國
國
國
國
立
立
立
立
交
交
交
交
通
通
通
通
大
大
大
大
學
學
學
學
生化工程研究所
生化工程研究所
生化工程研究所
生化工程研究所
碩
碩
碩
碩
士
士
士
士
論
論
論
論
文
文
文
文
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
性研究
性研究
性研究
性研究
Development and characterization of a
lipo-poly-complex in the induction of specific
immune responses
研 究 生:陳家弘
指導教授:廖光文博士
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
性研究
性研究
性研究
性研究
Development and characterization of a
lipo-poly-complex in the induction of specific immune
responses
研 究 生:陳家弘 Student:Chia-Hung Chen 指導教授:廖光文 Advisor:Kuang-Wen Liao 國 立 交 通 大 學 生化工程研究所 碩 士 論 文 A ThesisSubmitted to Institute of Biochemical engineering College of Biological Science and Technology
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Master
in
Biochemical Engineering
June 2009
Hsinchu, Taiwan, Republic of China
i
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
開發具有誘發特異性免疫反應之微脂體複合體與其特
性研究
性研究
性研究
性研究
研究生:陳家弘 指導教授: 廖光文 博士
國立交通大學生化工程研究所
中文摘要
中文摘要
中文摘要
中文摘要
抗原呈現細胞 (antigen presenting cell; APC) 利用 MHC 分子將病源體之抗 原和 T 細胞細胞膜表面的 TCR 交互作用後,並配合共同刺激因子,可有效地開 啟專一性的 T 細胞免疫反應。目前了解樹突細胞 (dendritic cell; DC) 是抗原呈現 細胞中,呈現抗原之能力最具有效率的。根據一些相關樹突細胞的研究,利用樹 突細胞能有效吞噬抗原、呈現抗原,及活化 T 細胞的能力,研究人員藉此已將 其發展成為一個很好的疫苗平台。但是,在其他研究中也發現到樹突細胞會受到 病源體相關因子影響,降低了樹突細胞之免疫功能。於是便發展人工化抗原呈現 細胞來解決樹突細胞所遭遇到問題。本篇論文研究成功建立了一個具有免疫調節 功能的新穎之微脂體。LPPC (Lipo-PEI-PEG Complex) 是本實驗室開發出來的微 脂體,因為它具有穩定吸附蛋白的能力,並且能維持該蛋白的活性。因此,利用 LPPC 新穎的特性,將具有可以調節免疫反應的單株抗體或 MHC 分子給予 LPPC 吸附後,賦予 LPPC 擁有抗原呈現細胞的能力。 研究結果顯示,LPPC 吸附 anti-CD3 和 anti-CD28 單株抗體後,仍保有原來 活性可刺激老鼠脾臟細胞和人類周邊單核球在細胞增生及細胞激素 (cytokine) 有較高表現。此外 LPPC 本身可以促進抗原呈現細胞對抗原之吞噬反應,也可以 刺激免疫細胞在前發炎反應激素表現及呈現抗原能力。因此 LPPC 在免疫功能上 具有佐劑一樣效應,可增強免疫細胞對抗原的反應。而當 LPPC 吸附來自於樹突 細胞富含許多 MHC 分子和共同刺激分子的膜蛋白時,一樣也保有該蛋白分子的 活性可誘發動物體內專一性的免疫反應。此外 LPPC 吸附帶有特定抗原的人類 MHC I (HLA-A2) 分子和 anti-CD28 單株抗體後,蛋白分子在 LPPC 平台上仍然 可以誘發動物體內專一性免疫反應。因此 LPPC 在本研究中,可以靈活變動地賦 予免疫功能之優勢去引發專一性的免疫反應,LPPC 未來將可以發展成一個很好 的免疫調控平台。
ii
Development and characterization of a
lipo-poly-complex in the induction of specific immune
responses
Student: Chia-Hung Chen Advisor: Dr. Kuang-Wen Liao Institute of Biochemical Engineering
National Chiao Tung University
ABSTRACT
Antigen presenting cells (APCs) can efficiently elicit specific T cell immune responses by presenting pathogen-derived peptides on the major histocompatibility complexs (MHCs) to T cell receptor (TCR) on the surface of T cells. Currently, dendritic cells (DCs) have been discovered as the most efficient APCs. Based on these findings, DCs have been developed as a good bio-reagent to activate host’s adaptive immunity by up-taking antigen, presenting antigen and stimulating T-cells. However, DCs would be inactivated in vivo by certain pathogen-derived antigens. Thus, artificial antigen presenting cells were developed to resolve this problem. In this study, a novel immuno-regulatory liposome was developed. Lipo-PEI-PEG-complex (LPPC) was a novel liposome could strongly absorb proteins on its surface and the bound proteins could maintain their activities. By these characters, LPPC was designed to adsorb certain monoclonal antibodies or MHC molecules which have the abilities to regulate immune responses and the immuno-LPPCs were used as APC.
The results showed that the LPPC adsorbed anti-CD3 and anti-CD28 monoclonal antibodies could increase the proliferation and cytokine secretions (IL-2, IFN-γ and TNF-α) of mouse splenocytes and human peripheral blood mononuclear cells (PBMC). In addition, LPPC can promote the abilities of APCs to up take antigen, induce the proinflammatory cytokine expressions, and present antigen. Thus, LPPC could provide a good adjuvant effect. Besides, the LPPC coated with membrane proteins of DCs performed as APCs to stimulate specific T-cell immune responses. Moreover, antigen-loaded HLA-A2 molecules and anti-CD28 monoclonal antibody were adsorbed on LPPC and they also maintained their specific activities, and induced the specific immune responses. The immuno-LPPC displays its flexible character and advantage to regulate immunity by combining certain immuno-regulatory antibodies and specific-antigen MHC molecules. Therefore, the immuno-LPPC may be developed as a good immunoregulatory platform.
iii
Acknowledgement
燈塔的夜晚,苦悶孤寂,一個人在實驗室敲打著鍵盤,拼湊這兩年來 瑣碎片段的記憶,嗅著濃烈咖啡因的味道,喝下這兩年來辛苦沉重的 心情,回憶如走馬燈在腦海跑了好幾回,猶記得兩年多前,風塵僕僕 孤身一人帶著豪情壯志揮軍北上,直闖風城裡腫瘤免疫絕學之深奧領 域中,盼望能學得一招半式,可以為自己和清水家鄉揚名立萬,誰知 層出不窮的實驗瓶頸,接踵而來的課業煩惱。 幸好,當初遇到了自 詡小諸葛的廖光文老師,讓我見識到如何靈活運用實驗室的資源及多 方面審視和分析每個人實驗結果,這些『非常人』之撇步,另外廖老 師認真指導加上不厭其煩地傳授資質駑鈍的我,使我可以快速累積和 汲取實質經驗。 一天,霎時任督打通茅塞頓開,領悟到免疫絕學之 奧妙,終於在今年六月順利通過廖老師和師叔姐們的考驗,成為一個 文武雙全的交大碩士生。 當然這求學拜師過程中還是需要其他師兄 弟和姊妹們的幫忙,像彥谷學長和于鈴學姐提攜指教,公主們小莉小 溫啾咪吐嘲求進步,姵姵小貓咪細心照顧幫忙,馬馬靜敏筱葳維瞳切 磋砥礪等眾人幫忙合作下,成就了今天的我,在下心中滿懷感激之情。 此外還有已畢業的學長姐們和其他實驗室有緣人們指教,真是很謝謝 大家。 『懂得屈膝,跳得更高』 家弘與有緣人共勉之iv
Contents
中文摘要 中文摘要 中文摘要 中文摘要... i ABSTRACT ... ii Acknowledgement ... iii Contents ... iv Tables ... vii Figures ... viii Abbreviations ... ix Chapter 1 Introduction ...1Chapter 2 Materials and Methods...6
2.1MATERIALS ...6 2.1.1 Reagents ... 6 2.1.2 Cell lines ... 6 2.1.3 Antibody ... 6 2.1.4 Kits ... 6 2.1.5 Animals ... 7 2.1.6 Others ... 7 2.2METHODS ...7 2.2.1 LPPC preparation ... 7 2.2.2 Adsorption of LPPC ... 8 Timing ... 8 Capacity ... 8 Competition... 8
v
2.2.3 PBMC isolation ... 8
2.2.4 Splenocyte isolation ... 9
2.2.5 The cytotoxicity of LPPC toPBMC or splenocyte ... 10
2.2.6 The activities of monoclonal antibodies adsorbed on LPPC ... 10
2.2.7 The stability of immunostimulatory monoclonal antibodies adsorbed on LPPC in RPMI ... 11
2.2.8 The dose-effect of monoclonal antibodies adsorbed on LPPC in immune cells ... 11
Cell proliferation ... 11
Cytokines secretion ... 12
Pro-inflammatory cytokine profiles secretion ... 12
2.2.9 The comparison of activities of monoclonal antibodies on LPPC ... 12
2.2.10 The uptake protein ability of P338D1 ... 13
2.2.11 DC harvest ... 13
Purification of DC membrane protein ... 15
DC surface marker staining ... 15
2.2.12 The strategy of animal immunization ... 17
Bovine serum albumin (BSA) ... 17
Heat shock protein 60 of Helicobacter pylori ... 18
HPV E7 YML ... 19
2.2.13 The enhancement of antigen presentation of APCs by LPPC ... 20
Cell proliferation ... 20
Cytpkines secretion ... 20
2.2.14 Membrane proteins with specific antigen coated LPPC ... 20
Cell proliferation ... 20
Cytpkines secretion ... 21
2.2.15 The specific peptide-loaded HLA-A2 adsorbed on LPPC ... 21
vi
Cytpkines secretion ... 22
2.2.16 The animal immunization of the immuno-LPPC in vivo ... 22
2.2.17 The animal immunization efficiency of immuno-LPPC in vivo ... 22
Cell proliferation ... 23
Cytpkines secretion ... 23
2.2.18 Statistical analysis ... 23
Chapter 3 Result ...25
3.1THE CHARACTERS OF LPPC ...25
3.2THE CHARACTERS OF LPPC FOR PROTEIN ADSORPTION ...25
3.3THE ACTIVITIES OF IMMUNOSTIMULATORY MONOCLONAL ANTIBODIES ADSORBED ON LPPC ...26
3.3.1 The cytotoxicity of LPPC to PBMCs or splenocytes ... 26
3.3.2 The effects of the bound antibodies in a dosage-dependent manner ... 26
3.4THE STABILITY OF IMMUNOSTIMULATORY MONOCLONAL ANTIBODIES ADSORBED ON LPPC IN RPMI ...28
3.5THE ENHANCEMENT OF UPTAKE PROTEIN ABILITY BY LPPC ...28
3.6THE SPECIFICITIES OF THE LPPC-BOUND PROTEINS ...29
3.7THE EFFICIENCY OF IMMUNIZATION IN VIVO ...30
Chapter 4 Discussion ...31
vii
Tables
TABLE 1. THE VARIOUS PROTEINS WERE ADSORBED BY LPPC. . ...41
viii
Figures
FIGURE 1. THE CHARACTERS OF LPPC. ...37
FIGURE 2. THE PROPERTIES OF LPPC ADSORPTION. ...39
FIGURE 3. THE CYTOTOXICITY OF LPPC FOR PBMC AND SPLENOCYTE. ...42
FIGURE 4. THE DOSE-DEPENDENT EFFECT OF CELL PROLIFERATIONS WAS
STIMULATED BY IMMUNOSTIMULATORY MONOCLONAL ANTIBODIES ON LPPC. ...43 FIGURE 5. THE DOSE-DEPENDENT FASHION OF CYTOKINE PROFILES SECRETION WAS
ACTIVATED BY MONOCLONAL ANTIBODIES ON LPPC. ...45 FIGURE 6. THE INDUCTION OF PRO-INFLAMMATORY CYTOKINE PROFILES WAS BY
THE LPPC. ...48 FIGURE 7. THE ACTIVITIES OF IMMUNOSTIMULATORY MONOCLONAL ANTIBODIES ON
LPPC. ...50
FIGURE 8. THE STABILITY OF IMMUNOSTIMULATORY MONOCLONAL ANTIBODIES ON
LPPC IN RPMI. ...53
FIGURE 9. THE INCREASE OF CELLULAR UPTAKE OF P338D1 WAS BY THE LPPC. ...55
FIGURE 10. THE ENHANCEMENT OF THE ANTIGEN PRESENTATION OF APC WAS BY
THE LPPC. ...57 FIGURE 11. THE SURFACE MARKER EXPRESSIONS OF DENDRITIC CELLS. ...59
FIGURE 12. THE SPECIFICTIES OF DCS’ MEMBRANE PROTEINS COATED LPPC. ...60
FIGURE 13. THE SPECIFICITIES OF ANTIGEN-LOADED HLA-A2COMBINED WITH
ANTI-CD28 MONOCLONAL ANTIBODY ON LPPC. ...62 FIGURE 14. THE ANIMAL IMMUNIZATION OF THE BOUND MEMBRANE PROTEINS
DERIVED FROM DCS ON LPPC. ...63 FIGURE 15. THE ANIMAL IMMUNIZATION OF THE LPPC COMBINED HLA-A2
ix
Abbreviations
APCs antigen-presenting cells
CTLs Cytotoxic T lymphocytes
DCs dendritic cells
LFA leukocyte function-associated antigen
ICAM intercellular adhesion molecule
IFN-γ interferon-γ
CEA carcinoembryonic antigen
rAAV recombinant adeno-associated virus
VEGF Vascular endothelial growth factor
HPV human papillomavirus
HIV human immunodeficiency virus
SIV simian immunodeficiency virus
IL-2 interleukin-2
IL-1β interleukin-1 β
IL-6 interleukin-6
IL-8 interleukin-8
IL-10 interleukin-10
TNF-α Tumor necrosis factor-α
MHC major histocompatibility complex
PBS phosphate buffer saline
mAb Monoclonal antibody
LPPC Lipo-PEI-PEG-Complex
x
PEG polyethylene glycols
TCR T cell receptor
PBMC peripheral blood mononuclear cell
DLS dynamic light scattering
HP Helicobacter pylori
CTLA-4 Cytotoxic T lymphocyte-associated
antigen 4
HSP Heat shock protein
LPS lipopolysaccharide
TEM Transmission electron microscopy
BSA Bovine serum albumin
HLA-A2 Human leukocyte antigen-A2
MART-1 melanoma-associated antigen
recognized by T cell-1
hTERT human telomerase reverse
1
Chapter 1 Introduction
A major aim in immunotherapy is to generate specific cell-mediate immune responses to regulate host immunity. Efficiently resist pathogens and virus that relies on T cell immune responses (1). The optimal T-cell stimulation requires engagement of the T-cell receptor (TCR) through the major histocompatibility complex (MHC) bound to peptide, together with at least one interaction of a costimulatory molecule with an appropriate ligand on the T cell (2, 3).
The most potent and best-investigated costimulatory molecules are B7-1 and B7-2, which bind to CD28 (4, 5) on the T cell and induce cell proliferations. In contrast, B7 molecules might also bind cytotoxic T lymphocyte antigen 4 (CTLA-4) molecules on activated T cells and induce apoptosis in those T cells. Additional interactions might also regulate T-cell stimulation, including T-cell subtype differentiation, induction of maximal proliferation and prevention of apoptosis. Potential molecules of these interactions are other costimulatory molecules such as ICOS, 4-1BBL and OX40 (6-8).
Another major group of molecules are the adhesion molecules, which include leukocyte function-associated antigen (LFA) and intercellular adhesion molecule (ICAM) (9, 10). These molecules increase the interaction time between the T cell and APC, and enhance efficient activation (11). As discuss above, over the past decade many
2
new costimulatory molecules have been identified, offering new insights into T-cell activation and regulation.
Naive T cells are activated to produce armed efficient T cells the first time they encounter their specific antigen in the form of a peptide:MHC complex on the surface of an activated antigen presenting cell (APC). Antigen presentation by APCs, most notably macrophages and dendritic cells (DCs), and infected B cells is critical for induction of specific T cells in the form of an adaptive immune response (12). Further, the induction of T cell-mediated immunity is controlled by antigen-presenting DCs, potent stimulators of specific T cell immunity (13). DCs in essence act as nature’s adjuvants and play an important role to generate adaptive immunity. They present immunogenic epitopes of antigens in the context of MHC class I and class II molecules in association with costimulatory molecules, and efficiently activate both cytotoxic T cells and T helper cells (14).
DCs are both efficient and specialized in antigen presentation, and they control the magnitude, quality, and memory of the ensuing immune response. Because of the exceptional ability of DCs to activate T-cell immunity in response to microbial pathogens and tumor cells, these cells have been exploited as ex vivo and in vivo tools for immunotherapy. For example, Dr. Lu demonstrate that a therapeutic vaccine made of inactivated SIV-pulsed DCs can elicit effective cellular and humoral immune responses against SIV, allowing the control of SIV replication in the secondary lymphoid tissues and the reduction of cell-associated viral
3
DNA and cell-free viral RNA in blood of SIV-infected macaques (15). In addition, HSP105-pulsed BM-DC vaccine could induce specific T cells to inhibit the growth of intestinal tumors overexpressing HSP105 (16). Moreover, Dr. Aldrich utilized rAAV with human tumor antigen, carcinoembryonic antigen (CEA), of gene to infect DC for induction of specific immunity (17). In another study, the coadministration of DNA vaccines encoding HPV16 E7 with siRNA targeting key proapoptotic proteins successfully prolongs the lives of DCs, enhances antigen specific CD8+ T-cell responses, and elicits potent antitumor effects against an E7-expressing tumor model (18).
As development of DC-based application, the challenges of these therapies need to be improved. Several virus- and nonvirus-based transduction methods have been used for DC-based therapy. However, all strategies result in different levels of gene expression depending on the transduction efficiency. Therefore, purification of expressing APCs is needed to avoid non-expect interaction (11). In addition, some articles indicated that pathogens or pathogens-derived factors, such as Candida albicans, Mycobacterium tuberculosis, mycobacterial LAM, and secretions of Candida respectively impact on efficiency of DCs and immune cells, affect cytokine expression and impair surface marker of DC (19-22). Moreover, mature DCs express high levels of costimulatory molecules such as B7.1 and B7.2, which could be contact to CTLA-4, an immuno-inhibitory ligand that suppresses T cell activation (23, 24). In addition, many tumors secrete immunosuppressive factors such as TGF-β, IL-10, or VEGF, which affect the function of DCs to influence
4
the efficiency of DC vaccine (11, 25-27).
As an alternative strategy, artificial antigen-presenting cell systems (aAPC) have been recently developed and are rapidly expanding. They encompass both cellular-based and acellular-based technologies (28). In cellular-based, they utilized genetic methods that transduced into murine fibroblast cells to express immunoregulatory molecules, such as antigen-loaded MHC molecules and B7 molecules (29). In addition, a nonspecific cell-based aAPC (K32 cells) has also been developed, that were transfected with the costimulatory molecule 4-1BBL and the low-affinity Fc gamma receptor CD32 (30).
In acellular-based, they employed chemical method to conjugate immunoregulatory molecules on nanoparticles, bead or liposome. For example, a acellular aAPC was developed that can be used to induce and expand clinically relevant amounts of highly enriched peptide-specific T cells based on HLA-A2–Ig molecules and anti-CD28 monoclonal antibody (mAb) coupled to a magnetic bead (31, 32). In this strategy, peptide resident in the HLA–Ig molecule with any HLA-A2-restricted antigenic peptide can be modified. Thus, a single batch of HLA-A2–Ig-based aAPCs can be loaded with various different antigenic peptides for expansion of cells with different antigenic specificities. These artificial APC-based strategies certainly diminished the effect of immunosuppression.
5
complex (LPPC), that was a biodegradable liposome with the characters that could strongly and rapidly adsorb proteins on its surface, and these proteins could maintain their activities. The purpose of this study, that manufacture artificial antigen presenting cells, or APC-liked liposome, which were combined the liposome with immunostimulatory molecules to develop as an immunoregulatory platform. Therefore, we exploited LPPC combined with immunostimulatory molecules as artificial antigen presenting cells to activate immunity. Here, we added anti-CD3 and anti-CD28 monoclonal antibodies (mAbs), or addition of DCs’ membrane proteins, or addition of specific peptide-HLA-A2 complex for the LPPC adsorption. The results showed that LPPC indeed exhibited ability of enhancement of the cell proliferations and cytokine secretions of human peripheral blood mononuclear cells (PBMCs) and murine splenocytes in vitro. We also showed that the LPPC with immuno-molecules induced specific immune responses in vivo. Moreover, LPPC might have the potential of an adjuvant that enhance immune responses of APCs. In this study, we indeed demonstrated that LPPC showed quickness and good flexibility to construct an immunoregulatory platform as an artificial antigen presenting cell.
6
Chapter 2 Material and method
2.1 Material
2.1.1 Reagent
The following reagents and chemicals were obtained as indicated: RPMI 1640, Fetal Bovine Serum (FBS), and BSA from Invitrogen. Penicillin/ streptomycin/ amphotericin (PSA) from Biological industries.
NaCl, Tris-HCl, Triton X-100, from Amresco. Ficoll-PaqueTM Plus from
GE healthcare. Propidium iodide (PI) from CE. EDTA and chloroform from TEDIA. NaOH, H3PO4, KH2PO4, Na2HPO4, tween 20, KHCO3,
NaN3, and KAc from SHOWA. KCl from Scharlau. Na2HPO4 from J. T.
Baker. Urea from USP. 2.1.2 Cell lines
P338D1 (Mouse macrophage-like cell line; ATCC number: CCL-46.)
2.1.3 Antibody
The following antibodies were obtained as indicated: Mouse anti-human CD3 (OKT3) and Mouse anti-mouse CD3 (2C11) were kindly provided from Dr. Steve R. Roffler (ACADEMIA SINICA, Institute of BioMedical Sciences). Anti-human CD28 monoclonal antibody, and anti-mouse CD28 monoclonal antibody were purchased from Biolegent.
2.1.4 Kits
7
IL-8, IL-2, IFN- γ and TNF-α ELISA kit, and mouse IL-4, IL-10, IL-2, IFN- γ, and TNF-α ELISA kit from R&D. MTT assay kit from Promega. 2.1.5 Animals
Six-eight weeks old female BALB/C mice were purchased from the National Laboratory Animal Center, Taipei, Taiwan, R.O.C. Six-eight weeks old female C57BL/6-Tg (HLA-A2.1) mice were kindly provided from Dr. Shih-Jen Liu (National Health Research Institutes).
2.1.6 Others
Peptide-HLA-A2 monomer and one identify epitope of HPV type 16 E7 protein (YMLDLQPETT) were kindly from Dr. Shih-Jen Liu (National Health Research Institutes).
2.2 Method
2.2.1 LPPC preparation
Briefly, added each DOPC and DLPC 500µl (50 mg/ml) into the round bottom flask, and then added 1000µl methanol into the same container and mix well. The mixture was placed the container of lipid
mixture to the rotary evaporator (37℃, without vacuum treatment,
minimum rotary speed) until dry (about 2 days). Hydrated the lipid film
by steam (about 37℃) for 2~3 hours. Added 5ml aqueous medium
(containing 0.675g PEI and 0.22g PEG in 5 ml filtered DDW) gently to the container of dry lipid and agitating gently. The container was vortexed violently for 10 minutes. After vortexed, the LPPC was placed at RT overnight. The turbid medium of LPPC extruded through 200nm mesh
8
nine times. The product stored into the container to 4℃ refrigerator.
2.2.2 Adsorption characters of LPPC Timing
Added 50 µg BSA-FITC into 40 µg/ml LPPC solution, and then centrifuged at 10,000 rpm for 5 min at different time. The fluorescence of LPPC pellet was estimated by Spectrofluorometer.
Capacity
Added different amounts of BSA into 40 µg/ml LPPC solution, and then centrifuged at 10,000 rpm for 5 min at 20 minutes. The amounts of BSA onto LPPC were measured by using coomassie plus reagent.
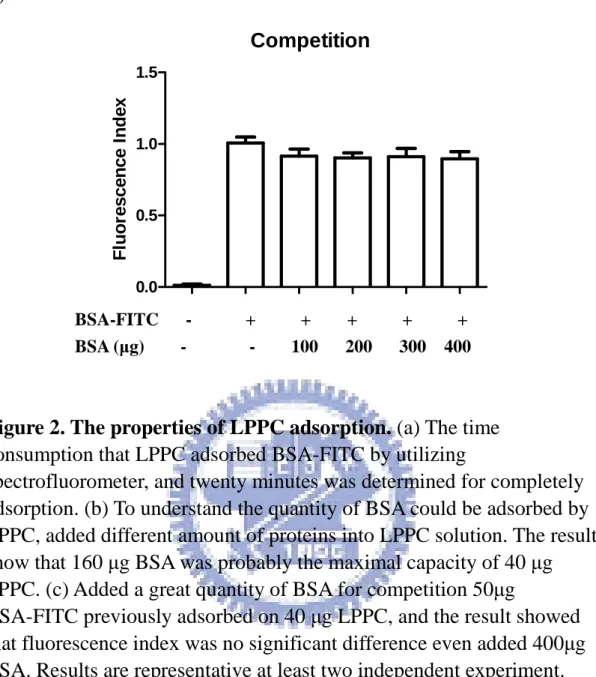
Competition
40 µg/ml LPPC prior to adsorb 50 µg BSA-FITC completely, and added different folds of BSA for competition in 20 minutes. And then centrifuged at 10,000 rpm for 5 min, the fluorescence of LPPC pellet was estimated by Spectrofluorometer. Positive control was the fluorescence of the LPPC solution without adding BSA. Negative control was the fluorescence of the LPPC alone solution.
2.2.3 PBMC isolation
Peripheral blood mononuclear cells (PBMCs) were separated from human white blood cell solution by using Ficoll-PaqueTM Plus. Dilute human white blood cells with equal volume of PBS. Add Ficoll-Paque PLUS (6 ml) to the 15 ml centrifuge tube and carefully layer the diluted blood sample (8 ml) on Ficoll-Paque PLUS. Centrifuge the tubes at 400g
9
for 40min at 18℃. Remove the plasma layer and collect the PBMC layer.
Wash the cells with 2 volume of PBS for centrifuging at 1500 rpm for 15 min. Discard the supernatant and lyse the red blood cells by ACK buffer
(0.15 M NH4Cl, 10 mM KHCO3, 0.1 mM EDTA in DDW) at room
temperature for 10min and followed by centrifuging at 1500 rpm for 15 min. Discard the supernatant and wash the cell with 10 ml PBS. Centrifuge for another 15 min. Discard the supernatant and count the cell number. For the cell proliferation of PBMC, cells were plated in a
96-well at 1 x 105 per well. For the cytokine profiles of PBMC, cells were
plated in a 24-well plate at 4 x 105 per well.
2.2.4 Splenocyte isolation
Mice were sacrificed by dislocation and their spleens were quickly harvested in a laminar flow hood. Spleens were placed in a 280 µm-pored mesh and chopped by scissors. 10 ml of RPMI 1640 (Invitrogen Co.,
USA) supplemented with 10% FBS, 0.2% NaHCO3 and 1% PSA. were
slowly added onto the mesh while spleens were being ground until the spleen tissue became white. Single cell suspension was collected in a
Petri dish and recovered by centrifugation at 1,200 rpm at 4℃ for 5 min.
Supernatant was discarded and 10 ml 1X ACK lysis buffer was added for 5 min at room temperature. 1X ACK buffer can lyse the red blood cells while leaving the rest of the lymphocytes and leucocytes. The mixture was then diluted by 10 ml of RPMI 1640 and cells were recovered by centrifugation at 1,200 rpm at 4℃ for 5 min. After the supernatant was discarded, the cells were rinsed by 10 ml PBS once more. Finally, cells were resuspended in RPMI 1640 and underwent cell calculation by trypan
10
blue exclusion. For the cell proliferation of splenocyte, cells were plated
in a 96-well at 2.5 x 105 per well. For the cytokine profiles of splenocyte,
cells were plated in a 24-well plate at 1 x 106 per well.
2.2.5 The cytotoxicity of LPPC to PBMCs or splenocytes
PBMC (1×105 cells per well) or splenocyte (2.5×105 cells per well) were respectively dispensed into 96-well culture plates and then except for control treated with different condition. After 72 hr, the cells were centrifuged at 400g for 15 min. Removed the medium, and added 100 µl MTT working solution per well. And then, the 96-well culture plates were put back incubator with 5% CO2 at 37℃ for 4 hr. The supernatant was removed, and added 100 µl DMSO to dissolve the purple crystal. Put plates on the shaker for 10 min. The optical density was determined by a microplate reader (Tecan) set to 595 nm and the data were analyzed by Magellan5 software.
2.2.6 The activities of monoclonal antibodies adsorbed on LPPC
In this study, anti-CD3 monoclonal antibody (2C11 or OKT3) was utilized as first signal for activation of T cell, and the other monoclonal antibody, anti-CD28 as second signal was for optimal activation of T cell.
PBMC (1×105 cells per well) or splenocyte (2.5×105 cells per well) were
respectively dispensed into 96-well culture plates and then except for control treated with different condition. 40 µg LPPC pre-adsorbed 100 µg BSA, and then adsorbed with 2.4 µg anti-CD3 mAb or with 2.4 µg anti-CD3 and 2.4 µg anti-CD28 mAb into 100 µl volume. After centrifuged, 2.5 µl LPPC complex treated PBMCs or splenocytes for
11
72hrs. By using MTT assay, and then the cell proliferation rate was calculated as O.D. value of sample divide into O.D. value of PBMC alone or splenocyte alone.
2.2.7 The stability of immunostimulatory monoclonal antibodies adsorbed on LPPC in RPMI
40 µg LPPC previously adsorbed 100 µg BSA, and then adsorbed with 2.4 µg anti-CD3 mAb or 2.4 µg anti-CD3 and 2.4 µg anti-CD28 mAb into 100 µl volume. After centrifuged, put the LPPC complex into
RPMI solution in 37 ℃ for 30 minutes. After 30 minutes, the solution was
centrifuged divide into LPPC pellet and the supernatant. The LPPC pellet was resuspend into 100 ul DDW. The 2.5 µl LPPC complex and the 2.5 µl
supernatant respectively treated PBMCs (1×105 cells per well) or
splenocytes (2.5×105 cells per well) in 96-well culture plate, and
estimated the cell proliferation of immune cells for investigating the efficiency of monoclonal antibodies on LPPC. By using MTT assay, and then the stimulation index was calculated as (O.D. value of sample –O.D. value of PBMC alone or splenocytes alone) / O.D. value of PBMC alone or splenocytes alone.
2.2.8 The dose-effect of monoclonal antibodies adsorbed on LPPC in immune cells
Cell proliferation
PBMC (1×105 cells per well) or splenocyte (2.5×105 cells per well) were respectively dispensed into 96-well culture plates and then except for control treated with different condition. Addition different amounts of
12
immunostimulatory monoclonal antibodies were combined with 1 µg LPPC to stimulate the proliferation of immune cells, and the cell numbers was counted by MTT assay at 72 hrs. The cell proliferation rate was calculated as O.D. value of sample divide into O.D. value of PBMC alone or splenocyte alone.
Cytokines secretion
PBMC (4×105 cells per well) or splenocyte (106 cells per well) were dispensed into 24-well culture plates and then except for control treated with different condition. 4 µg LPPC adsorbed different amounts of monoclonal antibodies respectively to treat PBMCs or splenocytes. And the supernatants were collected at 24h and 72 h and frozen at −80 ℃.
Supernatants concentrations of TNF-α, IL-2, and IFN-γ were measured
by Enzyme-Linked ImmunoSorbent Assay (ELISA). Pro-inflammatory cytokine profiles secretion
PBMC (4×105 cells per well) was dispensed into 24-well culture plates
and then except for control treated with different condition. 4 µg LPPC treated PBMCs and then the supernatants were collected at 24 h, 48 h and
72 h and frozen at −80℃. Supernatants concentrations of IL-1β, IL-6, and
IL-8 were measured by Enzyme-Linked ImmunoSorbent Assay (ELISA).
2.2.9 The comparison of activities of monoclonal antibodies on LPPC PBMC (1×105 cells per well) or splenocyte (2.5×105 cells per well) were respectively dispensed into 96-well culture plates for monitoring cell proliferation. 40 µg LPPC previously adsorbed 100 µg BSA, and then adsorbed with 2.4 µg anti-CD3 mAb or 2.4 µg anti-CD3 and 2.4 µg
13
anti-CD28 mAb into 100 µl volume. After centrifuged, 2.5 µl LPPC complex respectively treat PBMCs or splenocytes, and comparing to the same amount mAb that free from added into solution. The cell numbers was counted by MTT assay at 72 hrs. The cell proliferation rate was calculated as O.D. value of sample divide into O.D. value of PBMC alone or splenocyte alone.
On the other hand, PBMC (4×105 cells per well) or splenocyte (106 cells per well) were dispensed into 24-well culture plates for monitoring cytokines secretion. 40 µg LPPC previously adsorbed 100 µg BSA, and then adsorbed with 6 µg anti-CD3 mAb or 6 µg anti-CD3 and 6 µg anti-CD28 mAb into 100 µl volume. After centrifuged, 10 µl LPPC complex respectively treat PBMCs or splenocytes, and comparing to the same amount mAb that free from added into solution. And the
supernatants were collected at 24h and 72 h and frozen at −80 ℃.
Supernatants concentrations of TNF-α, IL-2, and IFN-γ were measured
by Enzyme-Linked ImmunoSorbent Assay (ELISA).
2.2.10 The uptake protein ability of P338D1
50 µg BSA-FITC as a green fluorescence protein was previously adsorbed by 150 µg LPPC or was not adsorbed, and then respectively
co-cultured two hours with 5 ×105 mouse macrophage, P338D1. Added
100 µl trypan blue to quench the green fluorescence from BSA-FITC that was not uptaken or only adhered to cell surface, and FACS analysis was performed. In addition, 50 µg BSA-FITC as a green fluorescence protein was previously adsorbed by 150 µg LPPC or 10 µg LPPC, and then
14
respectively co-cultured two hours with 5 ×105 mouse macrophage,
P338D1.
2.2.11 DC harvest
Balb/C mice were sacrificed by dislocation. Make a long transverse cut through the skin in the middle of the abdominal area. Reflect skin from the hindquarters and the hind legs. Removed the feet, and then removed all muscle from the femurs and tibiae. Separate the legs from the body at the hip joint (one leg each time). Transfer the bones to a 15 mL centrifuge tube containing cold RPMI. Place the bones in a 10 cm bacterial dish containing 70 % ethanol for less 2~5 min for disinfection, then washed with RPMI. Separate femurs and tibiae. Cut both ends of the bone with scissors and the marrow flushed with RPMI10 using a Syringe with a 25 G needle. Collect cell suspension in a 10 cm bacterial dish. Clusters within the cell suspension were disintegrated by vigorous pipetting. Transfer the cell suspension to a 15-mL centrifuge tube. Centrifuges at RT, 300g for 5 min and then discard the supernatant. Add 2 mL of ACK lysis buffer to lyse red cells for 45 sec. The mixture is then added with 10 mL of RPMI10and centrifuges at RT, 300g for 5 min to wash out ACK. Discard the supernatant, and then suspend the cell pellet and then add with 10 mL of RPMI. Transfer the suspension to another tube to remove the settled debris and clumps. Centrifuges at RT, 300g for 5 min and discard the supernatant. Count cell number and then BM leukocytes were
seeded at 2.5×106 per 100 mm dish in 10 mL R10 medium containing 200
U/mL rmGM-CSF. At day 3, another 10 mL RPMI10 medium containing 200 U/mL rmGM-CSF were added to the plates. At days 6, half of the
15
culture supernatant was collected (10 mL/dish), centrifuged at RT, 300g for 5 min, and the cell pellet resuspended in 10 mL fresh RPMI10 containing 200 U/mL rmGM-CSF/dish, and given back into the original plate. At days 8, half of the culture supernatant was collected (10 mL/dish), centrifuged at RT, 300g for 5 min, and the cell pellet resuspended in 10 mL fresh R10 containing 200 U/mL rmGM-CSF/dish, and given back into the original plate. At day 9 or 10, non-adherent cells were collected by gentle pipetting. Cells were centrifuged at 300g for 5
min at RT, and resuspended in 10 mL fresh R10 (106 per mL) into a fresh
100 mm tissue culture plastic dish containing 100 U rmGM-CSF and 0.5
µg/mL LPS (–20℃, A11, 100 µg/mL). Cells were then cultured for 1 or 2
days for further experiment (complete maturation). The mature dendritic cells were checked by staining with anti-mouse CD11 conjugated PE and analyzed by flow cytometry.
Purification of DC membrane protein
Harvested DC cells (1×107 cells) were by centrifuging the cell
suspension or culture at 900g for 10 min at 4℃. Resuspend the cell pellet
in 10 ml PBS buffer and centrifuged at 900g for 10 min at 4℃. Resuspend
the cells in 10 ml HEPES-KOH buffer. Homogenize the cells on ice to fine homogenate using an appropriate cell homogenizer. The cells were
centrifuged at 9000g for 15 min at 4℃. Transfer the supernatant into fresh
ultracentrifuge tubes and discard the pellet. The fresh ultracentrifuge tubes centrifuged at 50000g at 4℃. Discard the supernatant, briefly air dry, and save the membrane pellet. The membrane pellet was dissolve in the PBS buffer. The concentration of membrane proteins was analyzed by
16
using commassie plus test.
Dendritic cell surface marker staining
106 DCs were centrifuged the cells at 4000 rpm for 5min. Resuspend
the cells with 500 µl staining buffer (0.5% skim milk in PBS). Stain the cells with antibody on ice in the dark for 30min. After washing the cells with 500 µl staining buffer, centrifuge the cells at 4000 rpm for 5min. Repeat again. Analyze the cells on FACScan with dot plots with quadrant line. Figure 11 indicated that the surface marker expression of DCs, such as CD11c, MHC II, and CD86.
17
2.2.12 The experimental strategy of animal immunization Bovine serum albumin (BSA)
1st immunize (1mg/mouse) 2 weeks 1st boost (1mg/mouse) 1 week 2nd boost (1mg/mouse) 1 week 3rd boost (1mg/mouse) Experiment
Six- to eight-weeks old female BALB/c mice were purchased from the National Laboratory Center and housed in a temperature- and light-controlled room (12L:12D) at the Animal Maintenance Facility of National Chiao Tung University. The mice were first immunized by subcutaneously (s.c.) injection of 1mg/100µl BSA emulsified in CFA. The mice were boosted by subcutaneously (s.c.) injection of 1mg/100µl BSA emulsified in IFA, and all mice were sacrificed postchallenge. The experiment strategy of mice immunization followed above the protocol. The negative group was injected with 100 µl PBS alone.
18
Heat shock protein 60 of Helicobacter pylori
1st immunize (100µg/mouse) 2 weeks 1st boost (100µg/mouse) 1 week 2nd boost (100µg/mouse) 1 week 3rd boost (100µg/mouse) Experiment
Six- to eight-weeks old female BALB/c mice were purchased from the National Laboratory Center and housed in a temperature- and light-controlled room (12L:12D) at the Animal Maintenance Facility of National Chiao Tung University. The mice were first immunized by subcutaneously (s.c.) injection of 100µg/300µl HpHsp60 emulsified in CFA. The mice were boosted by subcutaneously (s.c.) injection of 100µg/300µl HpHsp60 emulsified in IFA, and all mice were sacrificed postchallenge. The experiment strategy of mice immunization followed above the protocol. The negative group was injected with 300 µl PBS alone.
19 HPV E7 epitope 1st immunize (30µg /mouse) 2 weeks 1st boost (30µg /mouse) 1 week Experiment
Six-eight weeks female C57BL/6-Tg(HLA-A2.1) mice were kindly provided from Dr. Shih-Jen Liu (National Health Research Institutes) and housed in a temperature- and light-controlled room (12L:12D) at the Animal Maintenance Facility of National Chiao Tung University. The mice were first immunized by subcutaneously (s.c.) injection of 30µg/100µl YML peptides emulsified in CFA. The mice were boosted by subcutaneously (s.c.) injection of 30µg/100µl YML peptides emulsified in IFA, and all mice were sacrificed postchallenge. The experiment strategy of mice immunization followed above the protocol. The negative group was injected with 100 µl PBS alone.
20
2.2.13 The enhancement of antigen presentation of APCs by LPPC Cell proliferation
First, 100 µg BSA proteins were adsorbed by 40 µg LPPC into 100 µl volume. Splenocytes that isolated from were prior immunized by BSA
(2.5×105 cells per well) were respectively dispensed into 96-well culture
plates for monitoring cell proliferation. The 2.5 µl LPPC-complex co-cultured with splenocytes, and MTT assay was used to estimate at 72 hrs. The cell proliferation rate was calculated as O.D. value of sample divide into O.D. value of splenocyte alone. Negative control was the splenocytes from naive mice.
Cytokines secretion
First, 100 µg BSA proteins were adsorbed by 40 µg LPPC into 100
µl volume. Splenocytes that isolated from were prior immunized by
HpHSP60 (106 cells per well) were respectively dispensed into 24-well culture plates for monitoring cytokines secretion. The 10 µl LPPC-complex co-cultured with splenocytes, and the supernatants were
collected at 24h and 72 h and frozen at −80 ℃. Supernatants
concentrations of TNF-α, IL-2, IL-10, IL-4, and IFN-γ were measured by
Enzyme-Linked ImmunoSorbent Assay (ELISA).
2.2.14 Membrane proteins with specific antigen coated LPPC Cell proliferation
50 µg membrane proteins contained BSA or HpHSP60 antigens isolated from DCs and 100 µg BSA proteins were for 40 µg LPPC adsorption into 100 µl volume. Splenocytes that isolated from were prior
21
immunized by HpHSP60 (2.5×105 cells per well) were respectively
dispensed into 96-well culture plates for monitoring cell proliferation. The 2.5 µl LPPC-complex co-cultured with splenocytes, and MTT assay was used to estimate at 72 hrs. The cell proliferation rate was calculated as O.D. value of sample divide into O.D. value of splenocyte alone. Negative control was the splenocytes from naive mice.
Cytokines secretion
50 µg membrane proteins contained BSA or HpHSP60 antigens isolated from DCs and 100 µg BSA proteins were for 40 µg LPPC adsorption into 100 µl volume. Splenocytes that isolated from were prior
immunized by HpHSP60 (106 cells per well) were respectively dispensed
into 24-well culture plates for monitoring cytokines secretion. The 10 µl LPPC-complex co-cultured with splenocytes, and the supernatants were
collected at 24h and 72 h and frozen at −80 ℃. Supernatants
concentrations of TNF-α, IL-2, IL-10, IL-4, and IFN-γ were measured by
Enzyme-Linked ImmunoSorbent Assay (ELISA).
2.2.15 The specific peptide-loaded HLA-A2 adsorbed on LPPC Cell proliferation
50 µg YML peptide-loaded HLA-A2 molecules and 100 µg BSA proteins were for 40 µg LPPC adsorption into 100 µl volume. Splenocyte
that immunized by YML antigen (2.5×105 cells per well) were
respectively dispensed into 96-well culture plates for monitoring cell proliferation. The 2.5 µl LPPC-complex co-cultured with splenocytes, and MTT assay was used to estimate at 72 hrs. The cell proliferation rate
22
was calculated as O.D. value of sample divide into O.D. value of splenocyte alone. Negative control was the splenocytes from naive mice. Cytokine secretion
50 µg YML peptide-loaded HLA-A2 molecules and 100 µg BSA proteins were for 40 µg LPPC adsorption into 100 µl volume. Splenocyte
that prior immunized by YML antigen (106 cells per well) were
respectively dispensed into 24-well culture plates for monitoring cytokines secretion. The 10 µl LPPC-complex co-cultured with splenocytes, and the supernatants were collected at 24h and 72 h and
frozen at −80 ℃. Supernatants concentrations of TNF-α, IL-2, and IFN-γ
were measured by Enzyme-Linked ImmunoSorbent Assay (ELISA).
2.2.16 The animal immunization of the immuno-LPPC in vivo
Six- to eight-weeks old female Balb/c mice were purchased from the National Laboratory Center and housed in a temperature- and light-controlled room (12L:12D) at the Animal Maintenance Facility of National Chiao Tung University. The 200µg LPPC previously adsorbed 250 µg peptide-loaded membrane proteins. And then the mice were immunized by intravenous (i.v.) injection of membrane proteins /LPPC complex and all mice were sacrificed after two weeks. The negative group was injected with 300 µl PBS alone.
Six-eight week female C57BL/6-Tg(HLA-A2.1) mice were kindly provided from Dr. Shih-Jen Liu (National Health Research Institutes) and housed in a temperature- and light-controlled room (12L:12D) at the Animal Maintenance Facility of National Chiao Tung University. The
23
200µg LPPC previously adsorbed 25 µg peptide-loaded HLA-A2 molecules and 25 µg anti-CD28 mAb. And then the mice were immunized by intravenous (i.v.) injection of LPPC complex and all mice were sacrificed after two weeks. The negative group was injected with 300 µl PBS alone.
2.2.17 The animal immunization efficiency of immuno-LPPC in vivo Cell proliferation
Splenocytes that isolated from were prior immunized by
immuno-LPPC or PBS (2.5×105 cells per well) were respectively
dispensed into 96-well culture plates for monitoring cell proliferation. HpHsp60 or YML peptides (2µg/ml) were co-cultured with splenocytes, and MTT assay was used to estimate at 72 hrs. The cell proliferation rate was calculated as O.D. value of sample divide into O.D. value of splenocyte alone. Negative control was the splenocytes from naive mice. Cytokine secretion
Splenocytes that isolated from were prior immunized by
immuno-LPPC or PBS (106 cells per well) were respectively dispensed
into 24-well culture plates for monitoring cytokines secretion. HpHsp60 or YML peptides (2µg/ml) were co-cultured with splenocytes, and the
supernatants were collected at 24h, 48 h and 72 h and frozen at −80 ℃.
Supernatants concentrations of TNF-α, IL-2, IL-10, IL-4, or IFN-γ were
measured by Enzyme-Linked ImmunoSorbent Assay (ELISA).
24
All figures are expressed as mean ± SD. All data were computed by student-test. All statistical significant was set at p < 0.05.
25
Chapter 3 Result
3.1 The characters of LPPC
As figure 1a shown, the shape of LPPC was approximately round and the particle size was about 200 nm. In addition, the dark shadow of LPPC was hair-like, which might be PEI and PEG polymers. LPPC is a cationic liposome, and it was found that LPPC can adsorb proteins on its surface. Therefore, DLS was utilized to investigate the particle size of LPPC with or without protein adsorption. The results showed that the diameters of LPPCs with protein adsorption were about 358 ± 16 nm, which was larger than the LPPC without protein adsorption (Figure 1b). Besides, the previous results have shown the empty LPPCs can be centrifuged and pelleted (Figure 1c) and the further experiments also indicated the protein adsorption did not affect this character. Because centrifugation is available for LPPC, unbound substances could be easily removed.
3.2 The characters of LPPC for protein adsorption
To understand the kinetic for protein adsorption to LPPC, the fluorescence of BSA-FITC was used to evaluate what time the LPPC need to adsorb proteins to their surface. The results showed that LPPC could adsorb 80% of proteins in ten minutes and reach the maximal adsorption in 20 minutes (Figure 2a). Moreover, the protein binding capacity of LPPC was estimated and the results revealed that the maximal adsorption of 40 µg LPPC was about 160 µg BSA (Figure 2b).
26
Surprisingly, the pre-adsorbed BSA-FITC on LPPC could not be replaced by the additions of different BSA dose (Figure 2c). The results showed that the pre-adsorbed proteins on LPPC were irreplaceable by the posterior added proteins.
3.3 The activities of immunostimulatory monoclonal antibodies adsorbed on LPPC
3.3.1 The cytotoxicity of LPPC to PBMCs or splenocytes
LPPC could adsorb proteins stably and remain their activities as previous experiments (Table 1). Therefore, to investigate whether LPPC could adsorb immunostimulatory monoclonal antibodies and stimulate immunity was further proceeded. First, the cytotoxicity of LPPC was determined for PBMCs or splenocytes at next experiment. The results indicated that 1 µg LPPC was an appropriate dosage for 105 PBMCs or
2.5×105 splenocytes, because the cells could survive without toxic
damage in this concentration (Figure 3).
3.3.2 The effects of the bound antibodies in a dosage-dependent manner
In this study, anti-CD3 and anti-CD28 monoclonal antibodies (mAbs) were utilized to activate T cell, which were used to determine whether the bound protein on LPPC could remain its biofunction. In order to understand the regulatory phenomenon of monoclonal antibodies on LPPC for activities, the different amounts of immunostimulatory mAbs were adsorbed on LPPC to stimulate the proliferation of PBMCs or
27
splenocytes. The results showed the cell proliferation rates of PBMCs and splenocytes were higher as anti-CD3 mAbs were increased. It would be more obvious when the anti-CD3 mAbs combined with anti-CD28 mAbs to work on immune cells (Figure 4). Therefore, the bound antibodies on LPPC could activate the cell in a dosage-dependent manner.
Further, whether the secretions of cytokines were triggered by the bound mAbs on LPPC in a dose-dependent manner was investigated and the results indicated that LPPC and the adsorbed immunostimulatory mAbs could stimulate PBMCs or splenocytes to secrete cytokines, such as IL-2, IFN-γ and TNF-α. In addition, the concentrations of cytokines in media were increased as the anti-CD3 mAbs were increased (Figure 5). Moreover, the expressions of cytokines were increased by the addition of anti-CD28 mAbs which could provide the costimulatory signal to enhance the T-cell response as previously reported. Besides, LPPC alone could activate TNF-α secretion of PBMCs and splenocytes but it could not trigger any the IL-2 and IFN-γ secretions (Figure 5). Therefore, the LPPC reagent was investigated further to analyze whether the inductions of other pro-inflammatory cytokine profiles were. The results showed that
the LPPC could stimulate IL-1β, IL-6 and IL-8 secretions of immune
cells, except for TNF-α expression (Figure 6).
Comparing to the activities of unbound mAbs, the bound mAbs showed there were no significant differences between the cell proliferation and cytokine secretions (IL-2, IFN-γ) (Figure 7). Nevertheless, LPPC with mAbs could enhance TNF-α secretion comparing to unbound mAbs, it should be due to the LPPC’s ability to facilitate the TNF-α secretion.
28
3.4 The stability of immunostimulatory monoclonal antibodies adsorbed on LPPC in RPMI
To investigate the stability of the bound mAbs on LPPC in 37℃, the activities of the dissociated antibodies in the medium were estimated. The results indicated that the antibodies on LPPC pellet remained their partial
activities after 37℃ treatment to induce 90% proliferation for PBMCs or
splenocytes (Figure 8). In addition, the antibodies in supernatant only displayed low activities to cause inferior proliferative index for PBMCs or splenocytes. These results revealed that the antibodies bound on LPPC would rather stably adhere than dissociate from the particle.
3.5 The enhancement of uptake protein ability by LPPC
The previous results indicated that LPPC alone has the ability to enhance pro-inflammatory cytokine secretions of PBMCs and splenocytes (Figure 5, 6). To understand whether LPPC has an adjuvant effect for the enhancement of antigen uptake by phagocytosis, the phagocytic rate for the fluorescent antigen was evaluated. As to the ability of phagocytosis, LPPC/BSA-FITC complexes indeed enhanced the uptake ability of P338D1 compared to BSA-FITC without LPPC adsorption (Figure 9a). In addition, as the additional amounts of LPPC were increasing, the uptake abilities were enhanced with the more efficiency (Figure 9b) and the results also showed that the phagocytic rates of P338D1 were in a dose-dependent manner (Figure 9c).
29
showed that the addition of BSA alone could be internalized by APC and present to the specific anti-BSA T-cells, which could trigger and increase the cell proliferation and cytokine secretions (including IL-2, IFN-γ, TNF-α, IL-4 and IL-10). By contrast, the results indicated that BSA adsorbed on LPPC could provide more efficacies to induce cell proliferation and cytokine secretions than BSA without LPPC adsorption (Figure 10).
3.6 The specificities of the LPPC-bound proteins
Certain immunostimulatory molecules on cell membrane, such as MHC or B7 molecules could trigger the specific immune response, so that DCs’ proteins on plasma membrane were determined whether they could perform their activities on LPPC surface as same as on the plasma membrane. Before isolated the plasma membrane proteins of DCs, anti-CD11c-PE, anti-MHC II-FITC, and anti-CD86-FITC were used to confirm that the surface markers of DCs performed (Figure 11). The membrane proteins derived from the DCs which were treated with HpHsp60 were bound to LPPC, which could react with the splenocytes derived from the mice had been prior immunized with HpHsp60 to induce the cell proliferation and cytokine secretions (Figure 12). However, neither the splenocytes without Hphsp60-immunized nor the DC’s membrane proteins without Hphsp60-treated could induce the cell proliferation or cytokine releases (Figure 12).
Moreover, YMLDLQPETT peptides (YML) derived from HPV E7 protein were loaded into the HLA-A2 molecules to verify the specificity of the LPPC-bound proteins again. The YML-loaded HLA-A2 molecules
30
on LPPC were interacted with the splenocytes derived from naïve or pre-immunizing E7 mice. The results showed that peptide-loaded HLA-A2 molecules on LPPC indeed remained their specificities to activate the splenocytes of pre-immunizing E7 mice and cause the cell proliferation and cytokine expressions but did not react with the naïve splenocytes (Figure 13). In addition, the anti-CD28 mAbs could facilitate the immune responses for the splenocytes of pre-immunizing E7 mice but have no effect on the naïve splenocytes (Figure 13).
3.7 The efficiency of immunization in vivo
The cell proliferations and cytokine expressions of the splenocytes which were i.v. immunized by immuno-LPPC estimated whether the induction of specific immune responses. The induction of the specific anti-HpHsp60 immune responses of splenocytes from membrane proteins /LPPC complex immunized was more efficient than that from HpHsp60 antigen (Figure 14). In addition, the results indicated that the splenocytes form the other immunization methods did not significantly react to HpHsp60.
Furthermore, the splenocytes that the LPPC combined
peptide-loaded HLA-A2 molecules with anti-CD28 mAbs immunized were efficiently activated immune responses against YML peptides, such as cell proliferations and cytokine secretions (Figure 15). No apparent immune responses were observed against YML peptides in the other immunization methods.
31
Chapter 4 Discussion
To summarize, we have established a novel platform for immunoregulation which utilized the LPPC to combine with certain immunostimulatory molecules, such as mAbs and MHC-loaded peptides molecules, and in this study the LPPC display its potential to be an artificial APC. First, the LPPC is easily adsorbing a variety of immunostimulatory proteins and the LPPC-bound proteins can remain their activities. Second, the LPPC have an adjuvant effects to enhance proinflammatory cytokine expression, antigen uptake and presentation of APC. Furthermore, our study provided the evidence that the regulatory
molecules/LPPC complexes dramatically increase the specific
anti-antigen activities of immune responses in vivo. As understanding the mechanism about immune responses, it is proposed that certain immunostimulatory molecules could be applied in this platform to regulate immunity for disease therapy. For example, the combination of microbeads with HLA-A2 monomers, anti-4-1BB antibodies and anti-CD28 antibodies can expand large numbers of antigen specific CTLs (33). Moreover, the addition of anti-LFA-1 mAbs with anti-CD3, anti-CD28 could enhance the T-cell expansion for adoptive cell therapy (34). Therefore, certain ligands or costimulatory molecules might be utilized for adsorption of LPPC to regulate immunity as artificial APCs.
The major defect for DC-based therapy was that the maturation of DCs was weaken by certain immunosuppressive substances such as
32
Moreover, Candida albicans, Mycobacterium tuberculosis, the secretions of mycobacterial, and the secretions of Candida respectively impact on efficiency of DCs and immune cells, affect cytokine expression and impair surface marker of DC (19-22). Many Comparing to our strategy, previous DC-based therapy strategies were DCs incubated with antigens such as tumor antigen HSP105, HPV E7 protein, breast tumor cell lysate, HIV, or SIV to successfully induce host immunity against tumor or pathogens (15, 16, 35-37). We modified these previous strategies in this study, the membrane proteins of DCs which were prior treated and up-take antigens (HpHsp60) were isolated and were adsorbed on LPPC, later the membrane antigen/LPPC complexes would be used as antigen to immunize animals. The results revealed the LPPC adsorbed membrane proteins could induce specific immune responses efficiently (Figure 13). The modified strategy excludes the usage of alive DC to prevent the above problem.
Moreover, immuno-molecules such MHC molecules are covalently coupled to beads or liposomes and used to expand CTL ex vivo (38, 39). However, such chemical modification might impair the original functions or activities of immunoregulatory or targeting molecules (40). In contrast, mAb non-covalently adsorbed on the nanoparticle surface could provide a better activity than covalent coupling (41). In this study, the LPPC provided an ability to strongly adsorb proteins on its surface, which was proposed that the activities of the bound immunoregulatory molecules could maintain higher activities without chemical damage. Moreover, figure 2c and figure 8 indicated that the pre-adsorbed proteins on the
33
surface of LPPC may be irreplaceable in serum without the dissociated doubt.
On the other hand, the results in this study indicated that the LPPC had the interesting characters, the effects of adjuvant. Figure 9 showed LPPC promoted antigen uptake ability of APC in a dose-dependent fashion. In addition, BSA adsorbed on the LPPC provided a better presentation of APC than free BSA to induce T-cell proliferations and cytokine secretions (Figure 10). Moreover, LPPC showed it could directly induced PBMCs or splenocytes to secret proinflammatory cytokines, IL-1β, IL-6, IL-8 or TNF-α. It is well-known that the function of the proinflammatory cytokines could recruit or activate macrophage or other immune cells (42). Similarly, the adjuvant MPL reagent, a liposome-like reagent could also improve the capacity of APCs by enhancements of antigen uptake and cytokine release (43). Furthermore, their primary adjuvant mechanism of cationic liposomes is to target to the membrane of APCs and to induce the uptake and increased presentation of antigen (44). Together these results indicated that the LPPC can be a good adjuvant.
In vivo experiments, the figure 13 and figure 14 showed that the proteins/ LPPC complexes induced a better specific immune response than free proteins of immunization. We proposed that the LPPC might provide a little positive charge to attract immune cells, and the higher density of the immunoregulatory molecules on the LPPC than free form of molecules might also provide good affinity. Moreover, the LPPC reagent had an adjuvant effect. Therefore, the LPPC should be a good
34
immune platform of enhancement of immune response.
Many investigators have started to develop artificial APCs and Dr. Mathias Oelke classified artificial APCs according to cell-based or non-cell-based in current approaches for immunotherapy (1). Early, murine MHC class I molecules with the costimulatory molecules B7-1 and ICAM-1 were co-expressed on drosophila cell to activate T cells, which resulted in specific killing for tumor cells in vitro and in vivo (45). However, the transfected drosophila cells were unstable in mouse body. Another cell-based approach was also used. The NIH/3T3 murine fibroblast cells were transduced with the costimulatory molecule B7-1, ICAM-1 and LFA-3, and a single HLA–peptide complex (HLA-A2) as APCs. These aAPCs have proved that they can induce specific CTLs to against tumor cells when the aAPC were pulsed with tumor antigens MART-1 or hTERT (29, 46). In acellular systems, magnetic beads were usually used to assemble as aAPCs, (31, 47, 48). In addition, latex microspheres (49) or liposomes (32) were also developed as aAPC. Generally, the immunoregulatory molecules on these aAPC are MHC molecules or immunoregulatory antibodies. For instance, HLA-A2–Ig molecules, biotinylated murine MHC class I or MHC class II-peptide molecules have been combined with costimulatory molecules such as anti-CD28 antibodies or B7 molecules, which were coupled on particle surface to induce and expand antigen-specific T cells (31, 32, 49).
In this study, the LPPC adsorbing with anti-CD3 and anti-CD28 mAbs or peptide–loaded HLA-A2 provided immunoregulatory function
35
to stimulate T cells, similar to other acellular-based APCs. But the difference between LPPC and other aAPC is that LPPCs do not need chemical modification and complicated purification. In addition, LPPC can also adsorb membrane proteins to trigger the specific T cells responses (Figure 11 and 13). As our knowledge, no other aAPC coupled with DC’s membrane proteins to induce specific immunity. Moreover, figure 11 and 12 showed that neither DC’s membrane proteins nor peptide-loaded HLA-A2 could induce stronger immune responses than these antigens alone except these molecules combined with LPPC. Figure 13 also revealed that LPPC/ DC’s membrane proteins complex could enhance antigen-induced Th-1 cytokine releases but its abilities to induce Th-2 cytokine releases were as same as antigen alone immunization. Therefore, LPPC should be developed as a convenient and efficient immunoregulatory platform to mediate host immunity.
Interestingly, the results showed that LPPC alone could stimulate immune cells to release pro-inflammatory cytokines (Figure 5 and 6). Its mechanism may result from PEI, one component of LPPC, has been proved it was an adjuvant effects to enhance the expressions of both Th1 and Th2 cytokines (50). However, LPPC alone could only induce the
proinflammatory cytokine releases including TNF-α (Figure 5), IL-1β,
IL-6 and IL-8 (Figure 6) but it had no effects on the releases of Th-1 and
Th-2 cytokines including IL-2 (Figure 5a and 5d) , IFN-γ (Figure 5b and
5e), IL-4 (Figure 10) and IL-10 (Figure 10). Because the proportion of
PEI in LPPC is 3%, only 1.62 µg PEI would interact with immune cells
36
(50), the difference between the dosages may result in different immunoregulation (51, 52). Except for the adjuvant effect, LPPC preferentially enhance Th-1 responses. Th-1 responses such as
Th1-CD4+T cell or CTL have been demonstrated that they are important
and necessary against malignant tumors or intracellular pathogens. Thus, the scientists attempted to design the strategies of vaccination by enhancement of Th-1 immunity (53-55). According to our results, LPPC seem to be a good adjuvant which can be developed as the vaccines against tumor or intracellular pathogens.
In conclusion, we demonstrated that LPPC could show its flexible and convenient characters as an artificial APC that stimulate immune responses and induce specific immunity in vitro and in vivo. The
immuno-LPPC has immuno-function by adsorbing certain
immunoregulatory molecules. Except activation of immunity, the LPPC may also combine with immunosuppressive molecules or apoptosis ligand, such as CTLA-4 and FasL, to provide different function to suppress abnormal immunity (11, 56). In the future, the immuno-LPPC may have broad functional immuno-regulatory abilities to treat human diseases by mediating host immunity.
37 Figure 1 (a) (b) (c)
Before adsorbing After adsorbing
0 20 40 60 80 100 0 100 200 300 400 500 particle size (nm) re la ti v e pe rc en ta g e (% ) LPPC alone LPPC-BSA complex
38
Figure 1. The characters of LPPC. (a) The TEM photo of LPPC; (b) The particle size distribution of LPPC with proteins or without proteins were estimated by utilizing DLS. The average particle size of LPPC
without BSA proteins was about 216 ± 4 nm, and the average particle size of LPPC with BSA proteins was about 358 ± 16 nm; (c) The centrifuged property of LPPC with or without proteins.
39 Figure 2 (a) Timing Course NC 5 10 15 20 25 30 0 50 100 150 Time (min) A d s o rb in g P e rs e n ta g e (% ) (b) Capacity 0 40 80 120 160 200 240 0 50 100 150 200 addition of BSA (µµµµg) d e te c ti o n o f B S A ( µµµµ g )