Reinwald, E., Risse, H.J., 1982. Study on a proteolytic enzyme from Trypanosoma congolense. Purification and some biochemical properties. Mol. Cell. Biochem. 47, 151–159.
Retzek, H., Steyrer, E., Sanders, E.J., Nimpf, J., Schneider, W.J., 1992. Molecular cloning and functional characterization of chicken cathepsin D, a key enzyme for yolk formation. DNA Cell Biol. 11, 661–672.
Riese, R.J., Wolf, P.R., Bromme, D., Natkin, L.R., Villadangos, J.A., Ploegh, H.L., Chapman, H.A., 1996. Essential role for cathepsin S in MHC class II-associated invariant chain processing and peptide loading. Immunity 4, 357–366. Riggio, M., Scudiero, R., Filosa, S., Parisi, E., 2002. Oestrogen-induced
expression of a novel liver-specific aspartic proteinase in Danio rerio (zebrafish). Gene 295, 241–246.
Sanger, F., Nicklen, S., Coulson, A.R., 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U. S. A. 78, 454–458. Santamaria, I., Velasco, G., Cazorla, M., Fueyo, A., Campo, E., Lopez-Otin, C.,
1998a. Cathepsin L2, a novel human cysteine proteinase produced by breast and colorectal carcinomas. Cancer Res. 58, 1624–1630.
Santamaria, I., Velasco, G., Pendas, A.M., Fueyo, A., Lopez-Otin, C., 1998b. Cathepsin Z, a novel human cysteine proteinase with a short propeptide domain and a unique chromosomal location. J. Biol. Chem. 273, 16816–16823. Shi, G.P., Munger, J.S., Meara, J.P., Rich, D.H., Chapman, H.A., 1992.
Molecular cloning and expression of human alveolar macrophage cathepsin S, an elastinolytic cysteine protease. J. Biol. Chem. 267, 7258–7262. Sivaraman, J., Nagler, D.K., Zhang, R., Menard, R., Cygler, M., 2000. Crystal
structure of human procathepsin X: a cysteine protease with the proregion covalently linked to the active site cysteine. J. Mol. Biol. 295, 939–951. Sloane, B.F., Honn, K.V., 1984. Cysteine proteinases and metastasis. Cancer
Metastasis Rev. 3, 249–263.
Tahtinen, V., Weber, E., Gunther, D., Ylonen, A., Kalkkinen, N., Olsen, R., Jarvinen, M., Soderstrom, K.O., Rinne, A., Bjorklund, H., Bogwald, J., 2002. Immunolocalization of cysteine proteinases (cathepsins) and cysteine proteinase inhibitors (salarin and salmon kininogen) in Atlantic salmon, Salmo salar. Cell Tissue Res. 310, 213–222.
Tezuka, K., Tezuka, Y., Maejima, A., Sato, T., Nemoto, K., Kamioka, H., Hakeda, Y., Kumegawa, M., 1994. Molecular cloning of a possible cysteine proteinase predominantly expressed in osteoclasts. J. Biol. Chem. 269, 1106–1109. Tisljar, K., Deussing, J., Peters, C., 1999. Cathepsin J, a novel murine cysteine
protease of the papain family with a placenta-restricted expression. FEBS Lett. 459, 299–304.
Troen, B.R., Gal, S., Gottesman, M.M., 1987. Sequence and expression of the cDNA for MEP (major excreted protein), a transformation-regulated secreted cathepsin. Biochem. J. 246, 731–735.
Uinuk-ool, T.S., Takezaki, N., Kuroda, N., Figueroa, F., Sato, A., Samonte, E., Mayer, W.E., Klein, J., et al., 2003. Phylogeny of antigen-processing enzymes: cathepsins of a cephalochordate, an agnathan and a bony fish. Scand. J. Immunol. 58, 436–448.
Velasco, G., Ferrando, A.A., Puente, X.S., Sanchez, L.M., Lopez-Otin, C., 1994. Human cathepsin O. Molecular cloning from a breast carcinoma, production of the active enzyme in Escherichia coli, and expression analysis in human tissues. J. Biol. Chem. 269, 27136–27142.
Vernet, T., Tessier, D.C., Richardson, C., Laliberte, F., Khouri, H.E., Bell, A.W., Storer, A.C., Thomas, D.Y., 1990. Secretion of functional papain precursor from insect cells. Requirement for N-glycosylation of the pro-region. J. Biol. Chem. 265, 16661–16666.
Villadangos, J.A., Riese, R.J., Peters, C., Chapman, H.A., Ploegh, H.L., 1997. Degradation of mouse invariant chain: roles of cathepsins S and D and the influence of major histocompatibility complex polymorphism. J. Exp. Med. 186, 549–560.
von der Decken, A., 1992. Physiological changes of skeletal muscle by maturation-spawning of non-migrating female Atlantic salmon, Salmo salar. Comp. Biochem. Physiol. B 101, 299–301.
Wallace, R.A., Selman, K., 1985. Major protein changes during vitellogenesis and maturation of Fundulus oocytes. Dev. Biol. 110, 492–498.
Wang, S.C., Huang, F.L., 2002. Carp ovarian cystatin binds and agglutinates spermatozoa via electrostatic interaction. Biol. Reprod. 66, 1318–1327. Wang, B., Shi, G.P., Yao, P.M., Li, Z., Chapman, H.A., Bromme, D., 1998. Human
cathepsin F. Molecular cloning, functional expression, tissue localization, and enzymatic characterization. J. Biol. Chem. 273, 32000–32008.
Yamashita, M., Konagaya, S., 1990. High activities of cathepsin B, D, H, and L in the white muscle of chum salmon in spawning migration. Comp. Biochem. Physiol. B 95, 149–152.
Yamashita, M., Konagaya, S., 1991. Increase in catheptic activity and appear-ance of phagocytes in the white muscle of chum salmon during spawning migration. Biomed. Biochem. Acta 50, 565–567.
Yoshizaki, N., Moriyama, A., Yoneyawa, S., 1998. Purification and properties of embryonic cysteine proteinase which participates in yolk-lysis of Xenopus laevis. Comp. Biochem. Physiol. B 119, 571–576.