REGULAR ARTICLE
Toxicity in leaves of rice exposed to cadmium is due
to hydrogen peroxide accumulation
Yi Ting Hsu&Ching Huei Kao
Received: 1 May 2007 / Accepted: 11 July 2007 / Published online: 7 August 2007 # Springer Science + Business Media B.V. 2007
Abstract The production of H2O2 in detached rice
leaves of Taichung Native 1 (TN1) caused by CdCl2
was investigated. CdCl2 treatment resulted in H2O2
production in detached rice leaves. Diphenyleneiodo-nium chloride (DPI) and imidazole (IMD), inhibitors of NADPH oxidase (NOX), prevented CdCl2-induced
H2O2 production, suggesting that NOX is a H2O2
-genearating enzyme in CdCl2-treated detached rice
leaves. Phosphatidylinositol 3-kinase inhibitors wort-manin (WM) or LY294002 (LY) inhibited CdCl2
-inducted H2O2 production in detached rice leaves.
Exogenous H2O2 reversed the inhibitory effect of
WM or LY, suggesting that phosphatidylinositol 3-phosphate is required for Cd-induced H2O2
produc-tion in detached rice leaves. Nitric oxide donor sodium nitroprusside (SNP) was also effective in reducing CdCl2-inducing accumulation of H2O2 in
detached rice leaves. Cd toxicity was judged by the decrease in chlorophyll content. The results indicated that DPI, IMD, WM, LY, and SNP were able to reduce Cd-induced toxicity of detached rice leaves. Twelve-day-old TN1 and Tainung 67 (TNG67) rice seedlings were treated with or without CdCl2. In
terms of Cd toxicity (leaf chlorosis), it was observed that rice seedlings of cultivar TN1 are Cd-sensitive and those of cultivar TNG67 are Cd-tolerant. On treatment with CdCl2, H2O2 accumulated in the
leaves of TN1 seedlings but not in the leaves of TNG67. Prior exposure of TN1 seedlings to 45oC for 3 h resulted in a reduction of H2O2accumulation, as
well as Cd tolerance of TN1 seedlings treated with CdCl2. The results strongly suggest that Cd toxicity of
detached leaves and leaves attached to rice seedlings are due to H2O2accumulation.
Keywords Cadmium . Hydrogen peroxide .
NADPH oxidase . Oryza sativa . Phosphatidylinositol 3-kinase . Phosphatiylinositol 3-phosphate
Abbreviations ASC Ascorbate
DAB 3,3-Diaminobenzidine DMSO Dimethyl sulfoxide
DPI Diphenyleneiodonium chloride HS Heat shock
IMD Imidazole
LY 294002
NO Nitric oxide
NOX NADPH oxidase (EC 1.6.99.6) PI3K Phosphatidylinositol 3-kinase
PI3P Phosphatidylinostolinositol 3-phosphate ROS Reactive oxygen species
SNP Sodium nitroprusside
DOI 10.1007/s11104-007-9357-7
Responsible Editor: Fangjie J. Zhao. Responsible Editor: Fangjie J. Zhao. Y. T. Hsu
:
C. H. Kao (*)Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
TN1 Taichung Native 1 TNG67 Tainung 67 WM Wortmannin
Introduction
Reactive oxygen species (ROS) have been character-ized as the key factor on the responses of plants to both biotic and abiotic stress (Apel and Hirt2004). Initially, ROS were only regarded as damaging to cells. More recently, ROS emerged as ubiquitous signaling mol-ecules participating in the recognition of and the response to stress factors (Foyer and Noctor2005).
Inside plant cells, ROS can be produced in chloroplasts, mitochondria, peroxisome, glyoxy-somes, byproducts of metabolic processes such as photosynthesis and respiration (Apel and Hirt 2004). Several biochemical mechanisms have been proposed to explain ROS production. Apoplastic H2O2
produc-tion can be mediated by cell-wall peroxidase, germin-like oxalate oxidases or amine oxidases (Apel and Hirt2004; Mittler et al.2004). However, it has been suggested that a NADPH oxidase (NOX), analogous to that which generated superoxide during the respiratory burst in mammalian phygocytes, can serve the source of the ROS detected in plants upon successful pathogen recognition (Auh and Murphy
1995; Doke 1983; Low and Merida 1996; Murphy and Auh 1996), in abscisic acid-mediated stomatal closure (Pei et al. 2000), in auxin-regulated gravi-tropic response (Joo et al.2005; Neill et al.2002), and under abiotic stresses such as temperature, wounding and ozone (Dat et al.1998; López-Delgado et al.1998; Orozco-Cárdenas and Ryan1999; Sharma et al.1996). Cadmium (Cd), a heavy metal toxic to humans, animals, and plants, is a widespread pollutant with a long biological half-life (Wagner 1993). It has been demonstrated that Cd can promote the generation of H2O2(Hsu and Kao2004; Kuo and Kao2004; Olmos
et al. 2003; Piqueras et al. 1999; Romero-Puertas et al.2004; Sandalio et al.2001; Schützendübel et al.
2001; Shah et al.2001). The involvement of NOX in Cd-induced H2O2 production has been suggested in
tobacco cells (BY-2 line; Olmos et al. 2003), pea leaves (Romero-Puertas et al. 2004), and pea roots (Rodriguez-Serrano et al.2006).
It has been shown that NOX is activated by binding phosphatidylinositol 3-phosphate (PI3P), a
product of phosphatidylinositol 3-kinase (PI3K), to the PX domain of p40phox (Ellson et al. 2001). In plant cells, PI3P is also known to be required for abscisic acid (ABA)-induced H2O2 production in
guard cells (Jung et al. 2002; Park et al. 2003) and in leaves (Hung and Kao 2004), methyl jasmonate-induced H2O2 production in leaves (Hung et al. 2006), and auxin-induced ROS production in roots (Joo et al. 2005).
We have previously shown that CdCl2 increases
the content of H2O2in detached leaves of rice cultivar
Taichung Native 1 (TN1; Hsu and Kao2004). In this article, we investigate the possibilities that NOX and PI3P, as found in animals cells and guard cells, activate H2O2 generation in CdCl2-treated detached
leaves of TN1 by using NOX inhibitors such as diphenyleneiodonium chloride (DPI) and imidazole (IMD), and PI3K inhibitors wortmanin (WM) and LY294002 (LY). Previously, we demonstrated that rice seedlings of cultivar TNG67 (TNG67) are more tolerant to Cd than those of cultivar TN1 (Hsu and Kao2003). Evidence has been provided to show that abscisic acid is involved in Cd tolerance of TNG67 rice seedlings (Hsu and Kao 2003). It appears that these two cultivars of rice seedlings with different tolerance to Cd provide a good system to study mechanism of Cd toxicity of rice plants. Thus, using detached rice leaves of TN1 and intact leaves attached to rice seedlings of TN1 and TNG67, the possible link of H2O2 between Cd and subsequent toxicity
(chlo-rophyll loss or leaf chlorosis) was also investigated.
Materials and methods Plant material and treatments
Rice (Oryza sativa L., cv. TN1, or TNG67) seeds were sterilized with 2.5% sodium hypochlorite for 15 min and washed extensively with distilled water. These seeds were then germinated in Petri dishes with wetted filter paper at 37°C under dark conditions. After 48 h incubation, uniformly germinated seeds were selected and cultivated in a 500 ml beaker containing half-strength Kimura B solution as de-scribed previously (Hsu and Kao 2003). The hydro-ponically cultivated seedlings were grown for 12 days in a Phytotron (Agricultural Experimental station, National Taiwan University, Taipei, Taiwan) with
natural sunlight at 30°C (day)/25°C (night) and 90% relative humidity. In our laboratory, detached leaves excised from the third leaf of TN1 rice seedlings have long been used as a model system for the studies of stress physiology. Thus, in experiments using de-tached rice leaves, the apical 3 cm of the third leaves of 12-day-old TN1 rice seedlings was used. A group of 10 segments was floated in a Petri dish containing 10 ml of test solution. Incubation was carried out at 27°C in the light (40 μmol m−2 s−1). Test solutions included CdCl2 (5 mM), H2O2 (1 mM), IMD
(50μM), nitric oxide (NO) donor, sodium nitroprus-side (SNP, 100 μM), and PI3K inhibitors, LY (100 μM) and WM (1 μM). LY and WM stock solutions were prepared in 100% dimethyl sulfoxide (DMSO). For experiments of using leaves attached to TN1 and TNG67 seedlings, CdCl2was added to
half-strength Kimura B solution at the time when the third leaf was fully expanded. CdCl2 concentrations at
0.5 mM and 30μM were applied over a short (2 days) and a longer period (6 days), respectively. For some experiments, seedlings exposed to 30°C and 45°C, respectively, for 3 h in darkness, to serve as non-heat shock (control) and heat shock (HS) treatments before the addition of CdCl2(0.5 mM or 30μM).
Evaluation of Cd toxicity
For detached rice leaves, Cd toxicity was judged by the decease in chlorophyll content. Based on our experience from the experiments of Cd effect on rice seedlings, chlorosis is first observed in the second leaf of TN1 seedlings. Thus, Cd toxicity in the second leaves attached to rice seedlings caused by excess Cd was assessed by chlorosis.
Determination of chlorophyll and H2O2
Chlorophyll content was determined according to Wintermans and De Mots (1965) after extraction in 96% (v/v) ethanol. H2O2was visually detected in the
leaves by using 3, 3-diaminobenzidine (DAB) as substrate (Orozco-Cárdenas and Ryan1999). Cd- and inhibitor-treated detached rice leaves were first rinsed with distilled water and were then supplied through the cut ends with DAB (1 mg ml−1) solution for 24 h under light at 27°C. Leaves were decolorized in boiling ethanol (95%) for 0.5 h. This treatment decolorized the leaves except for the brown
polymer-ization spots produced by DAB with H2O2. After
cooling, the leaves were extracted at room tempera-ture with fresh ethanol to visualize the brown polymerization product produced by DAB with H2O2. To verify the specificity of brown spots, before
staining with DAB some leaves were incubated for 2 h in 1 mM ascorbate (ASC), a H2O2scavenger. The
H2O2 staining was repeated four times with similar
results. The H2O2 content was also measured
color-imetrically as described by Jana and Choudhuri (1982). H2O2 was extracted by homogenizing leaf
tissue with phosphate buffer (50 mM, pH 6.5) containing 1 mM hydroxylamine. The homogenate was centrifuged at 6,000 g for 25 min. To determine H2O2content, the extracted solution was mixed with
0.1% titanium chloride in 20% (v/v) H2SO4. The
mixture was then centrifuged at 6,000 g for 25 min. The absorbance was measured at 410 nm. Using this method, we found that absorbance increased linearly with the amount of H2O2 and addition of H2O2 to
extracts resulted in the predicated increase of absor-bance, i.e., added H2O2was fully recovered (data not
shown). The H2O2 content in leaf extracts was
calculated using an extinction coefficient of 0.28μmol−1cm−1.
Statistical analysis
Statistical differences between measurements (n=4) on different treatments or on different times were analyzed following LSD test.
Results
CdCl2increases H2O2production and decreases
chlorophyll in detached rice leaves
In the present study, H2O2 production was first
visualized by a histochemical method with 3,3-diaminobenzidine (DAB) that is based on the forma-tion by H2O2of brown polymerization product. The
development of DAB-H2O2 reaction product in
detached rice leaves could be prevented by H2O2
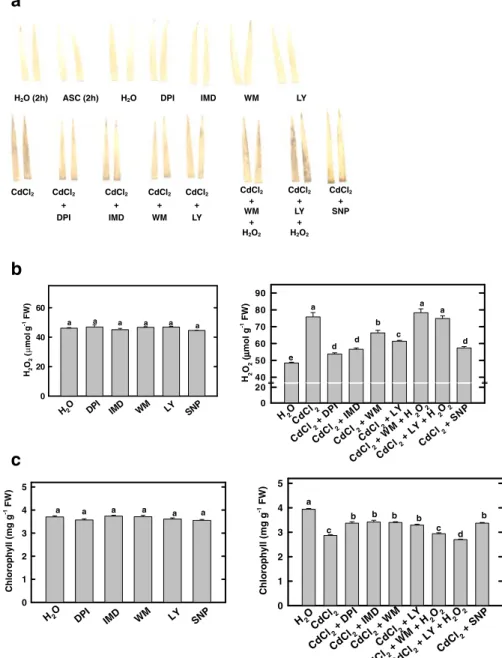
scavengers such as ASC, indicating that the DAB staining method for H2O2is specific (Fig.1a). CdCl2
treatment led to an accumulation of DAB-H2O2
reaction product (Fig.1a). When H2O2was measured
content in detached rice leaves (Fig.1b). As expected, CdCl2treatments resulted in a decrease in chlorophyll
content (Fig.1c).
The effect of NOX inhibitors
The role of NOX in the Cd-stimulated H2O2
production was investigated using NOX inhibitors such as diphenyleneiodonium chloride (DPI) and imidazole (IMD). When 1 μM DPI or 100 μM IMD
was added to detached rice leaves simultaneously with CdCl2, a reduction of Cd-induced H2O2
accu-mulation was observed (Fig. 1a,b). The decrease in chlorophyll content in detached rice leaves by CdCl2
was reduced by DPI and IMD (Fig.1c).
The effect of phosphatidylinositol 3-kinase inhibitors It has been shown that PI3P is important in NOX-mediated H2O2 production during ABA-induced
a
b
c
H2O (2h) ASC (2h) H2O DPI IMD WM LY
CdCl2 CdCl2 + DPI CdCl2 + IMD CdCl2 + WM CdCl2 + LY CdCl2 + WM + H2O2 CdCl2 + LY + H2O2 CdCl2 + SNP a H2 O2 ( μ mo l g -1 FW) 0 20 40 50 60 70 80 90 a a a b c d d d CdC l2 CdC l2 + DPI CdC l2 + IM D CdC l2 + WM CdC l2 + LY CdC l2 + WM + H2 O2 CdC l2 + LY + H2 O2 CdC l2 + SN P e H2O C h lo rop h y ll (mg g -1 FW ) 0 1 2 3 4 5 b b b b c c b CdC l2 CdC l2 + DPI CdC l2 + IMD CdC l2 + WM CdC l2 + LY CdC l2 + WM + H 2O 2 CdC l2 + LY + H2 O2 CdC l2 + SNP a d H2O C h loro phy ll (m g g -1 FW) 0 1 2 3 4 5 a a a a a H2O DPI IMD WM LY a SNP H2 O2 ( μ mol g -1 FW) 0 20 40 60 H2O DPI IMD WM LY SNP a a a a a a
Fig. 1 Effect of DPI, IMD, WM, LY, or SNP on the development of DAB-H2O2
reaction product (a), and the contents of H2O2(b) and
chlorophyll (c) in CdCl2
-treated detached rice leaves (cv. TN1) in the presence or absence of H2O2. The
con-centrations of CdCl2, DPI,
IMD, WM, LY, SNP, and ascorbate (ASC) were 5 mM, 10μM, 50 μM, 1μM, 100 μM, 100 μM, and 1 mM, respectively. DAB–H2O2reaction
prod-uct, for all treatments except ASC was visualized 24 h after treatment in the light. DAB–H2O2reaction
prod-uct for ASC treatment was visualized 2 h after treat-ment in the light. H2O2and
chlorophyll contents were measured 24 h after treat-ment in the light
stomatal closure (Jung et al. 2002), during ABA-promoted leaf senescence (Hung and Kao 2005), during methyl jasmonate-induced leaf senescence (Hung et al. 2006) and during auxin-induced root gravitropic responses (Joo et al. 2005). Thus, it is of great interest to understand whether PI3P is also important in CdCl2-induced H2O2production in rice
leaves. Wortmannin (WM) and LY294002 (LY) are inhibitors of PI3K, a product of which is PI3P. When detached rice leaves were treated with a solution WM (1 μM) or LY (100 μM), CdCl2-induced
accumula-tion of H2O2 in detached rice leaves was reduced
(Fig. 1a,b). WM or LY also reduced Cd-induced decrease in chlorophyll content (Fig.1c). Exogenous H2O2(1 mM) was observed to be able to reverse the
inhibitory effect of WM or LY on H2O2 production
and chlorophyll content (Fig.1a-c).
The effect of nitric oxide donor sodium nitroprusside Nitric oxide (NO) is a bioactive free radical implicated in a number of physiological functions, including intra-cellular mediation of some animal responses (Anbar1995). In plants, NO is involved in many physiological responses, such as pathogen response, programmed cell death, growth, germina-tion, root organogenesis, phytoalexin producgermina-tion, internal iron availability, and ABA-dependent sto-matal closure (Lamattina et al. 2003; Neill et al.
2003). We have previously demonstrated that Cd toxicity in detached rice leaves is reduced by NO (Hsu and Kao 2004). If H2O2is responsible for Cd
toxicity in detached rice leaves, then H2O2
accumu-lation is expected to be reduced by NO donor sodium nitroprusside SNP. As indicated in Fig. 1a, b, it is indeed the case. In the present study, we also show that the decrease in chlorophyll content in detached rice leaves caused by Cd was reduced by SNP (Fig.1c).
CdCl2induces H2O2accumulation in the leaves
of cultivar TN1 seedlings but not in cultivar TNG67 Figure 2 shows the effect of 0.5 mM CdCl2 on the
chlorosis of the second leaves of rice seedlings. It is clear that CdCl2treatment resulted in Cd toxicity of
the second leaves of TN1, but not in TNG67 seedlings (Fig. 2) at 2 days after Cd treatment. If H2O2 is important in regulating Cd toxicity, then
H2O2–DAB reaction product and H2O2 content are
expected to be more and higher, respectively, in CdCl2-treated TN1 seedlings than in TNG67. As
indicated in Fig.3, it is indeed the case.
The concentration of CdCl2used in the
aforemen-tioned study was 0.5 mM. We also conducted experi-ments with lower CdCl2 concentration, 30 μM,
applied over a longer period (6 days). Cd toxicity and H2O2 generation was also observed to be more
pronounced in TN1 seedlings than TNG67 treated with lower concentration (30 μM) CdCl2for 6 days
(Figs. 4 and 5). Thus, the responses to lower CdCl2
concentration are basically in accordance with those to higher CdCl2concentration.
TN1 TNG67
↑↑
CdCl2 – + – +
(0.5 mM, 2 d)
Fig. 2 Effect of CdCl2(0.5 mM) on the toxicity (chlorosis) in
the second leaves of TN1 and TNG67 rice seedlings. Rice seedlings were cultivated in half-strength Kimura B solution in a Phytotron with natural sunlight at 30°C (day)/25°C (night) and 90% relative humidity. CdCl2(0.5 mM) was added to
half-strength Kimura B solution at the time when the third leaves of both TN1 and TNG67 seedlings were fully expanded. Pictures were taken 2 days after the addition of CdCl2(0.5 mM). Arrow
Prior high temperature exposure of TN1 seedlings reduces Cd-induced H2O2 accumulation
Exposure in the range of 2 min to a few hours at temperature 5°C to 15°C above the normal growing temperature is usually called HS treatment. Prior exposure to HS has been shown to increase the tolerance of plants to subsequent Cd stress (Chen and
Kao1995; Orzech and Burke 1988). To test if prior high temperature exposure of TN1 seedlings affects subsequent Cd stress, TN1 seedlings were pretreated at 45°C for 3 h. It was observed that HS pretreatment induces tolerance of Cd stress (0.5 mM or 30μM) in TN1 seedlings (Figs. 6 and 8). Figures 7 and 9 also show that HS pretreatment resulted in the production of less H2O2in the second leaves of TN1 seedlings treated
with CdCl2than that non-HS (30°C) pretreatment.
Discussion
In the present study, 0.5 mM or 30 μM CdCl2was
used to evaluate Cd toxicity of leaves attached to rice
TN1 TNG67 CdCl2 – + – – – + (0.5 mM, 2 d) H2 O2 ( μ mol g -1 FW) 0 20 40 60 80 a bc c b
TN1
TNG67
b
CdCl2 + + (0.5 mM, 2 d)a
Fig. 3 Effect of CdCl2 (0.5 mM) on DAB–H2O2 reaction
product (a) and H2O2content (b) in the second leaves of TN1
and TNG67 rice seedlings. Rice seedlings were cultivated in half-strength Kimura B solution in a Phytron with natural sunlight at 30°C (day)/25°C (night) and 90% relative humidity. CdCl2(0.5 mM) was added to half-strength Kimura B solution
at the time when the third leaves of both TN1 and TNG67 seedlings were fully expanded. The second leaves were excised to visualize H2O2–DAB reaction product and measure H2O2
content 2 days after the addition of CdCl2(0.5 mM)
TN1 TNG67
CdCl2 – + – +
(30 μM, 6 d)
Fig. 4 Effect of CdCl2(30μM) on the toxicity (chlorosis) in
the second leaves of TN1 and TNG67 rice seedlings. Rice seedlings were cultivated in half-strength Kimura B solution in a Phytotron with natural sunlight at 30°C (day)/25°C (night) and 90% relative humidity. CdCl2(30μM) was added to
half-strength Kimura B solution at the time when the third leaves of both TN1 and TNG67 seedlings were fully expanded. Pictures were taken 2 days after the addition of CdCl2(30μM). Arrow
seedlings. In a recent study on 64 soils (urban, forest, and agricultural soils) containing various levels of Cd contamination, free dissolved Cd concentrations ranged from 0.1 to 2,000 nM (Sauvé et al. 2000). Thus. The 30 μM CdCl2 used in some of our
experiments can be considered to be very high. Basically, rice seedlings in our study were considered to be suffering from acute Cd toxicity.
It has been shown that Cd is able to produce H2O2
(Hsu and Kao2004; Kuo and Kao2004; Olmos et al.
2003; Piqueras et al. 1999; Romero-Puertas et al.
2004; Sandalio et al.2001; Schützendübel et al.2001; Shah et al. 2001). Here, we also show that CdCl2
induced H2O2 production in detached rice leaves by
histochemistry with DAB (Fig. 1a) and colorimetric methods (Fig. 1b). Wounding is known to induce H2O2production (Orozco-Cárdenas and Ryan 1999).
When detached rice leaves are used, wounding is always a problem. However in the present study, each long and narrow rice leaf was cut transversely; thus the area of wounding was very small. Therefore, H2O2 generation of detached rice leaves induced by
CdCl2 is unlikely to be complicated the wounding
effect.
A role for plasma membrane NOX in the produc-tion of the H2O2 has been a recent focus in ROS
signaling (Sagi and Flurhr2006). Here, we show that DPI and IMD, inhibitors of NOX, reduced CdCl2
-induced H2O2 accumulation (Fig. 1a,b). It has been
shown that a high concentration of DPI can affect ← ← Pretreatment HS – – + + Treatment CdCl2 (0.5 mM, 2 d) – + – +
Fig. 6 Effect of HS pretreatment on the toxicity (chlorosis) in the second leaves of rice seedlings (cv. TN1) in the presence or absence of CdCl2(0.5 mM). Rice seedlings at the time when
the third leaves fully expanded were transferred to 30°C (non-HS) or 45°C ((non-HS) in the dark for 3 h. Rice seedlings were then cultivated in a Phytotron with natural sunlight at 30°C (day)/ 25°C (night) and 80% relative humidity. Pictures were taken 2 days after the addition of CdCl2 (0.5 mM) to half-strength
Kimura B solution. Arrow indicated the second leaves
TN1 TNG67 CdCl2 – + – + (30 μμM, 6 d)
a
H 2 O2 ( μ mo l g -1 FW) 0 20 40 60 80 a c c bTN1
TNG67
b
CdCl2 – + – + (30 μM, 6 d)a
Fig. 5 Effect of CdCl2 (30 μM) on DAB–H2O2 reaction
product (a) and H2O2content (b) in the second leaves of TN1
and TNG67 rice seedlings. Rice seedlings were cultivated in half-strength Kimura B solution in a Phytron with natural sunlight at 30°C (day)/25°C (night) and 90% relative humidity. CdCl2(30μM) was added to half-strength Kimura B solution
at the time when the third leaves of both TN1 and TNG67 seedlings were fully expanded. The second leaves were excised to visualize H2O2–DAB reaction product and measure H2O2
other enzymes potentially involved in the production of ROS, including cell wall peroxidase and NO synthase (Bolwell et al. 1998). The fact that CdCl2-induced
H2O2 accumulation in detached rice leaves can be
inhibited by low concentration of DPI (1μM) and can be inhibited by both DPI and IMD (Fig.1a,b) strongly suggest that CdCl2-dependent H2O2 production
orig-inated, at least in part, from plasma membrane NOX. The involvement of NOX in Cd-induced H2O2
production has also been suggested in tobacco cells (Olmos et al.2003), pea leaves (Romero-Puertas et al.
2004), and pea roots (Rodriguez-Serrano et al.2006). It has been shown that zinc deficiency enhanced NAD(P)H-dependent superoxide radical production in
plasma membrane vesicles isolated from roots of bean plants (Pinton et al. 1994). Since cadmium toxicity has been shown to be associated zinc deficiency (Sandalio et al. 2001), thus the possibility that a Cd-induced reduction in Zn availability could stimulate superoxide radical production in leaves of rice seed-lings cannot be excluded.
The mechanism of ROS production and the molecules involved have been well investigated in animal cells, particularly in neutrophils. The NOX, which consists of many components, is responsible for ROS production in neutrophil cells and is activated by the binding of PI3P to one of the components (Ellson et al. 2001). In plant cells, PI3P is also known to be required for ABA-induced H2O2
production in guard cells (Jung et al.2002; Park et al.
2003) and in detached rice leaves (Hung and Kao
2005), methyl jasmonate-induced H2O2production in
detached rice leaves (Hung et al. 2006), and auxin-induced ROS production in roots (Joo et al.2005). It Pretreatment HS Treatment CdCl2 (0.5 mM, 2 d) Pretreatment HS Treatment CdCl2 (0.5 mM, 2 d) – – + + – + – + – – + + – + – + a H2 O2 ( μ mol g -1 FW) 0 20 40 60 a c bc b
b
a
Fig. 7 Effect of HS pretreatment on the H2O2–DAB reaction
product (a) and H2O2content (b) in the second leaves of rice
seedlings (cv. TN1) in the presence or absence of CdCl2
(0.5 mM). Rice seedlings at the time when the third leaves fully expanded were transferred to 30°C (non-HS) or 45°C (HS) in the dark for 3 h. Rice seedlings were then cultivated in a Phytotron with natural sunlight at 30°C (day)/25°C (night) and 80% relative humidity. The second leaves were were excised to visualize H2O2–DAB reaction product and measure H2O2
content, 2 days after the addition of CdCl2(0.5 mM) to
half-strength Kimura B solution
Pretreatment HS Treatment CdCl2 (30 μM, 6 d) – – + + – + – +
Fig. 8 Effect of HS pretreatment on the toxicity (chlorosis) in the second leaves of rice seedlings (cv. TN1) in the presence or absence of CdCl2(30μM). Rice seedlings at the time when the
third leaves fully expanded were transferred to 30°C (non-HS) or 45°C (HS) in the dark for 3 h. Rice seedlings were then cultivated in a Phytotron with natural sunlight at 30°C (day)/ 25°C (night) and 90% relative humidity. Pictures were taken 6 days after the addition of CdCl2(30 μM) to half-strength
appears that PI3P is important in CdCl2-induced H2O2
production based on two lines of evidence. First, LY or WM, inhibitor of PI3K, was able to reduce CdCl2
-induced H2O2 production (Fig.1a,b). Second,
exog-enous H2O2reversed the inhibitory effect of the PI3K
inhibitors on CdCl2-indcued H2O2 accumulation
(Fig. 1a,b). These results supported further that NOX is involved in CdCl2-induced H2O2production.
In neutrophlis, PI3P regulated H2O2 production by
binding the non-catalytic component p40phox of the NOX (Ellson et al. 2001). However, a rice homolog
of p40phox has not been reported. Therefore, the detailed mechanism of the action of PI3P during H2O2 production in the rice leaves needs further
investigation.
The fact that NOX and PI3K inhibitors, which reduced the Cd-induced H2O2production (Fig.1a,b),
were able to prevent Cd-decreased chlorophyll con-tent in detached rice leaves (Fig.1c) suggests that Cd toxicity of detached rice leaves is due to H2O2
accumulation.
Several reports convincingly demonstrate that NO is a able to counteract the toxicity of paraquat and diquat, which are known to generate ROS in potato and rice leaves (Beligni and Lamattina 1999; Hung et al. 2002), and block H2O2 production induced by
jasmonic acid in tomato leaves (Orozco-Cárdenas and Ryan 2002). Here we also showed that SNP, a NO donor, blocks H2O2 production and reduces Cd
toxicity (Fig. 1a,b). It has been shown that poly-amines are able to protect against oxidative damage caused by paraquat (Chang and Kao1997), acid rain (Velikova et al. 2000) and heavy metals such as Cd and Cu (Groppa et al. 2001). Recently, we reported that polyamines (spermidine and spermine) are able to protect Cd-induced toxicity of detached rice leaves and this protection is most likely related to the avoidance of H2O2 generation and Cd uptake
(Hsu and Kao2007). All these results support further that Cd toxicity is due to H2O2production in detached
rice leaves.In plants, the most general symptom of Cd toxicity is chlorosis (Das et al. 1997). Based on chlorosis and chlorophyll loss of the second leaves of rice seedlings, it was demonstrated that rice seedlings of cultivar TNG67 are more tolerant to Cd than those of cultivar TN1 (Hsu and Kao 2003; Kuo and Kao
2004). Evidence has also been provided to show that Cd content in the second leaf and roots of TNG67 seedlings remained unchanged and slightly increased, respectively, after Cd treatment (Hsu and Kao 2003; Kuo and Kao2004). In contrast, a marked increase in Cd content in Cd-treated TN1 leaves and roots was observed (Hsu and Kao 2003; Kuo and Kao 2004). We also observed that higher content of Cd in TN1 than TNG67 leaves results in more production of H2O2 in TN1 than TNG67 leaves (Kuo and Kao 2004). The fact that on treatment with CdCl2(0.5 mM
for 2 days or 30 μM for 6 days), the H2O2 content
increased in Cd-sensitive TN1 seedlings but not in Cd-tolerant TNG67 (Figs. 3 and5) supports the idea
Pretreatment HS Treatment CdCl2 (30 μμM, 6 d) Pretreatment HS Treatment CdCl2 (30 μM, 6 d) – – + + – + – + – – + + – + – + H2 O2 ( μ mol g -1 FW) 0 20 40 60 80 a c b c
b
a
Fig. 9 Effect of HS pretreatment on the H2O2–DAB reaction
product (a) and H2O2content (b) in the second leaves of rice
seedlings (cv. TN1) in the presence or absence of CdCl2
(30μM). Rice seedlings at the time when the third leaves fully expanded were transferred to 30°C (non-HS) or 45°C (HS) in the dark for 3 h. Rice seedlings were then cultivated in a Phytotron with natural sunlight at 30°C (day)/25°C (night) and 90% relative humidity. The second leaves were were excised to visualize H2O2–DAB reaction product and measure H2O2
content 6 days after the addition of CdCl2(30 μM) to
that Cd toxicity is due to H2O2accumulation caused
by Cd. Prior exposure to HS has been shown to increase tolerance of plants to subsequent Cd stress (Chen and Kao1995; Orzech and Burke1988). Here we also demonstrated that HS pretreatment of TN1 seedlings resulted in the reduction of H2O2
produc-tion as well as Cd toxicity of TN1 seedlings treated with CdCl2(Figs.6,7,8, and9). H2O2production is
believed to be the consequence of lower capacity of antioxidant system to avoid H2O2 accumulation. In
this connection, HS pretreatment may increase the capacity of antioxidant system to reduce H2O2
production under the subsequent Cd treatment. Our unpublished data indeed show that HS pretreatment of rice seedlings resulted in higher contents of reduced glutathione and ascorbate and higher activities of glutathione reductase and ascorbate peroxidase than non-HS.
Based on the results obtained from detached rice leaves and intact leaves attached to rice seedlings, we conclude that Cd toxicity in leaves of rice seedlings is due to H2O2 accumulation. This conclusion is
basically consistent with the results of Cho and Seo (2005), who reported that a lower H2O2accumulation
confers Cd-tolerance in Arabidopsis seedlings.
Acknowledgements This work was supported by a research grant from the National Science Council of the Republic of China (NSC 94-2313-B-002-028).
References
Anbar M (1995) Nitric oxide: a synchronizing chemical messenger. Experientia 51:545–550
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55:373–399
Auh C-K, Murphy TM (1995) Plasma membrane redox enzyme is involved in the synthesis of O2 and H2O2 by
Phytophthora elicitor-stimulated rose cells. Plant Physiol 107:1241–1247
Beligni MV, Lamattina L (1999) Nitric oxide protects against cellular damage produced by methyl viologen herbicides in potato plants. Nitric Oxide 3:199–208
Bolwell GP, Davies DR, Gerrish C, Auh CK, Murphy TM (1998) Comparative biochemistry of the oxidative burst produced by rose and French bean cells reveals two distinct mechanisms. Plant Physiol 116:1374–1385 Chang CJ, Kao CH (1997) Paraquat toxicity is reduced by
polyamines in rice leaves. Plant Growth Regul 22:163–168 Chen SL, Kao CH (1995) Prior temperature exposure affects subsequent Cd-induced ethylene production in rice leaves. Plant Sci 104:135–138
Cho U-H, Seo N-H (2005) Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation. Plant Sci 168:113–120
Das P, Sammantaray S, Rout GR (1997) Studies on cadmium toxicity: a review. Environ Pollut 98:29–36
Dat JF, Foyer CH, Scott IM (1998) Changes in salicylic acid and antioxidants during induced thermotolerance in mustard seedlings. Plant Physiol 118:1455–1461 Doke N (1983) Generation of superoxide anion by potato tuber
protoplasts during the hypersensitive response to hyphal cell wall components of Phytophthora infestans and specific inhibition of the reaction by suppressors of hypersensitivity. Physiol Plant Pathol 23:359–367 Ellson CD, Gobert-Gosse S, Anderson KE, Davidson K,
Erdjument-Bromage H, Tempst P, Thring JW, Cooper MA, Lim Z-Y, Holmes AB, Gaffney PRJ, Coadwell J, Chilvers ER, Hawkins PT, Steppheas LR (2001) PtdIns(3) P regulated the neutrophil oxidase complex by binding to the PX domain of p40phox. Nat Cell Biol 3:679–682 Foyer CH, Noctor G (2005) Oxidant and antioxidant signaling
in plants: a re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ 28: 1056–1071
Groppa MD, Tomaro ML, Benavides MP (2001) Polyamines as protectors against cadmium or copper-induced oxi-dative damage in sunflower leaf discs. Plant Sci 161:481–488
Hsu YT, Kao CH (2003) Role of abscisic acid in cadmium tolerance of rice (Oryza sativa L.) seedlings. Plant Cell Environ 20:867–874
Hsu YT, Kao CH (2004) Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul 42:227–238 Hsu YT, Kao CH (2007) Cadmium-induced oxidative damage
in rice leaves is reduced by polyamines. Plant Soil 291:27–37
Hung KT, Kao CH (2004) Phosphatidylinositol 3-phosphate is required for abscisic acid-induced hydrogen peroxide production in rice leaves. Plant Growth Regul 45:96–101 Hung KT, Kao CH (2005) Nitric oxide counteracts the senescence of rice leaves induced by hydrogen peroxide. Bot Bull Acad Sin 46:21–28
Hung KT, Chang CJ, Kao CH (2002) Paraquat toxicity is reduced by nitric oxide in rice leaves. J Plant Physiol 159:159–166 Hung KT, Hsu YT, Kao CH (2006) Hydrogen peroxide is
involved in methyl jasmonate-induced senescence of rice leaves. Physiol Plant 127:293–303
Jana S, Choudhuri MH (1982) Glycolate metabolism of these submerged aquatic angiosperms during aging. Aquat Bot 12:345–354
Joo JH, Yoo HJ, Hwang I, Lee JS, Nam KH, Bae YS (2005) Auxin-induced reactive oxygen species production re-quired the activation of phosphatidyliniaitol 3-kinase. FEBS Lett 579:1243–1248
Jung J-Y, Kim Y-W, Kwak JM, Hwang J-U, Young J, Schroeder JI, Hwang I, Lea Y (2002) Phosphatidyl-inostol 3- and 4-phosphate are required for normal stomatal movements. Plant Cell 147:2397–2412
Kuo MC, Kao CH (2004) Antioxidant enzyme activities are upregulated in response to cadmium in sensitive, but not in tolerant rice (Oryza sativa L.) seedlings. Bot Bull Acad Sin 45:291–299
Lamattina L, Garaia Mata C, Graziano M, Pagnussat G (2003) Nitric oxide: the versatility of an extensive signal molecule. Annu Rev Plant Biol 54:109–136
López-Delgado H, Dat JF, Foyer CH, Scoot IM (1998) Induction of tolerance in potato microplants by acetyl-salicylic acid and H2O2. J Exp Bot 49:713–720
Low PS, Merida JR (1996) The oxidative burst in plant defense: function and signal transduction. Physiol Plant 96:533–542
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F (2004) Tends Plant Sci 9:490–498
Murphy TM, Auh C-K (1996) The superoxide synthases of plasma membrane preparations from cultured rose cells. Plant Physiol 110:621–629
Neill SJ, Desikan R, Hancock J (2002) Hydrogen peroxide signaling. Curr Opin Plant Biol 5:388–395
Neill SJ, Desikan R, Hancock JT (2003) Nitric oxide signalings in plants. New Phytol 159:11–35
Olmos EO, Martínez-Solano JR, Piqueras A, Hellín E (2003) Early steps in the oxidative burst induced by cadmium in cultured tobacco cells (BY-2 line). J Exp Bot 54:291–301 Orozco-Cárdenas ML, Ryan CA (1999) Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid pathway. Proc Natl Acad Sci USA 96:6553–6557
Orozco-Cárdenas ML, Ryan CA (2002) Nitric oxide negatively modulate wound signaling in tomato plants. Plant Physiol 130:487–493
Orzech KA, Burke JJ (1988) Heat shock and the protection against metal toxicity in wheat leaves. Plant Cell Environ 11:711–714
Park K-Y, Jung J-Y, Park J, Hwang JU, Kim Y-W, Hwang I, Lee Y (2003) A role for phosphatidylinositol 3-phosphate in abscisic acid-induced reactive oxygen species genera-tion in guard cells. Plant Physiol 132:92–98
Pei Z-M, Murata Y, Benning G, Thomine S, Klüsener B, Allen GJ, Grill E, Schroeder JI (2000) Calcium channels activiated by hydrogen peroxide mediate abscisic acid signaling in guard cells. Nature 406:731–734
Pinton R, Cakmak I, Marschner H (1994) Zinc deficiency enhanced NAD(P)H-dependent superoxide radical produc-tion in plasma membrane vesicles isolated from roots of bean plants. J Exp Bot 45:45–50
Piqueras A, Olmos E, Martińez-Solano JR, Hellín E (1999) Cd-induced oxidative burst in tobacco BY2 cells: time course,
subcellular location and antioxidant response. Free Radic Res 31:S33–S38
Rodriguez-Serrano M, Romero-Puertas C, Zabalza A, Corpas FJ, Gómez M, del Rio LA, Sandalio LM (2006) Cadmium effect on oxidative metabolism of pea (Pisum sativum L.) roots. Imaging of reactive oxygen species and nitric oxide accumulation in vivo. Plant Cell Environ 291:1532–1544
Romero-Puertas MC, Rodriguez-Serrano M, Corpas FJ, Gómez M, del Rio LA, Sandalio LM (2004) Cadmium-induced subcellular accumulation of O2 and H2O2in pea
leaves. Plant Cell Environ 27:1122–1134
Sagi M, Flurhr R (2006) Production of reactive oxygen species by plant NADPH oxidase. Plant Physiol 141:336–340 Sandalio LM, Dalurzo HC, Gómez M, Romero-Puertas MC,
del Rio LA (2001) Cadmium-induced changes in the growth and oxidative metabolism of pea plant. J Exp Bot 52:2115–2126
Sauvé S, Norvell WA, McBride M, Hendershot W (2000) Speciation and complexation of cadmium in extracted soil solutions. Environ Sci Technol 34:291–296
Schützendübel A, Schwang P, Teichmann T, Gross K, Langenfeld-Heyer R, Godbold DL, Polle A (2001) Cadmium-induced changes in antioxidative systems, hy-drogen peroxide content, and differentiation in scots pine roots. Plant Physiol 127:887–898
Shah K, Kumar RG, Verma S, Dubey RS (2001) Effect of cadmium on lipid peroxidation, superoxide anion genera-tion and activities of antioxidant enzymes in growing rice seedlings. Plant Sci 161:1135–1144
Sharma Y, León J, Roskinn I, Davies KR (1996) Ozone-induced responses in Arabidopsis thaliana: the role of salicylic acid in the accumulation of defense-related transcripts and induced resistance. Proc Natl Acad Sci USA 93:5099–5104
Velikova V, Yordanov I, Edreva S (2000) Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Protective role of exogenous polyamines. Plant Sci 151:59–66
Wagner GJ (1993) Accumulation of cadmium in crop plants and its consequences to human health. Adv Agron 5:173– 212
Wintermans JFGM, De Mots A (1965) Spectrophotometric characteristics of chlorophyll a and b and their pheophy-tins in ethanol. Biochim Biophys Acta 109:448–453