Vol. 39: 13-19.1999 DISEASES OF AQUATIC ORGANISMS
Dis Aquat Org
1
Published December 22Studies on effective
PCR
screening strategies for
white spot syndrome virus (WSSV) detection in
Penaeus monodon
brooders
Hui-Chen Hsul, Chu-Fang LO',
Shan-Ching Linl,
Kuan-Fu
~ i u ~ ,
Shao-En
Peng',
Yun-Shiang Changl, Li-Li Chenl,
Wang-Jing Liu', Guang-Hsiung
K o u l ~ *'Department of Zoology, National Taiwan University, Taipei. Taiwan, ROC
'~ung-Kang Marine Laboratory, Taiwan Fisheries Research Institute, Tung-Kang. Ping-Tung, Taiwan, ROC
ABSTRACT: We re-tested stored (frozen) DNA samples in 5 independent polymerase chain reaction (PCR) replicates and confirmed that equivocal test results from a previous study on white spot syn- drome virus (WSSV) in brooders and their offspring arose because amounts of WSSV DNA in the test samples were near the sensitivity limits of the detection method. Since spawning stress may trigger WSSV replication, we also captured a fresh batch of 45 brooders for WSSV PCR testing before and after spawning. Replicates of their spawned egg batches were also WSSV PCR tested. For these 45 brood- ers, WSSV prevalence before spawning was 67 % (15/45 l-step PCR positive, 15/45 2-step PCR positive and 15/45 2-step PCR negative). Only 27 (60%) spawned successfully. Of the successful spawners, 56% were WSSV PCR positive before spawning and 74 % after. Brooders (15) that were heavily infected (i.e. l-step PCR positive) when captured mostly died within 1 to 4 d, but 3 (20%) did manage to spawn. All their egg batch sub-samples were l-step PCR positive and many failed to hatch. The remaining 30 shrimp were divided into a Lightly infected group (21) and a 2-step PCR negative group (9) based on replicate PCR tests. The spawning rates for these 2 groups were high (81 and 78%, respectively). None of the negative spawners (7) became WSSV positive after spawning and none gave egg samples posi- tive for WSSV. In the lightly infected group (21), 6 brooders were 2-step WSSV PCR negative and 15 were 2-step WSSV PCR positive upon capture. However, all of them were WSSV PCR positive in repli- cate tests and after spawning or death. Four died without spawning. The remaining 17 spawned but only 2 gave egg samples PCR negative for WSSV. The other 15 gave PCR positive egg samples, but they could be divided into 2 spawner groups: those (7) that became heavily infected (i.e. l-step PCR positive) after spawning and those (8) that remained lightly infected (i.e. became or remained 2-step PCR positive only). Of the brooders that became heavily infected after spawning, almost all egg sam- ple replicates (91%) tested 2-step PCR positive One brooder even gave heavily infected (i.e. l-step PCR positive) egg samples. For the brooders that remained lightly infected after spawning, only 27 % of the egg sample replicates were 2-step PCR positive. Based on these results, we recommend that to avoid false negatives in WSSV PCR brooder tests screening tests should be delayed until after spawn- ing. We also recommend, with our PCR detection system, discarding all egg batches from brooders that are l-step PCR positive after spawning. On the other hand, it may be possible with appropriate moni- toring to use eggs from 2-step PCR positive brooders for production of WSSV-free or lightly infected postlarvae. These may be used to stock shrimp ponds under low-stress rearing conditions.
KEY WORDS: WSSV
.
Carrier - Brooder. Penaeus monodonINTRODUCTION
Outbreaks of white spot syndrome virus (WSSV) have led to serious mortalities among populations of cultured shrimp in several Asian countries (Inouye et al. 1994, Momoyama et al. 1994, Nakano et al. 1994,
'Corresponding author. E-mail: [email protected]
Takahashi et al. 1994, Cai et al. 1995, Chou et al. 1995, Wang et al. 1995, Wongteerasupaya et al. 1995, Lo et al. 1996a, Peng et al. 1998). The principal clinical sign of this disease is the presence of white spots in the exoskeleton and epidermis of diseased shrimp, with the spots varying in size from barely visible to 3 mm
in diameter. Histopathological studies have demon- strated that the causative agent, WSSV, most fre- quently attacks the cuticular epidermis, where its pres- O Inter-Research 1999
14 Dis Aquat Or
ence is evident in degenerated cells with hypertro- phied n u c l e ~ (Wang et al. 1995). The effects of WSSV pathogenesis a r e often acute, with mortality typically reaching l 0 0 % within 2 to 7 d (Momoyama e t al. 1994, Chou e t al. 1995, Wang et al. 1995). Recently, WSSV has also been shown to cause disease, high mortality and/or latent infections not only in shrimp, but also in crabs a n d other arthropods (Lo et al. 199613, 1997, Lo & Kou 1998).
WSSV targets various tissues of captured Penaeus monodon brooders, including the reproductive organs. In t h e ovary, Lo et al. (1997) found WSSV particles in follicle cells, oogonia, developing oocytes a n d connec- tive tissue. Investigation of WSSV infection by 2-step polymerase chain reaction (PCR) revealed that some positive brooders produced larval batches that gave both 2-step WSSV PCR positive a n d negative test results with different sub-samples (Lo et al. 1997). A more recent study of a single WSSV-carrier brooder revealed that 1 0 % of its 13 mo old, outwardly healthy, adult offspring had tiny white spots on their carapace a n d exhibited WSSV-infected cells in the stomach, integument, gills a n d other organs by in situ DNA hybridization (Tsai et al. 1999). These results lndicate that the quantity of WSSV DNA in tested brooders may sometimes b e near the limits of detection methods. In this paper, w e further investigated WSSV infections in brooders, in a n attempt to find a method for more effective screening. As part of this process, we re- examined stored samples of DNA from 2-step WSSV positive brooders described i n a previous publication (Lo et al. 1997). We also applied a more rigorous screening procedure to a batch of 1998 brooders a n d their offspring.
MATERIALS AND METHODS
Re-examination of stored DNA samples from 1995 brooders. The Penaeus monodon DNA samples re- tested here were derived from a n earlier study of cap- tured brooders (Lo et al. 1997) a n d h a d been stored at -20°C. Specimens from the earlier study were selected only if they h a d previously tested negative for WSSV or positive by 2-step PCR. Specimens that were l - s t e p PCR positive were not included. Sub-fractions of each selected sample were checked in 4 more independent PCR reactions.
Detection of WSSV in newly captured brooders be- fore and after spawning. Penaeus monodon brooders w e r e captured from their natural environment in the coastal waters around southern Taiwan, either in April or between August a n d November 1998. They were immediately transported to the Tung Kang Marine Laboratory of the Taiwan Fisheries Research Institute
in Ping Tung Prefecture, and upon arrival, a pleopod was removed from each shrimp. To minimize handling stress a n d collateral damage during pleopod removal, each brooder was wrapped in a wet towel so that only the appendage to be removed remained exposed. The pleopod was then excised with a red-hot pair of forceps in a manner similar to that for eyestalk ablation. Some of the DNA extracted from the pleopod was then sub- jected immediately to 2-step WSSV diagnostic PCR (Lo et al. 1996a) and the remainder was stored a t -20°C for later use. All the brooders used were mature and ready to spawn, so eyestalk ablation was not used to induce spawning. Subsequently, the brooders were main- tained in reproduction facilities (1 brooder in each 500 1 fiber reinforced plastic (FRP) tank containing 450 1 aerated, filtered 33%0 seawater at 30
*
l 0 C , pH 8.3) for spawning. A few hours after spawning, the brooders were transferred to n e w tanks (1 brooder in each 67 1 tank containing 50 1 aerated, filtered 33%0 seawater at 30 2 1°C, pH 8.3) to prevent horizontal transmission of WSSV. At the time of transfer, another pleopod was excised and subjected to DNA extraction. Most of the extracted DNA was stored at -20°C, a n d only a portion was used immediately for WSSV diag- nostic PCR to determine the prevalence of WSSV in brooders after spawning. This meant that the first 2PCR tests for each brooder were m a d e only hours apart, and this contrasts with our earlier study (Lo et al. 1997), where several days elapsed between the pre- and post-spawning tests. Brooders that failed to spawn were not tested a second time, except after death. Stored DNA samples extracted pre- a n d post-spawn- ing (or post-death) were subsequently sub-sampled a n d subjected to 5 additional replications of 2-step WSSV diagnostic PCR. Samples of eggs from all the brooders that spawned were also extracted a n d tested for the presence of WSSV using 2-step diagnostic PCR. WSSV diagnostic PCR. A DNA extraction kit (Farm- ing IntelliGene Tech. Corp., Taipei, Taiwan) was used to isolate the template DNA for diagnostic PCR. This kit is based on a simplified extraction protocol (Lo et al. 1996a, 1998). Briefly, a n excised pleopod was placed in a n Eppendorf tube containing 0.6 m1 of a premixed 'DTAB' solution provided with the extraction kit, crushed with a disposable bamboo stick, and then incubated for 5 min at 75OC. This was followed by extraction with 0.5 m1 of chloroform once, brief vortex- ing a n d centrifugation at 12000 X g for 3 min. The upper aqueous phase was transferred to a new Eppen- dorf tube with 0.1 m1 'CTAB' (also pre-mixed a n d pro- vided with the kit) a n d 0.9 m1 double-distilled H 2 0 for 5 min at 75°C before being allowed to cool down to room temperature. After centrifugation at 12 000 X g f o r
10 min, the pellet was collected a n d resuspended in 150 p1 of the 'Dissolving Solution' provided with the kit
Hsu et al.: Effective PCR sc :reening for WSSV detection 15
for 5 min at 75'C. After further centrifugation at 12 000 X g for 5 min, the supernatant was transferred to a new Eppendorf tube with 300 1-11 95 % ethanol, mixed well and centrifuged again at 12 000 X g for 5 min. The pel- let was then washed with 200 p1 70% ethanol and resuspended in double-distilled H 2 0 at 65°C for 15 min before the DNA concentration was measured by ultra- violet absorbance at 260 nm using an RNA/DNA cal- culator (GeneQuantII, Pharmacia Biotech, Sweden). WSSV diagnostic PCR was then performed as de- scribed previously (Lo e t al. 1996a,b).
RESULTS
Re-examination of stored DNA samples from 1995 brooders
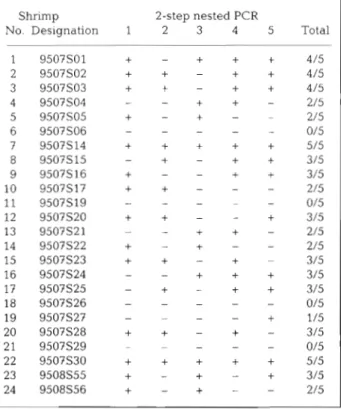
In the original replication of 2-step WSSV diagnostic PCR (Lo et al. 199?), 10 of the 24 tested DNA samples (41.67%) gave negative results (Table 1). However, after 4 more replications with the same templates, only 4 samples (16.67%) were consistently 2-step WSSV
Table 1. Penaeus rnonodon. Five replications of WSSV detection in brooders using 2-step WSSV diagnostic PCR. No specimens were originally l-step PCR positive. Designa- tion numbers used here consist of the year and month of capture followed by the specimen number originally used by Lo et a1 (1997). Data for the first replication are taken
from Lo et al. (1997) Shrimp 2-step nested PCR
No. Designation 1 2 3 4 5 Total
PCR negative and the other 20 gave a positive result at least once. This suggested that these 20 samples con- tained the WSSV template, although the number of positive PCR results obtained in each round of replica- tion only ranged from 42 to 58% (i.e. 10 to 14 out of 24)
Detection of WSSV in brooders before and after spawning
For the 45 newly captured brooders (Table 2), the immediate tests for WSSV after capture gave 15 (33 %) l-step PCR positive, 15 (33 %) 2-step PCR positive a n d 15 (33 %) 2-step PCR negative. The 15 brooders l-step WSSV positive upon capture were classified as 'heav- ily infected'. Three of these spawned but almost died afterwards. The other 12 failed to spawn and eventu- ally died. The 15 brooders 2-step WSSV positive upon capture were classified as 'lightly infected'. Of these, 3 failed to spawn and died, testing l-step positive after death. The 15 brooders 2-step negative upon capture fell into 2 groups, those that remained negative throughout testing and those that gave positive WSSV results after spawning or death. The latter were classi- fied a s 'lightly infected', like the brooders that were 2- step WSSV positive upon capture. Taken together, the 'lightly infected' brooders numbered 21. Four of these (1 being 2-step WSSV negative upon capture) did not spawn and died, testing l-step WSSV positive after death. The 17 remaining 'lightly infected' brooders spawned. After spawning, 4 of the initially 2-step neg- ative brooders became 2-step positive, and 1 became l-step positive, while 6 of the 2-step positive brooders became l-step positive and 6 remained 2-step positive. Of the 9 brooders that were 2-step negative, only 7 spawned and they gave WSSV negative tests after spawning. Only brooders that were 2-step negative after spawning were classified as truly 2-step WSSV negative.
Five additional 2-step PCR replicate tests (Table 3) were carried out using the stored parallel DNA extracts prepared upon shrimp capture and spawning. All 21 brooders assigned to the 'lightly infected' groups in Table 2 gave consistently positive results in 2-step PCR tests after spawning. However, results for their tests before spawning were somewhat variable. Assu- ming that the positive PCR result after spawning or death indicated a true infection, successful detection rates before spawning only ranged from 62 to 76 % for each round of replication (Table 3 ) . The 9 brooders classified as 2-step PCR negative in Table 2 were con- sistently 2-step WSSV PCR negative in all replicate tests, before a n d after spawning or without spawning. Indeed, the 2 brooders that did not spawn were still negative 1 mo after capture.
16 Dis Aquat Org 39: 13-19, 1999
Table 2. Penaeus monodon. Detection of WSSV by 2-step WSSV diagnostic PCR in 45 brooders before and after spatvnlng and eggs of these brooders. Designation numbers used here consist of the year and month of capture followed by the specimen num- ber. Brooders were sacrificed after spawning. Brooders that failed to spawn were not tested a second time or were tested only after death. Values for WSSV diagnostic PCR of eggs represent the number of DNA samples positive in the first or second step
PCR and the number of DNA samples examined. nd: not done
Shrimp Spawning Group WSSV diagnostic PCR of brooders WSSV diagnostic
No. Designation (Yes/No) Before spawning After spawning After death PCR of eggs
1st 2nd 1st 2nd 1st 2nd 1st 2nd Yes Yes Yes No No No No No No No No No No No No Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes No No No Yes Yes Yes Yes Yes No Yes Yes Yes Yes Yes Yes Yes No No Heavily infected Lightly infected Lightly infected 2-step PCR negative Not classified
Prevalence of WSSV in brooders after spawning Excluding the brooders that were l-step PCR posi- tive upon capture and examining only the brooders in the 'lightly infected' groups, there was evidence that spawning could result in increased severity of WSSV
infection. Excluding the 4 brooders that died, l l (65 % ) out of 17 brooders that spawned changed their WSSV Infection status either from 2-step PCR negative to 2 -
step positive (4/17 = 24 %) or l-step positi.ve (1/17 = 6 % ) or from 2-step positive to l-step positive (6/17 = 35 %).
Hsu et al.. Effective PCR screening for WSSV detection
Table 3. Penaeus rnonodon. Five repl~cat~ons of WSSV detection in brooders before and after spawnlng using 2-step WSSV diagnostic PCR. Designation numbers consist of the year and month of capture followed by the specimen number. Parentheses indicate that the specimens failed to spawn and were tested only after death. ++: specimens tested positive in both the first and second steps of the 2-step diagnostic PCR. Specimens 9810S67 and 9810S69 failed to spawn but survived through to the end of the 14 d experimental penod. Square brackets indicate that the stored D N A tested here was extracted 14 d after capture.
nd: not done
Shrimp Before spawning: 2-step PCR After spawning (after death): 2-step PCR
No. Designation 1 2 3 4 5 Total WSSV- 1 2 3 4 5 Total WSSV-
infected infected Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes nd nd nd No No No No No No Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes No No No No No No No No No
Negative impact of WSSV infection o n spawning brooders, those (6) that were l-step PCR positive after
spawning gave 89% 2-step positive egg samples, Of the 15 heavily infected brooders, only 3 ( 2 0 % ) while those that were only 2-step PCR positive after
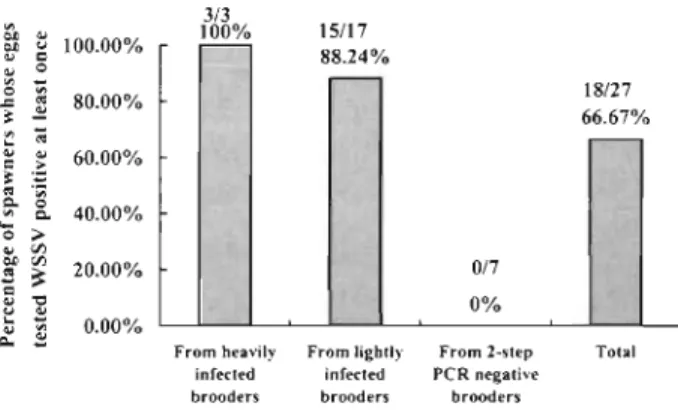
spawned successfully. Most of the others died within 1 spawning gave only 31 % (Table 2, Fig. 2). All of the to 4 d after capture without spawning. By contrast,
more than 80% (17 out of 21) of the lightly infected ~OO.OO% - 1712 l
brooders spawned successfully, as did 78 % (7 out of 9) 80.95%
717S%
80.00%
-
of the 2-step PCR negative brooders (Fig. 1). o 27/45 CI 60.00%
2
60.00%-
M C.-
Egg infectionsAll of the tested egg samples of the 3 heavily infected
brooders that spawned successfully were l-step PCR I
positive (Table 2, Fig. 2). Of the 17 lightly infected Heavily Lightly 2-step PCR Total
brooders that spawned, only 1 gave l-step PCR posi- infected infected negative
tive egg samples and only 2 gave 2-step PCR negative Fig. 1. Penaeus monodon. Negative impact of WSSV infection
Dls Aquat Org 39: 13-19, 1999
P I ;
F r o m heavily F r o m lightly F r o m 2-step Total infected infected PCR negative brooders brooders brooders
Fig 2. Penaeus monodon. Influence of brooder infection on egg Infection rate
tested eggs from the 2-step PCR negative brooders were 2-step PCR negative (Table 2 , Fig. 2 ) .
DISCUSSION
The inconsistency of 2-step PCR results from re- examination of stored DNA samples (Table 1) supports the hypothesis that equivocal PCR tests from offspring of 16 brooders in our earher study (Lo et al. 1997) resulted from detection sensitivity limits. Simllar results were obtained in a continuation study on the offspring of 1 brooder from the earlier study (Tsai et al. 1999). We therefore conclude that these samples h a d borderline concentrations of WSSV DNA and thus yielded different results with different sub-samples.
This sensltlvity limit raises doubts as to the reliability
of 2-step PCR for screening newly captured brooders, especially with respect to false negatives. Although the 2-step PCR has been shown to detect on the order of 20 copies of target plasmid in a PCR reaction mixture (Lo
& Kou 1998), this may translate to a requirement for u p to 1000 times more targets per gram of onginally extracted tissue, depending on the amount a n d con- centration of sample extract actually a d d e d to the PCR reaction vial. In addition, there is probably variation in numbers of viral particles in different pleopods from a single shrimp specimen. Because of these variables, it is likely that replicate PCR reactions from some lightly infected brooders would give inconsistent results such a s those in Table 2 or false-negative results such as some of the pre-spawning results in Table 3. On the other h a n d , the post-spawning results (Table 3) were completely consistent. The dramatic difference be- tween the pre- a n d post-spawning results supports our earlier finding that various kinds of stress - including
spawning stress - can trigger replication of WSSV (Lo
et al. 1997). Withln a few hours only, spawning stress brought virus q u a n t ~ t i e s to levels easily a n d reliably detected by 2-step PCR.
The value of a diagnostic test can be judged by its repeatabllity, tvhlch is an indication of reliability (Thrusfield 1995). For tests with a dichotomous out- come, such as the PCR tests used here, the repeatabil- ity can be determined by assessing the agreement between results when a test is repeated 2 or more tlmes. If a test is conducted 3 or more times, Cochran's Q-test can be used.
For the data in Table 3, the Cochran's Q-test was cal- culated for the replicates before a n d after spawning. Cochran's Q-test is calculated as:
where k = number of replicates, G = total number of positive results in each replicate, a n d L = total number of positive results in each animal. Thus, for the PCR data before spawning, Q = 4[(5 X 1019) - 5041]/[(5 X 71) - 3071 = 216/48 = 4.5. For the data after spawning, Q =
4[(5 X 2205) - 11 025]/[(5 X 105) - 5251 = 0.
The difference in the values of Q before a n d after spawning can b e clearly seen. Referring to tables of
x2
for 4 degrees of freedom the Q-statistic before spawn- ing gives a value which indicates a confidence level of around 50 %( x 2
= 4.352), demonstrating poor repeata- billty of the test in pre-spawning broodstock, whereas the 0 value for replicates after spawning indicates per- fect repeatability after spawning.It is possible to solve the problem of pre-spawning, false-negative WSSV tests for brooders by increasing the number of replicate PCR tests carried out for each specimen. Reference to Table 1 shows that 3 replicates detected all of the positive samples that were eventu- ally detected in 5 replicates, but in Table 3 some spec- imens required 5 replications. This is not a practical alternative d u e to the cost in time a n d money, so we propose that it would be more practical to screen brooders after spawning. At that time, it appears un- likely that borderline WSSV infections a r e obtained, a n d this should markedly decrease the probability of obtaining false-negative test results. Testing after spawning also has the added advantage of avoiding testing stress before spawning. This strategy may be
appropriate only when production throughput is rela- tively low or for shrimp species like Penaeus monodon
where e g g production per spawn is high and brooders and their offspring can b e individually reared. For example, production efficiency constraints necessitate pooling of P. vannamcli brooders and eggs. In this case individual broodstock holding, spawning and offspring rearing would not be practical, especially in high throughput hatcheries.
Although no l - s t e p WSSV PCR positive brooders in our previous study spawned successfully (Lo et al. 1997), 3 (20%) did here (Fig. 1 ) . Nonetheless, these
Hsu et al.: Effective PCR scl .eening for WSSV detection 19
3 brooders almost died after spawning. One (no. 9819S27) was even too weak to properly suspend its eggs and so they did not subsequently hatch into nau- plii. In addition, 100% of the egg samples were l-step PCR positive for WSSV. Obviously, such brooders should be discarded before spawning.
We have already shown that successful harvests can- not be expected from ponds stocked with postlarvae derived from WSSV-infected eggs (Lo et a l . 1998). Although the spawning rates of lightly infected brood- ers and the 2-step PCR negative brooders were similar (81 and 78 %, respectively), the lightly infected brood- ers gave 88 % WSSV-infected egg batches, some with intense infections or high prevalence (Table 2). Unfor- tunately, some of the lightly infected brooders tested 2- step PCR negative before spawning. Thus, the data argue strongly that brooders should be screened after spawning and that egg batches from l-step WSSV PCR positive spawners should be discarded. Under ideal circumstances, eggs from 2-step WSSV PCR positive brooders should also be discarded. However, under circumstances of low postlarvae availability, it may be possible with appropriate monitoring to utilize the eggs from lightly infected brooders since (1) some (2 = 2 0 % ) gave egg samples that were 2-step PCR nega- tive, (2) none gave egg samples that were l-step PCR positive and (3) the prevalence of 2-step WSSV PCR positive samples was only 30%. It may be possible to rear such eggs to become postlarvae that are 2-step PCR negative or at most 2-step PCR positive for WSSV. We have previously shown (Lo et al. 1998) that ponds stocked with 2-step PCR positive postlarvae can be brought through to successful harvest under low-stress cultivation conditions and with appropriate monitoring to reduce the risk of a WSSV outbreak.
Acknowledgements. This work was supported by the National Science Council under grant no. NSC 88-2317-B- 002-003. We thank Dr Liao, Director General, Taiwan Fish- erles Research Institute (TFRI), for his constructive sugges- tions. We are indebted to Paul Barlow for his helpful criticism of the manuscript and to D. F. Fegan for help with Cochran's Q-test.
LITERATURE CITED
Cai S, Huang J , Wang C, Song X , Sun X, Yu J , Zhang Y, Yang C (1995) Epidemiological studies on the explosive epi- demic disease of prawn in 1993-1994. J Fish China 19: 112-117
Chou HY. Huang CY, Wang CH, Chiang HC, Lo CF (1995) Pathogenicity of a baculovirus infection causing white spot syndrome in cultured penaeid shrimp in Taiwan. Dis Aquat Org 23:165-173
Editorial responsibility: Timothy Flegel, Bangkok, Thailand
Inouye K, Miwa S, Oseko N, Nakano H, IOmura T (1994) Mass mortalities of cultured kuruma shrimp, Penaeus japonicus. in Japan in 1993: electron microscop~c evidence of the causative virus. Fish Pathol 29:149-158 (in Japanese) Lo CF. Kou GH (1998) Virus-associated white spot syndrome
of shrimp in Taiwan: a review. Fish Pathol 33(4):365-371 Lo CF, Leu JH, Ho CH. Chen CH, Peng SE, Chen YT, Chou CM, Yeh PY, Huang CJ, Chou HY, Wang CH, Kou GH (1996a) Detection of baculovirus associated with w h ~ t e spot syndrome (WSBV) in penaeid shrimps uslng poly- merase chain reaction Dis Aquat Org 25:133-141 Lo CF. Ho CH. Peng SE, Chen CH. Hsu HC, Chiu YL, Chang
CF, Liu KF, Su MS, Wang CH, Kou GH (l996b) White spot syndrome baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthopods. Dis Aquat Org 27:215-225
Lo CF, Ho CH, Chen CH, Liu KF, Chiu YL, Yeh PY, Peng SE, Hsu HC, LIU HC, Chang CF, Su MS, Wang CH, Kou GH (1997) Detection a n d tissue tropism of white spot syn- drome baculovirus (WSBV) in captured brooders of Penaeus rnonodon with a special emphasis on reproduc- tive organs. Dis Aquat Org 30:53-72
Lo CF, Chang YS. Cheng CT, Kou GH (1998) PCR monitoring of cultured shrimp for white spot syndrome virus (WSSV) infection in growout ponds. In: Flegel TW (ed) Advances in shrimp biotechnology. National Center for Genetic Engineering and Biotechnology, Bangkok, p 281-286 Momoyama K. Hiraoka 114, Nakano H, Koube H , lnouye K.
Oseka N (1994) Mass mortalities of cultured kuruma shrimp, Penaeus japonicus, in Japan in 1993: histopatho- logical study. Fish Pathol 29 141-148 (in Japanese) Nakano H, Koube H, Umezaea S, Momoyama K, Hlraoka M,
Inouye K, Oseko N (1994) Mass mortalities of cultured kuruma shrimp, Penaeus japonicus, in Japan In 1993: epi- zootiological survey and infection trials. Fish Pathol 29. 135-139 (in Japanese)
Peng SE, Lo CF, Ho CH, Chang CF, Kou GH (1998) Detection of white spot baculovirus (WSBV) in giant freshwater prawn, Macrobrachlum rosenbergii, using polymerase chain reaction. Aquaculture 164:253-262
Takahashi Y, Itami T, Kondom M, Maeda M , Fujii R, Tomon- aga S, Supamattaya K, Boonyaratpahn S (1994) Electron microscopic evidence of bacilhform virus infection in Kuruma shrimp (Penaeus japonicus). Fish Pathol 29: 121-125
Thrusfield MV (1995) Veterinary epidemiology, 2nd edn. Blackwell Science Ltd, Oxford
Tsai MF, Kou GH, Liu HC, Liu KF, Chang CF, Peng SE, Hsu HC, Wang CH, Lo CF (1999) Long-term presence of white spot syndrome virus (WSSV) in a cultivated shrimp popu- lation without disease outbreaks. Dis Aquat Org 38: 107-114
Wang CH, Lo CF, Leu JH, Chou CM, Yeh PY. Chou HY, Tung MC, Chang CF, Su MS, Kou GH (1995) Purification and genomic analysis of baculovirus associated with white spot syndrome (WSBV) of Penaeus monodon. Dls Aquat Org 23:239-242
Wongteerasupaya C, Vickers JE, Sriurairatana S, Nash GL, Akarajamorn A, Boonsaeng V, Panyim S, Tassanakajon A, Withyachumnarnkul B, Flegel TW (1995) A non-occluded. systemic baculovirus that occurs in cells of ectodermal a n d mesodermal origin and causes high mortality in the black tiger prawn Penaeus rnonodon. Dis Aquat Org 21:69-77 Submitted: January 25, 1999; Accepted: September 6, 1999 Proofs received from author(s): December 7, 1999