MBD4 Gene is Associated with Rheumatoid Arthritis in Chinese Patients in Taiwan
Chung-Ming Huang1,4, Po-Hao Huang1, Chi-Lan Chen8, Lei Wan3, Chang-Hai Tsai6, Su-Ching Liu7, Wen-Liang Huang2, Fuu-Jen Tsai3,5
Division of Immunology and Rheumatology1, Department of Internal Medicine2, and Department of Medicine Genetics3, China Medical University Hospital, Taichung, Taiwan Graduate Institute of Integrated Medicine, College of Chinese Medicine, China Medical University4, Taichung, Taiwan
College of Chinese Medicine, China Medical University5, Taichung, Taiwan Taichung Asia University6, Taichung, Taiwan
Children’s Medical Center, China Medical University Hospital7, Taichung, Taiwan. Foo Yin Institute of Technology8, Kaohsiung, Taiwan
Address correspondence to: Fuu-Jen Tsai,
Department of Medical Genetics, China Medical University Hospital, No 2 Yuh Der Road, Taichung, Taiwan
TEL: 886-4-22052121 ext. 2038 Fax: 886-4-22034608
E-mail: [email protected]
Running title: Patients with RA
Abstract
This study examines whether or not MBD4 polymorphism is a marker for rhematoid arthritis (RA) susceptibility or severity for Chinese patients in Taiwan. This study included 193 patients with RA while 190 unrelated healthy individuals living in Central Taiwan served as controls. The relationship between MBD4 polymorphism and clinical manifestations of RA was evaluated. For the genotype and allelic frequency of MBD4-1057 polymorphism, there were no statistically significant differences between RA patients and controls.There were significant differences in the distribution of MBD4-8666 polymorphism frequencies between RA patients and controls [p = 0.013; Pcorrected(Pc)= 0.039]. There were also
significant relationships in the distribution of MBD4-9229 polymorphism genotype between RA patients and controls (p = 0.007; Pc = 0.021).However, we did not detect any
associations for MBD4-1057, -8666 or -9229 with rheumatoid factor (RF) presence, extraarticular involvement or bone erosion in RA patients. Results suggest that MBD48666 and -9229, but not -1057, gene polymorphisms are related to RA in Chinese patients in Taiwan.
Introduction
Rheumatoid arthritis (RA) is characterized by joint inflammation, along with several peripheral inflammatory manifestations [1]. It is known to involve chronic synovitis, leading to destruction of joint tissue (cartilage and bone) and consequently, serious impairment of joint functions [2]. There is evidence of immunopathogenesis in genetically susceptible hosts [3, 4]. However, the genetic basis of RA is largely unknown.
DNA methylation is an epigenetic regulator of several biological processes, including embryonic development, gene transcription, X-chromosome inactivation, genomic imprinting and chromatin modification [5–8]. Defective regulation of DNA methylation is associated with the pathogenesis of autoimmune diseases, such as systemic lupus erythematosus (SLE), RA, systemic sclerosis and subacute cutaneous lupus erythematosus, but not with
dermatomyositis [9-11]. Methylcytosine bases within promoter regions suppress transcription through association with methyl-CpG-binding domain proteins (MBDs) [13]. MBDs inhibit transcription factor binding by recruiting silencing complexes and histone deacetylases, thereby stabilizing and consolidating the heterochromatic state [14, 15]. Five genes belonging to the MBD family, each characterized by a methyl-binding domain, have been identified in mammalian genomes: MBD1, MBD2, MBD3, MBD4, and MeCP2 [16].
There is a growing body of evidence that supports the notion that MBD4 plays an
important role in DNA methylation and carcinogenesis. A few studies have demonstrated the relationship between MBD4 polymorphisms and susceptibility to cancer, including
esophageal squamous cell carcinoma and lung cancer [17-19]. However, little is known about the relationship between RA and MBD4 gene polymorphism. We hypothesize that genetic variants in MBD4 are associated with RA risk. This study examines polymorphisms for the MBD4-1057, -8666 and -9229 genes to determine whether or not they are markers for RA susceptibility or severity by focusing on single nucleotide polymorphisms (SNPs).
Patients and Methods
We enrolled 193 patients (168 females) diagnosed with RA (1987, revised American College of Rheumatology criteria) [20]. Their median age was 55 (range: 26-71). We also recruited 190 unrelated healthy individuals (169 females) living in Central Taiwan to serve as control subjects. Their median age was 53 (range: 23-69). This study was approved by the Institutional Review Board (IRB) at China Medical University Hospital prior to patient enrollment. Informed consent was obtained from all participants. Nephelometry was used for rheumatoid factor (RF) detection. Values ≧30 Iu/ml were deemed positive. Presence or history of extra-articular features for RA were noted [21]. Radiography for hands, wrists and feet for the RA patients were obtained. Bone erosion was evaluated by a radiologist and a rheumatologist. We noted only the presence or absence of erosion, with no calculation of radiological score.
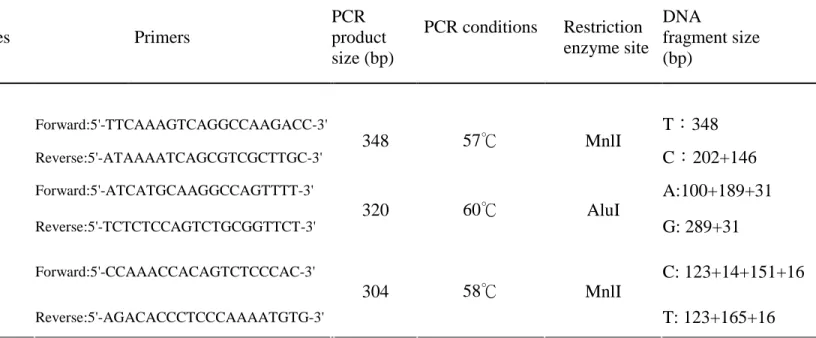
Genomic DNA was extracted from peripheral blood using a genomic DNA isolation reagent kit (Genomaker, Taiwan). Polymerase chain reaction (PCR) for MBD gene polymorphism was carried out to a total volume of 25 μl, containing genomic DNA (2-6 pmole of each primer); 1X Taq polymerase buffer (1.5 mM MgCl2); and 0.25 units of
AmpliTaq DNA polymerase (Perkin Elmer, Foster City, Calif, USA). For detection of MBD4-1057, a 348-bp fragment was amplified using nested PCR. The primer used was upstream 5’- TTCAAAGTCAGGCCAAGACC-3’and downstream 5’
-ATAAAATCAGCGTCGCTTGC-3’.PCR amplificationwas performed in a programmable thermal cycler [Gene Amp PCR System 2400 (Perkin- Elmer)]. Cycling conditions for
MBD4-1057 polymorphism were set as follows: one cycle at 94°C for 5 min, 35 cycles at 94° C for 30 sec, 35 cycles at 60°C for 30 sec, and 35 cycles at 72°C for 40 sec, and one final cycle of extension at 72°C for 7 min. PCR was followed by incubation with two units of Mnl I (New England Biolabs, Beverly,USA); 10lof the product was then loaded onto a 3%
agarose gel containing ethidium bromide for electrophoresis. As shown in Table 1, PCR procedures for MBD4-8666 and MBD4-9229 were the same as those for MBD4-1057.
Allelic frequencies were expressed as percentages of the total number of alleles. Results from controls and RA patients were compared using χ2(2 x 2 contingency tables). When the assumptions for the chi-squared test were violated and one cell had an expected count of <1, ormorethan 20% ofcellshad an expected countof<5,Fisher’sexacttestwasused.The distribution of MBD4 gene polymorphisms were evaluated for each group.Because of the multiple analysis conducted, the Bonferroni correction was applied to produce corrected probabilities (Pc).A p-value of less than 0.05 was considered statistically significant. Odds ratios (OR) were calculated from allelic frequency with a 95% confidence interval (95% CI) for MBD4 gene polymorphism.
Results
Genotype frequencies for RA and control groups are shown in Table 2. Among the 193 RA patients, 160 (90.9%) had MBD4-1057 genotype TT, 15 (8.5%) had T/C and one patient (0.6%) had CC. Among the 190 controls, the MBD4-1057 genotype TT was found in 158 (86.3%), T/C was found in 22 (12.0%), and CC was found in three (1.7%). There were no significant differences in MBD-1057 polymorphic distribution between controls and RA patients (p=0.33). There was also no significant association for MBD4-1057 allelic frequency between RA patients and control subjects (p=0.12), giving an odds ratio of 1.63 for the T allele (95% confidence interval: 0.88-3.04). However, 74 patients with RA (40.4%) had MBD4- 8666 genotype GG, while 80 (43.7%) had G/A and 29 (15.9%) had AA. Among controls, the MBD4- 8666 genotype GG was found in 49 subjects (26.5%), while G/A was noted in 99 (53.5%) and AA was found in 37 (20.0%). There were significant differences in the polymorphic distribution for MBD4- 8666 between controls and RA patients (p=0.018),
giving an odds ratio of 1.93 for the GG genotype (95% confidence interval: 1.05-3.53). There were also significant associations in allelic frequency for MBD4-8666 between RA patients and controls (p=0.013;Pc = 0.039), giving an odds ratio of 1.45 for the G allele (95% confidence interval 1.08-1.95). For MBD4-9229, 58 RA patients (30.7%) had genotype TT, while 84 (44.4%) had T/C and 47 (24.9%) had AA. Among controls, the MBD4- 9229 genotype TT was found in 29 subjects (16.7%), while T/C was found in 97 (56.1%) and CC was noted in 47 (27.2%). There was a significant difference in MBD4-9229 polymorphic distribution between controls and RA patients (p=0.007; Pc = 0.021), giving an odds ratio of 2.0 for the TT genotype (95% confidence interval: 1.10-3.65). There was also significant association in the allelic frequency for MBD4-9229 between RA patients and controls (p=0.03), giving an odds ratio of 1.38 for the T allele (95% confidence interval: 1.03-1.86).
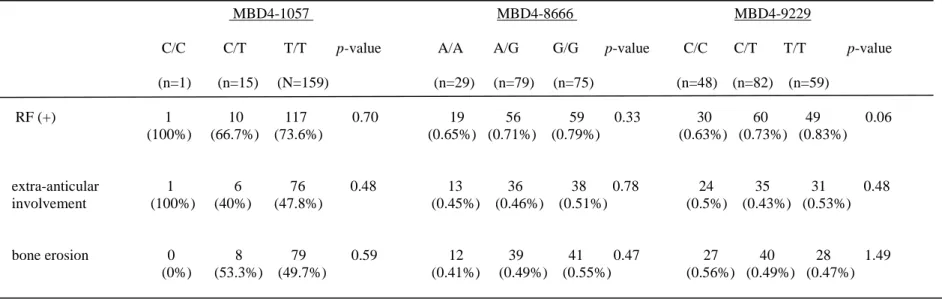
Clinical manifestations and laboratory findings for RA patients are shown in Table 3. The associations of MBD4 with particular clinical features of RA were examined. We did not detect any association between MBD4-1057 with the rheumatoid factor (RF), extra-articular involvement or bone erosion (p= 0.7, 0.48 and 0.59, respectively). We also did not find any relationships for MBD4-8666 and MBD4-9229 with the same clinical manifestations (p= 0.33, 0.78, 0.47, and p= 0.06, 0.48, 1.49, respectively). However, we did observe increased frequencies for RF among RA patients with the TT genotype (83.1%) when compared with CC (62.5%). However, this difference did not reach statistical significance (p=0.06).
Discussion
MBD4, also known as MED1 (methyl-CpG binding endonuclease 1), was identified as a methyl-CpG binding protein in 1998 [22]. This protein contains an N-terminal
5-methylcytosine–binding domain (MBD), which allows binding to methylated DNA, and a C-terminal catalytic domain with homology to bacterial DNA damage–specific
glycosylases/lyases. It is therefore thought to be an important component of a system that maintains the integrity of the methylation pattern throughout the genome [23–24]. Enzymatic removal of 5-methylcytosine from DNA by MBD4 can suppress CpG mutability [25]. MBD4 has also been shown to play an important role in genomic surveillance and apoptosis progress through interaction with the MLH1 repair protein and the Fas ligand protein [26-27].
Some medications that cause drug-induced lupus (procainamide, hydralazine), as well as ultraviolet light, can inhibit DNA methylation in cloned T-cell lines and can induce self-reactivity [28]. Such agents induce overexpression of lymphocyte function-associated antigen-1 (LFA-1), which confers an autoreactive status to T-cells [29]. CD4+ T-cells from patients with active lupus hypomethylate DNA and overexpress LFA-1 on an autoreactive subset, which spontaneously lyses autologous macrophages, which supports the hypothesis that methylation inhibition is sufficient to cause lupus-like illness [30-31]. Defective
regulation of DNA methylation is associated with the pathogenesis of autoimmune diseases, such as systemic lupus erythematosus (SLE), RA, systemic sclerosis and subacute cutaneous lupus erythematosus, but not dermatomyositis [9-12]. Regarding MBD4, its deficiency seems to reduce the apoptotic response [32]. In fact, MBD4 can directly participate in this process by interacting with the Fas-associated death domain protein [33]. An increase in MBD4 expression could induce an abnormal rate of cell death, which would account for the increased cell apoptosis and impaired clearance of dying cells frequently found in SLE patients. Balada et al., demonstrated that MBD4 levels in SLE patients are significantly increased and hypothesized that MBD4 may have an active role in the global demethylation observed in SLE patients [10].Richardson et al. demonstrated the relationship between DNA hypomethylation and RA [9]. That study proposed that chronic T-cell activation could affect T-cell DNA methylation, which remains a possibility for RA.
DNA damage and is a tumor suppressor gene candidate [26, 34-35]. MBD4 may play an important role in DNA methylation and carcinogenesis. A few studies have demonstrated the relationship between MBD4 polymorphism and susceptibility to cancer, including esophageal squamous cell carcinoma and lung cancer [17-19]. However, little is known about the
relationship between RA and MBD4 polymorphism.
To the best of our knowledge (MEDLINE literature search), an association between MDB4 polymorphism and RA has not been demonstrated before. We hypothesized MBD4 to be associated with RA risk. This study examined the MBD4-1057, -8666 and -9229 variants to determine whether or not they are markers for RA susceptibility or severity by focusing on SNPs. We compared allelic and genotypic frequencies between 193 Chinese RA patients and 190 healthy individuals in Taiwan.There were significant differences in the distribution of MBD4-8666 frequencies between RA patients and controls (p= 0.013; Pc = 0.039). There were also significant relationships in MBD4-9229 genotype between RA patients and controls (p= 0.007; Pc = 0.021).We conjecture that MBD4-8666 and -9229 gene
polymorphisms may promote MBD4 gene expression and cause the global demethylation in patients with RA.
This study suggests that MBD4-8666 and -9229, but not -1057, are related to RA in Chinese patients in Taiwan. However, there are no associations between the genotype of MBD4-1057, -8666, and -9229 and rheumatoid factor (RF), extra-articular involvement or bone erosion.
Acknowledgment
This study was supported by a grant (DMR-93-007) from China Medical University Hospital, Taiwan, R.O.C.
References:
1. Feldmann M, Brennan FM, Maini RN (1996) Rheumatoid arthritis. Cell 85:307-310. 2. Harris ED Jr (1990) Rheumatoid arthritis: pathophysiology and implications for therapy. N
Engl J Med 322:1277-1289.
3. Wordsworth BP, Bell JI (1991) Polygenic susceptibility in rheumatoid arthritis. Ann Rheum Dis 18:343-346.
4. Gomolka M, Menninger H, Saal JE, Kemmel EM, Albert ED, Niwa O, et al (1995) Immunoprinting: various genes are associated with increased risk to develop rheumatoid arthritis in different groups of adult patients. J Mol Med 73:19-29.
5 Okano M, Bell DW, Haber DA et al. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999; 99:247–57. 6 Reik W, Walter J. Genomic imprinting: parental influence on the genome. Nat Rev Genet
2001; 2:21–32.
7 Li E. Chromatin modification and epigenetic reprogramming in mammalian development. Nat Rev Genet 2002; 3:662–73.
8 Surani MA. Imprinting and the initiation of gene silencing in the germ line. Cell 1998; 93:309–12.
9. Richardson B, Scheinbart L, Strahler J, Gross L, Hanash S, Johnson M. Evidence for impaired T cell DNA methylation in systemic lupus erythematosus and rheumatoid arthr itis.Arthritis Rheum 1990;33:1665–73.
10. Balada E, Ordi-Ros J, Vilardell-Tarre´s M. DNA methylation and systemic lupus erythematosus. Ann N Y Acad Sci 2007;1108:127–36.
11. Lei W, Luo Y, Yan K, Zhao S, Li Y, Qiu X, Zhou Y, Long H, Zhao M, Liang Y, Su Y, Lu Q. Abnormal DNA methylation in CD4+ T cells from patients with systemic lupus erythematosus, systemic sclerosis, and dermatomyositis Scand J Rheumatol. 2009 ;14:1-6.
12. Luo Y, Li Y, Su Y, Yin H, Hu N, Wang S, Lu Q. Abnormal DNA methylation in T cells from patients with subacute cutaneous lupus erythematosus. Br J
Dermatol.2008 ;159:827-33.
13. Fujita N, Takebayashi S, Okumura K, Kudo S, Chiba T, Saya H, et al. Methylation-mediated transcriptional silencing in euchromatin by methyl-CpG binding protein MBD1 isoforms. Mol Cell Biol 1999;19:6415–26.
14. Bird AP, Wolffe AP. Methylation-induced repression: belts, braces, and chromatin. Cell 1999;99:451–4.
15. Leonhardt H, Cardoso MC. DNA methylation, nuclear structure, gene expression and cancer. J Cell Biochem Suppl 2000;35:78–83.
16. Hendrich B, Bird A. Identification and characterization of a family of mammalian methyl-CpG binding proteins. Mol Cell Biol 1998;18:6538–47.
17. Hao B, Wang H, Zhou K, Li Y, Chen X, Zhou G, et al. Identification of genetic variants in base excision repair pathway and their associations with risk of esophageal
squamous cell carcinoma. Cancer Res 2004;64:4378—84.
18. Shin MC, Lee SJ, Choi JE, Cha SI, Kim CH, Lee WK, et al. Glu346Lys polymorphism in the methyl-CpG binding domain 4 gene and the risk of primary lung cancer. Jpn J Clin Oncol
2006;36:483—8.
19. Miao R, Gu H, Liu H, Hu Z, Jin G, Wang H, Wang Y, Sun W, Ma H, Chen D, Tian T, Jin L, Wei Q, Lu D, Huang W, Shen H. Tagging single nucleotide polymorphisms in MBD4 are associated with risk of lung cancer in a Chinese population. Lung Cancer. 2008;62:281-6.
20. Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, et al (1988) The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum 3:315-324.
21. Yen JH, Chen JR, Tsai WJ, Tsai JJ, Liu HW (1995) HLA-DRB1 genotyping in patients with rheumatoid arthritis in Taiwan. J Rheumtol 22:1450-1454.
22. Hendrich B, Bird A. Identification and characterization of a family of mammalian methyl-CpG binding proteins. Mol Cell Biol 1998;18:6538—47.
23. Bellacosa A, Cicchillitti L, Schepis F, Riccio A, Yeung AT, MatsumotoY, et al. MED1, a novel human methyl-CpG-binding endonuclease, interacts with DNA mismatch repair protein MLH1. Proc Natl Acad Sci USA 1999;96:3969–74.
24. Riccio A, Aaltonen LA, Godwin AK, Loukola A, Percesepe A, Salovaara R, et al. The DNA repair gene MBD4 (MED1) is mutated in human carcinomas with microsatellite instability. Nat Genet 1999;23:266–8.
25. Millar CB, Guy J, Sansom OJ, Selfridge J, MacDougall E, Hendrich B, et al. Enhanced CpG mutability and tumorigenesis in MBD4-deficient mice. Science 2002;297:403-5. 26. Bellacosa A, Cicchillitti L, Schepis F, Riccio A, Yeung AT, Matsumoto Y, et al. MED1,
a novel human methyl-CpG-binding endonuclease, interacts with DNA mismatch repair protein MLH1. Proc Natl Acad Sci USA 1999;96:3969-74.
27. Screaton RA, Kiessling S, Sansom OJ, Millar CB, Maddison K, Bird A, et al. Fas-associated death domain protein interacts with methyl-CpG binding domain protein 4: a potential link between genome surveillance and apoptosis. Proc Natl Acad Sci USA 2003;100:5211-6
28. CORNACCHIA, E., J. GOLBUS, J. MAYBAUM, et al. 1988. Hydralazine and procainamide
29. YUNG, R., S. CHANG, N. HEMATI, et al. 1997. Mechanisms of drug-induced lupus. IV. Comparison of procainamide and hydralazine with analogs in vitro and in vivo. Arthritis Rheum. 40: 1436-1443.
30. RICHARDSON, B. C., J. R. STRAHLER, T.S. PIVIROTTO, et al. 1992. Phenotypic and functional similarities between 5-azacytidine-treated T cells and a T cell subset in patients with active systemic lupus erythematosus. Arthritis Rheum. 35: 647-662.
31. TAKEUCHI, T., K. AMANO, H. SEKINE, et al. 1993. Upregulated expression and function of integrin adhesive receptors in systemic lupus erythematosus patients with vasculitis. J. Clin. Invest. 92: 3008–3016.
32. SANSOM, O. J., J. ZABKIEWICZ, S.M. BISHOP, et al. 2003. MBD4 deficiency reduces the apoptotic response to DNA-damaging agents in the murine small intestine. Oncogene 22: 7130–7136.
33. SCREATON, R. A., S. KIESSLING, O. J. SANSOM, et al. 2003. Fas-associated death domain protein interacts with methyl-CpG binding domain protein 4: a potential link between genome surveillance and apoptosis. Proc. Natl. Acad. Sci. USA 100: 5211–5216. 34. Cortellino S, Turner D, Masciullo V, Schepis F, Albino D, Daniel R, et al. The base
excision repair enzyme MED1 mediates DNA damage response to antitumor drugs and is associated with mismatch repair system integrity. Proc Natl Acad Sci USA
2003;100:15071-6.
35. Bader SA, Walker M, Harrison DJ. A human cancer-associated truncation of MBD4 causes dominant negative impairment of DNA repair in colon cancer cells. Br J Cancer 2007;96:660-6.
Table 1. Primer panel for MBD4 gene application.
Gene SNPs rs # Alleles Primers
PCR product size (bp) PCR conditions Restriction enzyme site DNA fragment size (bp) Forward:5'-TTCAAAGTCAGGCCAAGACC-3' T:348 MBD4-1057 rs2311394 C/T Reverse:5'-ATAAAATCAGCGTCGCTTGC-3' 348 57℃ MnlI C:202+146 Forward:5'-ATCATGCAAGGCCAGTTTT-3' A:100+189+31 MBD4-8666 rs3138355 A/G Reverse:5'-TCTCTCCAGTCTGCGGTTCT-3' 320 60℃ AluI G: 289+31 Forward:5'-CCAAACCACAGTCTCCCAC-3' C: 123+14+151+16 MBD4 MBD4-9229 rs2005618 C/T Reverse:5'-AGACACCCTCCCAAAATGTG-3' 304 58℃ MnlI T: 123+165+16
Table 2. Comparison of MBD4 genotype distribution and allelic frequencies observed in RA patients and healthy control subjects.
RA Controls
Gene name, SNP database ID
Number (%) Number (%) p-value (Pcorrected) Odds Ratio (95% CI)
MBD4-1057 T/T 160(90.9) 158(86.3) 0.333 (0.999) 3.04 (0.31-29.52) T/C 15(8.5) 22(12.0) 2.05 (0.19-21.59) C/C 1(0.6) 3(1.7) 1.00 T allele 335(95.2) 338(92.3) 0.119 (0.357) 1.63 (0.88-3.04) C allele 17(4.8) 28(7.7) 1.00 MBD4-8666 G/G 74(40.4 %) 49(26.5%) 0.018 (0.054) 1.93 (1.05-3.53) G/A 80(43.7%) 99(53.5%) 1.03 (0.58-1.82) A/A 29(15.9 %) 37(20.0%) 1.00 G allele 228(62.3%) 197(53.2%) 0.013 (0.039)* 1.45 (1.08-1.95) A allele 138(37.7%) 173(46.8%) 1.00 MBD4-9229 T/T 58(30.7%) 29(16.7%) 0.007 (0.021)* 2.00 (1.10-3.65) T/C 84(44.4%) 97(56.1%) 0.87 (0.53-1.43) C/C 47(24.9%) 47(27.2%) 1.00 T allele 200(52.9%) 155(44.8%) 0.029 (0.087) 1.38 (1.03-1.86) C allele 178(47.1%) 191(55.2%) 1.00
Table 3. Relationships between MBD4-1057, -8666 & -9229 genotypes and clinical signs and findings for RA patients
MBD4-1057 MBD4-8666 MBD4-9229
C/C C/T T/T p-value A/A A/G G/G p-value C/C C/T T/T p-value
(n=1) (n=15) (N=159) (n=29) (n=79) (n=75) (n=48) (n=82) (n=59) RF (+) 1 10 117 0.70 19 56 59 0.33 30 60 49 0.06 (100%) (66.7%) (73.6%) (0.65%) (0.71%) (0.79%) (0.63%) (0.73%) (0.83%) extra-anticular 1 6 76 0.48 13 36 38 0.78 24 35 31 0.48 involvement (100%) (40%) (47.8%) (0.45%) (0.46%) (0.51%) (0.5%) (0.43%) (0.53%) bone erosion 0 8 79 0.59 12 39 41 0.47 27 40 28 1.49 (0%) (53.3%) (49.7%) (0.41%) (0.49%) (0.55%) (0.56%) (0.49%) (0.47%)