Elsevier Editorial System(tm) for Biomaterials Manuscript Draft
Manuscript Number: jbmt14835R1
Title: Mechanisms of cellular uptake and intracellular trafficking with chitosan/DNA/poly(γ-glutamic acid) complexes as a gene delivery vector
Article Type: FLA Original Research
Section/Category: Biomaterials & Gene Transfer
Keywords: lysosome; macropinocytosis; caveolae-mediated pathway: transfection efficiency; cellular uptake
Corresponding Author: Dr. Hsing-Wen Sung,
Corresponding Author's Institution: National Tsing Hua University First Author: Shu-Fen Peng
Order of Authors: Shu-Fen Peng; Michael T Tseng; Yi-Cheng Ho; Ming-Cheng Wei; Zi-Xian Liao; Hsing-Wen Sung
Abstract: Chitosan (CS)-based complexes have been considered as a vector for DNA delivery; nonetheless, their transfection efficiency is relatively low. An approach by incorporating poly(γ-glutamic acid) (γ-PGA) in CS/DNA complexes was developed in our previous study to enhance their gene expression level; however, the detailed mechanisms remain to be understood. The study was designed to investigate the mechanisms in cellular uptake and intracellular trafficking of CS/DNA/γ-PGA complexes. The results of our molecular dynamic simulations suggest that after forming complexes with CS, γ-PGA displays a free γ-glutamic acid in its N-terminal end and thus may be recognized by γ-glutamyl transpeptidase in the cell membrane, resulting in a significant increase in their cellular uptake. In the endocytosis inhibition study, we found that the internalization of CS/DNA complexes took place via macropinocytosis and caveolae-mediated pathway; by incorporating γ-PGA in complexes, both uptake pathways were further enhanced but the caveolae-mediated pathway played a major role. TEM was used to gain directly understanding of the internalization mechanism of test complexes and confirmed our findings obtained in the inhibition experiments. After
internalization, a less percentage of co-localization of CS/DNA/γ-PGA complexes with lysosomes was observed when compared with their CS/DNA counterparts. A greater cellular uptake together with a less entry into lysosomes might thus explain the promotion of transfection efficiency of CS/DNA/γ-PGA complexes. Knowledge of these mechanisms involving CS-based complexes containing γ-PGA is critical for the development of an efficient vector for DNA transfection.
AUTHOR DECLARATION
We the undersigned declare that this manuscript is original, has not been published before and is not currently being considered for publication elsewhere.
We wish to draw the attention of the Editor to the following facts which may be considered as potential conflicts of interest and to significant financial contributions to this work. [OR] We wish to confirm that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
We confirm that the manuscript has been read and approved by all named authors and that there are no other persons who satisfied the criteria for authorship but are not listed. We further confirm that the order of authors listed in the manuscript has been approved by all of us.
We confirm that we have given due consideration to the protection of intellectual property associated with this work and that there are no impediments to publication, including the timing of publication, with respect to intellectual property. In so doing we confirm that we have followed the regulations of our institutions concerning intellectual property.
We understand that the Corresponding Author is the sole contact for the Editorial process (including Editorial Manager and direct communications with the office). He/she is responsible for communicating with the other authors about progress, submissions of revisions and final approval of proofs. We confirm that we have provided a current, correct email address which is accessible by the Corresponding Author and which has been configured to accept email from [email protected].
[LIST AUTHORS AND DATED SIGNATURES ALONGSIDE]
Shu-Fen Peng Michael T. Tseng Yi-Cheng Ho Ming-Cheng Wei Zi-Xian Liao Hsing-Wen Sung
August 26, 2010
Professor D.F. Williams, F.R. Eng. Editor-in-Chief
Biomaterials
RE: jbmt14835 Revision
Dear Professor Williams:
Thank you very much for arranging the review process for our manuscript, jbmt14835. As suggested, the title of this manuscript has been re-organized as “Mechanisms of cellular uptake and intracellular trafficking with chitosan/DNA/poly( -glutamic acid) complexes as a gene delivery vector”. Thanks again.
Sincerely yours,
Hsing-Wen Sung, Ph.D. Deputy Dean, R&D Office Tsing Hua Chair Professor
Department of Chemical Engineering National Tsing Hua University Hsinchu, Taiwan 30013 Phone: +886-3-574-2540 Fax: +886-3-572-6832
E-mail: [email protected]
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Mechanisms of cellular uptake and intracellular trafficking with chitosan/DNA/poly(-glutamic acid) complexes as a gene delivery vector
Shu-Fen Peng1, Michael T. Tseng2, Yi-Cheng Ho3, Ming-Cheng Wei1, Zi-Xian Liao1, Hsing-Wen Sung1*
1
Department of Chemical Engineering, National Tsing Hua University, Hsinchu, Taiwan, R.O.C.
2
Department of Anatomical Sciences and Neurobiology, University of Louisville, Louisville, Kentucky, U.S.A.
3
Department of Biotechnology, Vanung University, Chungli, Taoyuan, Taiwan, R.O.C.
*
Correspondence to:
Hsing-Wen Sung, PhD Professor
Department of Chemical Engineering National Tsing Hua University Hsinchu, Taiwan 30013 Tel: +886-3-574-2504 Fax: +886-3-572-6832
E-mail: [email protected] *Title Page
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 Abstract
Chitosan (CS)-based complexes have been considered as a vector for DNA delivery; nonetheless, their transfection efficiency is relatively low. An approach by incorporating poly(γ-glutamic acid) (γ-PGA) in CS/DNA complexes was developed in our previous study to enhance their gene expression level; however, the detailed mechanisms remain to be understood. The study was designed to investigate the mechanisms in cellular uptake and intracellular trafficking of CS/DNA/γ-PGA complexes. The results of our molecular dynamic simulations suggest that after forming complexes with CS, γ-PGA displays a free γ-glutamic acid in its N-terminal end and thus may be recognized by γ-glutamyl transpeptidase in the cell membrane, resulting in a significant increase in their cellular uptake. In the endocytosis inhibition study, we found that the internalization of CS/DNA complexes took place via macropinocytosis and caveolae-mediated pathway; by incorporating γ-PGA in complexes, both uptake pathways were further enhanced but the caveolae-mediated pathway played a major role. TEM was used to gain directly understanding of the internalization mechanism of test complexes and confirmed our findings obtained in the inhibition experiments. After internalization, a less percentage of co-localization of CS/DNA/γ-PGA complexes with lysosomes was observed when compared with their CS/DNA counterparts. A greater cellular uptake together with a less entry into lysosomes might thus explain the promotion of transfection efficiency of CS/DNA/γ-PGA complexes. Knowledge of these mechanisms involving CS-based complexes containing γ-PGA is critical for the development of an efficient vector for DNA transfection.
Keywords: lysosome; macropinocytosis; caveolae-mediated pathway: transfection
efficiency; cellular uptake *Abstract
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 1. Introduction
Chitosan (CS), a cationic polysaccharide, is biodegradable, non-toxic and tissue compatible [1,2]. It has the potential to condense anionic DNA into a compact structure (CS/DNA complexes) through electrostatic interactions and has been considered as a non-viral vector for gene delivery [3,4]. Although advantageous for DNA packing and protection, CS-based complexes may lead to difficulties in DNA release once arriving at the site of action intracellularly, thus limiting their transfection efficiency. To overcome this problem, an approach that can modify the internal structure of CS/DNA complexes by incorporating a negatively charged poly(γ-glutamic acid) (γ-PGA) was developed in our previous study [5].
Analysis of the internal structure of CS/DNA/γ-PGA complexes by small angle X-ray scattering (SAXS) revealed that CS formed complexes with DNA and γ-PGA separately and yielded two types of domains, leading to the formation of “compounded nanoparticles” [5]. With this unique internal structure, the compounded nanoparticles might disintegrate into a number of even smaller subparticles after cellular internalization, thus improving the dissociation capacity of CS and DNA and enhancing the efficacy of gene expression [5]. In addition to improving the release of DNA intracellularly, the incorporation of γ-PGA in CS/DNA complexes markedly increased their cellular uptake. Similar observations were also reported by Kurosaki et al. [6], using cationic complexes coated with γ-PGA. However, the detailed mechanisms in endocytosis and intracellular routing of complexes incorporating with γ-PGA remain to be understood. Understanding the role of test complexes on their cellular uptake and intracellular fate is essential for the rational design of non-viral delivery devices.
The study was therefore designed to investigate the potential internalization mechanism of CS/DNA complexes with or without the incorporation of γ-PGA, using transmission electron microscopy (TEM) and the inhibitors specific to various endocytotic pathways. The role that γ-PGA may play in the cellular uptake of test complexes was modeling by molecular dynamic (MD) simulations. The intracellular routing of test complexes was *Manuscript
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
observed by a confocal laser scanning microscope (CLSM). Additionally, test complexes were characterized using dynamic light scattering (DLS), and their efficacy in gene expression was determined by luminance spectrometry and flow cytometry.
2. Materials and Methods 2.1. Plasmid DNA
The plasmid DNAs used in the study were pEGFP-N2 (4.7 kb, coding an enhanced green fluorescence protein reporter gene, Clontech, Palo Alto, CA, USA) and pGL4.13 (4.6 kb, coding a firefly luciferase reporter gene, Promega, Madison, WI, USA). pEGFP-N2 and pGL4.13 were amplified using DH5α and purified by Qiagen Plasmid Mega Kit (Valencia, CA, USA) according to the manufacturer’s instructions. The purity of plasmids was analyzed by gel electrophoresis (0.8% agarose), while their concentration was measured by UV absorption at 260 nm (Jasco, Tokyo, Japan).
2.2. Preparation of test complexes
The charge ratio (N/P/C) of test complexes was expressed as the ratio of moles of the amino groups (N) on CS to the phosphate groups (P) on DNA and the carboxyl groups (C) on γ-PGA. Test complexes at N/P/C molar ratios of 10/1/0 (CS/DNA complexes) and 10/1/4 (CS/DNA/γ-PGA complexes) were prepared by an ionic-gelation method [7]. Briefly, an aqueous DNA (pEGFP-N2 or pGL4.13, 33 μg) was mixed with an aqueous γ-PGA (20 kDa, 0 or 51.2 μg, Vedan, Taichung, Taiwan) with a final volume of 100 μl. Test complexes were obtained upon addition of the mixed solution, using a pipette, into an aqueous CS (15 kDa, with a degree of deacetylation of 85%, 0.2 μg/μl, 100 μl, pH 6.0, Challenge Bioproducts, Taichung, Taiwan) and then thoroughly mixed for 30–60 s by vortex and left for at least 1 h at room temperature. The morphology of the obtained complexes was examined by TEM (JEOL, Tokyo, Japan) [8].
2.3. In vitro transfection
HT1080 (human fibrosarcoma) cells were cultured in DMEM media supplemented with 2.2 g/l sodium bicarbonate and 10% fetal bovine serum (FBS). Cells were subcultured
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
according to ATCC recommendations without using any antibiotics. For transfection, cells were seeded on 12-well plates at 2 × 105 cells/well and transfected the next day at 50–80% confluency. Prior to transfection, the media were removed and cells were rinsed twice with transfection media (DMEM without FBS, pH 6.0). Cells were replenished with 0.6 ml transfection media containing test complexes at a concentration of 2 μg DNA/well.
At 2 h post transfection, the transfection media containing test complexes were removed, the cells rinsed twice with transfection media and refilled with FBS-containing media until analysis at 48 h after transfection. Cells were then observed under a fluorescence microscope (Carl Zeiss Optical, Chester, VA, USA) to monitor any morphological changes and to obtain an estimate of the transfection efficiency. Cells transfected with LipofectamineTM 2000 (Invitrogen, Carlsbad, CA, USA) were used as a positive control and those without any treatment were used as a negative control. Transfection efficiencies were presented by two numeric indicators: percentage of cells transfected and gene expression level [9].
2.4. Percentage of cells transfected
The percentage of cells transfected was quantitatively assessed at 48 h after transfection by flow cytometry. Cells were detached by 0.025% trypsin-EDTA. Cell suspensions were then transferred to microtubes, fixed by 4% paraformaldehyde and determined the transfection efficiency by a flow cytometer (Beckman Coulter, Fullerton, CA, USA) equipped with a 488-nm argon laser for excitation. For each sample, 10,000 events were collected and fluorescence was detected. Signals were amplified in logarithmic mode for fluorescence to determine the EGFP positive events by a standard gating technique. The percentage of positive events was calculated as the events within the gate divided by the total number of events, excluding cell debris.
2.5. Gene expression level
The gene expression levels of cells were assayed by quantifying the expressions of EGFP or luciferase. The expression level of EGFP was quantified by comparing mean fluorescence of 2 × 105 cells. Briefly, cells were treated with test complexes containing
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
pEGFP-N2. After 48 h, cells were detached and analyzed by flow cytometry as described in Section 2.4.
For the expression of luciferase, cells were plated on 24-well plates (with a seeding density of 1 × 105 cells) and transfected as described in Section 2.3 with the exception that 1 μg pGL4.13 was used. The cells transfected were lysed by 100 μl of passive lysis buffer (Promega). The cell lysate was transferred into a 1.5-ml microtube, while the cell debris was separated by centrifugation (14,000 rpm, 5 min). Subsequently, a 100 μl of the luciferase assay reagent (Promega) was added to a 20 μl of the supernatant and the relative luminescence of the sample was determined by a microplate luminometer (Berthold Technologies,Bad Wildbad, Germany) and normalized to the total cell protein concentration by the Bradford method.
2.6. Fluorescent complex preparation and flow-cytometry analysis
Cy3-labeled CS (Cy3-CS) and FITC-labeled CS (FITC-CS) were synthesized as per the methods described in the literature [10,11]. To remove the unconjugated Cy3 and FITC, the synthesized Cy3-CS and FITC-CS were dialyzed in the dark against deionized (DI) water and replaced on a daily basis until no fluorescence was detected in the supernatant. The resultant Cy3-CS and FITC-CS were lyophilized in a freeze dryer. Cy3- and FITC-labeled complexes were then prepared as described in Section 2.2.
To quantify the cellular uptake of test complexes, cells were plated on 12-well plates and transfected with FITC-labeled complexes at a concentration of 2 μg DNA/well for 2
h. After transfection, cells were detached by 0.025% trypsin-EDTA and transferred to
microtubes. Subsequently, cells were resuspended in phosphate buffered saline (PBS)
containing 1mM EDTA and fixed in 4% paraformaldehyde. Finally, the cells were analyzed
by flow cytometry as described in Section 2.4.
2.7. MD simulations
MD simulations of the self-assembly of CS and γ-PGA in complexation were performed by a MD method [12]. MD simulations were accomplished with the program NAMD [13] using parameters adapted from the CHARMM 27 force field [14]. The
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
models were minimized to remove unfavorable contacts, brought to 310 K by velocity rescaling and equilibrated for 1 ns. Before any MD trajectory was run, 40 ps of energy minimization were performed to relax the conformational and structural tensions. This minimum structure was the starting point for the MD simulations. For this purpose, the molecule was embedded into a cubic simulation box of 80 Å. A cutoff distance of 12 Å was employed for the nonbonded and electrostatic interactions. The heating process was performed from 0 to 310 K through Langevin damping with a coefficient of 10 ps-1. A time step of 2 fs was employed for rescaling the temperature. After 20 ps heating to 310 K, equilibration trajectories of 1 ns were recorded, which provided the data for the structural and thermodynamic evaluations. The equations of motion were integrated with the Shake algorithm with a time step of 1 fs. Figures displaying atomistic pictures of molecules with hydrogen bondings were generated using UCSF Chimera [15].
2.8. Endocytosis inhibition
To study the effect of various inhibitors on the uptake of test complexes, cells were pre-incubated with the following inhibitors individually at concentrations which were not toxic to the cells: 10 μg/ml of chlorpromazine [16], 50nM wortamannin [17], 5 μg/ml cytochalasin D [18], 5 μg/ml filipin [19] or 200μM genistein [19,20]; the MTT assay [16] was employed to confirm their toxicity. In the study, the group without any treatment was used as a background in the flow cytometry analysis, while the groups in the presence of test complexes but without inhibitor treatment were used as controls and their fluorescence intensities were expressed as 100%. Following pre-incubation for 30 min, the inhibitor solutions were removed, and the freshly prepared test complexes (FITC-labeled) in media containing inhibitors at the same concentrations were added and further incubated for 2 h. Subsequently, cells were washed three times with PBS, collected according to the methods described above and analyzed by flow cytometry.
2.9. Examination of internalization of test complexes by TEM
To directly observe the mechanism of cellular internalization, cells were incubated with test complexes at 37°C. After washing three times with PBS, cells were fixed for 30 min at
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
room temperature in 2% paraformaldehyde and 2.5% glutaraldehyde in 0.1M cacodylate buffer at pH 7.4. The cells were rinsed twice in the same buffer with 6.8% sucrose and subsequently postfixed in 1% OsO4. After rinsing followed by dehydration in graded
alcohol series, the cells were embedded in Spurr resin and polymerized at 70°C overnight. Ultrathin sections were then cut with a diamond knife and loaded onto TEM grids. The sections were examined by a Philips CM10 electron microscope at accelerating voltage of 120 kV and micrographs were taken [21].
2.10. Intracellular trafficking
To study the intracellular trafficking, cells were treated with the Cy3-labeled test complexes in the serum-free medium. After incubation for 1.5 h, cells were washed twice with the pre-warmed PBS and then treated with 50nM Lysotracker (HCK-123, Invitrogene, Carlsbad, CA, USA) for 30 min at 37°C following the supplier’s protocol and examined using CLSM (TCS SL, Leica, Germany).
2.11. Statistical analysis
Comparison between groups was analyzed by the one-tailed Student’s t-test (SPSS, Chicago, Ill, USA). All data are presented as a mean value with its standard deviation indicated (mean ± SD). Differences were considered to be statistically significant when the
P values were less than 0.05.
3. Results and Discussion
CS/DNA complexes (a binary system) generally transfect cells more efficiently than naked DNA but less than commercially available liposome formulations [22]. It has been suggested that the strength of electrostatic interactions between CS and DNA prevent their dissociation within cells, thus precluding transcription of DNA and resulting in low transfection [1,2]. In a previous study, we demonstrated that after the incorporation of γ-PGA in CS/DNA complexes (a ternary system), the percentage of cells transfected and their gene level expressed were significantly enhanced [5]. Additionally, Kurosaki et al. reported that cationic complexes coated with the negatively charged γ-PGA not only reduced
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
their cytotoxicity but also significantly improved their efficiency in gene expression [6]. Generally, anionic complexes are not taken up well by cells because of the electrostatic repulsion induced by the negatively charged cell membranes. Therefore, the ternary complexes containing γ-PGA must have different mechanisms in cellular uptake and intracellular fate from their binary counterparts.
3.1. Characterization of test complexes
In the study, test complexes were prepared with an N/P/C ratio of 10/1/0 (binary CS/DNA complexes) or 10/1/4 (ternary CS/DNA/γ-PGA complexes). The binding capacity of CS with DNA prepared at various N/P ratios was evaluated in our previous study using the gel retardation assay [5]. The results showed that as the N/P ratio was increased to 10/1, the migration of DNA was retarded completely. By incorporating γ-PGA in CS/DNA complexes, no significant DNA release was observed. However, as the amount of γ-PGA incorporated was increased to a critical value (N/P/C ratio of 10/1/6), the transfection efficiency of CS/DNA/γ-PGA complexes started to drop appreciably [5].
TEM was used to examine the morphology of test complexes. As shown in Fig 1, the binary CS/DNA complexes had an irregular shape, while the ternary CS/DNA/γ-PGA complexes were spherical in shape. Our previous SAXS results indicated that test complexes formed by the ternary system were composed of two types of domains containing CS/DNA and CS/γ-PGA complexes [5], as schematically illustrated in Fig 1. The particle size and zeta potential of CS/DNA (CS/DNA/γ-PGA) complexes measured by DLS were 140.2 ± 7.7 nm (152.5 ± 5.1 nm) and 31.7 ± 0.8 mV (28.7 ± 1.2 mV), respectively (n = 5 batches). The encapsulation efficiencies of DNA in test complexes for both studied groups were about the same and approached 100%.
3.2. Cellular uptake and transfection efficiency
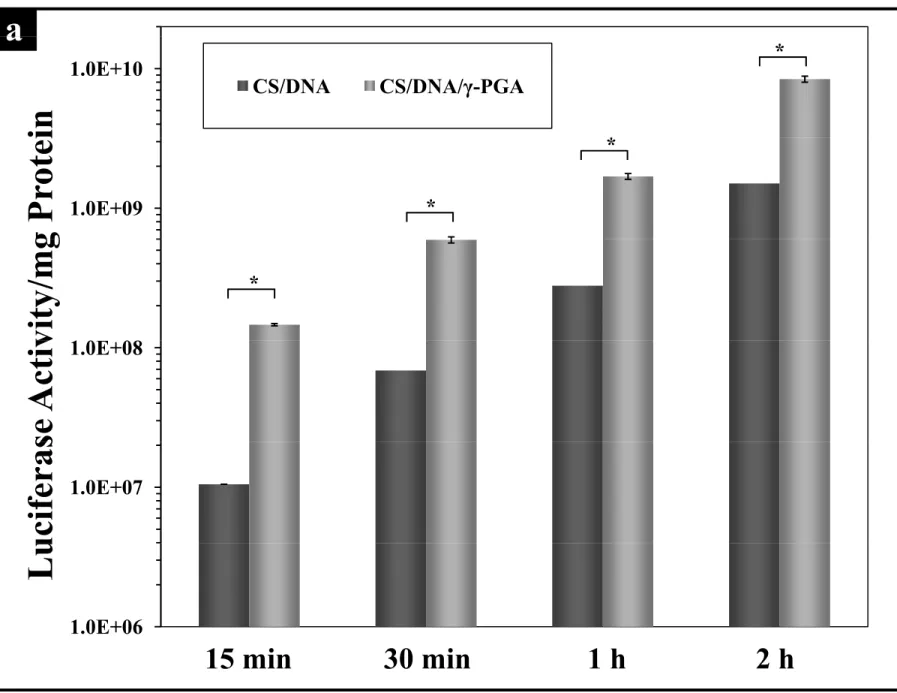
After pre-incubation of the cells with test complexes for different time periods (i.e., distinct durations of internalization or transfection), their successful expression of delivered DNA at 48 h post transfection was reflected by luciferase gene expression levels. As shown in Fig. 2a, cells transfected with CS/DNA or CS/DNA/γ-PGA complexes produced a
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
gradient increase of luciferase expression as the internalization time progressed. The luciferase gene expression levels for cells incubated with CS/DNA/γ-PGA complexes were consistently higher than their CS/DNA counterparts throughout the entire time course of the study (P < 0.05).
To visualize the cellular uptake, CS was fluorescently labeled with FITC. The percentage of cells that internalized the FITC-labeled test complexes (Fig 2b) and their fluorescence intensity (Fig. 2c) were quantified by flow cytometry at 2 h after transfection. Compared to those transfected with CS/DNA complexes, the percentage of fluorescent cells and their fluorescence intensity in the group treated with CS/DNA/γ-PGA complexes were significantly enhanced (P < 0.05), an indication of a greater cellular uptake.
To determine the percentage of cells that actually expressed the transgene, we counted the number of EGFP-positive cells using flow cytometry at 48 h post transfection. As shown in Fig. 2d, 15% of the cells produced EGFP when transfected with CS/DNA complexes. By incorporating γ-PGA in complexes (CS/DNA/γ-PGA), an approximately 4-fold increase in the percentage of EGFP-positive cells was found (55%, P < 0.05).
3.3. MD simulations
MD simulations were performed in a full-atom model to gain insight into the role that γ-PGA may play in assisting the cellular uptake. Both CS and γ-PGA molecules considered in the MD simulations contained 10 monomer units. The present atomistic simulation would allow the capture of the self-assembly of CS and γ-PGA in complexation. The methodology adopted has recently been applied to enable simulations of the self-assembly of protein and detergent into mixed micelles [23,24]. As shown in Fig. 3a, free γ-PGA forms an intramolecular hydrogen bonding between the amine group (−NH2) of
the N-terminal glutamyl unit and the carbonyl group (−C=O) on its neighboring unit; thus, the terminal amine group on γ-PGA is hidden. In contrast, after forming complexes, the amine groups on CS may form intermolecular hydrogen bondings with the carbonyl groups of γ-PGA; therefore, the hidden amine group in the N-terminal γ-glutamly unit on γ-PGA is exposed (Fig. 3b).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
The exposure of the free N-terminal γ-glutamly unit of γ-PGA on the surface of CS/γ-PGA complexes (Fig. 3b) may enhance its interaction with the enzyme γ-glutamyl transpeptidase (GGT), which is localized in the cell membrane and exerts the only specificity to those substrates containing free γ-glutamic acids in their N-terminal ends [25]. It has been reported that GGT can use a wide variety of γ-glutamyl compounds as substrates [26]. The most popular case of GGT substrates is glutathione; GGT may cleave the γ-glutamyl bond of extracellular glutathione, enabling the cell to use extracellular glutathione as a source of cysteine to increase the synthesis of intracellular glutathione [27]. In CS/γ-PGA complexes, γ-PGA displays a free γ-glutamic acid in its N-terminal end and thus may be recognized by GGT in the cell membrane, resulting in a significant increase in their cellular uptake (Fig. 2b and 2c). The detailed mechanism may need further investigation.
The results discussed in Sections 3.2 and 3.3 suggest that with the incorporation of γ-PGA, the cellular uptake and transgene expression of CS/DNA complexes were significantly enhanced. In a previous study, we demonstrated that there are different pathways involved in the internalization of CS/DNA and CS/DNA/γ-PGA complexes; the former is trypsin concentration-independent and the latter trypsin concentration-dependent [5]. However, their exact mechanisms in endocytosis and intracellular trafficking remain to be understood. Understanding the fate of complexes with respect to their uptake pattern and the intracellular localization is crucial in designing a new genetic or drug carrier [28].
3.4. Endocytosis pathways of CS/DNA and CS/DNA/γ-PGA complexes
A variety of forms of endocytosis have been demonstrated to be involved in the cellular uptake of polyplexes [16,20]. Current evidences suggest that endocytosis is the main mode of CS-based complexes entering into the cells [29]. To elucidate their potential cellular uptake pathways, the interactions between CS/DNA or CS/DNA/γ-PGA complexes and cell membranes were investigated by treating cells with different chemical inhibitors of clathrin-mediated endocytosis, caveolae-mediated endocytosis and macropinocytosis and then analyzed by flow cytometry. It has been reported that a narrow concentration range of specific inhibitory function and nonspecific toxicity exists [16]. We therefore first
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
determined the cytotoxicity of the concentration of each inhibitor used in the subsequent transfection experiments. As shown in Fig. 4, the concentration of each inhibitor used in the study did not significantly reduce the cell viability (> 94% of control). These data ensure that the reductions in cellular uptake to be discussed in Fig. 5a and 5b are specific to the inhibitors, rather than the cytotoxicity of inhibitors.
Fig. 5a and 5b presents the results of the internalization of CS/DNA and CS/DNA/γ-PGA complexes by cells in the presence of various inhibitors; their counterparts in the absence of inhibitors were used as controls. The inhibition of clathrin-mediated uptake was tested by using the cationic amphiphilic drug chlorpromazine, which causes clathrin to accumulate in late endosomes, thereby inhibiting coated pit endocytosis [20,30]. Compared with the controls, the cellular uptake of both test complexes increased relatively in the presence of chlorpromazine (Fig. 5b). Increase in cellular uptake after inhibitor treatment has also been reported by other investigators [20]. It is possible that other cellular uptake pathways that are not normally involved may be up-regulated in the presence of inhibitors. These results indicate that the clathrin-mediated pathway was not involved in the internalization of CS/DNA and CS/DNA/γ-PGA complexes.
Wortmannin is a phosphatidyl inositol-3-phosphate inhibitor [17], while cytochalasin D can inhibit actin polymerization and membrane ruffling [20]; both are known to be involved in macropinocytosis. Relative to the control, the reduction in internalization of CS/DNA complexes by cells pre-treated with wortmannin (cytochalasin D) was 3% (8%), implying that a minor fraction of test complexes could be internalized via micropinocytosis. In contrast, a greater degree of inhibition in cellular uptake of CS/DNA/γ-PGA complexes was observed (20% and 30% for the cells pre-treated with wortmannin and cytochalasin D, respectively, P < 0.05), suggesting that the incorporation of γ-PGA in CS/DNA complexes significantly enhanced their internalization by macropinocytosis.
Caveolae, a specialized type of lipid rafts, are flask-shaped invaginations in the plasma membrane enriched in proteins as well as cholesterol and spingolipids [31,32]. To discern any role that caveolae-mediated endocytosis may play in cellular uptake, two inhibitors,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
filipin and genistein, were used. Filipin (a sterol-binding pentaene macrolide antibiotic) selectively inhibits caveolae invagination by the formation of cholesterol precipitates, while genistein (a tyrosine kinase inhibitor) is used to block lipid raft-mediated endocytosis [16]. It has been reported that cholesterol and lipid rafts are involved in membrane trafficking [19,33].
As shown in Fig. 5a and 5b, treating cells with fillipin prior to incubation with CS/DNA or CS/DNA/γ-PGA complexes increased their cellular uptake. In contrast, cells pretreated with genistein resulted in a significant inhibition of the number of cells internalized [55% reduction for CS/DNA complexes and an enhanced reduction of 90% with the incorporation of γ-PGA (CS/DNA/γ-PGA)]. These data suggest that the internalization of both test complexes was caveolae-dependent and related with the lipid raft-mediated route, but not through the inhibition of invagination of caveolae.
The aforementioned results clearly show that the internalization of CS/DNA complexes took place by a combination of mechanisms, macropinocytosis and caveolae-mediated pathways at least; by incorporating γ-PGA in complexes (CS/DNA/γ-PGA), both uptake pathways were further enhanced. To further determine the effect of the internalization pathway on their transgene expression, the transfection efficiencies of CS/DNA/γ-PGA complexes (EGFP gene) in the presence of distinct inhibitors were studied. As shown in Fig. 5c, in the presence of wortmannin and cytochalasin D (macropinocytosis inhibitors), the transfection efficiencies of CS/DNA/γ-PGA complexes were reduced by 55% and 75%, respectively, while their transfection efficiency was suppressed by 90% when using a caveolae-mediated-pathway inhibitor, genistein (P < 0.05). These data suggest that caveolae-mediated pathway played a major role in the cellular uptake of CS/DNA/γ-PGA complexes and their subsequent transgene expression.
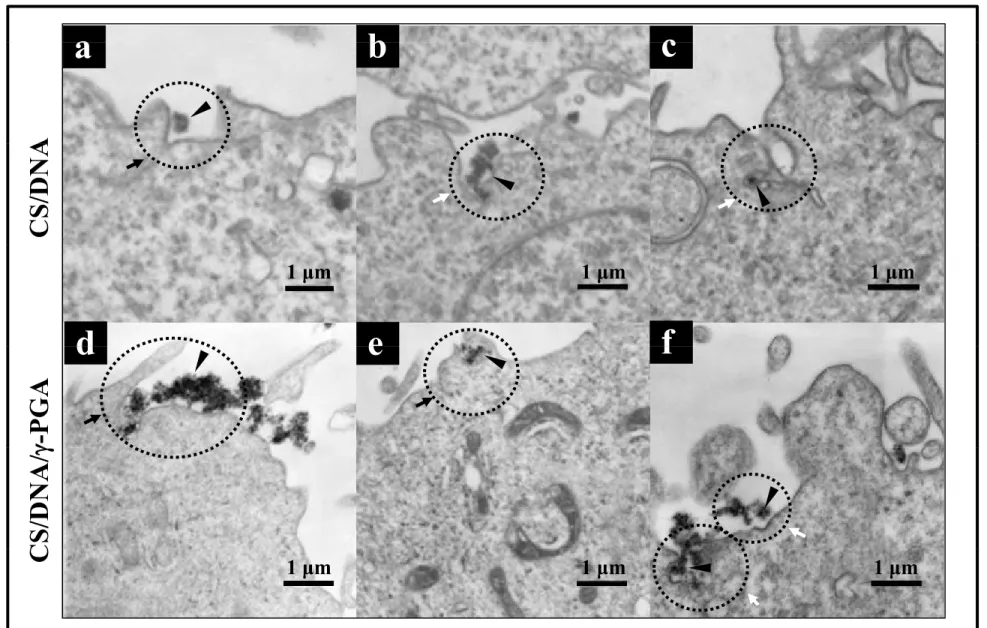
TEM was used to gain directly understanding of the mechanisms of internalization of CS/DNA and CS/DNA/γ-PGA complexes. As shown in Fig. 6a, macropinocytosis exerted its influence on the internalization of CS/DNA complexes. During macropinocytosis, membrane ruffling occurs; that is, the rims of the membrane folds extending from the
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
surface fuse back with the plasma membrane [34]. Additionally, TEM micrographs revealed flask-shaped caveolae in the plasma membrane (Fig. 6b and 6c), an indication of caveolae-dependent endocytosis [31]. Similar pathways were observed in the internalization of CS/DNA/γ-PGA complexes: the formation of lamellipodia (Fig. 6d and 6e) and caveolae-derived endocytic vesicles (Fig. 6f). These findings were consistent with those observed in the inhibition experiments (Fig. 5a and 5b).
3.5. Intracellular trafficking of test complexes
To track test complexes following their uptake, cells were treated with Cy3-labeled CS/DNA and CS/DNA/γ-PGA complexes individually and subsequently stained with Lysotracker; the intracellular localization of test complexeswas followed by CLSM. At 2 h after transfection, accumulation of Cy3-lableded complexes (red dots) was observed in most of the incubated cells in both studied groups (Fig. 7a); some complex aggregates were found entrapped within the lysosomal vesicles (green dots). The co-localization of test complexes with lysosomes produced a yellow fluorescence in the merged images. Interestingly, although there were more red dots (a greater cellular uptake) seen in the cytoplasm in the group treated with CS/DNA/γ-PGA complexes when compared with that transfected with CS/DNA complexes, a less percentage of their co-localization with lysosomes (yellow dots) was observed (50% for CS/DNA complexes vs. 30% for CS/DNA/γ-PGA complexes, Fig. 7b).
Degradation of complexes in lysosomes is one of the barriers for non-viral vectors mediated gene delivery [35]. It has been reported that macropinosomes can acidify but do not intersect with lysosomes, thus representing a potential alternative cell entry route of gene transfer for the avoidance of lysosomal degradation [36]. Additionally, the caveolae-mediated pathway has been proposed to be advantageous over the clathrin-mediated pathway for transfection DNA due to its possible avoidance of lysosomal degradation [35]. Some pathogens that use caveolae as their portal of entry escape delivery to and digestion in lysosomes [37]; it is thought that caveosomes lack the proper signal molecules required for interaction with other cellular compartments [38]. Compared with
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
their CS/DNA counterparts, a greater cellular uptake together with a less entry into lysosomes might explain the promotion of transfection efficiency of CS/DNA/γ-PGA complexes (Fig. 8).
4. Conclusions
The cellular uptake of CS-based complexes was significantly enhanced via the incorporation of γ-PGA. CS/DNA complexes were internalized via macropinocytosis and caveolae-mediated pathway. By incorporating γ-PGA in complexes (CS/DNA/γ-PGA), both pathways were significantly enhanced; however, the caveolae-mediated pathway played a major role. After internalization, the percentage of CS/DNA/γ-PGA complexes entry into lysosomes was significantly less than their CS/DNA counterpart and thus had an enhanced gene expression level. Knowledge of these mechanisms is critical for the development of efficient vectors for DNA transfection.
Acknowledgment
This work was supported by a grant from the National Science Council (NSC 99-2120-M-007-006), Taiwan, Republic of China.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 References
[1] Mao HQ, Roy K, Troung-Le VL, Janes KA, Lin KY, Wang Y, et al. Chitosan-DNA nanoparticles as gene carriers: Synthesis, characterization and transfection efficiency. J Control Release 2001;70:399–421.
[2] Kim TH, Park IK, Nah JW, Choi YJ, Cho CS. Galactosylated chitosan/DNA nanoparticles prepared using water-soluble chitosan as a gene carrier. Biomaterials 2004;25:3783–92.
[3] Morille M, Passirani C, Vonarbourg A, Clavreul A, Benoit JP. Progress in developing cationic vectors for non-viral systemic gene therapy against cancer. Biomaterials 2008;29:3477–96.
[4] Kim TH, Jiang HL, Jere D, Park IK, Cho MH, Nah JW, et al. Chemical modification of chitosan as a gene carrier in vitro and in vivo. Prog Polym Sci 2007;32:726–53.
[5] Peng SF, Yang MJ, Su CJ, Chen HL, Lee PW, Wei MC, et al. Effects of incorporation of poly( -glutamic acid) in chitosan/DNA complex nanoparticles on cellular uptake and transfection efficiency. Biomaterials 2009;30:1797–808.
[6] Kurosaki T, Kitahara T, Fumoto S, Nishida K, Nakamura J, Niidome T, et al. Ternary complexes of pDNA, polyethylenimine, and γ-polyglutamic acid for gene delivery systems. Biomaterials 2009;30:2846–53.
[7] Lee PW, Peng SF, Su CJ, Mi FL, Chen HL, Wei MC, et al. The use of biodegradable polymeric nanoparticles in combination with a low-pressure gene gun for transdermal DNA delivery. Biomaterials 2008;29:742–51.
[8] Adams CWM. Osmium tetroxide and the marchi method: Reactions with polar and non-polar lipids, protein and polysaccharide. J Histochem Cytochem 1960;8:262–7. [9] Ko IK, Ziady A, Lu S, Kwon YJ. Acid-degradable cationic methacrylamide polymerized
in the presence of plasmid DNA as tunable non-viral gene carrier. Biomaterials 2008:29:3872–81.
[10] Qaqish RB, Amiji MM. Synthesis of a fluorescent chitosan derivative and its application for the study of chitosan-mucin interactions. Carbohydr Polym *References
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 1999;38:99–107.
[11] Ho YP, Chen HH, Leong KW, Wang TH. Evaluating the intracellular stability and unpacking of DNA nanocomplexes by quantum dots-FRET. J Control Release 2006;116:83–9.
[12] Marrink SJ, de Vries AH, Mark AE. Coarse grained model for semiquantitative lipid simulations. J Phys Chem B 2004;108:750–60.
[13] Nelson MT, Humphrey W, Gursoy A, Dalke A, Kale LV, Skeel RD, et al. NAMD: A parallel, object oriented molecular dynamics program. Int J Supercomput Appl High Perform Comput 1996;10:251–68.
[14] Brooks BR, Bruccoleri RE, Olafson BD, States DJ, Swaminathan S, Karplus M. Charmm: A program for macromolecular energy, minimization, and dynamics calculations. J Comput Chem 1983;4:187–217.
[15] Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. UCSF Chimera-- A visualization system for exploratory research and analysis. J Comput Chem 2004;25:1605–12.
[16] Von Gersdorff K, Sanders NN, Vandenbroucke R, De Smedt SC, Wagner E, Ogris M. The internalization route resulting in successful gene expression depends on both cell line and polyethylenimine polyplex type. Mol Ther 2006;14:745–53.
[17] Araki N, Johnson MT, Swanson JA. A role for phosphoinositide 3-kinase in the completion of macropinocytosis and phagocytosis by macrophages. J Cell Biol 1996;135:1249–60.
[18] Parton RG, Joggerst B, Simons K. Regulated internalization of caveolae. J Cell Biol 1994;127:1199–215.
[19] Manunta M, Tan PH, Sagoo P, Kashefi K, George AJT. Gene delivery by dendrimers operates via a cholesterol dependent pathway. Nucleic Acids Res 2004;32:2730–9. [20] Perumal OP, Inapagolla R, Kannan S, Kannan RM. The effect of surface functionality
on cellular trafficking of dendrimers. Biomaterials 2008;29:3469–76.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
nanoparticles in paclitaxel-resistant human colorectal tumors. J Control Release
2006;112:312–9.
[22] Douglas KL, Piccirillo CA, Tabrizian M. Effects of alginate inclusion on the vector properties of chitosan-based nanoparticles. J Control Release 2006;115:354–61.
[23] Bond PJ, Holyoake J, Ivetac A, Khalid S, Sansom MSP. Coarse-grained molecular dynamics simulations of membrane proteins and peptides. J Struct Biol 2007;157:593–605.
[24] Bond PJ, Sansom MSP. Insertion and assembly of membrane proteins via simulation. J Am Chem Soc 2006;128:2697–704.
[25] Lieberman MW, Barrios R, Carter BZ, Habib GM, Lebovitz RM, Rajagopa-Ian S, et al. Gamma-glutamyl transpeptidase: What does the organization and expression of a multipromoter gene tell us about its functions? Am J Pathol 1995;147:1175–85.
[26] Magnan SDJ, Shirota FN, Nagasawa HT. Drug latentiation by -glutamyl transpeptidase. J Med Chem 1982;25:1018–21.
[27] Hanigan MH. Gamma-glutamyl transpeptidase, a glutathionase: Its expression and function in carcinogenesis. Chem Biol Interact 1998;111–112:333–42.
[28] Rejman J, Conese M, Hoekstra D. Gene transfer by means of lipo- and polyplexes: Role of clathrin and caveolae-mediated endocytosis. J Liposome Res 2006;16:237–47. [29] Douglas KL, Piccirillo CA, Tabrizian M. Cell line-dependent internalization pathways
and intracellular trafficking determine transfection efficiency of nanoparticle vectors. Eur J Pharm Biopharm 2008;68:676–87.
[30] Khalil IA, Kogure K, Akita H, Harashima H. Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol Rev 2006;58:32–45. [31] Parton RG, Simons K. The multiple faces of caveolae. Nat Rev Mol Cell Biol
2007;8:185–94.
[32] Mayor S, Pagano RE. Pathways of clathrin-independent endocytosis. Nat Rev Mol Cell
Biol 2007;8:603–12.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
biology and physics. Nat Cell Biol 2007;9:7–14.
[34] Mellman I. Endocytosis and molecular sorting. Annu Rev Cell Dev Biol 1996;12:575–625.
[35] Rejman J, Bragonzi A, Conese M. Role of clathrin- and caveolae-mediated endocytosis in gene transfer mediated by lipo- and polyplexes. Mol Ther 2005;12:468–74.
[36] Conner SD, Schmid SL. Regulated portals of entry into the cell. Nature 2003;422:37–44.
[37] Shin JS, Abraham S N. Caveolae as portals of entry for microbes. Microbes Infect 2001;3:755–61.
[38]Joiner KA, Fuhrman SA, Miettinen HM, Kasper LH, Mellman I. Toxoplasma gondii: fusion competence of parasitophorous vacuoles in Fc receptor-transfected fibroblasts. Science 1990;249:641–6.
Figure Captions
Figure 1. TEM micrographs of CS/DNA and CS/DNA/γ-PGA complexes; schematic illustrations of their internal structures obtained by small angle X-ray scattering (SAXS), modified from our previous study [5]. CS: chitosan; γ-PGA: poly(γ-glutamic acid).
Figure 2. (a) Kinetics of CS/DNA and CS/DNA/γ-PGA complexes mediated luciferase gene expressions (n = 3, *P < 0.05); (b) percentages of cellular uptake of FITC-labeled CS/DNA and CS/DNA/γ-PGA complexes (n = 3, *P < 0.05) and (c) their intracellular fluorescence intensities; (d) percentages of EGFP-expressing cells (n = 3, *P < 0.05). Lipofectamine: positive control; Control: the group without any treatment; CS: chitosan; γ-PGA: poly(γ-glutamic acid).
Figure 3. Results obtained by the molecular dynamic simulations showing the presence of (a) intramolecular hydrogen bondings in γ-PGA (the N-terminal amine group in γ-PGA is hidden) and (b) intermolecular hydrogen bondings between chitosan and γ-PGA (the N-terminal amine group in γ-PGA is exposed). Figure 4. Viability of the cells after being treated with distinct inhibitors, determined by the MTT assay (n = 5).
Control: the group without any inhibitor treatment.
Figure 5. Effects of inhibitors on the internalization of test complexes: (a) fluorescence intensities and (b) mean fluorescence intensities of intracellular uptake(n = 3, *P < 0.05); (c) EGFP intensities of cells pretreated with distinct inhibitors and then transfected with test complexes, determined by flow cytometry at 48 h after transfection (n = 3, *P < 0.05). Control: the group treated with test complexes only. CS: chitosan; γ-PGA: poly(γ-glutamic acid).
Figure 6. TEM images showing the internalization pathways of test complexes. Arrowheads indicate test complexes; black arrows macropinocytosis; and while arrows caveolae-mediated pathway. CS: chitosan; γ-PGA: poly(γ-glutamic acid).
Figure 7. (a) Images of the intracellular trafficking of test complexes observed by a confocal laser scanning microscope (scale bar: 8 μm); (b) quantitative analysis of Cy3-labeled test complexes colocalized with Lysotracker. CS: chitosan; γ-PGA: poly(γ-glutamic acid).
Figure 8. Schematic illustrations of potential mechanisms of internalization of (a) CS/DNA complexes and (b) CS/DNA/γ-PGA complexes. CS: chitosan; γ-PGA: poly(γ-glutamic acid).
Figure 1
CS/DNA
CS/DNA Complexes
DNA
CS/DNA
p
DNA
CS
γ-PGA
CS/DNA DomainCS/DNA/γ-PGA
CS/DNA/γ-PGA Complexes
(Compounded Complexes)
CS/γ-PGA Domain CS/DNA DomainCS/DNA and CS/γ-PGA Domains Bridged Domains Bridged
Figure 2
a
in
a
1.0E+10
CS/DNA
CS/DNA/γ-PGA
*
*
g Pr
ote
i
1.0E+09
*
*
vity/m
g
1 0E 08
*
se Acti
v
1.0E+08
u
cifera
s
1.0E+07
L
u
1.0E+06
15 min
30 min
1 h
2 h
Figure 2a100 110
Figure 2
b
*
d
60 70 80 90zed (%)
70
80
20 30 40 50 60l Internali
z
50
60
ted
*
0 10 20Cel
l
40
50
T
ransfec
t
Control CS/DNA90
c
20
30
of Cells
T
CS/DNA CS/DNA/γ-PGAE
vents
10
20
%
E
0
0
0
Figure 2bcdFigure 4
120
l)
100
110
f Contr
o
70
80
90
ty (%
of
50
60
70
ll V
iabili
30
40
Ce
l
0
10
20
0
Figure 4Figure 5
a
100 100Control (Cell only) CS/DNA CS/DNA+Chlorpromazine CS/DNA+Wortmannin CS/DNA/γ-PGA CS/DNA/γ-PGA+Chlorpromazine CS/DNA/γ-PGA+Wortmannin
Control (Cell only) CS/DNA CS/DNA+Cytochalasin D CS/DNA+Filipin CS/DNA/γ-PGA CS/DNA/γ-PGA +Cytochalasin D CS/DNA/γ-PGA +Filipin CS/DNA/γ-PGA +Genistein
Events
Events
CS/DNA+Wortmannin CS/DNA+Genistein γ 0 0 1 2 3 4 1000 101 102 103 104b
170 180 190Control Chlorpromazine Wortmannin
C t h l i D Fili i G i t i
l)
c
0 101 102 103 104 100 FL1 Fluorescence 0 FL1 Fluorescence 160 Control Wortmannin 100 101 102 103 104f Contr
ol)
120 130 140 150160 Cytochalasin D Filipin Genistein