EZH2 Regulates Neuronal Differentiation of Mesenchymal
Stem Cells through PIP5K1C-dependent Calcium Signaling
*
□SReceived for publication, September 14, 2010, and in revised form, January 6, 2011Published, JBC Papers in Press, January 7, 2011, DOI 10.1074/jbc.M110.185124
Yung-Luen Yu,abcd1,2Ruey-Hwang Chou,ac1Ling-Tzu Chen,cWoei-Cherng Shyu,efSu-Ching Hsieh,aChen-Shiou Wu,d
Hong-Jie Zeng,bSu-Peng Yeh,gDe-Ming Yang,hShih-Chieh Hung,iand Mien-Chie Hungabj3
From theaCenter for Molecular Medicine,eCenter for Neuropsychiatry, and thegDivision of Hematology and Oncology, Department of Medicine, China Medical University Hospital, Taichung 404, Taiwan, thebGraduate Institute of Cancer Biology,
fGraduate Institute of Immunology, and thedPh.D. Program for Cancer Biology and Drug Discovery, China Medical University,
Taichung 404, Taiwan, thecDepartment of Biotechnology, Asia University, Taichung 413, Taiwan, thehDepartment of Medical Research and Education, Taipei Veterans General Hospital, and Institute of Biophotonics, and theiStem Cell Laboratory,
Department of Medical Research and Education, Orthopaedics, and Traumatology, Taipei Veterans General Hospital, and Institute of Clinical Medicine, Institute of Pharmacology, National Yang-Ming University, Taipei 112, Taiwan, and thejDepartment of Molecular and Cellular Oncology, University of Texas M. D. Anderson Cancer Center, Houston, Texas 77030
Enhancer of zeste homolog 2 (EZH2) regulates stem cells renewal, maintenance, and differentiation into different cell lin-eages including neuron. Changes in intracellular Ca2ⴙ concen-tration play a critical role in the differentiation of neurons. How-ever, whether EZH2 modulates intracellular Ca2ⴙsignaling in
regulating neuronal differentiation from human mesenchymal stem cells (hMSCs) still remains unclear. When hMSCs were treated with a Ca2ⴙchelator or a PLC inhibitor to block IP3
-mediated Ca2ⴙ signaling, neuronal differentiation was
dis-rupted. EZH2 bound to the promoter region of PIP5K1C to sup-press its transcription in proliferating hMSCs. Interestingly, knockdown of EZH2 enhanced the expression of PIP5K1C, which in turn increased the amount of PI(4,5)P2, a precursor of
IP3, and resulted in increasing the intracellular Ca
2ⴙlevel,
sug-gesting that EZH2 negatively regulates intracellular Ca2ⴙ
through suppression of PIP5K1C. Knockdown of EZH2 also enhanced hMSCs differentiation into functional neuron both in
vitro and in vivo. In contrast, knockdown of PIP5K1C
signifi-cantly reduced PI(4,5)P2contents and intracellular Ca
2ⴙrelease
in EZH2-silenced cells and resulted in the disruption of neuro-nal differentiation from hMSCs. Here, we provide the first evi-dence to demonstrate that after induction to neuronal differen-tiation, decreased EZH2 activates the expression of PIP5K1C to evoke intracellular Ca2ⴙsignaling, which leads hMSCs to differ-entiate into functional neuron lineage. Activation of intracellu-lar Ca2ⴙsignaling by repressing or knocking down EZH2 might
be a potential strategy to promote neuronal differentiation from hMSCs for application to neurological dysfunction diseases.
Human mesenchymal stem cells (hMSCs)4 derived from bone marrow are easily obtained (1), safely expanded in vitro and are not susceptible to malignant transformation; thus, hMSCs are suitable for therapeutic applications (2). hMSCs can be induced to differentiate into multiple lineages, including bone, fat, cartilage, as well as neuron in vitro (1, 3–5). Trans-planted MSCs are able to differentiate into neuronal lineage and improve the functions of central nervous system (CNS) with ischemia (6), Parkinson disease (7), Alzheimer disease (8), spinal cord injury (9, 10), and other neurodegenerative disor-ders (11, 12) modeled in rodents. These findings demonstrate the potential of hMSCs for cell therapy in human CNS.
Spontaneous and transient elevation in intracellular Ca2⫹ concentration during an early period is required and critical for neuronal differentiation both in vitro and in embryonic neuron development (13). However, whether intracellular Ca2⫹ signal-ing is required for neuronal differentiation from hMSCs remains unclear. The expression of transient receptor potential (TRP) proteins, TRPC1 and TRPC3, is elevated to activate store-operated calcium entry (SOCE) after differentiation of H19 –7 hippocampal neuronal cells (14). It is also known that inositol 1,4,5-trisphosphate (IP3) stimulates a ligand-gated channel, IP3receptor (IP3R), to release Ca2⫹from intracellular stores in neurons (15). IP3 is generated by phospholipase C (PLC)-mediated hydrolysis of phosphatidylinositol 4,5-bis-phosphate [PI(4,5)P2], which is synthesized by type I phospha-tidylinositol-4-phosphate 5-kinase (PIP5K1) (16). PIP5K1 com-prises three isoforms namely␣ (PIP5K1A),  (PIP5K1B), and ␥ (PIP5K1C) (17). Unlike ubiquitously distributed PIP5K1A and PIP5K1B (18, 19), PIP5K1C is highly expressed in brain,
sug-*This study was supported, in whole or in part, by National Institutes of Health Grants R01 CA109311 and P01 CA099031. This work was also supported by The University of Texas M.D. Anderson-China Medical University and Hospital Sister Institution Fund, Cancer Center Research of Excellence DOH-99-TDC-111-05 (to M. C. H.); NSC-2632-B-039-001-MY3 (to M. C. H.); NSC-2320-B-039-032-MY3 (to Y. L. Y.); and NSC-3111-B-039 (to M. C. H., and Y. L. Y.).
□S
The on-line version of this article (available at http://www.jbc.org) contains supplemental Figs. S1 and S2.
1Both authors contributed equally to this work.
2To whom correspondence may be addressed. Tel.: 886-4-22052121; ext. 7933; Fax: 886-4-22333496; E-mail: [email protected].
3To whom correspondence may be addressed. Tel: 792-3668; Fax: 713-794-3270; E-mail: [email protected].
4The abbreviations used are: hMSC, human mesenchymal stem cell; EZH2, enhancer of zeste homolog 2; PIP5K1C, type I phosphatidylinositol-4-phosphate 5-kinase isoform C; PLC, phospholipase C; PI(4,5)P2, phospha-tidylinositol 4,5-bisphosphate; IP3, inositol 1,4,5-trisphosphate; DAG, dia-cylglycerol; ESCs, embryonic stem cells; CSCs, carcinoma stem cells; CNS, central nervous system; TRP, transient receptor potential; SOCE, store-op-erated calcium entry; PcG, polycomb group; PRCs, polycomb repressive complexes; SUZ12, suppressor of zeste-12; EED, embryonic ectoderm development; H3K27me3, histone H3 tri-methylation at lysine 27; H2AK119, histone 2A at lysine 119; ChIP, chromatin immunoprecipitation; shRNA, short-hairpin RNA; IP3R, IP3receptor; RyR, ryanodine-sensitive receptor.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 11, pp. 9657–9667, March 18, 2011 © 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
at China Medical University, on September 22, 2011
www.jbc.org
gesting that PIP5K1C may play a distinctive role in neuron functions (20, 21). Moreover, it has been shown that PIP5K1C is critical in the regulation of IP3-mediated Ca
2⫹signaling after stimulation of G protein-coupled receptor by histamine (22) and is required for cardiovascular and neuronal development (23). Nonetheless, the regulation of PIP5K1C and intracellular Ca2⫹transient during neuronal differentiation from hMSCs is still unclear.
The polycomb group (PcG) proteins consist of two function-ally distinct multimeric polycomb repressive complexes (PRCs) referred to as PRC1 and PRC2 (24). PRC2 binds to target genes and initiates tri-methylation at lysine 27 of histone H3 (H3K27me3); PRC1 recognizes the H3K27me3 through its chromodomain to mediate ubiquitylation of H2AK119 and maintain gene repression (25). The core components of PRC2 include suppressor of zeste-12 (SUZ12) and embryonic ecto-derm development (EED) for complex stability and enhancer of zeste homolog 2 (EZH2) for the methyltransferase activity (26). EZH2 not only catalyzes histone H3 tri-methylation at lysine 27 (27) but also recruits DNA methyltransferases to silence gene expression (28). The PcG proteins play a critical role in the regulation of gene expressions in stem cells maintenance and lineage specification (29). For example, EZH2 is highly expressed in embryonic stem cells (ESCs) and is required for the derivation of pluripotent ESCs (30). EZH2 is also involved in differentiation into different cell lineages in mice, including skeletal muscle (31, 32), hepatocytes (33), epidermis (34), as well as neuron (35). However, whether and how EZH2 regulates differentiation of hMSCs into neuronal lineage remain to be explored.
In the current study, we demonstrate that EZH2 targets PIP5K1C promoter to suppress its expression and negatively regulates intracellular Ca2⫹concentration to maintain homeo-stasis in proliferating hMSCs. After induction to neuronal dif-ferentiation, dissociation of EZH2 protein from PIP5K1C pro-moter induces the expression of PIP5K1C to elevate intracellular Ca2⫹contents and promote neuronal differentia-tion. This is the first evidence to show that PIP5K1C-mediated Ca2⫹signaling in neuronal differentiation from hMSCs is reg-ulated by EZH2.
EXPERIMENTAL PROCEDURES
Materials—Chemicals used in induction of neuronal
differ-entiation, dexamethasone, ascorbic acid-2-phosphate, indo-methacin, insulin, and 3-isobutyl-1-methyl-xanthine, an intracellular Ca2⫹ chelator, 1,2-bis(2-aminophenoxy)-ethane-N,N,N⬘,N⬘-tetraacetic acid tetrakis-acetoxymethyl ester (BAPTA-AM), polybrene, and puromycin were pur-chased from Sigma. The cell-permeant Ca2⫹indicator, Fluo-4-acetoxymethyl ester (Fluo4-AM) and Fura-2-Fluo-4-acetoxymethyl ester (Fura-2 AM) were obtained from Molecular Probes. The following antibodies were purchased from commercial compa-nies: CD105, MAP2, AlexFluor 488 conjugated anti-MAP2A and AlexFluor 647 conjugated anti--tubulin III (BD); anti--tubulin III (Covance); anti-Neu-N, clone A60 (Chemicon); anti--actin and ␣-tubulin (Sigma); anti-EZH2, clone AC22, and anti-PIP5K1C (Cell Signaling).
Cell Culture and Induction to Differentiation into Neuronal
Lineage—Human mesenchymal stem cells (hMSCs),
size-sieved stem cells from human bone marrow, were isolated and characterized previously, and the hMSCs had the capacity for multi-lineage potential to form bone, fat, cartilage (1), as well as electrically active neural cells (3). The hMSCs were immortal-ized without neoplastic transformation by transduction with HPV16 E6/E7 genes (5). The E6/E7-immortalized hMSC deriv-ative named 3A6 contains the human telomerase reverse tran-scriptase (hTERT) gene for more stem-like properties (36). The 3A6 cells were grown in Dulbecco’s modified Eagle’s medium-low glucose (DMEM-LG) (HyClone) with 10% fetal bovine serum (FBS), 100 units/ml penicillin, and 100g/ml streptomy-cin in a humidified incubator with 5% CO2at 37 °C. To induce neuronal differentiation, cells were seeded at a density of 4000 cells/cm2in the regular medium the day before experiment, and then treated with serum-free DMEM-high glucose (DMEM-HG) (HyClone) neuronal induction medium (NIM) containing 10⫺7Mdexamethasone, 50g/ml ascorbic acid-2-phosphate, 50Mindomethacin, 10g/ml insulin, and 0.45 mM
3-isobutyl-1-methyl-xanthine for 1–5 days, and the neuronal induction medium was changed per 3 days (37).
Western Blot Analysis—Cells were washed twice with
phos-phate-buffered saline (PBS, containing 137 mMNaCl, 2.7 mM
KCl, 10 mMNa2HPO4, 2 mMKH2PO4), and then lysed in RIPA
buffer (50 mMTris at pH 7.5, 150 mMNaCl, 1 mMEDTA, 0.25%
Na-deoxycholate, 1% Nonidet P-40, 1 mMNaF, 1 mMNa3VO4, 1 mM PMSF, 1g/ml aprotinin) by sonication. The soluble
extraction was collected from the supernatant after centrifuga-tion at 15,000⫻ g for 10 min at 4 °C. The extract was boiled at 100 °C for 5 min, separated by SDS-PAGE, and transferred to a PVDF membrane. Subsequently, the membrane was blocked with 5% skim milk in PBST buffer (PBS containing 0.1% Tween-20) for 1 h at room temperature, and then hybridized with pri-mary antibody with gentle agitation overnight at 4 °C. After washing with PBST, the membrane was incubated with HRP-conjugated secondary antibody (Chemicon) for 1 h at room temperature. The immunoreactive band was visualized by the enhanced chemiluminescence (ECL) detection reagent (GE Healthcare, Amersham Biosciences Place).
RNA Extraction and Reverse Transcriptase-Polymerase
Chain Reaction (RT-PCR)—Total RNA was extracted with
TRIzol reagent (Invitrogen) according to the manufacturer’s instruction. The complementary DNA (cDNA) was synthe-sized from 5g of total RNA in a reaction mixture containing 2.5Moligo (dT) primer, 0.5 mMdNTP mixture, 200 units of
SuperScript III reverse transcriptase, 40 units of RNaseOUT, an RNase inhibitor (all from Invitrogen). After incubation at 50 °C for 50 min, the reaction mixture was heat inactivated at 85 °C for 5 min and then treated with 2 units of RNase H at 37 °C for 20 min. The quantitative PCR (q-PCR) was performed by detec-tion of the hydrolyzed fluorescent probes from Universal Pro-beLibrary (UPL; Roche) using the LightCycler 480 equipment (Roche). Primers and UPL probes were designed using the ProbeFinder software. The primer sets and the matched UPL probe numbers are as following: PIP5K1C: 5 ⬘-CGCCACCGA-CATCTACTTTC-3⬘ (forward), 5⬘-ATAGTGGAGCGGGG-AGTACA-3⬘ (reverse), and UPL probe 17; -actin (ACTB):
at China Medical University, on September 22, 2011
www.jbc.org
5⬘-ATTGGCAATGAGCGGTTC-3⬘ (forward), 5⬘-GGATGC-CACAGGACTCCAT-3⬘ (reverse), and UPL probe 11. The q-PCR was examined by incubating the cDNA in a reaction mixture containing,0.5Mof each primer, 0.1MUPL probe, and 1-fold concentration of Probes Master reagent (Roche). The amplification conditions were initial denaturation at 95 °C for 10 min, followed by 45 cycles of 95 °C for 10 s, 55 °C for 30 s, and 72 °C for 1 s. The fluorescent signal was detected at 72 °C step of each cycle. The relative quantification of the gene of inter-est was normalized by-actin and calculated by the value of cross-point (CP) in each fluorescence curve of each gene. For traditional PCR, the reaction mixture containing 2l of cDNA, 0.2 mMdNTP
mixture, 2Mof each primers, 1 unit of TaqDNA polymerase, and
1-fold concentration of ThermalPol Buffer (New England Bio-Labs) was started by denatured 95 °C for 5 min, followed by ampli-fication of indicated cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s. The numbers of cycles for amplification of NSE, PITX-3, NURR1, and EZH2 were all 35, and that of-actin was 25. The specific primer sequences for these genes are as follows: NSE: 5 ⬘-CATCGACAAGGCTGGCTACACG-3⬘ (forward), 5⬘-GACAG-TTGCAGGCCTTTTCTTC-3⬘ (reverse); PITX-3: 5⬘-GTCTAT-CGGGACCCGTGTAA-3⬘ (forward), 5⬘-CCAGTCAAAATGA-CCCCAGT-3⬘ (reverse); NURR1: 5⬘-CAATGCGTTCGTG-GCT-3⬘ (forward), 5⬘-GGGTACGAAGTTCTGGG-3⬘ (reverse); EZH2: 5⬘-CAGTAAAAATGTGTCCTGCAAGAA-3⬘ (forward), 5⬘-TCAAGGGATTTCCATTTCTCTTTCGA-3⬘ (reverse); and -actin: GCACTCTTCCAGCCTTCCTTCC-3⬘ (forward), 5⬘-TCACCTTCACCGTTCCAGTTTTT-3⬘ (reverse).
Ca2⫹Influx ([Ca2⫹]
i) Measurement—Ca2⫹influx
measure-ment was carried out as previously described (3). The hMSCs were seeded on coverslips. After treatment, cells were incu-bated with 5MFura-2 AM for 1 h and then washed out the
excess Fura-2 AM. The coverslip was mounted on an inverted microscope (Olympus IX-70) under ⫻40 magnification. The excitation peak for Fura-2 shifts from 380 nm for the calcium-free chelator to about 340 nm for the calcium-saturated form. The excitation lights (340 nm and 380 nm) were provided by T.I.L.L. Polychrome IV spectrophotometer (T.I.L.L. Photon-ics), and the emission light (510 nm) was recorded by Princeton charged-coupled device camera CCD-130 (Roper Scientific). The [Ca2⫹]
iwas represented as a ratio (R340/380) between emis-sion intensities at 510 nm induced by 340 nm and 380 nm. For stimulation, loading buffer contained glutamate (25 M) or
high K⫹(the NaCl was replaced with an equimolar KCl) was puffed from a glass micropipette placed right beside the cell. The opening of the micropipette was⬃1.5m. The puff pres-sure was controlled by Eppendorf Microinjector 5242 (Eppen-dorf) with a P2 setting of 200 hPa.
In addition, we also used another method to measure [Ca2⫹]i
(38). Cells were seeded on the Biocoat poly-D-lysine 96-well
black/clear plate (BD) for 2 days, and then induced to neuronal differentiation for 5 days. Cells were pre-incubated with 5M
Fluo4-AM for 1 h. After washout of Fluo4-AM, cells were stim-ulated with 100Mglutamate (Sigma) in a calcium buffer
solu-tion containing 5 mMEGTA, 100 mMKCl, and 30 mMMOPS
with free 0.15 M Ca2⫹ (Sigma). The plates were measured
immediately at excitation (485 nm) and emission (528 nm)
every 40 s interval using Synergy 2 microplate reader (BioTek Instruments, Inc.).
Chromatin Immunoprecipitation (ChIP) Assay—ChIP assay
was performed with the EZ ChIP kit (Upstate). In brief, chro-matin and proteins from approximate 2⫻ 106cells were cross-linked with 1% formaldehyde for 10 min at room temperature. These cells were collected, lysed, and sonicated on ice to shear the chromatin DNA to a length between 200 bp and 1000 bp using Sonicator 3000 (Misonix) equipped with a microtip (set-ting: output level 4, 5 times of pulse for 10 s, and pause for 1 min). The sonicated chromatin lysate was immunoprecipitated with anti-EZH2 antibody, and collected with protein A/G-aga-rose beads (Pierce). The protein/DNA crosslinks of the immu-noprecipitated complexes were reversed by incubation in 0.2M
NaCl at 65 °C for 4 –5 h, and then the DNA was purified and applied to PCR as described above to determine the binding ability of EZH2 to its potential target gene, PIP5K1C. The sequences of the primers specific to the promoter of PIP5K1C are 5⬘-GACCTACACAGCACATGCCA-3⬘ (forward), and 5⬘-GCATGTATTGTGCATATCCG-3⬘ (reverse).
Gene Knockdown by Short-hairpin RNA (shRNA)
—Knock-down of genes were performed with the specific shRNAs deliv-ered by the lentiviral system from National RNAi Core Facility (Academia Sinica, Taipei, Taiwan) according to the instruction manual. Briefly, to generate the lentivirus containing specific shRNA, 293T cells were cotransfected with 2.25g of pCMV-⌬R8.91 plasmid harboring Gag and Pol genes, 0.25 g of pMD.G plasmid containing VSV-G gene for expression of envelope glycoprotein, and 2.5g of pLKO.1 plasmid bearing the specific shRNA for 16 h, and cells were then cultured in growth medium containing 1% BSA for another 24 h. The cul-tured medium containing lentivirus was collected and stored at ⫺80 °C as aliquots for further use. To deliver the specific shRNA construct, approximate 80% confluent cells were infected with the lentivirus bearing specific shRNA in growth medium containing 8g/ml polybrene and incubated at 37 °C for 24 h. Afterward, cells were subcultured and selected with 2 g/ml puromycin. The shRNA constructs targeting the inter-ested genes are as follows: PIP5K1C: TRCN0000037666 corre-sponding to the sequences, 5 ⬘-CGTGGTCAAGATGCACCT-CAA-3⬘; EZH2: TRCN0000040076 referring to the sequence, 5⬘-CGGAAATCTTAAACCAAGAAT-3⬘. The shRNA con-struct against luciferase (shLuc), TRCN0000072244 referring to the sequence, 5⬘-ATCACAGAATCGTCGTATGCA-3⬘ was used as negative control.
Detection of Intracellular Ca2⫹ by Flow Cytometer—
Fluo4-AM dissolved in DMSO was used to detect intracellular Ca2⫹(39), which bound to cytoplasmic-free Ca2⫹and emitted a green fluorescence (peak at 516 nm). Cells were seeded on 35-mm dishes at a density of 104cells/dish the day before exper-iment and incubated with 2MFluo4-AM at 37 °C for 30 min in the dark. Cells were then washed with PBS and incubated at 37 °C for further 1 h to allow for complete deesterification of the dye by cytosolic esterases inside cells. Subsequently, cells were collected and applied to detect the fluorescence of Ca2⫹bound to Fluo4 within cells using FL1 of the FACSCalibur flow cyto-metry (BD Biosciences). Simultaneously, cells were treated with DMSO in parallel as a negative control in each treatment. The
at China Medical University, on September 22, 2011
www.jbc.org
intensity of the fluorescence from Fluo4-AM treated cells was normalized by the background from DMSO-treated cells, and the geographic mean value (Geo Mean) of fluorescence in each experiment was calculated as the value of normalized Geo Mean using CellQuest program (BD Biosciences).
PI(4,5)P2Measurement—PI(4,5)P2mass was determined by
the PI(4,5)P2mass ELISA kit (Echelon Biosciences Inc.) accord-ing to the manual instruction (40). In brief,⬃2 ⫻ 106cells were collected, and lipids were extracted. The extracted lipids were incubated with a PI(4,5)P2detector protein at room tempera-ture for 1 h, and then transferred to the PI(4,5)P2-coated plate for competitive binding, and incubated at room temperature for another 1 h. After removing the reaction mixture and wash-ing three times with PBST, the peroxidase-linked secondary
detection reagent was added into the wells, and the plate was incubated at room temperature for 1 h to detect the PI(4,5)P2 detector protein bound to the plate. Finally, after discarding the reaction mixture from the plate, the 3,3 ⬘,5,5⬘-tetramethylben-zidine (TMB) substrate was added and incubated for 5–30 min to allow the color development, and the absorbance at 450 nm was measured by Synergy 2 microplate reader (BioTek Instru-ments, Inc.). The colorimetric signal is inversely proportional to the amounts of PI(4,5)P2.The amounts of PI(4,5)P2in each sample was calculated according to the standard curve from serious diluents of pure PI(4,5)P2.
Neuronal Differentiation from hMSCs in Animal Brains—
Adult male Sprague-Dawley (SD) rats (weight⬎ 300 ⫻ g; age, 7– 8 weeks) were used in this study. All animal experiments FIGURE 1. Characterization of neuronal differentiation from hMSCs. 3A6 hMSCs were seeded at a density of 4000 cells/cm2the day before induction of neuronal differentiation, and then treated with NIM for 1–5 days as described under “Experimental Procedures.” A, cell morphology at indicated time interval was observed at⫻100 magnification under an inverted phase microscope. B, protein expressions of a MSC marker, CD105, and neuron markers,-tubulin III, Neu-N, and MAP2 were examined. C, mRNA expressions of neuron markers, NSE, PITX3, and NURRI were determined as well.-Actin was used as an internal control. D, expressions of neuronal markers,-tubulin III and MAP2, were labeled by AlexFluor 647- and AlexFluor 488-conjugated antibodies, respectively, and then detected by flow cytometry. The-tubulin III- and MAP2-positive cells in comparison with the cells labeled with isotype IgG were margined as M1 region. The bottom bar plots show the percentage of cells in the M1 region in un-differentiated and neuronal differentiated hMSCs. E, hMSCs were treated with (solid
line) or without (dotted line) NIM for 5 days to induce differentiation into the neuron lineage. Change of [Ca2⫹]iafter stimulated with glutamate (upper panel) or
high K⫹buffer (lower panel) was used to determine the neuron-like function. The arrow indicates the time point of stimulation.
at China Medical University, on September 22, 2011
www.jbc.org
were approved by the Institutional Review Board of Animal Experiments, China Medical University Hospital. Prior to implantation, hMSCs with or without knockdown of EZH2 were incubated with 1 g/ml bis-benzimide Hoechst 33342 (Sigma) to label nuclei with blue fluorescence for 5 h at 37 °C. After washing three times with PBS, the labeled hMSCs were counted using a cytometer to ensure an adequate cell number for implantation. The SD rats were anesthetized with chloral hydrate (0.4 g/kg, intraperitoneal) and then were injected stereotaxically with approximate 1 ⫻ 106cells in 3–5l of DMEM medium through a 26-gauge Hamilton syringe into 3 cortical areas adjacent to the right middle cerebral artery, 3.0 to 5.0 mm below the dura mater. Cyclosporin A, an immu-nosuppressant drug, (CsA; 1 mg/kg/d, intraperitoneal; Novartis) injections were given daily to each experimental rat for 3 weeks (41).
Laser Scanning Confocal Microscopy for Immunofluorescence
Colocalization Analysis—To determine the neuronal
differen-tiation of hMSCs in rat brain, tissue sections from hMSCs implanted rat brains were stained with specific primary anti-bodies (1:300) against EZH2 and a neuronal specific marker, MAP2. Subsequently, the tissue sections were stained with Cy5- and Cy3-conjugated secondary antibodies (1:500, Jackson Immunoresearch). The immunofluorescent colocalization study with three-dimensional images was performed to test for the expression of hMSC (blue fluorescence), EZH2 (red fluo-rescence), and MAP2 (green fluorescence). The three-dimen-sional images were observed under a Carl Zeiss LSM510 laser-scanning confocal microscope.
RESULTS
Characterization of Neuronal Differentiation from 3A6
hMSCs—The immortalized hMSC line, KP-hMSCs, possesses
the characteristics for differentiation into several cell types, including fat, cartilage, and neuron (5). We further validated the capacity of its derivative line, 3A6, containing the human telomerase reverse transcriptase (hTERT) gene for more stem-like properties (36) and induction into neuronal lineage. Exten-sion of neurite was observed at 1 day after induction in the neuronal induction medium (NIM), and the length of neurite outgrowth was increased after 3–5 days of treatment (Fig. 1A). During the induction of neuronal differentiation, protein expression of neuron markers, -tubulin III, Neu-N, and MAP2, and mRNA expression of NSE, PITX3, and NURR1 increased with time (Fig. 1, B and C). However, the expression of MSC marker, CD105, was simultaneously reduced (Fig. 1B). The flow cytometry results showed that the percentage of cells expressing-tubulin III and MAP2 dramatically increased to 55⬃65% after induction of differentiation for 5 days (Fig. 1D). To further confirm that hMSCs possess the potential to differ-entiate into functional neuronal cell, neuron-like function was validated by measuring the [Ca2⫹]i change after stimulation
with glutamate or high K⫹. As shown in Fig. 1E, elevated [Ca2⫹]istimulated by glutamate or high K⫹was observed
sig-nificantly in differentiated hMSCs. These results demonstrated that the 3A6 hMSC line could be successfully induced to differ-entiate into functional neuron lineage.
EZH2 Negatively Regulates Intracellular Ca2⫹Required for
hMSCs Neuronal Differentiation—To investigate whether
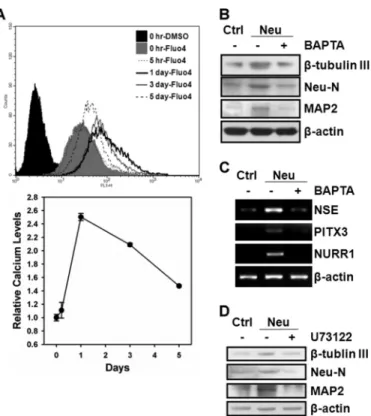
intracellular Ca2⫹change is required for neuronal differentia-tion from hMSCs, we measured the intracellular Ca2⫹contents with Fluo4-AM during differentiation and examined the effect of an intracellular Ca2⫹chelator, BAPTA-AM, on neuronal dif-ferentiation. As shown in Fig. 2A, intracellular Ca2⫹contents were transiently elevated during neuronal differentiation, which peaked at day 1 post-induction and declined afterward. hMSCs treated with BAPTA-AM significantly decreased the expression of neuron markers after induction to neuronal dif-ferentiation as demonstrated by both Western blot (Fig. 2B) and RT-PCR (Fig. 2C). To further show that hMSCs neuronal differentiation involves IP3-mediated Ca2⫹signaling, we used a PLC inhibitor, U73122, to disrupt PI(4,5)P2hydrolysis into IP3 and diacylglycerol (DAG) and examined its effect on neuronal differentiation. Cells that were treated with U73122 suppressed the expression of neuron markers after induction to neuronal differentiation (Fig. 2D), supporting that IP3-mediated Ca2⫹ signaling is required for inducing differentiation from hMSCs into neuronal lineage. We silenced the EZH2 gene by specific short hairpin RNA (shRNA) (Fig. 3A) to further understand the role of EZH2 in the regulation of intracellular Ca2⫹contents. Specifically, knockdown of EZH2 dramatically increased the intracellular Ca2⫹ FIGURE 2. hMSCs neuronal differentiation requires IP3-mediated
intra-cellular Ca2ⴙsignaling. A, intracellular Ca2⫹contents during neuronal
dif-ferentiation from 3A6 hMSCs was measured with Fluo4-AM by flow cytome-ter. The bottom plot is the relative intracellular Ca2⫹level comparing to that at the initial induction time (0 h) during neuronal differentiation. B and C, 3A6 hMSCs were pretreated with or without 10Mintracellular Ca2⫹chelator, BAPTA-AM, for 1 h, and then induced to neuronal differentiation for 5 days. The protein (B) and mRNA (C) expression of the indicated neuron markers in undifferentiated (lane 1) and neuron-differentiated (lanes 2 and 3) were deter-mined. D, 3A6 hMSCs were pretreated with or without 20MPLC inhibitor, U73122, for 30 min, and then induced to neuronal differentiation for 5 days. The expression of neuron markers were examined after each treatment.
at China Medical University, on September 22, 2011
www.jbc.org
content by 14-fold, indicating that EZH2 might be a negative reg-ulator of intracellular Ca2⫹content in proliferating undifferenti-ated hMSCs (Fig. 3B). In addition, treating EZH2-silenced hMSCs with U73122 inhibited about 70% of intracellular Ca2⫹contents compared with untreated cells (Fig. 3C), suggesting that EZH2 diminished the intracellular Ca2⫹content that is likely through repression of IP3-mediated Ca
2⫹release.
EZH2 Negatively Regulates Intracellular Ca2⫹through
Sup-pression of PIP5K1C—Because IP3-mediated Ca2⫹signaling is
required for neuronal differentiation from hMSCs (Fig. 2D), and PIP5K1C synthesizes a precursor of IP3, PI(4,5)P2, which is critical in the process (22) and highly expressed in brain (20, 21), we questioned whether EZH2 affects intracellular Ca2⫹ contents through regulation of PIP5K1C gene expression. To address whether EZH2 targets PIP5K1C gene promoter, we performed ChIP with anti-EZH2 antibody followed by amplifi-cation of the promoter region of PIP5K1C. EZH2 bound to the promoter of PIP5K1C in undifferentiated hMSCs but not
neu-ron-differentiated hMSCs. However, after induction to neuro-nal differentiation, binding of EZH2 on PIP5K1C promoter was significantly reduced (Fig. 4A). Knockdown of EZH2 dramati-cally increased the expression of PIP5K1C (Fig. 4, B and C), indicating that EZH2 targets the promoter of PIP5K1C and represses its expression in proliferating undifferentiated hMSCs. To further support our hypothesis that EZH2 nega-tively regulates intracellular Ca2⫹signaling through PIP5K1C, we knocked down PIP5K1C in EZH2-silenced cells (Fig. 4D) and determined the effect on intracellular Ca2⫹contents. Infec-tion of increasing amount of lentivirus carrying shRNA against PIP5K1C (1- versus 3-fold; middle panel, Fig. 4D) in EZH2-silenced hMSCs resulted in significant reduction of intracellu-lar Ca2⫹contents caused by knockdown of EZH2 in a dose-de-pendent manner (up to 90% of inhibition) (Fig. 4E). In addition, knockdown of EZH2 increased the amount of the product of PIP5K1C, PI(4,5)P2, which was abolished by further knock-down of PIP5K1C (Fig. 4F). Taken together, these results FIGURE 3. Negative regulation of IP3-mediated intracellular Ca
2ⴙcontents by EZH2 in hMSCs. A, 3A6 hMSCs were infected without (lane 1) or with
lentivirus carrying shRNAs against luciferase (lane 2) or EZH2 (lane 3) gene. The shRNA against luciferase was used as a negative control of shRNA. The expressions of EZH2 and␣-tubulin were examined. B, effect of EZH2 knockdown on intracellular Ca2⫹contents was determined by flow cytometer. The bottom
plot showed the ratio of intracellular Ca2⫹contents from shRNA-containing cells over mock control cells. Bars represented mean⫾ S.D., and the symbol, **, indicated p value⬍ 0.01 by t test. C, EZH2-silenced hMSCs were treated with or without 20MU73122 for 30 min, and then the intracellular Ca2⫹contents were determined. The bottom plot is the relative intracellular Ca2⫹level normalized by that from untreated cells.
at China Medical University, on September 22, 2011
www.jbc.org
showed that EZH2 negatively regulates intracellular Ca2⫹ con-tents by suppressing PIP5K1C expression.
PIP5K1C Is Required for hMSCs Neuronal Differentiation—
To explore the role of the EZH2 target gene, PIP5K1C, in neu-ronal differentiation from hMSCs, we silenced PIP5K1C gene expression by shRNA to determine its effect on neuronal dif-ferentiation. As shown in Fig. 5, A and B, the expression of PIP5K1C increased after inducing hMSCs to neuronal differen-tiation, which peaked at day 1 and declined at days 3 and 5 post-induction. The shRNAs against PIP5K1C showed 98% inhibition of its mRNA expression (Fig. 5C). Likewise, we also observed a significant decrease in PIP5K1C protein expression by Western blot (Fig. 5D). To clarify the influence of PIP5K1C on neuronal differentiation from hMSCs, we compared the expression of neuron markers in 3A6 hMSCs transfectants, which had been infected with or without shRNA against Luc (as negative control) or PIP5K1C, at day 0 and day 5 post-induction to neuronal differentiation. The expression of PIP5K1C as well as neuron markers, Neu-N and MAP2, was significantly
increased in mock-treated and Luc-silenced cells but not in PIP5K1C-silenced cells (Fig. 5E), and the intracellular Ca2⫹ content was not significantly changed in PIP5K1C-silenced cells (Fig. 5F), indicating that PIP5K1C is required for intra-cellular Ca2⫹ transient and neuronal differentiation from hMSCs.
Knockdown of EZH2 Enhances Neuronal Differentiation from
hMSCs in Vitro and in Vivo—To address the potential of
acti-vation of Ca2⫹signaling to enhance hMSCs differentiation into functional neuron, we compared the capabilities of neuronal differentiation from hMSCs with or without knockdown of EZH2. As shown in Fig. 6A, knockdown of EZH2 increased the expression of neuron markers (NSE, PITX3, and NURR1) and PIP5K1C (upper panel), and the intracellular Ca2⫹content was also elevated (lower panel). Further knockdown of PIP5K1C in EZH2-silenced cells disrupted neuronal differentiation as indi-cated by loss of neuron marker expression and attenuated the elevation of Ca2⫹ transient. The neuron-like function mea-sured by change of [Ca2⫹]iafter glutamate stimulation was
sig-FIGURE 4. The role of PIP5K1C in EZH2-mediated calcium signaling. A, undifferentiated and neuron-differentiated 3A6 hMSCs were collected and applied to ChIP assay with anti-EZH2 antibody or immunoglobulin (IgG, negative control). The immunoprecipitated chromatin DNA was amplified by PCR with primers specific to the promoter region of PIP5K1C gene. B and C, effects of EZH2 knockdown on the expression of PIP5K1C was determined by quantitative RT-PCR (bar3 in B) and Western blotting (lane 3 in C). D, 3A6 hMSCs were infected with or without lentivirus containing shRNAs against luciferase or EZH2, and EZH2-silenced cells were infected with increased amounts of lentivirus harboring shRNA to PIP5K1C (1- and 3-fold amounts). Total RNA was extracted and the expressions of EZH2, PIP5K1C, and-actin were determined by RT-PCR. E, effect of PIP5K1C knockdown in EZH2-silencing cells on intracellular Ca2⫹contents was examined by flow cytometer. The bottom plot was the relative quantity of intracellular Ca2⫹contents. The bars represented mean⫾ S.D., and the symbol, **, indicated p value ⬍ 0.01 by t test. F, 3A6 hMSCs were infected with or without lentivirus containing shRNAs against luciferase or EZH2, and PIP5K1C was further knocked down in EZH2-silenced cells. The PI(4,5)P2mass after each treatment was determined as described under “Experimental Procedures.”
at China Medical University, on September 22, 2011
www.jbc.org
nificantly higher in EZH2-silenced hMSCs than that in parental hMSCs, and knockdown of PIP5K1C in both parental and EZH2-silenced hMSCs disturbed the neuron-like function (Fig. 6, B and C). These results suggest that knockdown of EZH2 enhanced hMSCs differentiation into functional neuron, and this is likely due to the activation of PIP5K1C-mediated Ca2⫹ signaling. To further investigate the effect of EZH2 knockdown in neuronal differentiation in vivo, we implanted hMSCs with or without knockdown of EZH2 to the brains of SD rats for 3 weeks and examined the expression of MAP2 in their tissue sections. Images from the colocalization study (three-dimen-sional image) showing exogenous implanted hMSCs and MAP2 (green fluorescence) positive cells in implanted hMSCs (blue fluorescence) as indicated by the arrows. Quantitative analysis of the implanted MAP2-positive cells in EZH2-silenced hMSCs was significantly more than that in the mock-treated hMSCs (Fig. 6E), suggesting that knock-down of EZH2 in hMSCs might be a potential strategy to enhance neuronal differentiation in vivo.
DISCUSSION
Although the involvement of Ca2⫹in neuronal differentia-tion has been reported from 1990s (42), it remains unclear whether intracellular Ca2⫹transient is required for differenti-ation from hMSCs into neuronal lineage. In the current study, we demonstrated that intracellular Ca2⫹transient peaks at day 1 post-induction (Fig. 2A) and is required for neuronal differ-entiation from hMSCs (Fig. 2, B and C). We also showed that
the effect of intracellular Ca2⫹on neuronal differentiation from hMSCs, at least in part, is through IP3-mediated calcium signal-ing (Figs. 2D and 3C). Recently, a study reported that a ryano-dine-sensitive receptor (RyR) agonist, caffeine, stimulates Ca2⫹ response that increases throughout neuronal differentiation in embryonic P19 carcinoma stem cells (CSCs) and adult murine MSCs (43). Intracellular Ca2⫹concentration has been demon-strated to be controlled by multiple mechanisms, including well characterized Ca2⫹ influx through voltage-gated channels, ligand-gated channels (44, 45), and non-voltage-gated channels (14), as well as Ca2⫹releasing from the endoplasmic reticulum (ER) via intracellular RyR and IP3R (15). Here, we uncovered for the first time that the intracellular Ca2⫹contents could be reg-ulated by a PcG protein, EZH2, through modulating the gene expression of PIP5K1C in addition to above mentioned calcium channels (Figs. 3 and 4). The expression profile of PKP5K1C (Fig. 5, A and B) is positively correlated to that of intracellular Ca2⫹contents (Fig. 2A) during induction of neuronal differen-tiation from hMSCs.
PcG proteins epigenetically repress transcription and their target genes have been genome-wide-mapped in murine ESCs (46) and human embryonic lung fibroblast TIG3 cell line (47), revealing that numerous development regulators and differen-tiation-related genes are repressed by binding of PcG proteins. Cells deficient in PRC2 component, EED, de-repress PcG target genes and activate neuronal differentiation in murine ESCs (46). Inactivation of PcG protein by knock-out of EED or EZH2 FIGURE 5. Neuronal differentiation from hMSCs requires PIP5K1C. A and B, 3A6 hMSCs were induced to differentiation into neuronal lineage. The expres-sion of PIP5K1C at each time interval was determined by quantitative RT-PCR (A) and Western blotting (B). C and D, cells were infected with or without lentivirus containing shRNAs against luciferase or PIP5K1C. The effect of these shRNAs on PIP5K1C expression was examined by detecting its mRNA (C) and protein (D) level. E, shRNA containing cells were induced to neuronal differentiation. The expressions of indicated neuron markers were determined at day 0 and day 5 after induction.-Actin was used as an internal control. F, intracellular Ca2⫹contents in hMSCs with PIP5K1C knockdown during induction of neuronal differenti-ation at indicated time interval were measured by flow cytometry.
at China Medical University, on September 22, 2011
www.jbc.org
promotes neurogenesis of neural precursor cells (NPCs, or neu-ral stem cells, NSCs), (48). Our study showed that EZH2 targets the promoter of PIP5K1C gene in proliferating hMSCs, and the binding could not be detected in neuron-differentiated hMSCs. Knockdown of EZH2 enhances PIP5K1C expression, PI(4,5)P2 generation, elevates intracellular Ca2⫹ contents (Fig. 4), and promotes hMSCs differentiating into functional neuron lineage (Fig. 6, A–C). Furthermore, knockdown of EZH2 in hMSCs also enhanced neuronal differentiation in the rat brain (Fig. 6, D and E). Although the mRNA expression of EZH2 was almost no different during induction to neuronal differenti-ation in hMSCs (Fig. 6A), the protein level of EZH2 was decreased after day 1 (supplemental Fig. S1). This is likely the cause of increased amount of PIP5K1C mRNA after day 1 post-induction to neuronal differentiation. Likewise, de-creased level of EZH2 has been reported in ESCs differenti-ation (49). In addition, a previous report demonstrated that EZH2 protein is highly expressed in proliferating NSCs from embryonic mice and decreased after NSCs differentiating into neurons and astrocytes (35).
It has been well characterized that PIP5K1s catalyze the pro-duction of PI(4,5)P2 in membrane lipid metabolism (50). In terms of their cellular functions, PIP5K1s are also involved in the regulation of actin reorganization and focal adhesion
dynamics, which are critical in cell migration (51) and neurite outgrowth (52, 53). Among all PIP5K1s, PIP5K1C is mainly expressed in brain and required for cardiovascular and neuro-nal development. The PIP5K1C-null embryos result in exten-sive prenatal lethality (23). A common regulatory mechanism of PIP5K1s is phosphorylation at specific site to alter the asso-ciation with binding partners. For instance, phosphorylation at Tyr-649 of human PIP5K1C by Src increases the affinity of its C-terminal for talin, and phosphorylation at Ser-650 by cyclin B1/Cdk1 blocks this interaction in focal adhesion (54, 55). Here, we provide evidence to establish that PIP5K1C is transcription-ally regulated by EZH2 (Fig. 4, A–C), and knockdown of PIP5K1C disrupts neuronal differentiation from hMSCs (Fig. 5E) and EZH2-silenced hMSCs (Fig. 6, A–C), indicating that PIP5K1C is essential for neuronal differentiation from hMSCs, and silencing EZH2 enhanced neuronal differentiation might be mediated via activation of PIP5K1C.
In summary, the current study demonstrates a novel regula-tory mechanism of intracellular Ca2⫹ signaling by EZH2 in hMSCs. A proposed model is shown in Fig. 7. After induction of neuronal differentiation, disassembly of EZH2 protein from the promoter of PIP5K1C gene increases the expression of PIP5K1C and generation of PI(4,5)P2, leading to elevated intra-cellular calcium signaling and advance neuronal differentiation FIGURE 6. Knockdown of EZH2 enhances neuronal differentiation from hMSCs in vitro and in vivo. A, mRNA expression profiles of neuron markers (NSE, PITX3, and NURR1), EZH2, PIP5K1C, as well as-actin, were determined by RT-PCR at indicated time intervals post-induction to neuronal differentiation in parental and shRNA-containing hMSCs. The bottom plot shows the relative intracellular Ca2⫹contents at indicated time point during neuronal differentiation of each hMSC transfectant. The original flow cytometry profiles were shown insupplemental Fig. S2. B, neuron-like function in each hMSC transfectant after induction of differentiation for 5 days was determined by change of Ca2⫹influx after stimulation of 100Mglutamate. The plot shows the kinetic profile of glutamate stimulated [Ca2⫹]i. C, difference of fluorescence intensity between the basal (before stimulation) and maximum (during stimulation) values (mean⫾
S.D.) from plot B. The symbols, * and ** indicate p value⬍0.05 and ⬍ 0.01, respectively by t test. D, representative three-dimensional image of bisbenzimide-labeled hMSCs (blue fluorescence) implantation in rat brain, significant down-regulated expression of EZH2 (red fluorescence) was found in EZH2 knockdown hMSCs-treated rats compared with that of control mock hMSCs-treated rats. The white arrows indicate the implanted MAP2 (green fluorescence)-positive hMSCs in rat brains. Scale bar: 50m. E, quantitative analysis of the implanted MAP2-positive cells numbers in both the EZH2 knockdown hMSCs-treated rats and control mock hMSCs-treated rats.
at China Medical University, on September 22, 2011
www.jbc.org
from hMSCs. To the best of our knowledge, this is the first report that demonstrates intracellular Ca2⫹signaling could be modulated by a PcG protein, EZH2, through transcriptional regulation of PIP5K1C. Our study provides a new insight to the role of EZH2 in neuronal differentiation from hMSCs. Activa-tion of intracellular Ca2⫹ signaling, which is suppressed by EZH2, might be a potential strategy to promote neuronal dif-ferentiation for an application to cure neurodegenerative dis-eases or spinal cord injury.
Acknowledgment—We thank the National RNAi Core Facility (Aca-demia Sinica, Taipei, Taiwan) for providing the shRNAs.
REFERENCES
1. Hung, S. C., Chen, N. J., Hsieh, S. L., Li, H., Ma, H. L., and Lo, W. H. (2002)
Stem Cells 20,249 –258
2. Bernardo, M. E., Zaffaroni, N., Novara, F., Cometa, A. M., Avanzini, M. A., Moretta, A., Montagna, D., Maccario, R., Villa, R., Daidone, M. G., Zuf-fardi, O., and Locatelli, F. (2007) Cancer Res. 67, 9142–9149
3. Hung, S. C., Cheng, H., Pan, C. Y., Tsai, M. J., Kao, L. S., and Ma, H. L. (2002) Stem Cells 20, 522–529
4. Hung, S. C., Lu, C. Y., Shyue, S. K., Liu, H. C., and Ho, L. L. (2004) Stem
Cells 22,1321–1329
5. Hung, S. C., Yang, D. M., Chang, C. F., Lin, R. J., Wang, J. S., Low-Tone Ho, L., and Yang, W. K. (2004) Int. J. Cancer 110, 313–319
6. Zhao, L. R., Duan, W. M., Reyes, M., Keene, C. D., Verfaillie, C. M., and Low, W. C. (2002) Exp. Neurol. 174, 11–20
7. Dezawa, M., Kanno, H., Hoshino, M., Cho, H., Matsumoto, N., Itokazu, Y.,
Tajima, N., Yamada, H., Sawada, H., Ishikawa, H., Mimura, T., Kitada, M., Suzuki, Y., and Ide, C. (2004) J. Clin. Invest. 113, 1701–1710
8. Lee, J. K., Jin, H. K., Endo, S., Schuchman, E. H., Carter, J. E., and Bae, J. S. (2010) Stem Cells 28, 329 –343
9. Cho, S. R., Kim, Y. R., Kang, H. S., Yim, S. H., Park, C. I., Min, Y. H., Lee, B. H., Shin, J. C., and Lim, J. B. (2009) Cell Transplant 18, 1359 –1368 10. Gu, W., Zhang, F., Xue, Q., Ma, Z., Lu, P., and Yu, B. (2010)
Neuropathol-ogy 30,205–217
11. Jin, H. K., Carter, J. E., Huntley, G. W., and Schuchman, E. H. (2002) J. Clin.
Invest. 109,1183–1191
12. Bae, J. S., Han, H. S., Youn, D. H., Carter, J. E., Modo, M., Schuchman, E. H., and Jin, H. K. (2007) Stem Cells 25, 1307–1316
13. Spitzer, N. C., Lautermilch, N. J., Smith, R. D., and Gomez, T. M. (2000)
Bioessays 22,811– 817
14. Wu, X., Zagranichnaya, T. K., Gurda, G. T., Eves, E. M., and Villereal, M. L. (2004) J. Biol. Chem. 279, 43392– 43402
15. Banerjee, S., and Hasan, G. (2005) Bioessays 27, 1035–1047 16. van den Bout, I., and Divecha, N. (2009) J. Cell Sci. 122, 3837–3850 17. Kanaho, Y., Kobayashi-Nakano, A., and Yokozeki, T. (2007) Biol. Pharm.
Bull. 30,1605–1609
18. Ishihara, H., Shibasaki, Y., Kizuki, N., Katagiri, H., Yazaki, Y., Asano, T., and Oka, Y. (1996) J. Biol. Chem. 271, 23611–23614
19. Loijens, J. C., and Anderson, R. A. (1996) J. Biol. Chem. 271, 32937–32943 20. Ishihara, H., Shibasaki, Y., Kizuki, N., Wada, T., Yazaki, Y., Asano, T., and
Oka, Y. (1998) J. Biol. Chem. 273, 8741– 8748
21. Giudici, M. L., Emson, P. C., and Irvine, R. F. (2004) Biochem. J. 379, 489 – 496
22. Wang, Y. J., Li, W. H., Wang, J., Xu, K., Dong, P., Luo, X., and Yin, H. L. (2004) J. Cell Biol. 167, 1005–1010
23. Wang, Y., Lian, L., Golden, J. A., Morrisey, E. E., and Abrams, C. S. (2007)
Proc. Natl. Acad. Sci. U.S.A. 104,11748 –11753
24. Schwartz, Y. B., and Pirrotta, V. (2008) Curr. Opin. Cell Biol. 20, 266 –273 25. Sparmann, A., and van Lohuizen, M. (2006) Nat. Rev. Cancer 6, 846 – 856 26. Pasini, D., Bracken, A. P., Jensen, M. R., Lazzerini Denchi, E., and Helin, K.
(2004) EMBO J. 23, 4061– 4071
27. Cao, R., Wang, L., Wang, H., Xia, L., Erdjument-Bromage, H., Tempst, P., Jones, R. S., and Zhang, Y. (2002) Science 298, 1039 –1043
28. Vire´, E., Brenner, C., Deplus, R., Blanchon, L., Fraga, M., Didelot, C., Mo-rey, L., Van Eynde, A., Bernard, D., Vanderwinden, J. M., Bollen, M., Es-teller, M., Di Croce, L., de Launoit, Y., and Fuks, F. (2006) Nature 439, 871– 874
29. Rajasekhar, V. K., and Begemann, M. (2007) Stem Cells 25, 2498 –2510 30. O’Carroll, D., Erhardt, S., Pagani, M., Barton, S. C., Surani, M. A., and
Jenuwein, T. (2001) Mol. Cell. Biol. 21, 4330 – 4336
31. Caretti, G., Di Padova, M., Micales, B., Lyons, G. E., and Sartorelli, V. (2004) Genes Dev. 18, 2627–2638
32. Juan, A. H., Kumar, R. M., Marx, J. G., Young, R. A., and Sartorelli, V. (2009) Mol. Cell 36, 61–74
33. Aoki, R., Chiba, T., Miyagi, S., Negishi, M., Konuma, T., Taniguchi, H., Ogawa, M., Yokosuka, O., and Iwama, A. (2010) J. Hepatol. 52, 854 – 863
34. Ezhkova, E., Pasolli, H. A., Parker, J. S., Stokes, N., Su, I. H., Hannon, G., Tarakhovsky, A., and Fuchs, E. (2009) Cell 136, 1122–1135
35. Sher, F., Ro¨ssler, R., Brouwer, N., Balasubramaniyan, V., Boddeke, E., and Copray, S. (2008) Stem Cells 26, 2875–2883
36. Tsai, C. C., Chen, C. L., Liu, H. C., Lee, Y. T., Wang, H. W., Hou, L. T., and Hung, S. C. (2010) J. Biomed. Sci. 17, 64
37. Chu, M. S., Chang, C. F., Yang, C. C., Bau, Y. C., Ho, L. L., and Hung, S. C. (2006) Cell Signal. 18, 519 –530
38. Tondreau, T., Dejeneffe, M., Meuleman, N., Stamatopoulos, B., Del-forge, A., Martiat, P., Bron, D., and Lagneaux, L. (2008) BMC Genomics
9,166
39. Gee, K. R., Brown, K. A., Chen, W. N., Bishop-Stewart, J., Gray, D., and Johnson, I. (2000) Cell Calcium. 27, 97–106
40. Gray, A., Olsson, H., Batty, I. H., Priganica, L., and Peter Downes, C. (2003)
Anal. Biochem. 313,234 –245
41. Shyu, W. C., Liu, D. D., Lin, S. Z., Li, W. W., Su, C. Y., Chang, Y. C., Wang, H. J., Wang, H. W., Tsai, C. H., and Li, H. (2008) J. Clin. Invest. 118, FIGURE 7. A proposed model of EZH2-mediated PIP5K1C-dependent
neu-ronal differentiation from hMSCs. In proliferating undifferentiated hMSCs,
EZH2 protein binds to the promoter of PIP5K1C gene to repress its transcrip-tion to maintain the homeostasis of intracellular Ca2⫹contents in relative low level. After induction to neuronal differentiation, EZH2 protein is dissociated from the promoter of PIP5K1C to enhance its gene expression and results in increase of PI(4,5)P2generation, leading to activation of IP3-mediated Ca2⫹ signaling to promote neuronal differentiation from hMSCs.
at China Medical University, on September 22, 2011
www.jbc.org
2482–2495
42. Holliday, J., and Spitzer, N. C. (1990) Dev. Biol. 141, 13–23
43. Resende, R. R., da Costa, J. L., Kihara, A. H., Adhikari, A., and Lorenc¸on, E. (2010) Stem Cells Dev. 19, 379 –394
44. Tsien, R. W., Lipscombe, D., Madison, D., Bley, K., and Fox, A. (1995)
Trends Neurosci. 18,52–54
45. Jackson, M. B. (1999) Adv. Neurol. 79, 511–524
46. Boyer, L. A., Plath, K., Zeitlinger, J., Brambrink, T., Medeiros, L. A., Lee, T. I., Levine, S. S., Wernig, M., Tajonar, A., Ray, M. K., Bell, G. W., Otte, A. P., Vidal, M., Gifford, D. K., Young, R. A., and Jaenisch, R. (2006) Nature
441,349 –353
47. Bracken, A. P., Dietrich, N., Pasini, D., Hansen, K. H., and Helin, K. (2006)
Genes Dev. 20,1123–1136
48. Hirabayashi, Y., Suzki, N., Tsuboi, M., Endo, T. A., Toyoda, T., Shinga, J., Koseki, H., Vidal, M., and Gotoh, Y. (2009) Neuron 63, 600 – 613
49. de la Cruz, C. C., Fang, J., Plath, K., Worringer, K. A., Nusinow, D. A., Zhang, Y., and Panning, B. (2005) Chromosoma 114, 183–192
50. Heck, J. N., Mellman, D. L., Ling, K., Sun, Y., Wagoner, M. P., Schill, N. J., and Anderson, R. A. (2007) Crit. Rev. Biochem. Mol. Biol. 42, 15–39 51. Ling, K., Schill, N. J., Wagoner, M. P., Sun, Y., and Anderson, R. A. (2006)
Trends Cell Biol. 16,276 –284
52. van Horck, F. P., Lavazais, E., Eickholt, B. J., Moolenaar, W. H., and Dive-cha, N. (2002) Curr. Biol. 12, 241–245
53. Yamazaki, M., Miyazaki, H., Watanabe, H., Sasaki, T., Maehama, T., Fro-hman, M. A., and Kanaho, Y. (2002) J. Biol. Chem. 277, 17226 –17230 54. Ling, K., Doughman, R. L., Iyer, V. V., Firestone, A. J., Bairstow, S. F.,
Mosher, D. F., Schaller, M. D., and Anderson, R. A. (2003) J. Cell Biol. 163, 1339 –1349
55. Lee, S. Y., Voronov, S., Letinic, K., Nairn, A. C., Di Paolo, G., and De Camilli, P. (2005) J. Cell Biol. 168, 789 –799
at China Medical University, on September 22, 2011
www.jbc.org