Investigating neurochemical techniques for assessment of nerve
regenerates within polymer guides
Chung-Chia Chen1,2, Chu-Ying Chan2, Li-Shu Chen2,
Fuu-Jen Tsai2, Chun-Hsu Yao2,3†,Chin-Chuan Tsai4,5†, Yueh-Sheng Chen2,3*†
1
Department of Chinese Medicine, Taipei City Hospital, Taipei, Taiwan 2
Lab of Biomaterials, School of Chinese Medicine, China Medical University, Taichung, Taiwan
3
Department of Biomedical Imaging and Radiological Science, China Medical University, Taichung, Taiwan
4
School of Chinese Medicine for Post-Baccalaureate, I-Shou University, Kaohsiung, Taiwan
5
Chinese Medicine Department, E-DA Hospital, Kaohsiung, Taiwan
Running title: Neurochemical assessment of regenerated nerves
†These authors contributed equally to this work.
*Corresponding author: Yueh-Sheng Chen, PhD
Lab of Biomaterials, School of Chinese Medicine,
China Medical University, Taichung, Taiwan
Tel.: 886-4-22053366 ext. 3308; Fax: 886-4-22032295
Abstract
This research evaluated the use of proteins associated with neurite outgrowth,
synapse formation, and Schwann cell proliferation as surrogate measures for
peripheral nerve regeneration. Rat sciatic nerve regeneration was across a 10-mm gap
in silicone rubber conduits (SRCs), genipin-crosslinked gelatin conduits (GGCs), and
porous GGCs (PGGCs). After 8 weeks, axonal growth of regenerated nerves was
determined using light microscopy and computer-based quantitative image analysis.
Expression of the axonal growth associated protein 43 (GAP-43), the synaptic protein
synapsin I, and the transforming growth factors type β (TGF-βs) in regenerated
nerves were assessed simultaneously by Western blot. As a result, the nerve
regenerates in the silicone rubber conduits had a relatively larger area containing more
myelinated axons compared to those in the PGGCs and the GGCs. The levels of
GAP-43 and synapsin I, but not TGF-β, correlated well with axonal growth in the
regenerated nerves. These data suggest that SRCs could provide a more stable and
suitable growth environment for regenerating axons than PGGCs and GGCs. In
addition, GAP-43 and synapsin I may be a useful indicator of the status of
regenerating axons in bridging conduits with different construction and physical
properties.
1. Introduction
Tissue engineering has grown as an interdisciplinary field combination of cells,
engineering and biomaterials [1]. Specific biochemical functions were performed
using cells within an artificially-created support system to improve or replace
biological tissues, such as heart [2], skin [3], and bone tissues [4]. Recently, the use of
in vitro systems to assess developmental neurology is gaining momentum, and a
number of neuronal models have been used for examining potential biomaterials for
nerve regeneration. In particular, phenochromocytoma and Schwann cells have been
widely used to screen biocompatible materials as a model of neuronal proliferation
and differentiation [5-8]. However, there are limitations to the use of these cell lines.
First, they do not develop true axons, dendrites, or synapses with each other. In
addition, genetic drift with respect to repeated cell passage with a concomitant change
in phenotype could happen [9]. Therefore, in vivo models are necessary to be used for
revealing the real regeneration process in the developing nervous system.
Nerve bridge conduits have been used to study nerve regeneration in vivo [10-12].
The nerve bridge technique is the introduction of both ends of the injured nerve
stumps into a tubular chamber, which can aid guidance of growing nerve fibers along
appropriate paths and can enhance the precision of stump approximation. Several
conduit. In the nondegradable materials, silicone rubber is the most acceptable
material applied to construct the nerve guide, mainly for its stable properties. It has
been demonstrated that silicone rubber conduits (SRCs) are well tolerated in humans
even after 3 years of implantation [13]. Numerous biodegradable materials have been
used to construct nerve guides to repair injured nerves, such as poly-lactic acid and
poly-glycolic acid [14,15]. Recently, a biodegradable material composed of
genipin-crosslinked gelatin has also been used to make nerve regeneration chambers
[16]. Gelatin, which is essentially denatured collagen, has a myriad of applications in
the food, pharmaceutical, and cosmetic industries [17]. As for the genipin, it is a
naturally occurring and low-cytotoxic crossing agent, which can be obtained from its
parent compound geniposide isolated from the fruits of Gardenia jasminoides Ellis
[18].
Neurochemical measurements of specific proteins such as neurotypic and
glialtypic proteins have been used to detect injury in the developing nervous system in
vivo [19]. Although not widely applied to date, the use of neurochemical measures of
nerve regeneration has the potential to add to data obtained from the use of the
aforementioned morphological measures. A number of neurotypic and glialtypic
proteins have been associated with PC12 cell differentiation and Schwann cell
membrane-associated proteins, such as synapsin, and type β transforming growth
factors (TGF-βs) [20].
The present study examined the use of proteins associated with neurite
outgrowth, synapse formation, and Schwann cell proliferation and myelination as
surrogate measures of rat peripheral nerve regeneration in SRCs, genipin-crosslinked
gelatin conduits (GGCs), and porous GGCs (PGGCs). This work tested the hypothesis
that GAP-43, synapsin I, and TGF- β are sensitive to chemical disruption of
regenerated nerves in the three types of bridging conduits. The expression of GAP-43,
synapsin I, and TGF-β were determined at the same period of time and compared
with axonal growth in the regenerated nerves assessed using the morphological and
electrophysiological methods.
2. Materials and Methods
2.1. Manufacturing of PGGCs and GGCs
PGGCs and GGCs were constructed as described previously [21,22]. A
polypropylene tubing (4.3 mm ID) was used as a mold to control the exterior shape of
the PGGC. First, using a syringe, 50 ml of a 10% (w/w) solution of gelatin (300
bloom number, Sigma #G2500) in distilled water with a fine powder of gelatin was
blocked with a plug. A glass rod (1.6 mm OD), used as a mandrel, was vertically
pushed along the wall of the polypropylene tubing through the opening without the
plug and into the gelatin paste. The open end of the polypropylene tubing was then
sealed with another plug. Next, lyophilization was performed for 3 days and the
specimens were retrieved from the polypropylene tubing, immersed in 0.5% (w/w)
solution of genipin (Challenge Bioproducts Co., Taichung, Taiwan) for 48 h for
cross-linking. The PGGC was then manually de-molded from the glass mandrel. To
clean the residual genipin, the PGGCs were immersed in distilled water for 30 min,
washed with distilled water for 10 times, and freeze-dried again before storage. To
allow fixation of the nerve tissue to the conduit, two small holes were drilled at both
ends of the PGGCs. Finally, the PGGCs were sterilized with 25 kGy ofγ-ray for subsequent implantation. As for the GGCs, a 10% (w/w) solution of gelatin in
distilled water at 60°C was prepared by magnetic stirring. A silicone rubber tube (1.96
mm OD; Helix Medical, Inc., Carpinteria, CA) was used as a mandrel vertically
dipped into the gelatin solution. The mandrel was then withdrawn slowly and allowed
to stand for 30 min for air-drying. Eight coating steps were used to obtain a gelatin
tube. The gelatin-coated mandrels were then cross-linked in a 0.5% (w/w) solution of
genipin for 48 h. The GGCs were then de-molded, cleaned, air-dried, and sterilized
2.2. Cross-linking degree of PGGCs and GGCs
Ninhydrin assay was used to evaluate the cross-linking degree of PGGCs and
GGCs. Ninhydrin (2,2-dihydroxy-1,3-indanedione) was used to determine the amount
of amino groups of each test sample. The test PGGCs and GGCs were heated with a
ninhydrin solution for 20 min. After being heated with ninhydrin, the optical
absorbance of the solution was recorded using a spectrophotometer (Model
Genesys™ 10,SpectronicUnicam,New York,NY)at570 nm (wavelength ofthe blue-purple color) using gelatin at various known concentrations as standard. The
amount of free amino groups in the residual gelatin, after heating with ninhydrin, is
proportional to the optical absorbance of the solution. The cross-linking degree of
PGGCs and GGCs was then determined.
2.3. Bridging conduit implantation
A total of 36 adult Sprague-Dawley rats underwent placement of SRCs (1.47 mm
ID, 1.96 mm OD; Helix Medical, Inc., Carpinteria, CA), GGCs, and PGGCs (12 rats
per each tube group), which were removed upon sacrifice at 8 weeks. The animals
were anesthetized with an inhalational anesthetic technique (AErrane®, Baxter, USA).
dissection, and the right sciatic nerve was severed into proximal and distal segments.
The proximal stump was then secured with a single 9-0 nylon suture through the
epineurium and the outer wall of the nerve conduits. The distal stump was secured
similarly into the other end of the chamber. Both the proximal and distal stumps were
secured to a depth of 2.5 mm into the chamber, leaving a 10-mm gap between the
stumps. The muscle layer was re-approximated with 4-0 chromic gut sutures, and the
skin was closed with 2-0 silk sutures. All animals were housed in temperature (22°C)
and humidity (45%) controlled rooms with 12-hour light cycles, and they had access
to food and water ad libitum. All animals were maintained in facilities approved by
China Medical University for Accreditation of Laboratory Animal Care, and
according to the regulations and standards of the National Science Council of Health
of the Republic of China.
2.4. Electrophysiological techniques
After 8 weeks of regeneration, all the animals with apparent nerve regeneration
were re-anaesthetized and the sciatic nerve exposed. The stimulating cathode was a
stainless-steel monopolar needle, which was placed directly on the sciatic nerve trunk,
5-mm proximal to the transection site. The anode was another stainless-steel
conductive velocity (NCV) of the evoked muscle action potentials (MAP) were
recorded from gastrocnemius muscles with micro-needle electrodes linked to a
computer system (Biopac Systems, Inc., USA). The amplitude and the area under the
MAP curve from the baseline to the maximal negative peak were calculated. The
MAP was used to calculate the NCV, which was carried out by placing the recording
electrodes in the gastrocnemius muscles and stimulating the sciatic nerve proximally
and distally to the bridging conduit. The NCV was then calculated by dividing the
distance between the stimulating sites by the difference in latency time.
2.5. Histological measures
Immediately after the recording of muscle action potential, sciatic nerve sections
were extracted from the middle of the regenerated nerve in the chamber. The rest of
the nerve tissues were used for later neurochemical measurements. Following fixation,
the nerve tissue was post-fixed in 0.5% osmium tetroxide, dehydrated, and embedded
in Spurr’sresin.The tissue was then cut to a thickness of 5 µm using a microtome (Leica EM UC6, Leica Biosystems, Mount Waverley, Australia) with a dry glass knife,
stained with Toluidine Blue. All tissue samples were observed under a light
microscope (Olympus IX70, Olympus Optical Co., Ltd., Japan). An image analyzer
counted the blood vessels and calculated the cross-sectional area of each nerve section
at magnifications between 40x and 400x. At least 30 to 50% of the nerve section area
was randomly selected from each nerve specimen at a magnification of 400x to count
the axons. The axon counts were extrapolated by using the area algorithm to estimate
the total number of axons in each nerve. Axon density was then obtained by dividing
the axon counts by the total nerve areas. All data are expressed as mean ± standard
deviation. Statistical comparisons between groups were made using the one-way
ANOVA. A p value of < 0.05 was considered statistically significant.
2.6. Neurochemical measures
GAP-43, synapsin I, and TGF-β were quantified using electrophoresis and
Western blot analysis. Protein concentration of sciatic nerve extracts was determined
by the Bradford method (Bio-Rad Protein Assay, Hercules, CA). Protein samples (50 μg/lane) were separated on a 10% SDS polyacrylamide gel electrophoresis (SDS-PAGE) with a constant voltage of 75 V. Electrophoresed proteins were
transferred to polyvinylidene difluoride (PVDF) membrane (Millipore, Bedford, MA, 0.45 μm pore size) with a transfer apparatus (Bio-Rad Protein Assay, Hercules, CA). PVDF membranes were incubated in 5% milk in TBS buffer. Primary antibodies
including GAP-43 (Abcam, Cambridge, MA), synapsin I (Millipore, Bedford, MA),
and TGF-β(Millipore, Bedford, MA) and α-tubulin (Neo Markers, Fremont, CA) were diluted to 1:500 in antibody binding buffer overnight at 4°C. The immunoblots
antibody solution containing goat anti-mouse IgG-HRP, goat anti-rabbit IgG-HRP, or
donkey anti goat IgG-HRP for 1 hour and diluted 500-fold in TBS buffer. The
immunoblots were then washed in TBS buffer for 10 min three times. For repeated
blotting, nitrocellulose membranes were stripped with Restore Western blot stripping
buffer (Pierce Biotechnology, Rockford, IL) at room temperature for 30 min. The
immunoblotted proteins were visualized using an enhanced chemiluminescence ECL
Western blotting luminal reagent (Santa Cruz, CA) and quantified using a Fujifilm
LAS-3000 chemiluminescence detection system (Tokyo, Japan). All data are
expressed as mean ± standard deviation. Statistical comparisons of Western blot data
between groups were made using the one-way ANOVA. A p value of < 0.05 was
considered statistically significant.
3. Results
3.1. Macroscopic observation of bridging conduits
The SRCs had a semitransparent chamber lumen (Fig. 1a) whereas the GGCs
(Fig. 1b) and the PGGCs (Fig. 1c) were dark blue in appearance caused by the
reaction between genipin and amino acids or proteins. The GGCs were concentric and
round with rough outer surface and the inner lumen was smooth. By comparison, the
PGGCs featured an outer surface with pores of variable size homogeneously
traversing their wall and a partially fenestrated inner surface connected by an open
3.2. Physical characteristics of PGGCs and GGCs
The cross-linking index of PGGCs and GGCs, expressed as a percentage of free
amino groups lost during cross-linking, was 36±3%. It means that 1.0 wt.% genipin
was sufficient to cross-link about 36% of the amino groups.
3.3. Macroscopic observation of nerve regenerates
After 8 weeks of implantation, all the SRCs were intact with no swelling or
deformation. Brownish fibrous tissue encapsulation covered all over the chamber and
the parts of the nerve stumps in the chamber openings. However, the regenerated
nerve could be seen through the chamber lumen after trimming the fibrous tissue
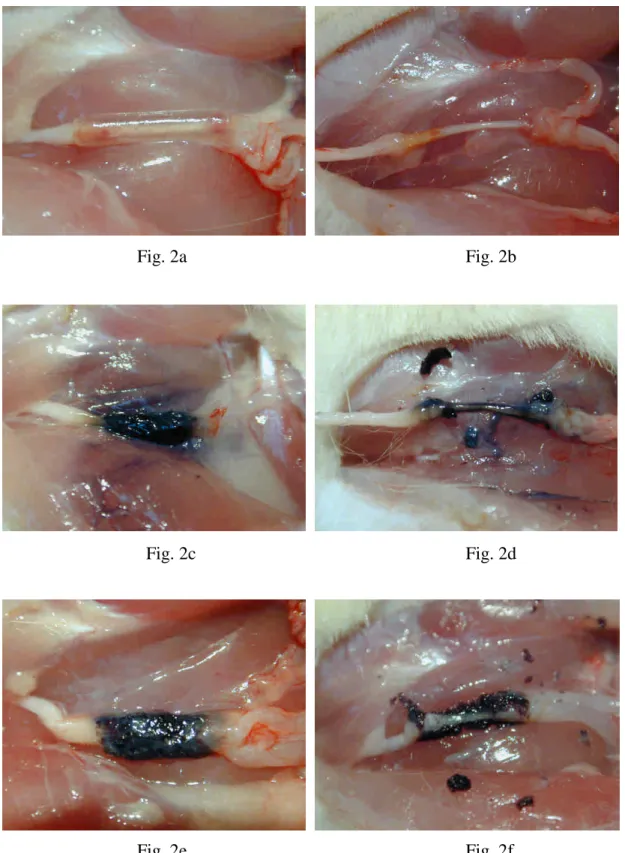
(Figs. 2a, 2b). No nerve dislocation was noted and the regenerated nerve, which was
surrounded by fluid, occupied a central location within the chamber. By comparison,
the GGCs featured a partially fenestrated outer layer, however, they still remained
circular with a round lumen (Figs. 2c, 2d). As for the PGGCs, their process of
swelling and degradation was dramatic (Figs. 2e, 2f). Only a small amount of wall
residues was seen surrounding the regenerating nerve.
3.4. Morphological evaluation of regenerated nerves
consisted mainly of a collagenous connective tissue matrix in which circumferential
cells resembling perineurial cells and fibroblasts were seen (Fig. 3a). The endourium
components included fibroblasts, an occasional macrophage, and collagenous and,
mainly, reticular fibers which surrounded Schwann cells ensheathing the axons. The
nerve fibers were packed in the nerve bundles with oval appearances. Nuclei of
Schwann cells were interspersed among these nerve fibers. Axons in the endoneurium
were easily defined by their surrounding myelin sheaths stained dark blue by the
toluidine blue (Fig. 3b). Blood vessels were numerous in the epineurium as well as in
the endoneurial areas of the nerve. By comparison, the regenerated nerves in the
GGCs displayed a structure with a thin epineurium surrounding by thick fibrous
tissues (Fig. 3c). The endoneurium was cellular and vascularized in which thinly
myelinated fibers grouped in regenerative clusters were evident. In addition, Schwann
cells organized in clusters surrounding groups of unmyelinated axons were present
(Fig. 3d). These axon-Schwann cluster formations, termed as regeneration units, are
common organization structures seen under nerve cuff bridging conditions. As seen in
the GGCs, it is still difficult to discriminate between the epineurium from the
surrounding thick fibrous tissues of the regenerates in the PGGCs (Fig. 3e). However,
the regenerated nerves in the PGGCs were relatively more mature compared to those
myelinated axons had been seen (Fig. 3f).
3.5. Morphometric and electrophysiological measurements of regenerated nerves
Morphometric studies revealed constant data in regenerated nerves in the three
types of bridging conduits for their mean values of myelinated axon number, axon
density, and endoneurial nerve area. Specifically, the differences of the axon number
(Fig. 4a) and the axon density (Fig. 4b) in the GGCs compared to the SRCs and the
PGGCs reached the significant level at p = 0.05. In addition, the nerve regenerates in
the GGCs had a significantly smaller endoneurial area compared to those in the SRCs
(Fig. 4c). With respect to electrophysiology, morphometric observations were not
correlated with electromyography in the regenerated nerves (Figs. 5a-5c). Only the
nerves in the PGGCs had a significantly larger NCV compared to those in the SRCs
and GGCs (p < 0.05).
3.6. Neurochemical measurements of regenerated nerves
In the Western blot analysis, the nerve regenerates in the SRCs (p < 0.001) and
the PGGCs (p < 0.001) had a significantly higher expression of GAP-43 compared to
those in the GGCs (Fig. 6a). In addition, the nerve regenerates in the PGGCs (p =
compared to those in the SRCs (Fig. 6b). On the contrary, the TGF-β expression was
significantly higher in the groups of PGGC (p < 0.001) and GGC (p < 0.001) as
compared to the SRC group (Fig. 6c).
4. Discussion
While designing polymer chambers used for nerve regeneration, investigators
must evaluate the effect of tube characteristics on the quality of regenerated nerves.
Nondegradable tubes can provide an isolated and stable environment for the nerves to
regenerate across the gap and toward the distal stump. However, they may also block
the nutrient and metabolic exchange between the lumen and the outside environment.
To improve this disadvantage, investigators have developed biodegradable conduits
which could maintain their structure integrity, permitting cell infiltration and
subsequent tissue ingrowth during the regenerative processes [23,24]. In the present
study, we used nondegradable SRCs and degradable GGCs and PGGCs to repair
dissected rat peripheral nerve. The dynamic nature of the developing nervous system
is so complex that it is not easy to assess nerve recovery in the animal just based on a
single criterion. Therefore, my group tried to use electrophysiological methods
accompanied with morphological observations to evaluate regenerated rat sciatic
electrophysiological measurements as seen in the present study, which may result
from serious gastrocnemius muscle atrophy even though the muscle fibers had been
reinnervated [25]. Gait analysis is another popular way to evaluate the nerve
regenerates. However, the missing toes caused by automutilation due to nerve injury
could result in inconsistent walking patterns [26]. We therefore developed the
neurochemical ways in this study to assist in the morphometric measurements.
In the neurochemical analyses, the expression of GAP-43, synapsin I, and
TGF-β was determined over the same time period of nerve regeneration in the bridging
conduits. GAP-43 is a marker for growth cones and elongating axon of developing
neurons [27]. High levels of GAP-43 expression are correlated with the beginning of
neurite outgrowth. As for the synapsins, they are abundant phosphoproteins essential
for regulating neurotransmitter release and increase in level with the formation of
mature synapses in developing cell cultures [28]. In the present study, the expression
of GAP-43 and synapsin I of nerve regenerates was significantly higher in the group
of SRCs as compared to the GGCs. This result is similar to the morphological
measurements of nerve regeneration in that the regenerates in the GGCs had a less
mature microstructure with a significantly smaller area containing fewer myelinated
axons. These results imply that though the SRCs were nondegradable, they still
toward the distal stump. On the contrary, the GGCs had fewer myelinated axons with
numerous degenerating fibers even after 8 weeks of regeneration, though these
bridging conduits were degradable. Interestingly, regenerates in PGGCs had more
myelinated axons as compared to those in GGCs. Accordingly, these results indicate
that how to select a suitable material used for nerve entubulization is a serious
challenge. We believe that the nondegradable material, such as the silicone rubber
used in this study, is the best candidate in constructing the bridging conduit for its
superior properties which can provide a continuous and stable growth environment for
the regenerating nerves. By comparison, a delay in temporal events of nerve
regeneration was noted in the GGCs compared to those in the SRCs. It is most likely
that the GGCs degraded at a rate slower than expected, resulting in their degraded
materials and the metabolites produced by the regenerating nerves were sealed up in
the tube lumens. This could trigger the cellular activity vigorously, causing deleterious
effects on the regenerating nerves. The controversial result was dramatically improved
by using the PGGCs, suggesting that the progressive increase in wall permeability of
bridging conduits could accelerate collapse of the tube walls, allowing evacuation for
the hazardous materials and entrance of more nutrients and cells that could promote
nerve regeneration.
can stimulate DNA synthesis in quiescent cells and increase their proliferation rate
[29]. In adult animals, Schwann cells can provide their basal lamina as a substratum
for the regenerating axons to adhere and grow [30]. In the present study, the
expression of TGF-βof nerve regenerates was significantly higher in the groups of
GGCs and PGGCs as compared to the SRCs. This result is not consistent with our
morphometric measurements since if the Schwann cells are beneficial to regenerating
axons, the expression of TGF-βof nerve regenerates in the SRCs should be higher
than that in the GGCs. This result inspired us to look deeper into the TGF-βfor its
role playing in the nerve regeneration. In the literature, we found that the TGF-βhad
versatile functions. In addition to the Schwann cell proliferation-promoting capability,
the TGF-βcan also activate monocytes, generating fibroblast growth-promoting
factors to induce fibrosis [31]. Based on these theories, we believe the high levels of
TGF-β in the PGGCs and GGCs were coming from the thick fibrous tissues at the
outer layers of the nerve regenerates. By comparison, the nerve regenerates in the
SRCs had no such mass with only a thin epineurium surrounding the inner
endoneurium.
5. Conclusions
axonal outgrowth and neuronal maturation have the potential to serve as surrogate
markers of peripheral nerve regeneration. In particular, changes in GAP-43 and
synapsin I expression correlated well with axon growth in bridging conduits made by
different types of biomaterials. For TGF-β, its versatile biochemical functions may
deviate histological observations from morphological measures of the regenerated
nerves.
Acknowledgements
The authors would like to thank China Medical University and Hospital
(CMU96-172; CMU97-139; DMR-97-055; DMR-98-054) and Taiwan Department of
Health Clinical Trial and Research Center of Excellence (Contract No.
DOH100-TD-B-111-004) for financially supporting this research.
References
[1] R. Langer, “Editorial: tissue engineering: perspectives, challenges, and future directions,”Tissue Eng. Part A, 13: 1-2, 2007.
[2] H. Hosseinkhani, M. Hosseinkhani, S. Hattori, R. Matsuoka and N. Kawaguchi,
“Micro and nano-scale in vitro 3D culture system for cardiac stem cells,”J.
[3] F. Tian, H. Hosseinkhani, M. Hosseinkhani, A. Khademhosseini, Y. Yokoyama, G.
G. Estrada and H. Kobayashi, “Quantitativeanalysisofcelladhesion on aligned micro- and nanofibers,”J. Biomed. Mater. Res., 84A: 291-299, 2008.
[4] A. Akkouch, Z. Zhang and M. Rouabhia, “novel collagen /hydroxyapatite
/poly(lactide-co-ε-caprolactone) biodegradable and bioactive 3D porous scaffold
for bone regeneration,”J. Biomed. Mater. Res., 96A: 693-704, 2011.
[5] S. Berski, J. van Bergeijk, D. Schwarzer, Y. Stark, C. Kasper, T. Scheper, C.
Grothe, R. Gerardy-Schahn, A. Kirschning and G. Dräger, “Synthesis and
biological evaluation of a polysialic acid-based hydrogel as enzymatically
degradable scaffold material for tissue engineering,”Biomacromolecules, 9:
2353-2359, 2008.
[6] V. T. Ribeiro-Resende, B. Koenig, S. Nichterwitz, S. Oberhoffner and B.
Schlosshauer, “Strategies for inducing the formation of bands of Büngner in
peripheral nerve regeneration,”Biomaterials, 30: 5251-5259, 2009.
[7] C. M. Valmikinathan, S. Defroda and X. Yu, “Polycaprolactone and bovine serum
albumin based nanofibers for controlled release of nerve growth factor,”
Biomacromolecules, 10: 1084-1089, 2009.
[8] J. D. Yuan, W. B. Nie, Q. Fu, X. F. Lian, T. S. Hou and Z. Q. Tan, “Novel
Schwann cells,”Chin. J. Traumatol., 12: 133-137, 2009.
[9] K. P. Das, T. M. Freudenrich and W. R. Mundy, “Assessment of PC12 cell
differentiation and neurite growth: a comparison of morphological and
neurochemical measures,”Neurotoxicol. Teratol., 26: 397-406, 2004.
[10] I. P. Clements, Y. T. Kim, A. W. English, X. Lu, A. Chung and R. V. Bellamkonda,
“Thin-film enhanced nerve guidance channels for peripheral nerve repair,”
Biomaterials, 30: 3834-3846, 2009.
[11] C. M. Lu, C. J. Chang, C. M. Tang, H. C. Lin, S.C. Hsieh, B. S. Liu and W. C.
Huang, “Effectsofasymmetric polycaprolactone discs on co-culture nerve conduit model,”J. Med. Biol. Eng., 29: 76-82, 2009.
[12] H. Jiao, J. Yao, Y. Yang, X. Chen, W. Lin, Y. Li, X. Gu and X. Wang,
“Chitosan/polyglycolic acid nerve grafts for axon regeneration from prolonged axotomized neurons to chronically denervated segments,”Biomaterials, 30:
5004-5018, 2009.
[13] G. Lundborg, B. Rosén, S. O. Abrahamson, L. Dahlin and N. Danielsen, “Tubular
repair of the median nerve in the human forearm. Preliminary findings,”J. Hand
Surg., 19: 273-276, 1994.
[14] J. H. Huang, D. K. Cullen, K. D. Browne, R. Groff, J. Zhang, B. J. Pfister, E. L.
engineered nervous tissue constructs promotes peripheral nerve regeneration,”
Tissue Eng. Part A, 15: 1677-1685, 2009.
[15] Y. Xiong, Y. S. Zeng, C. G. Zeng, B. L. Du, L. M. He, D. P. Quan, W. Zhang, J.
M. Wang, J. L. Wu, Y. Li and J. Li, “Synaptic transmission of neural stem cells
seeded in 3-dimensional PLGA scaffolds,”Biomaterials, 30: 3711-3722, 2009.
[16] Y. S. Chen, J. Y. Chang, C. Y. Cheng, F. J. Tsai, C. H. Yao and B. S. Liu, “An in
vivo evaluation of a biodegradable genipin-cross-linked gelatin peripheral nerve
guide conduit material,”Biomaterials, 26: 3911-3918, 2005.
[17] Y. S. Chen, S. M. Kuo and C. H. Yao, “A review for gelatin used for artificial
nerve and bone implants- 10-year retrospection,”Biomed. Eng. Appl. Basis
Comm., 21: 233-238, 2009.
[18] M. C. Chen, C. T. Liu, H. W. Tsai, W. Y. Lai, Y. Chang and H. W. Sung,
“Mechanical properties, drug eluting characteristics and in vivo performance of a genipin-crosslinked chitosan polymeric stent,” Biomaterials, 30: 5560-5571,
2009.
[19] H. A. El-Fawal and J. P. O'Callaghan, “Autoantibodies to neurotypic and
gliotypic proteins as biomarkers of neurotoxicity: assessment of trimethyltin
(TMT),”Neurotoxicology, 29: 109-115, 2008.
Nagy, “Bcl-2 or bcl-XL gene therapy increases neural plasticity proteins nestin
and c-fos expression in PC12 cells,”Neurochem. Int., 55: 349-353, 2009.
[21] M. C. Lu, S. W. Hsiang, T. Y. Lai, C. H. Yao, L. Y. Lin and Y. S. Chen,
“Influence of cross-linking degree of a biodegradable genipin-cross-linked gelatin guide on peripheral nerve regeneration,”J. Biomater. Sci. Polym. Ed., 18: 843-863,
2007.
[22] J. Y. Chang, T. Y. Ho, H. C. Lee, Y. L. Lai, M. C. Lu, C. H. Yao and Y. S. Chen,
“Highly permeable genipin-cross-linked gelatin conduits enhance peripheral nerve regeneration,”Artif. Organs, 33: 1075-1085, 2009.
[23] Y. Z. Bian, Y. Wang, G. Aibaidoula, G. Q. Chen and Q. Wu, “Evaluation of
poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) conduits for peripheral nerve
regeneration,”Biomaterials, 30: 217-225, 2009.
[24] P. Plikk, S. Målberg and A. C, ”Albertsson. Design of resorbable porous tubular
copolyester scaffolds for use in nerve regeneration,”Biomacromolecules, 10:
1259-1264, 2009.
[25] H. T. Chen, C. H. Yao, P. D. Chao, Y. C. Hou, H. M. Chiang, C. C. Hsieh, C. J.
Ke and Y. S. Chen, “Effect of serum metabolites of Pueraria lobata in rats on
peripheral nerve regeneration: in vitro and in vivo studies,”J. Biomed. Mater. Res.
[26] M. F. Meek, J. Ijkema-Paassen and A, ”Gramsbergen. Functional recovery after transection of the sciatic nerve at an early age: a pilot study in rats,”Dev. Med.
Child Neurol., 49:377-379, 2007.
[27] J. J. Xu, E. Y. Chen, C. L. Lu and C. He, “Recombinant ciliary neurotrophic
factor promotes nerve regeneration and induces gene expression in silicon
tube-bridged transected sciatic nerves in adult rats,”J. Clin. Neurosci., 16:
812-817, 2009.
[28] W. L. Coleman and M. Bykhovskaia, “Synapsin I accelerates the kinetics of
neurotransmitter release in mouse motor terminals,”Synapse, 63: 531-533, 2009.
[29] E. O. Johnson, A. Charchanti and P. N. Soucacos, “Nerve repair: experimental
and clinical evaluation of neurotrophic factors in peripheral nerve regeneration,”
Injury, 39 Suppl. 3: S37-S42, 2008.
[30] A. Pierucci, E. A. Duek and A. L. de Oliveira, “Expression of basal lamina
components by Schwann cells cultured on poly(lactic acid) (PLLA) and
poly(caprolactone) (PCL) membranes,”J. Mater. Sci. Mater. Med., 20: 489-495,
2009.
[31] K. R. Cutroneo, “TGF-beta-induced fibrosis and SMAD signaling: oligo decoys
as natural therapeutics for inhibition of tissue fibrosis and scarring,”Wound
Captions
Fig. 1. Macrographs of (a) SRC, (b) GGC, and (c) PGGC.
Fig. 2. (a) SRC, (c) GGC, and (e) PGGC 8 weeks of implantation. Regenerated
nerve exposed after trimming the remained bridging materials of (b) SRC, (d)
GGC, and (f) PGGC.
Fig. 3. Photomicrographs of regenerated sciatic nerve sections 8 weeks after
surgery stained with Toluidine Blue. (a) Regenerated nerve repaired with
SRC had a well-defined structure in which the endoneurium (circle inside)
was highly cellular and vascularized. In the epineurium, circumferential cells
resembling perineurial cells (asterisks in insert and (a)) and fibroblasts
(small arrow in insert and (a)) were seen. By comparison, regenerated nerve
repaired with (c) GGC formed thick fibrous tissues and a thin epineurium
(regions between arrows in insert and (c)) surrounding the inner
endoneurium (circle inside). (e) Similarly, thick fibrous tissues of the
regenerates were seen in the PGGCs. Abundant myelinated axons were seen
in the regenerated nerves repaired with (b) SRC and (f) PGGC. (d) In the
GGC, thinly myelinated fibers grouped in regenerative clusters (arrows)
were evident accompanied by numerous degenerating fibers In addition,
axons were present (arrowheads in insert and (d)). Scale bars in a, c and e =
100 µm; b, d and f = 30 µm; insert in (a), (d) = 20 µm, (c) = 30 µm.
Fig. 4. Morphometric analysis from regenerated nerves in bridging conduits,
including (a) myelinated axon count, (b) axon density, and (c) endoneurial
area. Data are means ± S.E. (n=12). *Significantly different at p<0.05.
Fig. 5: Analysis of the evoked muscle action potentials, including (a) peak
amplitude, (b) MAP area, and (c) NCV. *Significantly different at p<0.05.
Fig. 6. Representative immunoblots for (a) GAP-43, (b) synapsin I, and (c) TGF-β,
and quantification of the three protein levels in regenerated nerves.
Fig. 1a
Fig. 1b
Fig. 2a Fig. 2b
Fig. 2c Fig. 2d
Fig. 3a
Fig. 3b
Fig. 3c
Fig. 3e
Fig. 4a Fig. 4b * * * *
Fig. 4c *
Fig. 5a
Fig. 5c
* *
SRC PGGC GGC Fig. 6a ← 43 KDa ← 42 KDa β-actin GAP43 * *
SRC PGGC GGC Fig. 6b ← 42KDa ← 70KDa synapsin I β-actin * *
SRC PGGC GGC Fig. 6c ← 25KDa ← 42KDa TGF-β1 β-actin * *