行政院國家科學委員會專題研究計畫 成果報告
過敏原對樹突細胞活化及訊息傳導的影響(2/2)
計畫類別: 個別型計畫
計畫編號: NSC93-2314-B-002-045-
執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日
執行單位: 國立臺灣大學醫學院臨床醫學研究所
計畫主持人: 江伯倫
計畫參與人員: 黃心穎 洪維鍊
報告類型: 完整報告
處理方式: 本計畫可公開查詢
中 華 民 國 94 年 10 月 31 日
行政院國家科學委員會補助專題計畫成果報告
計畫名稱:過敏原對樹突細胞活化及訊息傳導的影響
計畫類別:個別型計畫
計畫編號:NSC 92-2314-B-002-201 及
NSC 93-2314-B-002-045-
執行日期:民國九十二年八月一日至民國九十四年七
月三十一日
計畫主持人:江伯倫 教授
計畫參與人員:黃心穎 洪維鍊
執行單位:台大醫學院臨床醫學研究所
中文摘要
關鍵詞:樹突細胞、訊息傳導、過敏原
樹突細胞被認為是在免疫反應的產生上扮演著一個最關鍵的角色,有愈
來愈多的證據顯示樹突細胞可能在決定身體免疫系統接觸到外來抗原時的免疫
反應的走向。也因此,如果我們能夠對樹突細胞的活化和成熟機轉,和活化後
凋亡的機制有更進一步的了解,將有助於我們研究免疫反應的機轉。尤其是在
調控免疫反應的走向上,有著關鍵的影響。所以我們將分別研究不同刺激下的
樹突細胞,是否會產生不同型的樹突細胞。目前知道樹突細胞如果接受不同的
刺激,也會在輔助分子和細胞激素的分泌上有著不同的表現,而這些不同表現
的樹突細胞便會主導不同免疫反應的進行。而這些不同表現型的樹突細胞可能
在活化途徑和凋亡現象上可能有著相當大的差異,如果能夠釐清這些現象將讓
我們對疾病的機轉有更清楚的認識。我們在這段期間分別建立了試管內模式,
能夠利用樹突細胞來評估各種不同刺激對樹突細胞的影響。結果我們發現樹突
細胞的試管內表現的確也可以表現出不同的細胞表面標記和細胞激素,如果
IL-12 的表現量較高則會讓免疫反應往第一型免疫反應的方向發展。我們也成功地
利用由靈芝衍生來的多醣研究樹突細胞的訊息傳導途徑,得到相當不錯的研究
成果,已經發表在 Journal of Leukocyte Biology。
Abstract
Dendritic cells (DCs) are the most potent antigen-presenting cells (APCs) in the
immune system, which can trigger the immune response. They not only express high
level of the MHC-peptide complex, but also the other accessory molecules that interact
with the receptors on the T cells to enhance adhesion and signalling. More and more
evidences suggest that dendritic cells play the critical role in the initiation and
development of immune response. In this project, we aim to clarify the activation
pathway involved in the different types of dendritic cells with the stimulation of mite
allergens. In this project, we aim to clarify the functional changes and activation signals
of dendritic cells after activation with mite allergens. The information generated in the
project will help us in understanding the basic mechanisms of activation pathway of the
dendritic cells and also their roles in the pathogenesis of the diseases. In this project, we
have established an in vitro assay for the assessment of allergen, lipopolysaccharide and
CpG motif on the function of dendritic cells. Further, we also like to correlate the
phenotypes of dendritic cells and in vivo function of these molecules. Finally, we have
also established the method for the assay of signal pathway of dendritic cell activation.
We believe the information here will provide a very novel system in studying the
mechanisms of allergic diseases, also other immunological diseases.
前言
此一研究計畫主要是要建立一個模式來研究是否可以建立一個模式來評
估過敏原對樹突細胞的影響,再了解過敏原引起特定的第二型 T 輔助細胞的機
制。在過去一段時間,我們已經分別建立了樹突細胞的培養,並利用各種不同
的刺激來了解是否可以來經由樹突細胞的表面分子表現和細胞激素的製造來達
到評估免疫反應的目的。我們在過去一段時間利用了由細菌分離出來的多醣、
靈芝和過敏原 mite 來進行這方面的研究,並進一步來研究樹突細胞的訊息傳導
途徑,這部份一直是較少看到相關的研究。而樹突細胞被認為在整個免疫反應
中扮演了一個最重要的角色,所以如果我們能夠有效地將樹突細胞培養出,再
評估其細胞表面分子和細胞激素的分泌情形,便可以再進一步到體內來評估是
否能夠誘發相關的免疫反應,就可以將試管內的樹突細胞表現與體內的免疫反
應來加以分析。同時,有關樹突細胞的訊息傳導研究也是這幾年來才逐步有較
多的研究,所以進一步研究這些不同刺激的訊息傳導,也將是一個有興趣的課
題。
研究目的
此ㄧ研究計畫的主要目的是要建立一個試管內樹突細胞培養的方式來評
估不同刺激對樹突細胞的影響,尤其是我們將同時評估在過敏反應中較重要的
一些刺激,如過敏原、CpG motif 和相關的多醣。
文獻探討
而在抗原呈現細胞中,樹突細胞(dendritic cells, DC)又被稱為專業的抗原
呈現細胞,且被認為是控制免疫反應極為關鍵的細胞。這幾年由於樹突細胞的
培養愈來愈方便,所以有相當的研究及治療可以利用培養的樹突細胞來達到。
近年來的研究顯示,樹突細胞在誘發與調控免疫反應上扮演重要角色。而且,
來源不同的兩種樹突細胞 - myeloid 樹突細胞和 lymphoid 樹突細胞,對於 T 細
胞的的發育會導致完全不同的影響。樹突細胞被認為是體內負責加工並呈獻抗
原最重要的抗原呈獻細胞。目前已證明樹突細胞加入 GM-CSF、TNF- 和 IL-4
等細胞激素後便可成功的在活體外培養。有許多實驗已驗證活體外培養的樹突
細胞在許多模式中都可以誘發第一類和第二類 MHC 分子限制的免疫反應。以
往的觀念中,將 T 細胞的活化分成兩訊息傳導。最近的研究顯示樹突細胞在 T
細胞的活化上更扮演了一個主導 T 細胞發育的角色,所以抗原在與樹突細胞接
觸時便已經決定了 T 細胞的功能。舉例來說,CpG motif 會刺激第一型 T 輔助
細胞的發生,主要是因為 CpG motif 會刺激樹突細胞分泌較高的 IL-12 而幫助 T
細胞的發育。所以如果利用樹突細胞來進行這些過敏原的研究,便可以了解過
敏原本身的特性。
所以我們利用此ㄧ樹突細胞的模式來進行研究,並進一步來探討是否過敏
原和相關的環境因子,或是治療開發的製劑上會對樹突細胞有特定的影響,而
達到我們研究的目的。同時,對樹突細胞的訊息傳導目前的研究也正是方興未
璦,所以如果建立相關的方法在未來的研究上應該會有相當大的助益。
也因此,我們能夠利用這些培養的樹突細胞來進行相關的研究。利用樹突
細胞當成一個技術平台來評估各種抗原,能夠應用在促進免疫功能,進而可以
用在腫瘤的免疫治療上。舉例而言,一個特定的製劑可刺激樹突細胞分泌細胞
激素如跟第一型 T 輔助細胞相關的 IL-12,或是增加與免疫反應相關的輔助分子
(accessory molecules),如第二型 MHC 分子和 B7.1 分子,也可能會將增進免疫
反應,而達到治療過敏疾病的效果。如此,可以利用目前已經相當成熟的樹突
細胞培養,來檢視一些相關的過敏原和分子在過敏疾病中所扮演的角色。
研究方法
動物在此一研究計畫中所使用的小鼠主要是雌性的Balb/c小鼠,是來自台大
醫學院的動物中心。
骨髓-衍生的樹突細胞培養
為了要培養骨髓-衍生的樹突細胞,我們將小鼠的腿骨兩端剪開後以細胞
培養液將其中的幹細胞沖洗出。將大約每 ml 培養液有 106 細胞的濃度加入細胞
激素 GM-CSF (500 U/ml) and IL-4 (1000 U/ml) (Pepro Tech Inc., Rocky Hill, NJ)來
一道培養。培養基為 RPMI-1640 medium supplemented with 5% heat-inactivated
FCS, 4 mM L-glutamine, 25 mM Hepes (pH 7.2), 50 µM 2-mercaptoethanol, 100 U/ml
penicillin, 100 µg/ml streptomycin and 0.25 µg/ml amphotericin. 每隔一天,我們便會
加入有 GM-CSF and IL-4 的新培養基. 在培養的第八天時將這些骨髓衍生的樹突
細胞(BM-DC)收集後進一步測定其 MHC class II, B7-1, B7-2, CD11c and 33D1 的表
現。這些 BM-DCs 也會被進一步測定其刺激 T 細胞的能力。
以不同刺激來刺激樹突細胞
為了進一步了解是否可以利用樹突細胞來評估不同刺激在試管內的情形,
和這些表現與體內免疫反應的相關性,我們將分別利用過敏原、CpG motif 和與
免疫反應相關的一些多醣。我們將前面培養出來的樹突細胞與
樹突細胞
為了進一步分析樹突細胞的表面標記,我們分別利用認識 MHC class II,
Mac-1, CD11c, B7-1, B7-2, CD40 and OX40L 的抗體來進行細胞表面標記的染色。
樹突細胞將置於 0.5 ml of PBS with 0.1% sodium azide 中來進行螢光流體計數儀的
分析。我們將分別在螢光流體計數儀上分析 10,000 顆細胞,分別利用適當的軟體
來加以分析 (FACScan, Becton Dickinson, Mountain View, CA). 對照組則是細胞不
加入這些特定的抗體。
細胞激素的分泌情形
我們將樹突細胞與這些不同的刺激培養後,分別在 24, 48 and 72 小時後將
培養的上清液取出, 將分別利用 Quantikine M ELISA Kit (R&D Systems, MN,
USA)來測定如 IL-10 和 IL-12 的分泌情形。同時,在測定樹突細胞刺激 T 細胞進
行細胞增殖的同時,我們也將進一步分析這些 T 細胞分泌細胞激素的情形,所以
將分別測定這些細胞分泌如 IFN-g 和 IL-2 的濃度。
樹突細胞激媒的活性分析
樹突細胞受到刺激後,將分別收集細胞來分析其 ERK, JNK and MAP
kinase 的活性。我們會將樹突細胞以不同刺激一道培養後,收取其 200µg 溶解物
質加入 1µg of anti-ERK, anti-JNK1 or 1µg of anti-p38a 的抗體來處理,再利用 20µl
of protein G-Sepharose 來處理。而這些酵素的受質分別為 myelin basic protein 用在
ERK assay, GST-c-Jun1-7 用在 JNK assay, GST-ATF-21-109 用在 p38 MAPK assay.
這些作用的免疫複合物將進一步加以跑膠,再利用 densitormeter 來測定其活性。
NF-kB 的活性分析
樹突細胞培養後將利用 Lipofectamine (Promega)將相關帶有reporter的質體送
入細胞內。質體, 3xkB-L (kindly provided by Dr. Nakano, Juntendo University), 帶有
NF-kB 在minimal thymidine kinase promoter and a luciferase gene的上游 in pGL-2
vector (Promega)。在培養和刺激24小時後將相關的細胞溶解物取出後利用冷光分
析儀來測定其冷光強度。
樹突細胞的 NF-kB 磷酸化分析
為了進一步了解樹突細胞活化時 NF-kB 的相關酵素的磷酸化情形,我們在
樹突細胞活化後,利用抗體將相關的蛋捕捉後,這些免疫沈澱物將分別與 1µg of
GST-I
B
(1
-100), GST-I
-100AA), GST-I B (1
B
(1
-120), GST-I B (1
-120AA), GST-I
-61), or GST-I B (1
B
(1
-61AA) and [ -32P]ATP (10µCi)及激
媒受質一道作用。此一反應再加入 Laemmli’s sample buffer 來終止。這些產物則
也是進一步跑電泳來分析其強度。而這些蛋白都會進一步利用西方墨點法確定其
蛋白質的正確性。
TLR 在樹突細胞活化中所扮演的角色
為了進一步了解這些刺激影響樹突細胞的途徑究竟是來自哪一個受體,我
們將分別利用抗 TLR-2、TLR-4 和 TLR-9 的抗體來加入培養中,以了解在何種抗
體的阻斷下,樹突細胞的細胞激素製造會受到最明顯的影響。在這種情形下,我
們便可以了解哪一個受體在樹突細胞的活化中扮演了一個最重要的角色。
結果與討論
一.我們利用樹突細胞的培養,已經成功地建立一個試管內模式,可以經由樹
突細胞在試管內的表現來評估此一試劑進入體內的反應。我們在初期分別利用
由靈芝衍生來的多醣和 CpG motif 來進行相關研究。我們發現樹突細胞在經由
刺激後會表現較高量的輔助分子,同時其細胞激素的分泌也分別與引起的免疫
反應有著密切的關係。
我們更進一步研究樹突細胞的訊息傳導途徑,結果能夠成功地分析其刺
激後的與 NF-kB 和特定的酵素激媒有著密切的關係。此一研究成果已經發表在
今年的 Journal of Leukocyte Biology (Lin, Y.-L. et al Polysaccharide purified from
Ganoderma lucidum induced activation and maturation of human monocyte-derived
dendritic cells by the NF-kB and p38 mitogen-activated protein kinase pathways. J
Leuko Biol 2005; 78:533-543)
二.我們也同時利用過敏原來進行對樹突細胞的分析,但是在目前的結果發現
對樹突細胞的細胞激素分泌的影響並未與其他抗原有著明顯的差別。所以,我
們目前還在研究其他的相關免疫調節分子,如 DcR3,目前有研究顯示 DcR3 分
子可能在第二型 T 輔助細胞發育的調控上也扮演著重要的角色,在一段時間後
應該會有更清楚的研究結果。
文獻探討
Baeuerle PA and Baltimore D. NF-kB: Ten years after. Cell 1996, 87:13-20.
Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature 1998;
392:245-52.
Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka
K. Immunobiology of dendritic cells. Annu Rev Immunol 2000; 18:767-811.
Bell D, Young J.W.,Banchereau J. Dendritic cells. Adv in Immunol 1999, 72:255-324
Brightbill H D & Modlin R L, Toll-like receptors: molecular of the mammalian
immune response. Immunology 2000;101:1-10.
Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A and Alber G.
Ligation of CD40 on dendritic cells triggers production of high levels of
interleukin-12 and enhances T cell stimulatory capacity: T-T help via APC
activation. J. Exp. Med. 1996, 184: 747-752.
Cella M, Sallusto F and Lanzavecchia A. Origin, maturation and antigen presenting
function of dendritic cells. Current Opin Immunol 1997, 9:10-16.
Cella M, Jarrossay D, Facchetti F, Alebardi O, Nakajima H, Lanzavecchia A and
Colonna M. Plasmacytoid monocytes migrate to inflamed lymph nodes and
produce large amounts of type I interferon. Nature Medicine1999;5:919-923.
Chen A.I., McAdam A.J.,Buhmann J.E.,Scott S.,Lupher M.L.,Jr.,Greenfield E.A.,Baum
P.R.,Fanslow G.J.,Calderhead D.M.,Freeman G.J.,Sharpe A.H. Ox 40-ligand has a
critical costimulatory role in dendritic cell :T cell interactions Immunuty
1999,11:689-698.
Flores-Romo L, In vivo maturation and migration of dendritic cells. Immunology
2001;102:255-262.
Gagliardi MC, Sallusto F, Marinaro M, Langenkamp A, Lanzavecchia A, De Magistris
MT. Cholera toxin induces maturation of human dendritic cells and licences them
for Th2 priming. Eur. J. Immunol. 2000, 30:2394-2403.
Gallucci S, Lolkema M, Matzinger P. Natural adjuvants: endogenous activators of
dendritic cells. Nature Med 1999; 5:1249-55.
Girolomoni G, Ricciardi-Castagnoli P. Dendritic cells hold promise for immunotherapy.
Immunology Today 1997; 18:102-104.
Goerdt S and Orfanos CE. Other functions, other genes: alternative activation of
antigen-presenting cells. Immunity 10:137-142.
Hsieh, K. H. & Shen, J. J. Prevalence of childhood asthma in Taipei, Taiwan, and other
Asian Pacific countries. J. Asthma 1988, 25: 73-82.
Huang F-P, Platt N, Wykes M, James R. Major, Timothy J.Powell, Jenkins C D, and
MacPherson G G. A discrete subpopulation of dendritic cells to T cell areas of
mesenteric lymph nodes.J.Exp.Med 2000;191:435-433.
Inaba K, Inaba M, Romani N, Aya H, Deguchi M, Ikehara S, Muramatsu S, Steinman
RM. Generation of large numbers of dendritic cells from mouse bone marrow
cultures supplemented with granulocyte / macrophage colony-stimulating factor. J
Exp Med 1992; 176:1693-1702.
Inaba K, Turley S, Yamaide F, Iyoda T, Mahnke K, Inaba M, Pack M, Subklewe M,
Sauter B, Sheff D, Albert M, Bhardwaj N, Mellman I, Steiman RM. Efficient
presentation of phagocytosed cellular fragments on the major histocompatibility
complex class II products of dendritic cells. J. Exp. Med. 1998; 188:2163-2173.
Ingulli E, Mondino A, Khoruts A, Jenkins MK. In vivo detection of dendritic cell
antigen presentation to CD4+ T cells. J. Exp. Med. 1997; 185:2133-2141.
Iwasaki A and Kelsall BL. Freshly isolated Peyer’s patch, but not spleen, dendritic cells
produce interleukin-10 and induce the differentiation of T helper type 2 cells. J.
Exp. Med. 1999, 190: 229-239.
Iwasaki A and Kelsall B L. Localization of Distinct Peyer’s Patch Dendritic Cell
Subsets and Their Recruitment by Chemokines Macrophage Inflammatory Protein
(MIP)-3a, MIP-3b, and Secondary Lymphoid Organ Chemokine. J.Exp.Med
2000;191:1381-1393.
Kaisho T, Akira S, Dendritic-cell function in Toll-like receptor-and MyD88-knockout
mice. Trends in Immunol 2001;22:78-83.
Knight SC, Iqball S, Roberts MS, Macatonia S, Bedford PA. Transfer of antigen
between dendritic cells in the stimulation of primary T cell proliferation. Eur. J.
Immunol. 1998; 28:1636-1644.
Koch F, Stanzl U, Jennewein P, Janke K, Heufler C, Kampgen E, Romani N and
Schuler G. High level IL-12 production by murine dendritic cells: upregulation via
MHC class II and CD40 molecules and downregulation by IL-4 and IL-10. J. Exp.
Med. ……..
Kooten C V and Banchereau J. Functions of CD40 on B cells, dendritic cells and other
cells. Curr Opin Immunol 1997;9:330-337.
Lane P JL and Brocker T. Developmental regulation of dendritic cell function. Curr
Opin in Immunol 1999;11:308-313.
Langenkamp A, Messi M, Lanzavecchia A, & Sallusto F, Kinetics of dendritic cell
actvation:impact on priming of T
H1, T
H2 and nonpolarzed T cells.Nature
Immunology 2000;1:311-316.
Ludewig B, Odermatt O, Landmann S, Hengartner H, and Zinkerngel R M. Dendritic
cells induce autoimmune diabetes and maintain disease via de novo formation of
local lymphoid tissue. J.Exp.Med 1998;188:1493-1501.
Melero I, Vile R G and Colombo M P. Feeding dendritic cells with tumor antigens:
self-service buffet or a la carte? Gene Therapy 2000;7:1167-1170
Moser M, and Murphy K.M, Dendritic cell regulation of T
H1- T
H2 development.
Nature Immunology 2000;199-205.
Muzio M, Bosisio D, Polentarutti N, D’amico G, Stoppacciaro A, Mancinelli R, Veer C
V, Penton-Rol G, .Ruco L P, Allavena P, and Mantovani A. Differential
expression and regulation of Toll-like receptors(TLR) in human leukocytes:
Selective expression of TLR3 in dendritic cells. J. Immunol 2000;164:5998-6004.
Nakano H, Shindo M, Sakon S, Nishinaka S, Mihara M, Yagita H and Okumura K.
Differential regulation of IkB kinase a and b by two upstream kinases,
NF-kB-inducing kinase and mitogen-activated protein kinase/ERK kinase kinase-1. Proc
Natl Acad Sci USA 1998, 95:3537-3542.
Nobes C and Marsh M. Dendritic cells:New roles for Cdc42 and Rac in antigen uptake?
Current Biol 2000;10:R739-R741.
Patterson S. Flexibility and cooperation among dendritic cells. Nature Immunol 2000,
1:273-274.
Palucka K and Banchereau J. Dendritic cells: a link between innate and adaptive
immunity. J. Clin. Immunol. 1999, 19: 12-25.
Palucka K and Banchereau J. Linking innate and adaptive immunity. Nature
Medicine1999;5:868-870.
Porgador A, Snyder D, Gilboa E. Induction of antitumor immunity using bone
marrow-generated dendritic cells. J. Immunol. 1996; 156:2918-2926.
Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, Granucci F,
Kraehenbuhi J-P and Ricciardi-Castagnoli P. Dendritic cells express tight junction
proteins and penetrate gut epithelial monolayers to sample bacteria. Nature
Immunity 2001, 2:361-367.
Schuler G, Steinman RM. Dendritic cells as adjuvants for immune-mediated resistance
to tumors. J. Exp. Med. 1997; 186:1183-1187.
Schulz O, Edwards AD, Schito M, Aliberti J, Manickasingham S, Sher A and e Sousa
CR. CD 40 triggering of heterodimeric IL-12 p70 production by dnedritic cells in
vivo requires a microbial priming signal. Immunity 2000, 13:453-462.
Steinman R M, Turley S S, Mellman I, and Inaba K. The induction of tolerance by
dendritic cells that have captured apoptotic cells. J.Exp.Med 2000;191:411-416.
Tang H.L.and Cyster J G Chemokine up-regulation and activated T cell Attraction by
maturing dendritic cells. Science 1999;284:819-821.
Viney JL, Mowat AM, O’Malley JM, Williamson E and Fanger NA. Expanding
dendritic cells in vivo enhances the induction of oral tolerance. J. Immunol. 1998,
160:5815-5825.
Williamson E, Westrich G M, and Viney J L. Modulating dendritic cells to optimize
mucosal immunization protocols. J. Immunol.1999;163:3668-3675.
Whelan M, Harnett M M, Houston K M, Patel V, Harnett W, .and Rigley. K P. . A
filarial nematode-Sscreted product signals dendritic cells to acquire a phenotype
that drives development of Th2 cells. J. Immunol. 2000;164;6453-6460.
Wong B.R.,Josien R,Choi Y. TRANCE is a TNF family member that regulates
dendritic cell and osteoclast function J. Leukocyte Biol. 1999,65:715-724.
Polysaccharide purified from Ganoderma lucidum induced
activation and maturation of human monocyte-derived
dendritic cells by the NF-
B and p38 mitogen-activated
protein kinase pathways
Yu-Li Lin,* Yu-Chih Liang,
†Shiuh-Sheng Lee,
‡and Bor-Luen Chiang*,
1*Graduate Institute of Clinical Medicine, College of Medicine, National Taiwan University, Taipei, Republic of
China;
†Graduate Institute of Biomedical Technology, College of Medicine, Taipei Medical University, Taiwan,
Republic of China; and
‡Department of Biochemistry, National Yang-Ming University, Taipei, Taiwan,
Republic of China
Abstract:
Ganoderma lucidum, a fungus native
to China, has been widely used to promote health
and longevity in the Chinese. The polysaccharide
component with a branched (1
36)--D-glucan
moiety of G. lucidum (PS-G) has been reported to
exert anti-tumor activity and activation of
natu-ral killer cells. In this study, we investigated the
effects of PS-G on human monocyte-derived
den-dritic cells (DC). Treatment of DC with PS-G
resulted in the enhanced cell-surface expression
of CD80, CD86, CD83, CD40, CD54, and
hu-man leukocyte antigen (HLA)-DR, as well as the
enhanced production of interleukin (IL)-12p70,
p40, and IL-10 and also IL-12p35, p40, and
IL-10 mRNA expression, and the capacity for
endocytosis was suppressed in DC. In addition,
treatment of DC with PS-G resulted in enhanced
T cell-stimulatory capacity and increased T cell
secretion of interferon-
␥ and IL-10.
Neutraliza-tion with antibodies against Toll-like receptor
(TLR)-4 inhibited the PS-G-induced production
of IL-12 p40 and IL-10, suggesting a vital role
for TLR-4 in signaling DC upon incubation with
PS-G. Further study showed that PS-G was able
to augment inhibitor of
B (IB) kinase and
nuclear factor (NF)-
B activity and also IB␣
and
p38
mitogen-activated
protein
kinase
(MAPK) phosphorylation. Further, inhibition of
NF-
B by helenalin and p38 MAPK by SB98059
prevented the effects of PS-G in the expression
of CD80, CD86, CD83, CD40, CD54, and
HLA-DR and production of IL-12p70, p40, and
IL-10 in various degrees. Taken together, our
data demonstrate that PS-G can effectively
pro-mote the activation and maturation of immature
DC, suggesting that PS-G may possess a potential
in regulating immune responses. J. Leukoc. Biol.
78: 533–543; 2005.
Key Words:
PS-G
䡠signal transduction
䡠T cells
䡠IL-10
䡠IL-12
INTRODUCTION
Ganoderma lucidum, a native fungus from China, has been
widely used in China and other Asian countries. G. lucidum
has been reported to be effective in modulating immune
func-tions and inhibiting tumor growth and in the treatment of
chronic hepatopathy, hypertension, and hyperglycemia [1]. The
polysaccharide from G. lucidum (PS-G) is a branched
(136)--D-glucan moiety. Studies have demonstrated the
antineo-plastic action of G. lucidum and attributed it to the activated
host immune response [2, 3]. PS-G has been reported to
enhance the cytotoxic activity of natural killer (NK) cells and
to increase tumor necrosis factor
␣ (TNF-␣) and interferon-␥
(IFN-
␥) release from macrophages and lymphocytes,
respec-tively [4, 5]. The polysaccharide component from G. lucidum
also has been reported to elicit antiapoptotic effects on
neu-trophils, and this action primarily depends on the activation of
Akt-regulated signaling pathways [6].
Dendritic cells (DC) are the most professional
antigen-pre-senting cells (APCs), whose primary function is to capture,
process, and present antigens to unprimed T cells [7].
Imma-ture DC reside in nonlymphoid tissues, where they can capImma-ture
and process antigens. Thereafter, DC migrate to the T cell
areas of lymphoid organs, where they lose the
antigen-process-ing activity and mature to become potent immunostimulatory
cells [8]. The induction of DC maturation is critical for the
induction of antigen-specific T lymphocyte responses and may
be essential for the development of human vaccines relying on
T cell immunity. Fully mature DC show a high surface
expres-sion of major histocompatibility complex (MHC) class II and
costimulatory molecules (CD40, CD80, and CD86) but a
de-creased capacity to internalize antigens [9]. Up-regulation of
CD83, a specific marker for DC maturation, also occurs [10].
Various stimuli, such as proinflammatory cytokines [e.g.,
TNF-
␣ and interleukin (IL)-1], CD40 ligation, bacterial
prod-1Correspondence: Department of Pediatrics, National Taiwan University
Hospital, No. 7, Chungshan South Road, Taipei, Taiwan, R.O.C. E-mail: [email protected]
ucts [e.g., lipopolysaccharide (LPS) and unmethylated DNA
CpG motif], and contact sensitizers, can induce DC maturation
in vivo and in vitro [11, 12]. Several reports have already
indicated that the nuclear transcription factor (NF)-
B also
plays an important role in DC maturation [13]. Another
intra-cellular component involved in DC maturation, the three major
mitogen-activated protein kinase (MAPK) signaling pathways
in mammals, including p38 MAPK, extracellular
signal-regu-lated kinases (ERK), and c-Jun N-terminal kinases (JNK), are
activated in DC on maturation induced by LPS or TNF-
␣ [14].
The exact effects of PS-G on human DC are yet to be defined.
In the present study, we first examined the molecular
mecha-nisms of PS-G on the activation and maturation of human
monocyte-derived DC.
MATERIALS AND METHODS
Reagents
Escherichia coli LPS (L8274, E. coli) and lipoteichoic acid (LTA; L2515, from Staphylococcus aureus) were purchased from Sigma Chemical Co. (St. Louis,
MO). Isotopes were obtained from Amersham Corp. (Arlington Heights, IL). Neutralization antibodies (without sodium azide) against Toll-like receptor (TLR)-2 and TLR-4 were purchased from eBiosciences (San Diego, CA), and helenalin, SB203580, PD98059, and JNK inhibitor II were purchased from Calbiochem (Germany). Treatment of immature DC with these inhibitors (helenalin, SB203580, PD98059, and JNK inhibitor II) before stimulation was performed for 60 min. These inhibitors were dissolved in dimethyl sulfoxide (DMSO), where a 0.1% (v/v) concentration of DMSO was used as a negative control whenever indicated.
PS-G purification from G. lucidum
As in our previous study [2], fruiting bodies of G. lucidum were washed, disintegrated, and extracted with boiling water for 8 –12 h. Hot water extract of
G. lucidum was fractionated into a polysaccharide fraction (alcohol-insoluble)
and nonpolysaccharide fraction (alcohol-soluble). The crude polysaccharide obtained was then passed through a gel-filtration Sephadex G 50 column (Pharmacia, Uppsala, Sweden) and was further purified by anion exchange chromatography with a column of diethylaminoethyl cellulose [1]. The PS-G was a protein-bound polysaccharide consisting of⬃95% polysaccharide and 5% peptides. To rule out possible endotoxin LPS contamination of PS-G samples, we determined LPS content by the chromogenic Limulus amebocyte lysate assay. We found that there was no detectable level of endotoxin (⬍0.10 endotoxin units/ml) in the PS-G samples.
Human DC generation
DC were generated from peripheral blood mononuclear cells (PBMC), as described previously [15, 16], with some modification. Briefly, PBMC were obtained from healthy donors by centrifugation with Ficoll-Hypaque (Pharma-cia), and the light-density fraction from the 42.5–50% interface was recovered. CD14⫹cells were purified by positive selection using anti-CD14⫹microbeads in conjunction with the MiniMACS system by following the manufacturer’s instructions (Miltenyi Biotec, Auburn, CA). The DC14⫹cells were cultured at 1⫻ 106cells per 1 ml RPMI 1640 containing 10% fetal calf serum in 24-well
plates (Costar, Cambridge, MA) with granulocyte macrophage-colony stimu-lating factor (GM-CSF; 800 U/ml) and IL-4 (500 U/ml). Fresh medium con-taining GM-CSF and IL-4 was added every 2–3 days. Human monocyte-derived DC were used routinely at day 6 of culture.
Determination of cytokine levels
The IL-12 p70, IL-12 p40, IL-10, and IFN-␥ in the culture supernatant from DC or T cell were assayed with an enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN), as per the manufacturer’s
Reverse transcriptase-polymerase chain reaction
(RT-PCR)
Total RNA was isolated from DC using TRIzol reagent (Life Technologies, Gaithersburg, MD) following the manufacturer’s instructions. Total RNA was converted to cDNA with Moloney-murine leukemia virus RT (Life Technolo-gies) at 42°C for 1 h. The amplification of IL-12 p35, IL-12 p40, and IL-10 cDNA was performed by incubating equivalents of cDNA with Super Taq DNA polymerase. The IL-12 p35 primers used were the forward primer 5 ⬘-GAGTC-CCGGGAAAGTCCTGCC-3⬘ and the reverse primer 5⬘-TCTGGCCTTCTG-GAGCATGTT-3⬘. The IL-12 p40 primers used were the forward primer 5⬘-GGGGTGACGTGCGGAGCTGCT-3⬘ and the reverse primer 5⬘-TCTTGC-CCTGGACCTGAACGC-3⬘. The IL-10 primers used were the forward primer 5⬘-TTTCTCTTGGAGCTTATTAAAG-3⬘ and the reverse primer 5⬘-AA-GACTTTCTTTCAAATGAAGG-3⬘ (Invitrogen, Carlsbad, CA). The cDNA se-quence of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was also amplified as a control using the following primers: 5 ⬘-CTCATGACCACAGTC-CATGC-3⬘ and 5⬘-CCCTGTTGCTGTAGCCAAAT-3⬘. These primers produced a 450-base pair product. A thermal cycle of 30 s at 94°C, 30 s at 52°C, and 1 min at 72°C was used for 35 cycles for IL-12 p35 and IL-12 p40. A thermal cycle of 30 s at 94°C, 30 s at 55°C, and 1 min at 72°C was used for 35 cycles for IL-10.
Intracellular staining of IL-10
Intracellular cytokine staining was performed by using the BD Cytofix/Cyto-perm kit (BD Biosciences, San Diego, CA). Briefly, 1⫻ 105CD3⫹T cells and
DC were incubated at a ratio of 5:1 at 37°C for 2 days. After adding the transport inhibitor monesine, the culture was incubated at 37°C for 2.5 h. After washing with staining buffer, cells were labeled with CD3-fluorescein isothio-cyanate (FITC) and permeabilized. Intracellular staining was performed with phycoerythrin-labeled IL-10 antibodies or isotype control.
Flow cytometric analysis
DC were harvested and washed with cold buffer [phosphate-buffered saline (PBS) containing 2% fetal calf serum (FCS) and 0.1% sodium azide]. Cells were then incubated in cold buffer. Subsequent stainings with monoclonal antibodies (mAb) or isotype-matched controls were performed for 30 min on ice. Stained cells were then washed twice and resuspended in cold buffer and analyzed with a FACSort cell analyzer (Becton Dickinson, San Jose, CA). More than 1 ⫻ 104cells were analyzed for each sample, and the results were
processed by using Cellquest software (Becton Dickinson).
FITC-labeled dextran uptake
Cultured DC were washed twice and resuspended in 1 ml RPMI 1640 sup-plemented with 10% FCS, 2 mM L-glutamine, 100 U/ml penicillin, 100 U/ml streptomycin, and 25 mM HEPES. The cells were then incubated with FITC-labeled dextran (0.2 mg/ml) at 4°C or 37°C for 1 h. Finally, the cells were washed thrice with cold buffer and analyzed with a FACSort cell analyzer, as described above.
Allogeneic mixed leukocyte reaction (MLR)
PBMC were obtained as described above, and CD3⫹T cells were purified from PBMC using magnetic beads (Miltenyi Biotec). Theallogeneic CD3⫹T cells obtained were distributed at 1⫻ 105cells per well and incubated for 5 days
in the presence of graded numbers of irradiated DC (3000 rad,137Cs source).
Tritiated thymidine (1Ci/well, New England Nuclear, Boston, MA) incorpo-ration for 6 h was determined with a liquid counter.
Neutralization experiments
Human DC were preincubated for 1 h with 20g/ml antibody solution of TLR-2 and TLR-4. LPS, LTA, and PS-G were then added for 15 h. The cell culture supernatants were collected and were analyzed for IL-12 p70, IL-12 p40, and IL-10 by ELISA.
Inhibitor of
B (IB) kinase (IKK) activity assay
[10% glycerol, 1% Triton X-100, 1 mM sodium orthovanadate, 1 mM EGTA, 5 mM EDTA, 10 mM NaF, 1 mM sodium pyrophosphate, 20 mM Tris-HCl, pH 7.9, 100M -glycerophosphate, 137 mM NaCl, 1 mM phenylmethylsulfonyl fluoride (PMSF), 10g/ml aprotinin, and 10 g/ml leupeptin] for 30 min at 4°C. The cell lysate was clarified by centrifugation at 12,000 g for 10 min at 4°C. Equal amounts of total cellular protein (100g) were immunoprecipitated with IKK1- and IKK2-specific antibody (Santa Cruz Biotechnology, CA) and protein A/G-PLUS agarose for 12 h at 4°C. Kinase assay was carried out in 45 l kinase buffer [40 mM Tris-NaOH, pH 7.5, 500 mM NaCl, 0.1% Nonidet P-40 (NP-40), 6 mM EDTA, 6 mM EGTA, 10 mM-glycerophosphate, 10 mM NaF, 10 mM p-nitrophenyl phosphate, 300M sodium orthovanadate, 1 mM benzamidine, 2M PMSF, 10 g/ml aprotinin, 1 g/ml leupeptin, and 1 mM dithiothreitol (DTT)] containing 5M cold adenosine 5⬘-triphosphate (ATP), 10 Ci [␥-32P] ATP (5000Ci/mmol, Amersham), and 1 g glutathione
S-transferase (GST)-IB␣ fusion protein (Santa Cruz Biotechnology) as
sub-strate and incubated for 20 min at 25°C. Each sample was mixed with 8l 5⫻ Laemmli’s loading buffer to stop the reaction, heated for 10 min at 100°C, and subjected to 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel electro-phoresis. The gels were dried, visualized by autoradiography, and quantified by densitometry (IS-1000, Digital Imaging System).
Western blotting
Total cellular extract was prepared using Gold lysis buffer. Total protein (50 g) was separated on 10% SDS-polyacrylamide minigels and transferred to Immobilon polyvinylidene difluoride membrane (Millipore, Bedford, MA). The membrane was incubated overnight at 4°C with 10% bovine serum albumin in PBS to block nonspecific immunoglobulins (Igs) and then incubated with anti-IB-P polyclonal and anti-␣-tubulin mAb (Santa Cruz Biotechnology) and p38-P, p42/44-P, p46/54-P, and total p38 polyclonal anti-body (Cell Signaling Technology, Beverly, MA).
Preparation of nuclear extracts and
electrophoretic mobility shift assay (EMSA)
Nuclear and cytoplasmic extracts were prepared as described previously [18]. At the end of the culture, the cells were suspended in hypotonic buffer A (10 mM HEPES, pH 7.6, 10 mM KCl, 0.1 mM EDTA, 1 mM DTT, 0.5 mM PMSF) for 10 min on ice and vortexed for 10 s. Nuclei were pelleted by centrifugation at 12,000 g for 20 s. The supernatants containing cytosolic proteins were collected. Pellets containing nuclei were resuspended in buffer C (20 mM HEPES, pH 7.6, 25% glycerol, 0.4 M NaCl, 1 mM EDTA, 1 mM DTT, 0.5 mM PMSF) for 30 min on ice. The supernatants containing nuclear proteins were collected by centrifugation at 12,000 g for 20 min and stored at –70°C. For EMSA, each 5g nuclear extract was mixed with the labeled, double-stranded NF-B oligonucleotide, 5⬘-AGTTGAGGGGACTTTCCCAGGC-3⬘, and incu-bated at room temperature for 20 min. The incubation mixture included 1g poly (dI-dC) in a binding buffer (25 mM HEPES, pH 7.9, 0.5 mM EDTA, 0.5
mM DTT, 1% NP-40, 5% glycerol, and 50 mM NaCl). The DNA-protein complex was electrophoresed on 4.5% nondenaturing polyacrylamide gels in 0.5⫻ Tris-boric acid EDTA buffer (0.0445 M Tris, 0.0445 M borate, 0.001 M EDTA). A double-stranded, mutated oligonucleotide, 5
⬘-AGTTGAGGC-GACTTTCCCAGGC-3⬘, was used to examine the specificity of the binding of NF-B to DNA. The specificity of binding was also examined by comparison with the unlabeled oligonucleotide.
Statistical analysis
The Student’s t-test was used to analyze the results, and a P value of less than 0.05 was considered to be statistically significant.
RESULTS
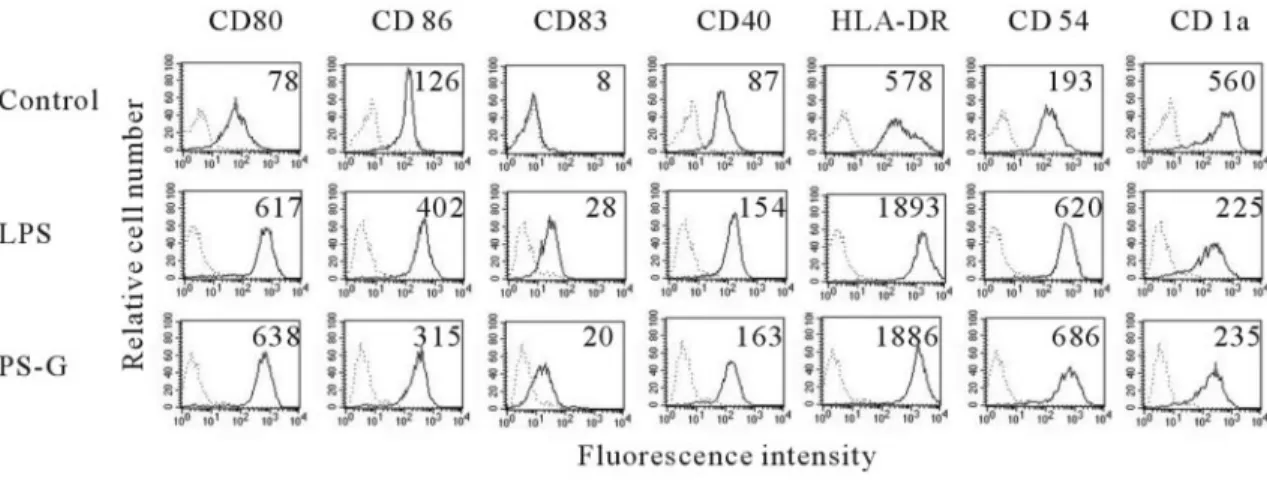
PS-G induces maturation of human
monoctye-derived DC
LPS has been described as an inducer of DC activation and
maturation [19]. Therefore, we use LPS as a positive control in
this study. To determine whether PS-G also can modulate the
development of human DC in vitro, we compared the phenocyte
of human DC treated with or without PS-G for 24 h. Our data
demonstrated that PS-G increased the presentation of CD80,
CD86, CD83, CD40, CD54, and MHC class II molecules on the
cell membrane of human DC (Fig. 1).
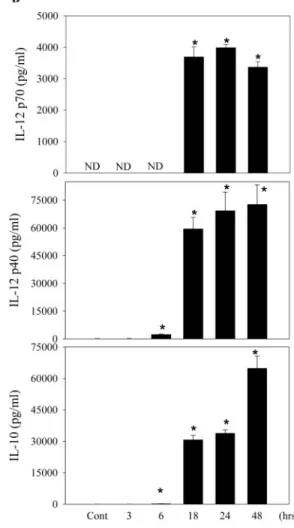
PS-G induces IL-12 p70, IL-12 p40, and IL-10
production in human DC
To determine whether PS-G can affect the cytokine production in
human DC, we compared cytokine concentrations in the
super-natants of DC cultured with different doses of PS-G, which
en-hanced the production of IL-12 p70, IL-12 p40, and IL-10 (Fig.
2A
). When human DC were treated with 10
g/ml PS-G for 3, 6,
18, 24, and 48 h, we found that PS-G significantly enhanced the
production of IL-12 p70, IL-12 p40, and IL-10 at 18, 24, and 48 h
(Fig. 2B). It was clear that the stimulatory effect of PS-G on IL-12
p70, IL-12 p40, and IL-10 production was dose- and
time-depen-dent in manner. To determine whether PS-G could affect IL-12
p35, IL-12 p40, and IL-10 mRNA expression, human DC were
activated with PS-G at indicated periods of time and assayed for
Fig. 1. The effect of PS-G and LPS on DC phenotype. Human DC were treated with PS-G (10g/ml), LPS (100 ng/ml), or medium alone for 24 h, and surface markers were analyzed by flow cytometry (dotted line, isotype control; solid line, specific mAb). The values shown in the flow cytometry profiles are the mean
IL-12 p35, IL-12 p40, and IL-10 mRNA expression by RT-PCR.
We found that significantly higher levels of IL-12 p35, IL-12 p40,
and IL-10 mRNA were expressed at 6 h, 6 h, and 18 h,
respec-tively, in human DC, especially highly expressed IL-12 p40
mRNA (Fig. 2C). In unstimulated DC, there was no detectable
IL-12 p35, IL-12 p40, and IL-10 mRNA.
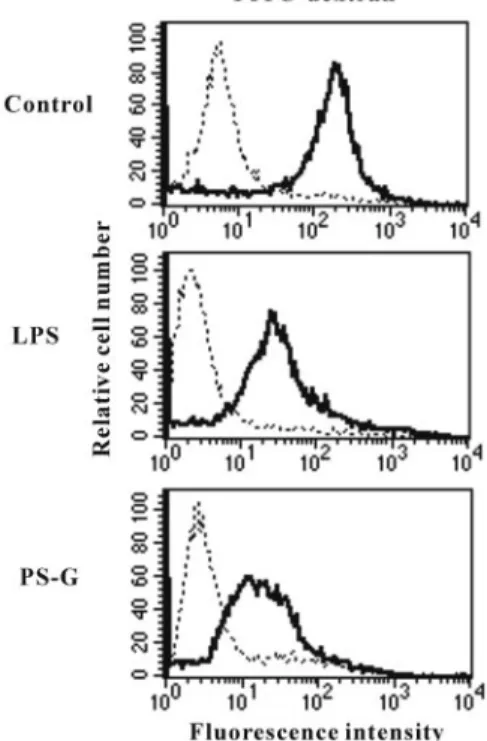
PS-G down-regulation of endocytotic activity of
human DC
Immature DC capture and process antigens via their high
activities of antigens and mature into potent
immunostimu-latory APCs during differentiation [7]. The uptake of
FITC-dextran is known to be maximal in the immature
monocyte-derived DC and occurs by a combination of
macropinocy-tosis and binding to the mannose receptor. Previous studies
have shown that the endocytic capacity of DC is suppressed
by LPS during their maturation process. Thus, we tested
whether PS-G affected the uptake of FITC-labeled dextran
by human DC. In our study, we demonstrated a reduction in
FITC-dextran uptake when human DC were matured with
Fig. 2. PS-G induces IL-12 p70, IL-12 p35, and IL-10 productionin human monocyte-derived DC. (A) Human DC were cultured for 24 h in the presence of 100 ng/ml LPS or various concentrations of PS-G. At the end of the incubation time, the culture medium was collected for cytokine assay by ELISA. (B) Human DC were incu-bated with PS-G (10g/ml) for the indicated period of time. At the end of the incubation time, IL-12 p70, IL-12 p35, and IL-10 production was subsequently analyzed by ELISA. Each data repre-sent the mean⫾SEfor three determinations. Statistical analysis concerns unstimulated versus stimulated DC. *, P⬍ 0.05. ND, Not determined. (C) RT-PCR analysis of mRNA expression of IL-12 p35, IL-12 p40, and IL-10. DC was incubated in the presence of PS-G (10 g/ml) for 3, 6, 18, and 24 h. This experiment was repeated three times with similar results. GAPDH, glyceraldehyde-3-phosphate dehydrogenase. Lane M, marker.
Enhancement of T cell activation by
PS-G-treated human DC
Mature DC have the capacity to induce proliferation in
allo-genic T cells at a much higher level than immature DC [8]. In
human DC, we found that PS-G up-regulated cell-surface
markers, increased IL-12 production, and induced the
activa-tion of NF-
B. To test whether this maturation is sufficient to
promote activation of naive T cells, DC were treated with LPS
or PS-G. These cells were then used to activate allogenic, naive
T cells. The results presented in Figure 4A show that
PS-G-treated DC enhanced T cell activation, as evidenced by the
secretion of IFN-
␥ in the culture supernatant (Fig. 4B). The
IFN-
␥ production induced under these experimental
condi-tions was far higher than that seen following LPS treatment of
DC, especially at low DC/T cells ratios. It is interesting that we
demonstrated that PS-G-treated DC enhanced T cell secretion
of IL-10, except for LPS-treated DC, which were not able to
enhance T cell secretion of IL-10 in the supernatant (Fig. 4C).
To investigate if IL-10 is produced by the activated T cells in
this experiment, we performed intracellular cytokine staining
for IL-10 on CD3
⫹T cells (
⬎95% purity by flow), which were
cocultured with autologous DC. Intracellular cytokine staining
results showed that PS-G-treated DC enhanced the T cell
production of IL-10, such that the percentage gated of
double-positive CD3 and IL-10 is 6.04%. In DC alone and LPS-treated
DC, the percentages gated of double-positive CD3
⫹and IL-10
are 1.74% and 1.51%, respectively. Therefore, intracellular
IL-10 staining of T cells in the PS-G-treated DC group is
significantly higher than in DC alone or in the LPS-treated DC
group. In the LPS group or the PS-G-treated group, DC could
not induce T cell secretion of IL-4 cytokine (data not shown).
Fig. 3. PS-G on the endocytotic capacities of human DC. At day 6, immatureDC were stimulated with medium alone, LPS (100 ng/ml), or PS-G (10g/ml) for 24 h, and cells were then incubated with FITC-dextran for 1 h at 4°C (dotted lines) or 37°C (solid lines). This experiment was repeated three times with similar results.
Fig. 4. PS-G enhances T cells response. (A) Immature DC were stimulated with LPS (100 ng/ml) or PS-G (10g/ml) for 24 h. Allogeneic T cell proliferation was measured after 5 days of coculture with DC. These data are means⫾SEMof triplicates and representative of three independent experi-ments. Supernatants were analyzed for (B) IFN-␥ and (C) IL-10, produced by activated T cells after 2 days of culture.
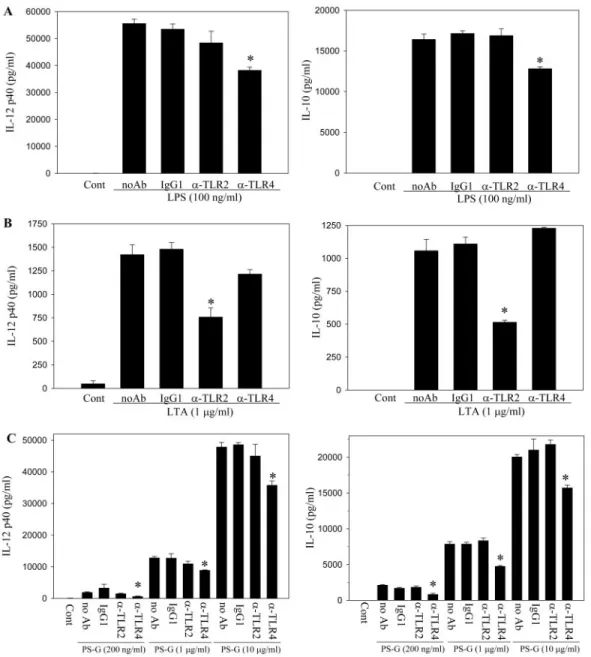
PS-G induces IL-12 p40 and IL-10 synthesis
through TLR-4
TLRs have been shown to be involved in the antifungal defense
mechanism in Drosophila and the antibacterial defense in
humans. To determine the involvement of these receptors in
the interaction of DC with PS-G, neutralization experiments
were performed. Cell-surface TLR-2 and TLR-4 receptors were
blocked by neutralizing concentrations of their respective
an-tibodes before DC were treated with LPS, LTA, or PS-G. We
showed positive and negative controls for the neutralization of
TLR-4 (by using LPS) and TLR-2 (by using LTA) in Figure 5,
A
and B. In Figure 5C, we demonstrated that the addition of an
anti-TLR-4 mAb to human DC blocked PS-G (100 ng/ml, 1
g/ml, and 10 g/ml)-induced IL-12 p40 production ⬃70%,
31%, and 47%, respectively, and IL-10 production,
⬃55%,
39%, and 20%, respectively, but the addition of an anti-TLR-2
mAb failed to inhibit PS-G-induced IL-12 p40 and IL-10
production.
PS-G induces IKK activity and phosphorylation
of I
B␣ in human DC
As the activation of IKK activity is necessary for I
B␣
phos-phorylation, the effect of PS-G on IKK activity was likewise
studied. Human DC were treated with PS-G (10
g/ml) for the
indicated periods of time. To measure IKK1 activity directly in
human DC, IKK1 and IKK2 proteins were immunoprecipitated
from cell extracts, and the kinase activity in the
immunocom-plex was assayed using recombinant GST-I
B␣ (1–317) as a
substrate. Figure 6A illustrates the relative effect on IKK
activity. After stimulation with PS-G, the GST-I
B␣ fusion
Fig. 5. Neutralization with TLR-4 mAb inhibits the synthesis of IL-12 p40 and IL-10 in PS-G-treated, human DC, which were preincubated with 20g/ml anti-TLR-2, TLR-4, and IgG1 antibodies separately for 1 h. DC were then challenged with LPS (A), LTA (B), or PS-G (C) for 15 h. The cell culture supernatants were collected for IL-12 p40 and IL-10 analysis. Data are represented as mean⫾SE. Significant difference between DC treated with antibodies and no antibodies
protein was strongly phosphorylated at 30 min, indicating the
stimulation of IKK activity in human DC.
NF-
B is one molecular family whose activation is
associ-ated with DC maturation. NF-
B normally binds to IB␣,
which impedes NF-
B nuclear translocation from the
cyto-plasm to the nucleus. Once cells are exposed to inflammatory
stimuli, including LPS and TNF-
␣, IB␣ is phosphorylated,
leading to I
B␣ degradation and nuclear translocation of
NF-B. We thus examined whether PS-G had any effect on IB␣
phosphorylation. The cytoplasmic levels of I
B␣-P protein
were examined by Western blot analysis. LPS and PS-G
in-duced the phosphorylation of I
B␣. After 60 min from the
activation of human DC with PS-G, the cytosolic I
B␣ protein
was significantly phosphorylated (Fig. 6B).
PS-G induces NF-
B activation
DC maturation derived by LPS has been clearly associated with
NF-
B activation. To determine whether PS-G uses a similar
activation pathway, we monitored its ability to activate NF-
B
translocation into the nucleus. DC were cultured in the
pres-ence of PS-G for 2 h, and nuclear extracts were analyzed for
NF-
B binding by the EMSA. As shown in Figure 7, PS-G
was able to induce NF-
B translocation and activation.
Iden-tical results were obtained after treatment of DC with LPS. The
binding of NF-
B was specific and could be blocked by
unla-beled, competing NF-
B oligonucleotide.
PS-G induces phosphorylation of members of
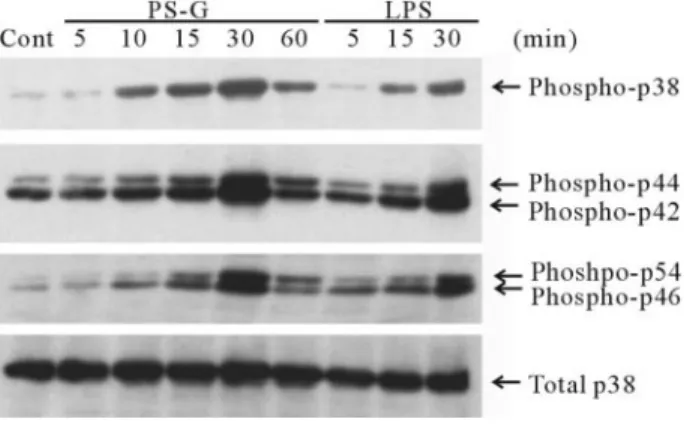
the three MAPK families in human DC
MAPK is a serine and threonine protein kinase whose activities
are up-regulated through tyrosine and threonine residue
phos-phorylation by its upstream regulators [20, 21]. This
experi-ment focused on p38 MAPK, p42/44 ERK, and p46/54 JNK to
PS-G signaling. Human DC were stimulated with PS-G or none
at all, and the level of MAPK phosphorylations was assessed by
Western blotting with respective antityrosine-phosphorylated
MAPK mAb. Total p38 mAb was used for internal control.
Results presented in Figure 8 show that PS-G induced the
phosphorylation of all MAPK tested, especially in inducing a
higher p38 phosphorylation at 30 min. The total amount of p38
was unchanged following stimulation.
Fig. 6. PS-G induced IKK activity and IB␣ phosphorylation in human DC. (A) Human DC were treated with PS-G (10g/ml) for the indicated time periods and collected the total cell lysates for IKK activity assay. Immuno-precipitated IKK was incubated with [␥-32P] ATP and GST-IB fusion protein,
as substrates performed the kinase activity assay as described in Materials and Methods, and P-GST-IB␣ is shown. (B) Human monocyte-derived DC were treated with LPS (100 ng/ml) for 45 min or PS-G (10g/ml) for the indicated time periods. Cytosolic fractions were prepared and analyzed for the phosphor-ylation level of IB by Western blotting. The lower panel shows the blot probed for␣-tubulin to demonstrate equal loading of samples. This experiment was repeated three times with similar results.
Fig. 7. PS-G induces NF-B activation. Human monocyte-derived DC were treated with LPS (100 ng/ml) or PS-G (10 g/ml) for 2 h or remained unstimulated, and nuclear fractions were prepared and analyzed for NF-B binding activity by EMSA. To assess the specificity of the binding, 100-fold excess of cold NF-B probe or mutant probe was added to the LPS condition. This experiment was repeated three times with similar results.
Fig. 8. PS-G induces the phosphorylation of p38 MAPK, p42/44 ERK, and p46/52 JNK kinase. Human monocyte-derived DC were treated with PS-G (10 g/ml) inhibitors for the indicated time periods and then collected the cell lysate, and the level of MAPK phosphorylations was assessed by Western blotting with respective antityrosine-phosphorylated MAPK mAb, and total
Inhibition of NF-
B and MAPK prevents the
maturation changes induced by PS-G
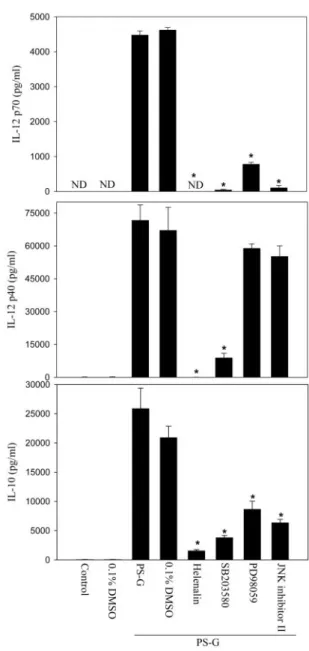
PS-G-treated DC produced many cytokines, including IL-12
p70, IL-12 p40, and IL-10, during maturation (Fig. 2, A and
B). We investigated whether the PS-G-induced secretions of
IL-12 p70, IL-12 p40, and IL-10 were affected by the
inhibi-tors of NF-
B, p38 MAPK, p42/44 ERK, and p46/54 JNK.
Immature human DC were pretreated with helenalin (a specific
blocker of NF-
B), SB203580 (a specific blocker of p38
MAPK), PD98059 (an inhibitor of the ERK pathway), or JNK
inhibitor II (an inhibitor of the JNK pathway) for 1 h and
subsequently stimulated with PS-G for 24 h. The production of
IL-12 p70, IL-12 p40, and IL-10 was quantified by means of
ELISA. PS-G induced significant production of IL-12 p70,
IL-12 p40, and IL-10, and these cytokine productions were
abrogated significantly by helenalin and SB203580 (Fig. 9). In
contrast, PD98059 and JNK inhibitor II down-regulated IL-12
p70 and IL-10 production but only had little effect on the
inhibition of IL-12 p40 production induced by PS-G.
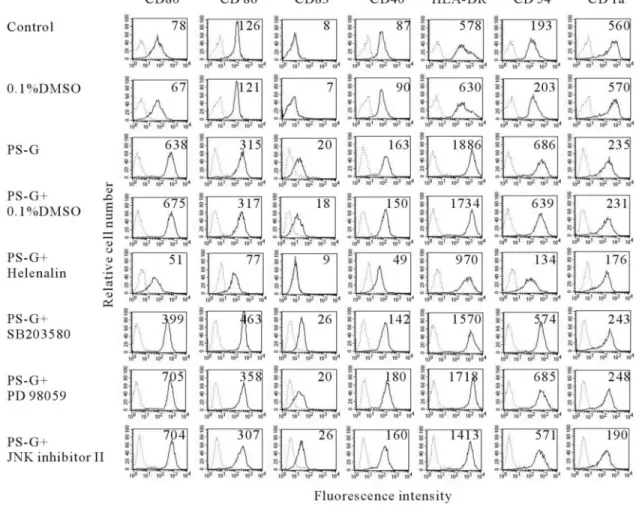
To further examine the involvement of NF-
B, p38 MAPK,
ERK, and JNK in the PS-G-induced expression of
costimula-tory and antigen-presenting molecules, NF-
B inhibitor,
hel-enalin, p38 MAPK inhibitor, SB203580, ERK pathway
inhib-itor, PD98059, and JNK pathway inhibinhib-itor, JNK inhibitor II, in
the expression of costimulatory and adhesion molecules as well
as HLA-DR, were investigated. Blocking the NF-
B pathway
with helenalin significantly inhibited the PS-G and induced the
up-regulation of CD80, CD86, CD83, CD40, CD54, and
HLA-DR (Fig. 10). In contrast, blocking the p38 MAPK and
JNK pathway with SB203580 and JNK inhibitor II,
respec-tively, had little effect on CD80, CD86, CD83, CD40, CD54,
and HLA-DR expression. PD98059, a specific inhibitor of
ERK, had no effect on these costimulatory molecules and MHC
class II expression. These results show that certain features of
human monocyte-derived DC maturation are regulated by
sig-naling via NF-
B and p38 MAPK and imply that different
aspects of the maturation process induced by PS-G may be
regulated by distinct signal transduction pathways.
DISCUSSION
The biological process of DC maturation represents a crucial
step in the initiation of adaptive immune responses [22]. This
process is regulated by various extracellular stimuli, including
cytokines, bacterial products, and membrane-bound ligands
[10, 23]. DC maturation is accompanied by changes in their
morphological, phenotypic, and functional properties [24].
Re-cently, several researchers have demonstrated the
immuno-modulatory effects of polysaccharides purified from G. lucidum
on T lymphocytes [25]. Cao and Lin [26] showed that G.
lucidum polysaccharides could promote the maturation and
function of murine DC. These results are similar to our finding
in human DC. However, little is known about the molecular
mechanisms responsible for the regulation of DC in their
activation and maturation states by PS-G. In this study, we
were the first to demonstrate the PS-G-induced morphological,
derived DC. PS-G promoted the maturation of DC, and mature
DC demonstrated characteristic morphology with enlarged size
and numerous cytoplasmic processes, which gave rise to a
stellate appearance (data not shown). Maturation of DC was
characterized by a decreased antigen-processing capacity, an
increased cell-surface expression of MHC class II molecule
and costimulatory molecules CD80, CD86, and CD40, and the
secretion of IL-12, which primed a strong stimulation of T
lymphocyte growth and differentiation. The CD83 marker for
Fig. 9. The effect of inhibiting the NF-B, p38 MAPK, ERK1/2, or JNK pathways on the PS-G-induced up-regulation of IL-12 p70, IL-12 p40, and IL-10 production in human monocyte-derived DC. Human DC were pretreated with 0.1% DMSO, 10M helenalin (a specific blocker of NF-B), 20 M SB203580 (a specific blocker of p38 MAPK), 50M PD98059 (an inhibitor of the ERK1/2 pathway), or 20M JNK inhibitor II (an inhibitor of the JNK pathway) for 1 h and then incubated with 10g/ml PS-G for 24 h. At the end of the incubation time, the supernatant was collected for IL-12 p70, IL-12 p 35, and IL-10 production by ELISA. Significant difference between DC treated with PS-G alone and pretreated with inhibitors is indicated by P⬍ 0.05 (*). ND, Not determined.In immune responses, IL-12 plays a central role as a link
between the innate and adaptive immune systems [27]. Thus,
IL-12 induces and promotes NK and T cells to generate IFN-
␥
and lytic activity. In addition, IL-12 polarizes the immune
system toward a primary T helper cell type 1 (Th1) response. In
this study, we found out that LPS and PS-G can induce IL-10
and IL-12 production in human DC. IL-10 is a pleiotropic
cytokine produced by DC, T cells, and macrophages and
anti-inflammatory and immunosuppressive properties [24]. We
suggested that when IL-12 p40 is overexpressed in
PS-G-treated human DC, IL-10 could act as a feedback regulatory
role, although LPS has the similar effect in human DC, but the
cytotoxicity of LPS is higher than PS-G. Therefore, PS-G is a
safe immune modulator for human DC. IFN-
␥ and IL-10
cy-tokines were induced in MLR by PS-G-treated human DC. In
contrast to PS-G, only IFN-
␥ cytokine was induced in MLR by
LPS-treated, human DC. Therefore, LPS was described as a
Th1 inducer. Our experimental data show that under some
conditions, PS-G can induce a Th1 differentiation or promote
the differentiation of naı¨ve T cells for the secretion of IL-10.
However, although DC are widely regarded as the most potent
tant role in inducing immune tolerance [28] and regulating
Th1/Th2 immunity balance [29]. Furthermore, these immature
DC could also be alternatively activated and induced to exert
suppressive effects [30].
TLRs have been identified in humans as an important
com-ponent of innate immunity against microbial pathogens. LPS is
recognized by TLR-4 in DC. The effects of TNF-
␣ are
medi-ated by two distinct cell-surface receptors, TNF-R1 and
TNR-R2, where TNF-R1 has been implicated in TNF-
␣-induced
phenotypic and functional changes in DC [10]. Recent reports
show that LPS and TNF-
␣, two potent DC maturation factors,
induced the NF-
B activation and phosphorylation of p38,
ERK1/2, and p46/54 JNK in monocyte-derived DC [31]. Our
results demonstrated that PS-G activated NF-
B and all three
MAPK pathways during maturation. Neutralization
experi-ments with antibodies blocking TLR-2 and TLR-4 further
demonstrated that TLR-4 played a critical role in the signal
transduction cascade induced by PS-G. The blocking effect in
high concentration of PS-G (10
g/ml)-treated DC is not better
than in low concentration (200 ng/ml). However, the
percent-age of inhibition by anti-TLR-4 antibody was similar between
Fig. 10. The effect of inhibiting the NF-B, p38 MAPK, ERK1/2, or JNK pathways on the PS-G-induced up-regulation of CD80, CD86, CD83, CD40, CD54, and HLA-DR in human monocyte-derived DC. Day 6 immature DC were pretreated with 0.1% DMSO, 10M helenalin (a specific blocker of NF-B), 20 M SB203580 (a specific blocker of p38 MAPK), 50M PD98059 (an inhibitor of the ERK1/2 pathway), or 20 M JNK inhibitor II (an inhibitor of the JNK pathway) for 1 h before the addition of PS-G (10g/ml) for 24 h. The cell-surface expression of CD80, CD86, CD83, CD40, CD54, and HLA-DR was then measured using the flow cytometry (dotted line, isotype control; solid line, specific mAb). The values shown in the flow cytometry profiles are the MFI indexes. These results are representative of three independent experiments with similar results.zymosan, as zymosan particles are recognized simultaneously
by dectin-1 and TLR-2 [32–34]. Recent reports show that
NF-
B is responsible for LPS-induced DC maturation in an in
vitro murine model [35] and that cytokine-induced maturation
of human DC results in increased NF-
B nuclear translocation
[36]. Many proinflammatory cytokines displayed NF-
B-re-sponsive elements in their promoters, conferring a major role
on immune responses [37]. Moreover, the p38 MAPK pathway
has been shown to contribute to NF-
B-mediated
transactiva-tion [38, 39]. Little is known about the signal transductransactiva-tion
pathways involved in the maturation of human
monocyte-de-rived DC by PS-G. We demonstrated that the NF-
B, p38
MAPK, ERK1/2, and p46/54 JNK pathways are activated when
immature human DC are exposed to PS-G, suggesting a role of
these pathways in the maturation process. The promoters of
human (h)IL-12 p35 and hIL-12 p40 gene contain
B-binding
sites [40]. It likely that NF-
B is also involved in the IL-12 p35
and IL-12 p40 expression. The lack of
B-binding sites in the
hIL-10 promoter makes it unlikely that NF-
B is involved in
IL-10 regulation [41]. Recently, it has been suggested that p38
MAPK is involved in the regulation of IL-10 production [42].
Early phosphorylation of p38 MAPK, ERK1/2, and p46/54
JNK was investigated in PS-G-treated, human
monocyte-de-rived DC. Our results corroborate recent reports using the
murine models, as well as human DC in vitro models showing
activation of all three MAPK pathways during maturation [35,
43]. The availability of specific inhibitory drugs for the NF-
B,
p38 MAPK, ERK, and JNK pathways prompted us to
investi-gate the respective roles of the NF-
B and these MAPK in DC
maturation. In cytokines analysis, pretreatment of helenalin
and SB203580 significantly inhibited the IL-12 p70, IL-12
p40, and IL-10 productions in PS-G-treated, human DC. In
contrast, PD98059 and JNK inhibitor II were shown to inhibit
IL-12 p70 and IL-10 production, although we only observed a
little inhibitory effect of these compounds in the up-regulation
of IL-12 p40 in the process of DC maturation triggered by
PS-G. Concerning costimulatory molecules and MHC class II
expression, helenalin-pretreated human DC were able to
com-pletely suppress these molecules’ expression induced by PS-G.
The inhibition of p38 MAPK and p46/54 JNK by SB203580
and JNK inhibitor II, respectively, before PS-G stimulation had
a weak effect on the CD80, CD86, CD83, CD40, CD54, and
MHC class II expression induced during DC maturation.
PD98059 had no effect on the costimulatory molecules and
MHC class II expression in the process of DC maturation
triggered by PS-G. Moreover, the inhibitory effects of these
inhibitors were not a result of nonspecific toxicity, as these
inhibitors did not modify the viability of DC (data not shown).
Collectively, these results show that the NF-
B and p38 MAPK
pathways play critical roles in the initiation of DC maturation.
The human CD86 promoter has been cloned recently, and two
canonical NF-
B binding sites have been revealed [44]. One of
them is essential for the Th-induced CD86 gene transcription.
Moreover, NF-
B activation has been shown previously to
drive CD 80 transcription [45]. A recent report describes the
generation of MAPK kinase 3-deficient mice to study the role
of the p38 MAPK pathway in vivo [46]. Using this animal
model, the authors showed that p38 MAPK is required for the
different signal transduction pathways regulate the different
aspects of DC maturation.
In conclusion, we demonstrated that PS-G can effectively
and rapidly induce the significant activation and maturation of
human DC by the NF-
B and p38 MAPK pathways. Therefore,
PS-G is a good and potential part of the treatment regimen to
regulate host immune responses.
ACKNOWLEDGMENTS
This study was supported by the National Science Council NSC
92-2314-B-002-201. The authors extend their gratitude to
Ya-Hui Chuang for the discussions and comments about this
work and manuscript.
REFERENCES
1. Miyazaki, T., Nishijima, M. (1981) Studies on fungal polysaccharides, XXVII. Structural examination of a water-soluble, anti-tumor polysaccha-ride of Ganoderma lucidum. Chem. Pharm. Bull. (Tokyo) 29, 3611–3616. 2. Wang, S. Y., Hsu, M. L., Hsu, H. C., Tzeng, C. H., Lee, S. S., Shiao, M. S., Ho, C. K. (1997) The anti-tumor effect of Ganoderma lucidium is mediated by cytokines released from activated macrophages and T lymphocytes. Int.
J. Cancer 70, 699 –705.
3. Furusawa, E., Chou, S. C., Furusawa, S., Hirazami, A., Dang, Y. (1992) Anti-tumor activity of Ganoderma lucidum, an edible mushroom, on intraperitoneally implanted Lewis lung carcinoma in synergenic mice.
Phytother. Res. 6, 300 –304.
4. Lee, S. S., Wei, Y. H., Chen, C. F., Wang, S. Y., Chen, K. Y. (1995) Antitumor effects of Ganoderma lucidum. J. Chin. Med. 6, 1–12. 5. Won, S. J., Lee, S. S., Ke, Y. H., Lin, M. T. (1989) Enhancement of spenic
NK cytotoxic activity by extracts of Ganoderma lucidum mycelium in mice. J. Biomed. Lab. Sci. 2, 201–213.
6. Hsu, M. J., Lee, S. S., Lin, W. W. (2002) Polysaccharide purified from
Ganoderma lucidum inhibits spontaneous and Fas-mediated apoptosis in
human neutrophils through activation of the phosphatidylinositol 3 kinase/ Akt signaling pathway. J. Leukoc. Biol. 72, 207–216.
7. Banchereau, J., Steinman, R. M. (1998) Dendritic cells and the control of immunity. Nature 392, 245–252.
8. Cella, M., Sallusto, F., Lanzavecchia, A. (1997) Origin, maturation and antigen presenting function of dendritic cells. Curr. Opin. Immunol. 9, 10 –16.
9. Cella, M., Pinet Engering, A., Pieters, J., Lanzavecchia, A. (1997) Inflam-matory stimuli induce accumulation of MHC class II complexes on den-dritic cells. Nature 388, 782–787.
10. Sallusto, F., Cella, M., Danieli, C., Lanzavecchia, A. (1995) Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the major histocompatibility complex class II compart-ment: downregulation by cytokines and bacterial products. J. Exp. Med.
182,389 – 400.
11. Caux, C., Massacrier, C., Vanbervliet, B., Dubois, B., van Kooten, C., Durand, I., Banchereau, J. (1994) Activation of human dendritic cells through CD40 cross-linking. J. Exp. Med. 180, 1263–1272.
12. Jakob, T., Walker, P. S., Krieg, A. M., Udey, M. C., Vogel, J. C. (1998) Activation of cutaneous dendritic cells by CpG-containing oligode-oxynucleotides: a role for dendritic cells in the augmentation of Th1 responses by immunostimulatory DNA. J. Immunol. 161, 3042–3049. 13. Yoshimura, S., Bondeson, J., Foxwell, B. M., Brennan, F. M., Feldmann,
M. (2001) Effective antigen presentation by dendritic cells is NF-B dependent: coordinate regulation of MHC, co-stimulatory molecules and cytokines. Int. Immunol. 13, 675– 683.
14. Ardeshna, K. M., Pizzey, A. R., Devereux, S., Khwaja, A. (2000) The PI3 kinase, p38 SAP kinase, and NF-B signal transduction pathways are involved in the survival and maturation of lipopolysaccharide-stimulated human monocyte-derived dendritic cells. Blood 96, 1039 –1046. 15. Sallusto, F., Lanzavecchia, A. (1994) Efficent presentation of soluble
antigen by cultured human dendritic cells is maintained by granulocyte/ macrophage colony-stimulation factor plus interleukin 4 and