234
*To whom correspondence and reprint requests should be addressed.
D
iurnal raptors often consume prey large enough to be identified at a distance. Yet, study-ing raptor diets can be difficult and time-consum-ing given the large home ranges, low population densities, and low rates of prey capture in many species. Some raptor species use regular pluck-ing sites where they remove the feathers or other-wise handle their prey before consuming it (Newton 1986, Selås 1993, Watson et al. 1998). The diet of such species can be studied by moni-toring the plucking sites. However, not all raptor species have regular plucking sites, and many species pluck their prey near the site of prey cap-ture, resulting in widely dispersed sites. Accipitersin Taiwan are not known to have regular plucking sites. The frequent rain and high humidity in Taiwan would also quickly alter the appearance of a plucking site, greatly reducing the probability of detecting such a site. Thus, studying prey provi-sioning to nestlings is perhaps the most manage-able and useful approach to gaining a basic under-standing of accipiters in Taiwan. Direct observa-tion of prey capture or consumpobserva-tion (e.g., Collopy 1983, Newton 1986, Simmons et al. 1991), collect-ing prey remains (e.g., Newton 1986, Joy et al. 1994, Mañosa 1994, Toyne 1998, Thrailkill et al. 2000, Rutz 2003), and collecting pellets (e.g., Mañosa 1994, Toyne 1998, Sharp et al. 2002) all
Comparison of Three Common Methods for Studying the Diet of
Nestlings in Two Accipiter Species
Kuang-Ying Huang1, Yao-Sung Lin1, and Lucia Liu Severinghaus2,*
1Institute of Ecology and Evolutionary Biology, National Taiwan University, Taipei, Taiwan 106, R.O.C. E-mail: [email protected]
2Research Center for Biodiversity, Academia Sinica, Taipei, Taiwan 11528, R.O.C. E-mail: [email protected]
(Accepted July 15, 2005)
Kuang-Ying Huang, Yao-Sung Lin, and Lucia Liu Severinghaus (2006) Comparison of three common
meth-ods for studying the diet of nestlings in two Accipiter species. Zoological Studies 45(2): 234-243. Food provi-sioning to 4 Accipiter virgatus nests and 2 A. trivirgatus nests was monitored during 1998 and 2000 in Yangmingshan National Park, Taipei, Taiwan. We made direct observations of prey deliveries and collected prey remains and pellets throughout the breeding seasons in order to compare the effectiveness and efficiency of the different diet analysis methods for these 2 raptor species. The method of using prey remains worked best for A. virgatus for all measurements, producing 45.23% of all prey items found, 45.23% of all prey items identified to prey taxa, and 38.87% of all prey items identified to species. The observation method worked best for A. trivirgatus, providing 41.05% of all prey items and 18.95% of prey items identified to species, but the pro-portion of prey items identified to prey taxa was highest using the pellet method (37.37%). Time efficiency was consistently the highest for the prey remains method in A. virgatus. In contrast, for A. trivirgatus, the pellet method was most efficient for assessing the number of prey items delivered and the number identified to prey taxa, but the prey remains method worked best for identifying prey to species. Combining the prey remains and pellet methods increased the prey taxa identified to 81.63% for A. virgatus and 58.95% for A. trivirgatus. Although the value of direct observation has been stressed by many researchers, its effectiveness varied between these 2 species studied in Taiwan. http://www.sinica.edu.tw/Journals/45.2/234.pdf
entail different time and energy demands. Each method requires researchers to overcome different difficulties including preventing disturbance to par-ents and young, cleaning and identifying fragmpar-ents of prey, and accessing raptor nests positioned high above the ground. Pellet analysis is frequently used for owls because owls usually swallow prey whole or in large fragments and leave less-identifi-able remains (Marti 1987). Owls are also noctur-nal which makes prey identification by observation difficult without sophisticated photographic equip-ment. The usefulness of the pellet method for diurnal raptors is limited by the greater ability of the Falconiformes to digest bones (Reynolds and Meslow 1984). However, tools to examine the micromorphological features of hair and feathers have increased the range of prey items identifiable from pellets (Squires 2000). Direct observation is rarely used because it is highly time-consuming. In contract, analyzing prey remains has been wide-ly used to assess the diets of raptors. In recent years, videography has replaced direct observa-tions (Rutz 2003, Lewis et al. 2004), but this method is very expensive and also time-intensive (Booms and Fuller 2003).
Traditional methods of diet analysis (Collopy 1983, Simmons et al. 1991, Bosakowski and Smith 1992, Sharp et al. 2002, Rutz 2003, Lewis et al 2004) have been evaluated to some extent for temperate raptors. Comparisons between pellet analysis, prey remains analysis, and direct obser-vations are available only for the Hen Harrier (Circus cyaneus; Redpath et al. 2001) Bonelli,s Eagle (Hieraaetus fascistus; Real 1996), and Northern Goshawk (A. gentilis, Lewis et al. 2004, who substituted videos for direct observations). These comparisons concluded that no one method was sufficient to cover the entire diet of a raptor species. Biases varied with prey size, prey taxon, the frequency and intervals of observation, prey remains, and pellet collection.
Given the differences in temperature, precipi-tation, vegetation structure, foliar density, speed of decomposition, and abundance of ants between the subtropics and temperate zone, we decided to evaluate the usefulness of these methods for rap-tors in Taiwan. We compared the relative effi-ciencies and accuracies of direct observation, prey remains analysis, and pellet analysis for 2 Accipiter species breeding in Yangmingshan near Taipei. Accipiter virgatus regularly preys upon medium-sized birds, insects, and mammals (Huang et al. 2004). Accipiter trivirgatus con-sumes reptiles, rats and other mammals, and
medium to large birds (Huang unpublished data). Both species usually process bird and mammal prey before consuming it by plucking the items. With reptiles and insects, A. trivirgatus often swal-lows them whole, whereas A. virgatus usually processes them first. The findings of this study should be of value to future diet studies on sub-tropical raptors.
MATERIALS AND METHODS Species description and study area
Accipiter virgatus and A. trivirgatus are similar in appearance, but differ in size (A. virgatus female 224.4 ± 14.3 g, male 124.2 ± 6.0 g; A. trivirgatus female 563.0 ± 29.8 g, male 352.3 ± 13.9 g). These species are difficult to tell apart in flight, especially at a distance. The 2 species are sym-patric in the lowlands of Taiwan. Normal clutch size is 3-4 eggs for A. virgatus and 2 for A. trivirga-tus. Accipiter virgatus usually nests in woods with dense vegetation on gentle slopes, while A. trivir-gatus nests either in relatively open woods on fair-ly steep slopes or in open vegetation on gentle slopes.
We studied these 2 species in the Yang-mingshan region (25
°
10'N, 121°
33'E), at 350-750 m in elevation. Annual rainfall averages 3000 mm at low elevations and up to 4500 mm at higher ele-vations. The study area was covered with a native evergreen broadleaf forest and some introduced conifer plantations, containing more than 1224 species of plants belonging to 170 families, includ-ing at least 2 species of gymnosperms, 294 species of monocotyledons, 747 species of dicotyledons, and 181 species of ferns. Dominant woody plants include Trochodendron aralioides, Machilus thunbergii, M. kusanoi, Prunus phaeosticta, Itea parviflora, Pseudosasa usawai, Cleyera japonica, and Hydrangea angustipetala. In both accipiter nesting areas, the dominant tree species were Cryptomeria japonica, Machilus thunbergii, Acacia confusa, Meliosma rhoifolia, Weldlandia formosana, Turpinia formosana, Bischofia javanica, and Cleyera japonica.Field data collection
We collected data on food provisioning to nestlings between May and July 1998 and 2000 at 4 A. virgatus and 2 A. trivirgatus nests, using all 3 methods described below. We searched for prey
remains and pellets upon arrival at the nests early in the morning; we then observed food provision-ing by the parents for the remainder of the day. Details of the methods are given here.
1. Direct observation: We built a blind at a location which afforded a clear lateral or downward view of the nest. We observed nest provisioning with a spotting scope (25 x 77 power) and binocu-lars (8 x 30 power). We spent 4 to 23 d (at least 5 h/d) observing each nest per year. We recorded prey transfers from male to female, the number and type of prey delivered, and hunting attempts by the fledglings. We made an effort to identify all prey items to class or species, and the size, color, and shape of each item were noted when possible. 2. Collection and identification of prey remains: We strung a 10 x 10 m net with a 0.3 cm mesh size below each nest to capture falling food remains. Every 1-3 d throughout the breeding cycle, we spent 1 h collecting all prey remains at each nest from the net and the ground within a 50 m radius of the nest. Every 1-7 d, we spent 3 h climbing each nest tree to collect prey remains left in the nest. Because both species of accipiters continued to add fresh green nesting material to the nest before the nestlings fledged, we searched among the nest surface material for prey remains. We removed all prey remains from the collecting nets and the nests at each visit to avoid double counting.
We stored the collected prey remains from each nest separately in sealed envelopes with the date and nest number clearly marked. These envelopes were stored in a freezer after the prey items were identified. Prey remains included feathers, bones, lower jaw, teeth, fur, skin, other unconsumed portions, and insect exoskeletons. Based on the distribution of prey items in the study area and using study skins and the skeleton col-lection of Yangmingshan National Park as refer-ence material, we constructed a prey identification key. The identification key included the color and shape of the bill, tarsus, remiges, and rectrices, the color and other characteristics of body feathers of birds, special features of insect exoskeletons, the tails of mammals and lizards, and the color of mammal hair, skin, and fur, etc. Using this identifi-cation key, prey remains were identified to class or species, and the number of individuals was record-ed.
3. Pellet collection and analysis: Pellets were collected simultaneously with the prey remains. Pellets were individually stored in sealed envelopes with the date and nest number clearly
marked. Pellets were softened and washed with water, teased apart with fingers and forceps, then dried in an oven at 45
°
C (Sabo and Laybourne 1994). Pellets usually contained feathers, bones, bristles, exoskeletons of insects, and scales of rep-tiles. We visually identified the prey items within a pellet with the identification key we developed, and when necessary, we used a magnifying glass. We recorded the contents of each pellet and counted the number of pellets.Data analysis
We followed Rutz (2003) and adopted a con-servative method for counting the frequencies of prey remains. We paired body parts, plumage, pelage, and skin of the prey remains collected from the same nest on the same day to estimate the minimum number of individuals consumed. For the pellet analysis, we followed Redpath et al. (2001) and considered all parts from the same taxon in a pellet to be from a single individual, while parts in different pellets were considered to be from different individuals.
We obtained the average biomass of each prey species from museum specimens, reference books, or by weighing a few captured individuals. Prey items that could not be identified to species were identified to class and separated into the size categories of large, medium, and small.
The efficiency of each method was assessed by calculating the number of individual prey items identified per hour of field data collection effort. The method that produced the highest mean num-ber of individual prey items, prey taxa, or prey species was considered the most efficient method. The effectiveness of the methods referred to the proportion of total prey types found through using each method.
RESULTS
Of the 4 A. virgatus and 2 A. trivirgatus nests we monitored in 1998 and 2000, incubation lasted 28 d for A. virgatus and about 39 d for A. trivirga-tus. During incubation, females of both species generally left the nest to receive food from males at non-specific locations nearby and consumed it away from the nest. In both years of the study, A. virgatus nestlings began hatching in late May, and A. trivirgatus nestlings began hatching in early to mid-May. For both species, we observed very few prey deliveries and found very few prey remains or
pellets during the incubation period and the 1st week after hatching. Furthermore, we were unable to identify what was delivered in this period because the parents of both species often removed the feathers or hair before returning to the nests and fed the nestlings small pieces of meat that did not produce pellets. Young A. virga-tus fledged at 22-23 d, while young A. trivirgavirga-tus fledged at 34-36 d. After fledging, the parents con-tinued to deliver food to the nest, so the fledglings of both species remained near the nest. Thus we were able to continue collecting information on their diets for about 10 d after fledging.
Prey delivered to nestlings
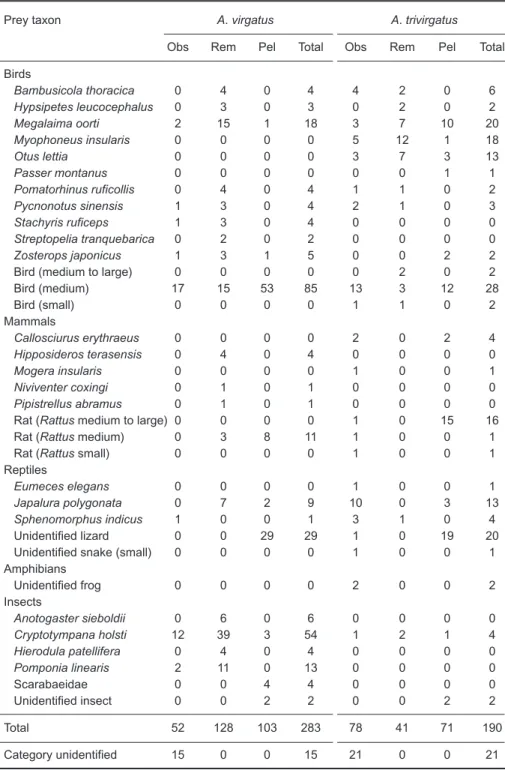
Altogether we observed 52 food deliveries in 29 d (134.5 h) for A. virgatus. During the same period, we conducted 59 searches (74 h) and found 128 prey remains and 62 pellets containing 103 individual prey items. In total, 268 individual prey items consisting of 22 prey taxa were identi-fied. Among these, 4 taxa were identified by all 3 methods, 5 were detected by 2 methods, and 1 only by observation, 9 only using prey remains and 3 only from pellet analysis (Table 1).
Altogether we observed 78 food deliveries in 46 d (323 h) for A. trivirgatus. During the same period, we conducted 44 searches (59 h) and found 41 prey remains and 34 pellets containing 71 individual prey items. In total, 169 individual prey items consisting of 26 prey taxa were identi-fied. Among these, 5 taxa were identified through all 3 methods, 8 were detected through 2 methods, and 7 only by direct observation, 2 only using prey remains, and 4 only from pellet analysis (Table 1).
Each diet analysis method detected a differ-ent proportion of individual prey items (Table 2). The prey remains method identified the greatest number of individuals for A. virgatus, while the direct observation method worked best for A. trivir-gatus (Table 2A). The prey remains method also identified the greatest proportion of individuals to taxon and species for A. virgatus (Table 2B, C). In contrast, the pellet method identified a greater pro-portion of individuals to taxon, while the observa-tion method identified a greater proporobserva-tion of indi-viduals to species for A. trivirgatus (Table 2B, C). Combining the prey remains and pellet methods identified more individuals within each category than any single diet analysis method for both A. virgatus and A. trivirgatus.
We compared the observed prey items brought back to the nest with the prey remains and
pellets found the following day. The taxa con-tained within the pellets of A. virgatus differed from what had been delivered to the nest the day before, and only 6.25% of prey types delivered the day before appeared in A. trivirgatus. About 18% of prey remains for A. virgatus were seen being delivered to the nest the day before, and only 40% of the prey remains were seen being delivered to A. trivirgatus nests. Apparently the prey types found in prey remains and pellets did not corre-spond well to those observed at the nests the day before.
Biomass delivered to nestlings
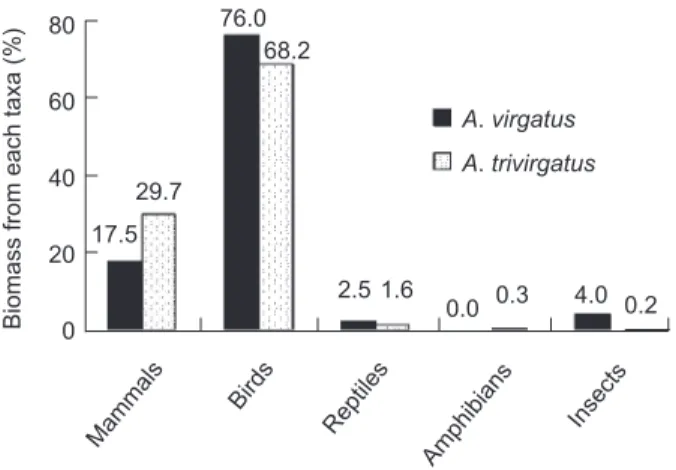
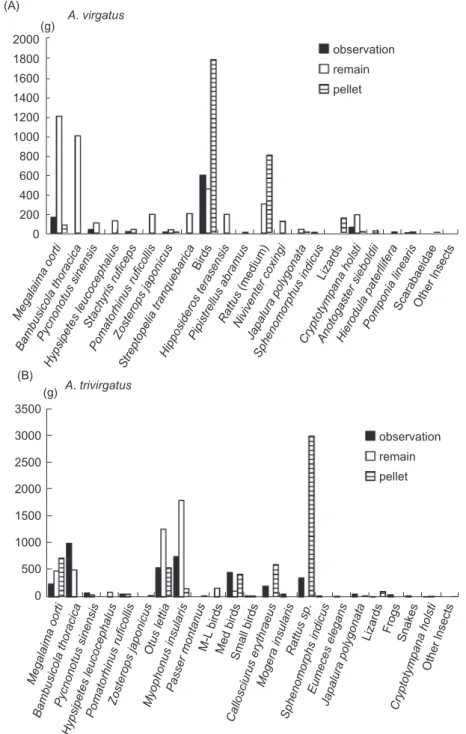
We compared the total biomass delivered to the nest by prey taxa. For A. virgatus, birds con-tributed the greatest percent of biomass followed by mammals (Fig. 1). Using pellets or direct observations alone would have underestimated the significance of certain rare but heavy bird species in A. virgatus diets, such as Megalaima oorti, Bambusicola thoracica, Pomatorhinus ruficollis, and Streptopelia tranquebarica (Fig. 2A). A species of bat (Hipposideros terasensis) was also identified only in the prey remains. Pellet analyses added to our understanding of the biomass contri-bution of prey taxa (e.g., medium-sized rodents), but not to the contribution of individual species. For A. trivirgatus, birds also contributed the great-est percent of biomass followed by mammals (Fig. 1). Although direct observation identified the greatest number of prey items to species in A. trivirgatus diets, biomass data based on direct observation alone would have either missed or underestimated the significance of Myophoneus
Biomass from each taxa (%)
Mam mal s A. virgatus A. trivirgatus Bird s Rep tiles Am phib ians Inse cts 80 60 40 20 0 76.0 68.2 29.7 17.5 2.5 1.6 0.0 0.3 4.0 0.2
Fig. 1. Relative proportion of biomass of each taxon
con-sumed by Accipiter virgatus and A. trivirgatus during 1998 and 2000 in Yangmingshan National Park, Taipei, Taiwan.
insularis, B. thoracica, M. oorti, Hypsipetes leuco-cephalus, Otus lettia, and medium to large rodents (Fig. 2B). For both Accipiter species, pellet analy-ses added to our understanding of the biomass contribution of prey taxa (e.g., medium-sized
rodents).
Efficiency comparisons
To assess the time economy of each method,
Table 1. Number of individuals per prey taxon identified using direct
obser-vation (Obs), prey remains (Rem), pellets (Pel), and all methods combined (Total) for Accipiter virgatus and A. trivirgatus during 1998 and 2000 in Yangmingshan National Park, Taipei, Taiwan
Prey taxon A. virgatus A. trivirgatus
Obs Rem Pel Total Obs Rem Pel Total
Birds Bambusicola thoracica 0 4 0 4 4 2 0 6 Hypsipetes leucocephalus 0 3 0 3 0 2 0 2 Megalaima oorti 2 15 1 18 3 7 10 20 Myophoneus insularis 0 0 0 0 5 12 1 18 Otus lettia 0 0 0 0 3 7 3 13 Passer montanus 0 0 0 0 0 0 1 1 Pomatorhinus ruficollis 0 4 0 4 1 1 0 2 Pycnonotus sinensis 1 3 0 4 2 1 0 3 Stachyris ruficeps 1 3 0 4 0 0 0 0 Streptopelia tranquebarica 0 2 0 2 0 0 0 0 Zosterops japonicus 1 3 1 5 0 0 2 2
Bird (medium to large) 0 0 0 0 0 2 0 2
Bird (medium) 17 15 53 85 13 3 12 28 Bird (small) 0 0 0 0 1 1 0 2 Mammals Callosciurus erythraeus 0 0 0 0 2 0 2 4 Hipposideros terasensis 0 4 0 4 0 0 0 0 Mogera insularis 0 0 0 0 1 0 0 1 Niviventer coxingi 0 1 0 1 0 0 0 0 Pipistrellus abramus 0 1 0 1 0 0 0 0
Rat (Rattus medium to large) 0 0 0 0 1 0 15 16
Rat (Rattus medium) 0 3 8 11 1 0 0 1
Rat (Rattus small) 0 0 0 0 1 0 0 1
Reptiles
Eumeces elegans 0 0 0 0 1 0 0 1
Japalura polygonata 0 7 2 9 10 0 3 13
Sphenomorphus indicus 1 0 0 1 3 1 0 4
Unidentified lizard 0 0 29 29 1 0 19 20
Unidentified snake (small) 0 0 0 0 1 0 0 1
Amphibians Unidentified frog 0 0 0 0 2 0 0 2 Insects Anotogaster sieboldii 0 6 0 6 0 0 0 0 Cryptotympana holsti 12 39 3 54 1 2 1 4 Hierodula patellifera 0 4 0 4 0 0 0 0 Pomponia linearis 2 11 0 13 0 0 0 0 Scarabaeidae 0 0 4 4 0 0 0 0 Unidentified insect 0 0 2 2 0 0 2 2 Total 52 128 103 283 78 41 71 190 Category unidentified 15 0 0 15 21 0 0 21
we calculated the number of individual prey items identified, the number of prey identified to taxa, and the number of prey identified to species per hour of field data collection (Table 3). The prey remains method was the most efficient in all cases for A. virgatus. For A. trivirgatus, the pellet method was the most efficient for determining the number of individuals and number of prey identified to taxa, but the prey remains method worked best for iden-tifying prey to species. For both species and in each category, combining prey remains with pel-lets was the most efficient method.
We compared the physical characteristics of prey items which influenced the efficiency of each method for identifying prey to species within taxa. Species possessing special morphological charac-teristics in the exoskeleton or bones, or had col-ored feathers were usually identified by all 3 meth-ods. Direct observation identified some birds but
few mammals, reptiles, or insects to species unless they had distinct morphological traits. The prey remains method consistently identified the greatest number of individuals to species within each taxon (birds, mammals, reptiles, and insects). The pellet method provided very limited species information except when a prey item had obvious morphological features such as the coarse scales of Japalura polygonata. Most prey found in the pellets could only be identified to general taxa.
DISCUSSION
Our results showed that for both Accipiter species, different methods produced different com-binations of prey types and prey species. This finding is consistent with results for other raptors (e.g., Purger 1998, Watson et al 1998, Sharp et al.
Table 2. Comparison of the proportion of individuals identified using direct
observation, prey remains, pellets, all methods (Total), and prey remains and pellets combined (Rem + Pel) for Accipiter virgatus and A. trivirgatus during 1998 and 2000 in Yangmingshan National Park, Taipei, Taiwan
Species Observation Prey remains Pellets Total Rem + Pel
A. Percent (%) of total individuals identified by each method
A. virgatus 18.37 45.23 36.40 100.00 81.63
A. trivirgatus 41.05 21.58 37.37 100.00 58.95
B. Percent (%) of total individuals identified to prey taxon by each method
A. virgatus 13.07 45.23 36.40 94.70 81.63
A. trivirgatus 30.00 21.58 37.37 88.95 58.95
C. Percent (%) of total individuals identified to species by each method
A. virgatus 7.07 38.87 3.89 49.82 42.76
A. trivirgatus 18.95 18.42 12.11 49.47 30.53
Table 3. Prey identification efficiency of direct observation, prey remains,
pellets, and the combination of prey remains and pellets (Rem + Pel) per hour of field data collection effort for Accipiter virgatus and A. trivirgatus dur-ing 1998 and 2000 in Yangmdur-ingshan National Park, Taipei, Taiwan
Species Observation Prey remains Pellets Rem + Pel
A. Number of individual prey items identified/h of effort*
A. virgatus 0.39 1.73 1.39 3.12
A. trivirgatus 0.24 0.69 1.20 1.90
B. Number of prey items identified to taxon/h of effort
A. virgatus 0.28 1.73 1.39 3.12
A. trivirgatus 0.18 0.69 1.20 1.90
C. Number of prey items identified to species/h of effort
A. virgatus 0.15 1.49 0.15 1.64
A. trivirgatus 0.11 0.59 0.39 0.98
2002, Boom and Fuller 2003, Lewis et al 2004). Sharp et al. (2002) attributed these differences to differential digestibility, body size, and handling and consumption behaviors associated with differ-ent prey species.
Effectiveness of the methods compared
We found that for A. virgatus, all 3 methods
worked relatively well at detecting bird prey, but observation under-represented mammals and rep-tiles, whereas for A. trivirgatus, observation worked relatively well for all prey taxa.
The prey remains method under-represented reptiles in the A. virgatus diet and under-represent-ed mammals and reptiles in the A. trivirgatus diet. These prey types were mostly small in size in our study; thus although we carefully searched for prey
Fig. 2. Estimates of the total biomass contribution by prey taxa obtained using direct observation, prey remains, and pellets for A.
trivir-gatus during 1998 and 2000 in Yangmingshan National Park, Taipei, Taiwan.
Ba mb usi cola tho raci ca Me ga laim a o ort i Pyc non otu s si nen sis Hyp sip ete s le uco cep ha lus Sta chyr is r ufic ep s Po ma torh inu s ru fico llis Zo ste rop s ja po nic us Str ep top elia tra nqu eba rica Bir ds Hip posi dero s te rase nsi s Pip istr ellu s a bra mu s Niv ive nte r co xin gi Sp he nom orp hu s in dic us Cry pto tym pan a h ols ti Ra ttu s (m ed ium ) Jap alu ra p oly go nata Liz ard s An oto gast er sie bold ii Hie rod ula pa terl life ra Po mp on ia li nea ris Sca rab aeid ae Oth er lnse cts Me ga laim a o ort i Ba mb us ico la t ho rac ica Py cn on otu s s ine ns is Hy ps ipe tes le uc oc ep ha lus Po ma torh inu s r ufic olli s Zo ste rop s ja po nic us Otu s le ttia My op ho nu s i ns ula ris M-L bir ds Sm all bir ds Mo ge ra i ns ula ris Pa sse r m on tan us Me d b ird s Ca llos ciu rus ery thra eu s Ra ttu s s p. Sp he no mo rph s in dic us Eu me ce s e leg an s Ja pa lura po lyg on ata Liz ard s Fro gs Sn ak es Cry pto tym pa na ho lsti Oth er lns ec ts observation remain pellet observation remain pellet (g) (g) A. virgatus (A) (B) A. trivirgatus 2000 1800 1600 1400 1200 1000 800 600 400 200 0 3500 3000 2500 2000 1500 1000 500 0
remains each time, the probability of overlooking something inconspicuous was still higher than for larger items. Redpath et al. (2001) also discov-ered that large prey of Hen Harriers (Circus cya-neus), a raptor of comparable size to A. trivirgatus, were over-represented and small prey under-rep-resented when analyzed using prey remains. At our study site, some prey remains may have been removed by scavengers (e.g., ants and rodents), but our use of collecting nets below nest trees should have decreased the severity of this prob-lem.
Regarding the number of prey taxa or species identified per hour of effort, prey remains analysis was the most efficient single method for A. virgatus. For A. trivirgatus, prey remains worked well for prey species identification. Our analysis of the efficiency of the prey remains or pellets did not include the amount of time spent examining and identifying the prey items, because this part of the work was done at our convenience. However, if the time spent working in the laboratory were included, the efficiency of the prey remains method would have been somewhat lower.
The pellet method underestimated insects in the A. virgatus diet and birds in the A. trivirgatus diet. Similar patterns were found for the pellet analysis of Hen Harriers (Redpath et al. 2001). The pellet method worked efficiently for the num-ber of prey individuals identified and for the prey types identified per hour of effort for A. trivirgatus. If the time spent working in the laboratory were included, the efficiency of the pellet method would have been much lower.
The combination of the prey remains and pel-let methods was the most efficient in terms of the number of prey types and species identified per hour of effort.
Suitability of methods varied with species
Direct observation at the nest should provide the most accurate information on nestlings,diet, as proposed by a number of authors who considered direct observations necessary when quantifying the diet of nestlings (Marti 1987, Watson et al. 1998, Redpath et al. 2001). However, observation did not work equally well for the 2 accipiters we studied. At A. virgatus nests, observation involved difficult observation angles and long distances to the nests, as well as a larger brood size and greater visual blockage caused by the dense foliage. All these factors contributed to greater dif-ficulty in making observations of A. virgatus than of
A. trivirgatus nests.
Raptors often pluck the feathers of their prey before delivering it to the nest. This habit may reduce the ability to identify bird prey items by the observation method, and a portion of the birds we observed were plucked fairly clean. Rutz (2003) reported that Northern Goshawks plucked large prey more thoroughly than smaller prey at the cap-ture site. In our study, both species plucked their prey when their nestlings were small but did not do so after the nestlings were large enough to handle partially plucked whole prey themselves. We do not think our observation data were biased by size-determined plucking of prey.
The suitability of the prey remains and pellet methods also differed between the 2 accipiters due to differences in their body sizes, prey types, and behavior. There appeared to be a size threshold for each raptor with regard to which prey types were usually swallowed whole and which were partially left as food remains. Rodent remains were found at A. virgatus nests but never at A. trivirgatus nests, while they were found in pellets of both species. Many small bird remains were found around A. virgatus nests but there were fewer at A. trivirgatus nests, possibly because A. trivirgatus nestlings could consume small bird prey whole, leaving few remains. Accipiter trivirgatus adults often ate the prey remains left by the nestlings, including the skin and bones, or carried the prey remains away to consume it elsewhere. These habits most probably reduced the amount of prey remains found around the nest for this species. This appears to be similar to the Northern Goshawk, which was also found to remove food remains from the nest after a successful capture (Rutz 2003, Lewis et al. 2004). Unlike the Northern Goshawk (Lewis et al. 2004), A. virgatus parents rarely carried prey remains away from the nest. The larger brood size of A. virgatus probably also meant that more prey items were delivered to the nests. Both adult and juvenile A. virgatus often tossed their heads sideways as they fed on prey, increasing the chance that loose feathers and other prey fragments fell from the nest. Competition among 4 nestlings probably also caused some feathers and body parts to fall down. Overall, a higher number of prey remains were found around A. virgatus than A. trivirgatus nests.
Our pellet records should be fairly complete, because unlike other raptors, such as the Golden Eagle (Collopy 1983), neither accipiter ate or removed the pellets regurgitated by nestlings. Pellets are best at revealing small prey often
missed by the observation and food remains meth-ods, such as most mammals and reptiles.
Redpath et al. (2001) suggested that pellets were useful for estimating prey diversity and as an index of the frequency of certain prey types in the diet. In our study, they were useful when consider-ing prey at the level of animal classes, but provid-ed only limitprovid-ed information at finer resolution due to the difficulties in separating fragments into species. This lack of usefulness is similar to that found by Newton (1986) for Sparrowhawks (Accipiter nisus), and by Purger (1998) for Red-Footed Falcons (Falco vespertinus). An added problem in Taiwan is that pellets begin to mildew and decompose very quickly during the rainy sea-son, which coincides with the breeding season of the 2 raptors we studied.
Both raptor species we studied are primarily bird eaters (48% vs. 59% of prey items and 76% vs. 68% of prey biomass for A. virgatus and A. trivirgatus, respectively). Thus, a method,s power in identifying bird prey should be especially impor-tant. The observation method worked well in sep-arating birds from other prey classes for both Accipiter species. Both prey remains and pellets revealed fewer species of birds than expected for A. virgatus, compared to the proportion identified by direct observation. In contrast, A. trivirgatus prey remains revealed significantly more bird species than expected.
Sources of bias of the methods
Each of the methods we examined had some shortcomings or biases. The direct observation method recorded prey items only when an observ-er was present, while prey remains and pellet sam-ples represented a sample of the diet for the entire time between collections. For direct observation, even with high-intensity coverage, it is still likely that some prey species were missed for purely sto-chastic reasons. In our study, the numbers of prey items observed and species identified for A. virga-tus through the observation method were both very small, while comparable results for A. trivirgatus were larger. This difference most likely resulted from stochastic factors.
Direct observations can provide the most accurate dietary records if the coverage is relative-ly complete, but the number of nests observed is usually very small compared to those available to the other methods. In addition, the time and labor required to observe just a few nests often render this method unfeasible. Although videography can
lower the time and labor demands of directly moni-toring a nest, reviewing the videotapes afterwards still requires extensive amounts of time and man-power.
When sample sizes are small, the prey remains and pellet methods may also be biased due to stochastic reasons. Many researchers have pointed out that data from pellets and prey remains are often biased (Marti 1987, Redpath et al. 2001, Booms and Fuller 2003). Prey remains are potentially biased towards the more-colorful and conspicuous avian species, towards bird species plucked near nest sites, and against less-conspicuous mammals, small reptiles, amphibians, and arthropods (Reynolds and Meslow 1984, Bielefeldt et al. 1992). In our study, both species of accipiters ate a fairly high number of lizards, yet very few lizard remains were found. Obviously with prey remains, our samples did not represent the entire diet but only the larger or harder prey items. Lizards and perhaps other small or soft items were totally eaten and no remains were left, especially for A. trivirgatus.
The accuracy of the pellet method is greatly influenced by the diet of the predator. A diet pieced together through pellet analysis might not be representative because of differential digesta-bilities of different prey items. Booms and Fuller (2003) further pointed out that different prey types have different surface-to-volume ratios which pos-sibly influence the amount of material left in the pellets. Another source of bias with the pellet method in our study was our ability to identify prey fragments. We used a magnifying glass to exam-ine prey fragments. Our results could probably have been improved by examining fragments under a dissecting microscope or with molecular techniques. However both these methods of iden-tification would have greatly added to the effort and cost required for the pellet method.
In our study, the proportion of prey items iden-tified could have been improved by combining the prey remains and pellet methods. These 2 meth-ods seemed to compensate for each other some-what by producing large-sized prey remains and finding small-sized prey in pellets, potentially underestimating only the small and easily digested prey items. Nevertheless, Redpath et al. (2001) found that combining these methods could not completely eliminate the biases for Hen Harriers.
Using a combination of pellets, prey remains, and videography is the best way to reach a proxi-mate representation of the diet of raptors (Booms and Fuller 2003, Lewis et al. 2004).
CONCLUSIONS
During incubation, the females of both species generally left the nests to feed on prey brought back by males. Thus none of the 3 prey monitoring methods worked well before the eggs hatched. With nestlings present, we found that none of the 3 methods alone provided complete prey information for either species of accipiter. The combination of prey remains and pellets reduced the proportion of prey types omitted to 18% and 41% for A. virgatus and A. trivirgatus, respectively, and provided the largest proportion of prey items identified to species and type. Our study shows that the return on the investment with direct observation varied greatly between species.
Acknowledgments: We wish to thank Ms. S.S.
Hsieh, Mr. R.C. Wang, Mr. T.F. Lee, Mr. R.H. Yu, and Mr. M.L. Jung for help in the field and labora-tory.
REFERENCES
Bielefeldt J, RN Rosenreld, JM Papp. 1992. Unfounded assumptions about the diet of the Cooper,s Hawk. Condor 94: 427-436.
Booms TL, MR Fuller. 2003. Gyrfalcon diet in central West Greenland during the nesting period. Condor 105: 528-537.
Bosakowski T, DG Smith. 1992. Comparative diets of sym-patric nesting raptors in the eastern deciduous forest biome. Can. J. Zool. 70: 984-992.
Collopy MW. 1983. A comparison of direct observations and collections of prey remains in determining the diet of Golden Eagles. J. Wildlife Manage. 47: 360-368. Huang KY, YS Lin, LL Severinghaus. 2004. The diet of Besra
Sparrowhawk (Accipiter virgatus) in Yangmingshan area, Northern Taiwan. Taiwania 49: 149-158.
Joy SM, RT Reynolds, RL Knight, RW Hoffman. 1994. Feeding ecology of Sharp-shinned Hawks nesting in deciduous and coniferous forests in Colorado. Condor
96: 455-467.
Lewis SB, MR Titus, Fuller K. 2004. A comparison of 3
meth-ods for assessing raptor diet during the breeding season. Wildlife Soc. B. 32: 373-385.
Mañosa S. 1994. Goshawk diet in a Mediterranean area of northeastern Spain. J. Raptor Res. 28: 84-92.
Marti CD. 1987. Raptor food habits studies. In BA Pendleton, BA Millsap, KW Chine, DM Bird, eds. Raptor manage-ment techniques manual. Washington, DC: National Wildlife Federation, pp. 67-80.
Newton I. 1986. The Sparrow Hawk. Calton, UK: Poyser. Purger JJ 1998. Diet of Red-footed Falcon Falco vespertinus
nestlings from hatching to fledging. Ornis Fennica 75: 185-191.
Real J. 1996. Biases in diet study methods in the Bonelli,s Eagle. J. Wildlife Manage. 60: 632-638.
Redpath SM, R Clarke, M Madders, SJ Thirgood. 2001. Assessing raptor diet: comparing pellets, prey remains, and observational data at Hen Harrier nests. Condor 103: 184-188.
Reynolds RT, EC Meslow. 1984. Partitioning of food and niche characteristics of coexisting Accipiters during breeding. Auk 101: 761-779.
Rutz C. 2003. Assessing the breeding season diet of Goshawks Accipiter gentilis: biases of plucking analysis quantified by means of continuous radio-monitoring. J. Zool. 259: 209-217.
Sabo AB, RC Laybourne. 1994. Preparation of avian material recovered from pellets and as prey remains. J. Raptor Res. 28: 192-193.
Selås V. 1993. Selection of avian prey by breeding Sparrowhawks Accipiter nisus in south Norway: the importance of size and foraging behaviour of prey. Ornis Fennica 70: 144-154.
Sharp A, L Gibson, M Norton, A Marks, B Ryan, L Semeraro. 2002. An evaluation of the use of regurgitated pellets and skeletal material to quantify the diet of Wedge-tailed Eagles, Aquila audax. Emu 102: 181-185.
Simmons RE, DM Avery, G Avery. 1991. Biases in diets deter-mined from pellets and remains: correction factors for a mammal and bird-eating raptor. J. Raptor Res. 25: 63-67. Squires JR. 2000. Food habits of Northern Goshawks nesting
in south central Wyoming. Wilson Bull. 112: 536-539. Thrailkill JA, LS Andrews, RM Claremont. 2000. Diet of
breed-ing Northern Goshawks in the Coast Range of Oregon. J. Raptor Res. 34: 339-340.
Toyne EP. 1998. Breeding season diet of the Goshawk Accipiter gentilis in Wales. Ibis 140: 569-579.
Watson JW, DW Hays, SP Finn, P Meehan-Martin. 1998. Prey of breeding Northern Goshawks in Washington. J. Raptor Res. 32: 297-305.